Preparation Of Libraries Of Protein Variants Expressed In Eukaryotic Cells And Use For Selecting Binding Molecules
McCafferty; John ; et al.
U.S. patent application number 16/788039 was filed with the patent office on 2020-05-28 for preparation of libraries of protein variants expressed in eukaryotic cells and use for selecting binding molecules. This patent application is currently assigned to IONTAS LIMITED. The applicant listed for this patent is IONTAS LIMITED. Invention is credited to Michael Dyson, John McCafferty, Kothai Parthiban.
| Application Number | 20200165597 16/788039 |
| Document ID | / |
| Family ID | 50980562 |
| Filed Date | 2020-05-28 |







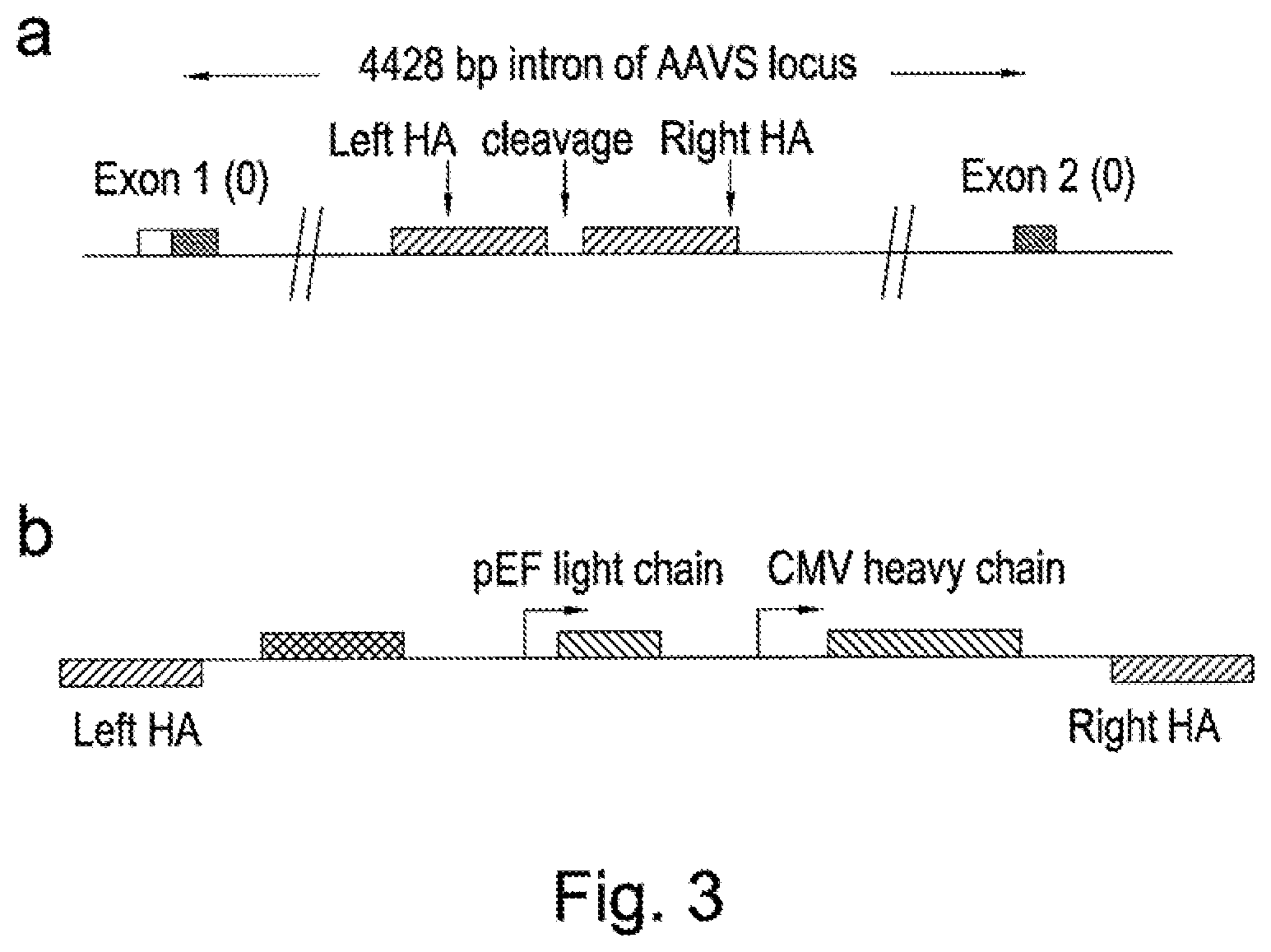


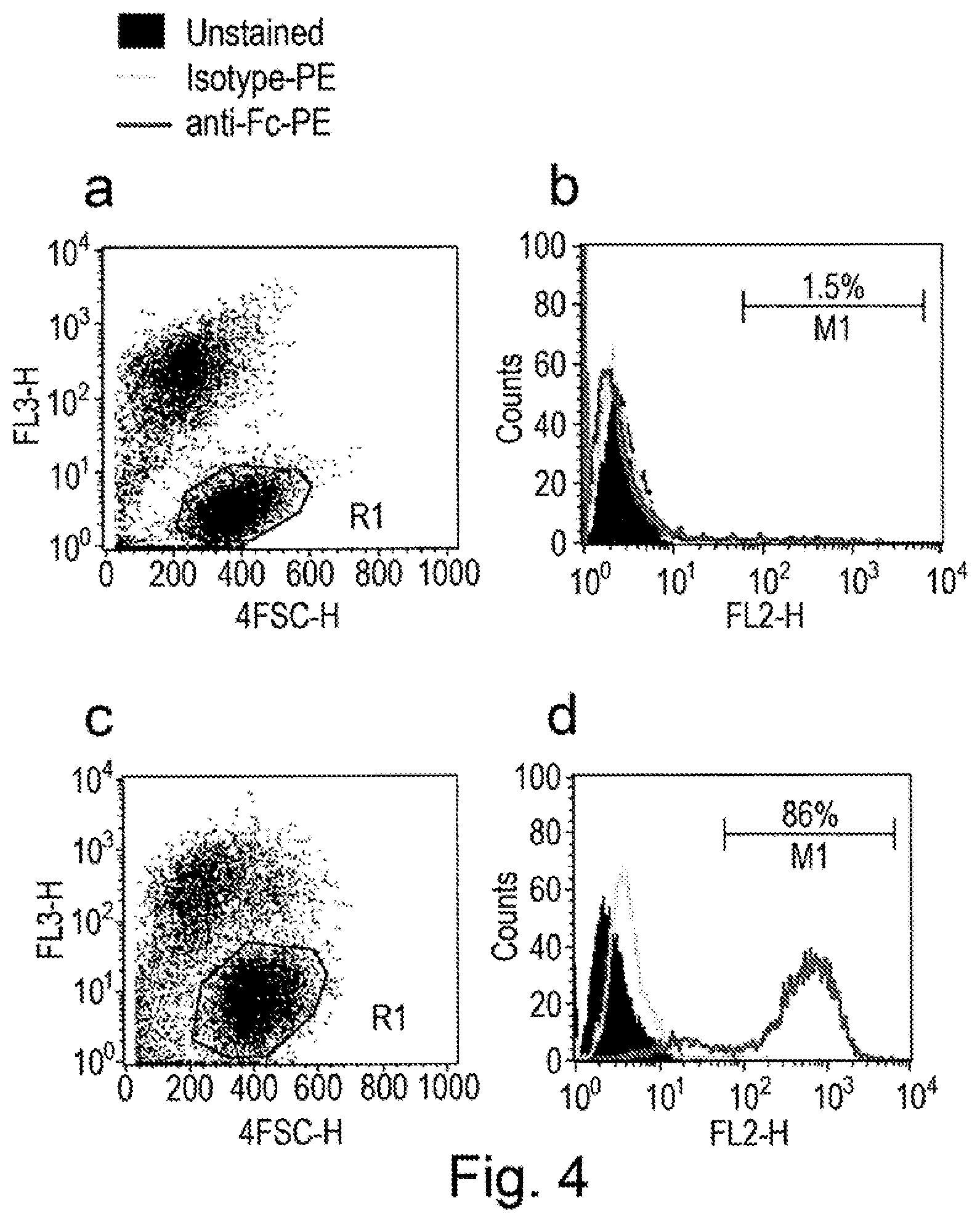

View All Diagrams
| United States Patent Application | 20200165597 |
| Kind Code | A1 |
| McCafferty; John ; et al. | May 28, 2020 |
PREPARATION OF LIBRARIES OF PROTEIN VARIANTS EXPRESSED IN EUKARYOTIC CELLS AND USE FOR SELECTING BINDING MOLECULES
Abstract
The invention relates to methods of producing eukaryotic cell libraries encoding a repertoire of binding molecules ("binders"), wherein the methods use a site-specific nuclease for targeted cleavage of cellular DNA to enhance site-specific integration of binder genes through endogenous cellular repair mechanisms. Populations of eukaryotic cells are produced in which a repertoire of genes encoding binders are integrated into a desired locus in cellular DNA (e.g., a genomic locus) allowing expression of the encoded binding molecule, thereby creating a population of cells expressing different binders.
| Inventors: | McCafferty; John; (Babraham, GB) ; Dyson; Michael; (Cambridge, GB) ; Parthiban; Kothai; (Cambridge, GB) | ||||||||||
| Applicant: |
|
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Assignee: | IONTAS LIMITED Cambridge GB |
||||||||||
| Family ID: | 50980562 | ||||||||||
| Appl. No.: | 16/788039 | ||||||||||
| Filed: | February 11, 2020 |
Related U.S. Patent Documents
| Application Number | Filing Date | Patent Number | ||
|---|---|---|---|---|
| 15308570 | Nov 2, 2016 | |||
| PCT/GB2015/051287 | May 1, 2015 | |||
| 16788039 | ||||
| Current U.S. Class: | 1/1 |
| Current CPC Class: | C12N 15/907 20130101; C07K 2317/622 20130101; C07K 2317/64 20130101; C07K 2319/036 20130101; C07K 2319/035 20130101; C12N 15/1037 20130101; C07K 16/005 20130101; C07K 2319/03 20130101; C12N 15/102 20130101; C12N 2800/80 20130101; C12N 2800/30 20130101; C12N 2310/20 20170501; C07K 2317/24 20130101 |
| International Class: | C12N 15/10 20060101 C12N015/10; C07K 16/00 20060101 C07K016/00; C12N 15/90 20060101 C12N015/90 |
Foreign Application Data
| Date | Code | Application Number |
|---|---|---|
| May 2, 2014 | GB | 1407852.1 |
Claims
1. A method of integrating donor DNA into genomic DNA of a higher eukaryotic cell a having a genome size of greater than 2.times.10.sup.7 base pairs, comprising providing single-stranded donor DNA, wherein the donor DNA is packaged in an adeno-associated viral (AAV) vector and comprises a large transgene encoding a binder, wherein the binder is an antibody molecule, a receptor or a protein, introducing the donor DNA into a cell, and cleaving the genomic DNA with a site-specific nuclease, whereby the transgene is incorporated into the genomic DNA.
2. The method of claim 1, wherein the binder is an engineered protein, a T cell receptor (TCR) or a chimeric antigen receptor (CAR).
3. The method of claim 1, wherein the binder is a full length immunoglobulin, IgG, Fab, scFv-Fc or scFv.
4. The method of claim 1, wherein the site-specific nuclease is a meganuclease, a zinc finger nuclease, a TALE nuclease or a nucleic acid-guided nuclease.
5. The method of claim 4, wherein the site-specific nuclease is a CRISPR/Cas nuclease.
6. The method of claim 5, wherein the site-specific nuclease is introduced as a protein:RNA complex.
7. The method of claim 1, wherein the cell is a mammalian cell.
8. The method of claim 7, wherein the cell is a HEK293 cell, a Chinese hamster ovary cell, a T lymphocyte lineage cell or a B lymphocyte lineage cell.
9. The method of claim 8, wherein the cell is a primary T cell or a T cell line.
10. The method of claim 8, wherein the cell is a primary B cell, a B cell line, a pre-B cell line or a pro-B cell line.
11. The method of claim 1, the method further comprising culturing the eukaryotic cell comprising the transgene incorporated into its genomic DNA under conditions for expression of the binder, and isolating the binder.
12. A method of producing a library of eukaryotic cell clones containing DNA encoding a diverse repertoire of binders, wherein the binders are antibody molecules, receptors or proteins, comprising providing single-stranded donor DNA molecules encoding the binders, wherein the donor DNA molecules are packaged in AAV vectors, providing higher eukaryotic cells with a genome size of greater than 2.times.10.sup.7 base pairs, introducing the donor DNA into the cells and providing a site-specific nuclease within the cells, wherein the nuclease cleaves a recognition sequence in cellular DNA to create an integration site at which the donor DNA becomes integrated into the cellular DNA, integration occurring through DNA repair mechanisms endogenous to the cells, thereby creating recombinant cells containing donor DNA integrated in the cellular DNA, and culturing the recombinant cells to produce clones, thereby providing a library of eukaryotic cell clones containing donor DNA encoding the repertoire of binders.
13. The method of claim 12, wherein the binders are full length immunoglobulins, IgG, Fab, scFv-Fc or scFv.
14. The method of claim 12, wherein the binders are T cell receptors or chimeric antigen receptors (CARs).
15. A method of screening for a binder that recognises a target, comprising: producing a library by the method of claim 12, culturing cells of the library to express the binders, exposing the binders to the target, allowing recognition of the target by one or more cognate binders, if present, and detecting whether the target is recognised by a cognate binder.
16. The method of claim 15, further comprising detecting target recognition by a cognate binder, and recovering cells of a clone containing DNA encoding the cognate binder.
17. The method of claim 16, further comprising isolating nucleic acid encoding the binder from the recovered clone, thereby obtaining nucleic acid encoding a binder that recognises the target.
18. The method of claim 17, comprising optionally introducing mutation or converting the nucleic acid to modified nucleic acid encoding a restructured binder, and introducing DNA encoding the binder into a host cell.
19. The method of claim 18, further comprising culturing the cells to express the binder, and purifying the binder.
Description
CROSS-REFERENCE TO RELATED APPLICATIONS
[0001] This application is a continuation of U.S. patent application Ser. No. 15/308,570, filed Nov. 2, 2016, which is a 35 U.S.C. .sctn. 371 application of International Application Serial No. PCT/GB2015/051287, filed May 1, 2015; which claims the benefit of Great Britain Application Number 1407852.1 that was filed on May 2, 2014. The entire content of the applications referenced above are hereby incorporated by reference herein.
SEQUENCE LISTING
[0002] The instant application contains a Sequence Listing which has been submitted in ASCII format via EFS-Web and is hereby incorporated by reference in its entirety. Said ASCII copy, created on Oct. 31, 2016, is named 00130_003US1_SL.txt and is 92.9 kilobytes in size.
FIELD OF THE INVENTION
[0003] This invention relates to methods of producing eukaryotic (e.g., mammalian) cell libraries for screening and/or selection of binding molecules such as antibodies. Libraries can be used to contain and display a diverse repertoire of binders, allowing binders to be screened to select one or more binders having a desired property such as specificity for a target molecule. The invention especially relates to methods of introducing donor DNA encoding the binders into eukaryotic cells to provide a cell library in which a desired number of donor DNA molecules are faithfully integrated at a desired locus or loci in the cells.
INTRODUCTION
[0004] Protein engineering techniques permit creation of large diverse populations of related molecules (e.g., antibodies, proteins, peptides) from which individual variants with novel or improved binding or catalytic properties can be isolated. The ability to construct large populations of eukaryotic cells, particularly mammalian cells, where each cell expresses an individual antibody, peptide or engineered protein would have great value in identifying binders with desired properties.
[0005] The basic principle of display technology relies on the linkage of a binding molecule to the genetic information encoding that molecule. The binding properties of the binding molecule are used to isolate the gene which encodes it. This same underlying principles applies to all forms of display technology including, bacteriophage display, bacterial display, retroviral display, baculoviral display, ribosome display, yeast display and display on higher eukaryotes such as mammalian cells [1, 2, 3, 4].
[0006] Display technology has best been exemplified by display of antibodies on filamentous bacteriophage (antibody phage display) which over the last 24 years has provided important tools for discovery and engineering of novel binding molecules including the generation of human therapeutic antibodies. Using phage display antibody molecules are presented on the surface of filamentous bacteriophage particles by cloning the gene encoding an antibody or antibody fragment in-frame with the gene encoding a phage coat protein. The antibody genes are initially cloned into E. coli such that each bacterium encodes a single antibody. Generation of bacteriophage from the bacteria using standard methods results in the generation of bacteriophage particles displaying an antibody fragment on their surface and encapsulating the encoding antibody gene within the bacteriophage. The collection of bacteria or the bacteriophage derived from them is referred to as an "antibody library". Using antibody phage display, antibodies and their associated genes can be enriched within the population by exposing antibody-presenting bacteriophage to a target molecule of interest.
[0007] To allow recovery of bacteriophage displaying a binder recognising a target of interest, the target molecule needs to be immobilised onto the surface of a selection vessel or needs to be recoverable from solution by secondary reagents, e.g., biotinylated target protein, recovered from solution using streptavidin-coated beads. Following incubation of the library of binder-displaying bacteriophage with the target molecule, unbound phage are removed. This involves washing the matrix to which the target (and associated bacteriophage) is attached to remove unbound bacteriophage. Bound bacteriophage with their associated antibody gene can be recovered and/or infected into host bacterial cells. Using the approach outlined above it becomes possible to enrich a subset of bacteriophage clones capable of binding a target molecule of choice. Phage display libraries have been shown to provide a rich source of antibody diversity, providing hundreds of unique antibodies to a single target [5,6,7].
[0008] Historically, display systems for isolating novel antibody binding specificities have been based in prokaryotic systems and in particular on display of single chain Fvs (scFv) and to a lesser extent as Fabs on bacteriophage. Display of binders on the surface of bacteria has been described but has not been widely used and applications have largely been limited to peptide display or display of antibody fragments pre-enriched for binders through immunisation [8]. Despite the power of prokaryotic display systems including phage display there are limitations. Following selection by phage or ribosome display the genes encoding individual binding molecules are identified by introducing the selected gene population into bacteria, plating the bacterial populations, picking colonies, expressing binding molecules into the supernatant or periplasm and identifying positive clones in binding assays such as enzyme or fluorescence linked immunosorbent assays (ELISA). Although binding molecules are identified this approach does not resolved information on the extent of expression and the binding affinity of the resultant clones. Thus although it is possible to generate potentially thousands of binders, the ability to screen the output is limited by the need for colony picking, liquid handling etc., coupled with limited primary information on relative expression level and affinity.
[0009] Display of binding molecules on the surface of eukaryotic cells has the potential to overcome some of these problems. In conjunction with flow cytometry, eukaryotic display allows rapid, high throughput selection. It becomes possible to survey millions of cellular clones expressing different binding molecules on their surface. Cell surface display has best been exemplified for the display of antibody fragments formatted as scFvs on the surface of yeast cells. A commonly used modality for yeast surface display makes use of the yeast agglutinin proteins (Aga1p and Aga2p). As described by Chao et al. [9], genes encoding a repertoire of scFvs are genetically fused with the yeast agglutinin Aga2p subunit. The Aga2p subunit then attaches to the Aga1p subunit present in the cell wall via disulphide bonds. Yeast cells expressing a target-specific binding molecule can be identified by flow cytometry using directly or indirectly labelled target molecule. For example biotinylated target can be added to cells and binding to the cell surface can be detected with streptavidin-phycoerythrin. Within a population it becomes possible, using limiting target concentrations, to distinguish those clones which express higher affinity binding molecules since these clones will capture more target molecules and will therefore exhibit brighter fluorescence. Typically, each yeast cell will display 10,000 to 100,000 copies of a single scFv on the surface of the cell. To control for variation in scFv surface expression in different cells Chao et al used a fluorescently labelled anti-tag antibody to measure antibody expression level on the surface of each cell allowing normalisation for variation in expression level. This approach therefore allows yeast cells displaying high affinity binding molecules to be differentiated from those cells expressing high levels of a lower affinity antibody. Thus using fluorescence activated cell sorting (FACS) it is possible to separate cell clones according to the affinity and/or expression level of the encoded binding molecule.
[0010] Eukaryotic systems have also proven to be more effective than prokaryotic systems for the display of multi-chain antibody fragments and in particular with larger fragments such as full IgGs, FAbs or fusions of scFv with Fc domains (scFv-Fc fusions). Bead-based or flow sorting-based methods as described above for yeast cells could also be used to select antibodies from display libraries based on higher eukaryotes such as mammalian cells. The ability to format display libraries and select directly as IgGs, Fabs or as scFv-Fc fusions in mammalian cells would be a further advantage over yeast display. The glycosylation, expression and secretion machinery of bacterial and yeast cells is different from higher eukaryotes giving rise to antibodies with different post-translational modifications than those produced in mammalian cells. Since the manufacture of antibodies for research, diagnostic and therapeutic application is typically carried out in mammalian cells, display on mammalian cells (or other higher eukaryotic cells such as invertebrate, avian or plant cell lines) could give a better indication of potential issues or benefits for downstream manufacturing, e.g., identifying clones with optimal expression properties. In addition, antibodies discovered within the context of display on higher eukaryotes and particularly mammalian cells could be applied directly into cell-based reporter assays without extensive purification and without the complicating effect of contaminants from bacteria and yeast cells. Further, libraries of binders could be expressed directly in eukaryotic reporter cells such as mammalian cells to identify clones which directly affect cellular phenotype.
[0011] Despite the above advantages promised by eukaryotic display libraries, there remain significant problems with creation of libraries of binders in eukaryotic cells, especially higher eukaryotic cells. Introduction of a repertoire of exogenous genes ("transgenes") for expression in higher eukaryotes is more difficult than in yeast and bacteria. The cells of higher eukaryotes are more difficult to handle and scale up and transformation efficiencies are lower. Typical library sizes achieved are much smaller. In addition, introduced DNA integrates randomly within the genome leading to position effect variegation. Further, donor DNA introduced into mammalian cells by standard transfection or electroporation methods integrates as a linear array with variable copy number of the transfected transgene. The introduction of DNA encoding a repertoire of antibody genes therefore has the potential to introduce multiple antibody genes into each cell resulting in expression of multiple distinct antibodies per cell. In addition the presence of multiple antibody genes will reduce the relative expression of any given antibody and will lead to the isolation of many passenger antibody genes reducing the rate of enrichment of specific clones.
[0012] Although display of a library of binders on the surface of higher eukaryotes is more challenging, some examples have previously been described. In an early publication using mammalian display of IgGs derived from human immunisation, 3 rounds of selection (involving transient transfection, cell sorting, DNA recovery and re-transfection) were required to achieve a 450 fold enrichment of antigen-specific cells, averaging 7.6 fold enrichment per round [10]. Similarly transient expression from immunised libraries expressed within episomally-replicating vectors has also been described with antibodies formatted as scFvs [11, 12] or IgGs [13].
[0013] A number of approaches have been described to introduce a single or limited number of antibody genes into each cell. This includes dilution of DNA or mixing with carrier DNA [13] but this is a relatively uncontrolled method for managing copy number of introduced genes and reducing DNA input will have a detrimental effect on library size. Introduction of antibody genes by viral vectors has provided another solution to control the introduction of multiple antibody genes per cell. A cell surface display library has been generated in this way from several hundred human B lymphocytes generated by immunization and further enriched by flow sorting of antigen-specific B cells [14]. The antibody genes from this enriched pool were formatted as scFvs, cloned into a Sindbis alphavirus expression system and introduced into BHK cells using a low multiplicity of infection.
[0014] Breous-Nystrom et al. [15] used sequential retroviral infection to introduce a limited repertoire of 91 V kappa antibody genes followed by a heavy chain genes repertoire from 6 healthy donors into a murine pre-B cell line (1624-5). Infectious retrovirus was generated using the V-Pack system based on Moloney Murine Leukemia Virus (Stratagene). In order to bias towards single copy insertions, a multiplicity of infection was chose which led to infection of approximately 5% of cells. A major disadvantage of these approaches is that integration within the genome is random, leading to potential variation in transcription level based on the transcriptional activity of the site of integration. Another disadvantage in all these cases is that the integration of the antibody genes is controlled by limited infection or transfection which impacts on library size.
[0015] Site-specific integration of transgenes directed by recombinases has previously been described. Recombinases are enzymes that catalyse exchange reactions between DNA molecules containing enzyme-specific recognition sequences. For example Cre recombinase (derived from the site specific recombination system of E. coli) or Flp recombinase (utilising a recombination system of Saccharomyces cerevisiae) act on their specific 34 bp loxP recognition sites and 34 bp Flp Recombination Target (FRT) site respectively [16]. Recombinases have mainly been used in cellular engineering to catalyse site-specific integration. A number of studies from the work of Chen Zhou [17, 18, U.S. Pat. No. 7,884,054] have described the recombinase-mediated site-specific integration of antibody genes into the genome of mammalian cells using Flp recombinase within the "Flp-In" system, (http://tools.lifetechnologies.com/content/sfs/manuals/flpinsystem_man.pd- f). The Flp-In system utilises a variety of cell lines which have previously had a single FRT site introduced within their genome. By expressing the enzyme Flp recombinase it is possible to direct integration of expression plasmids, incorporating a FRT recombination site, into this pre-integrated FRT site in target cells.
[0016] Using the Flp-In system Zhou et al. [17] introduced an incoming antibody expression plasmid containing a FRT site into Chinese Hamster Ovary (CHO) cell line incorporating a FRT site (CHOF cells). Their work describes construction of a display library where 4 residues within an existing anti-OX40 ligand antibody were mutagenised. The library was screened using FACS to identify antibodies with anti-ligand affinity on the cell surface. The overall success in generating improved antibodies was limited to the isolation of a single improved antibody. The number of unique mammalian cell clones achieved was not reported.
[0017] A follow-on paper by Li et al. in 2012 [18] utilised lymphocytes from a hepatitis B patient to construct an antibody display library. Separate libraries were produced with the heavy and light chain genes obtained from a donor who had been immunised with HBsAg, individually reported to be libraries of size 1.02.times.10.sup.6 and 1.78.times.10.sup.5, respectively. A secondary library was then produced including both the heavy and light chains which reportedly had a size of 4.32.times.10.sup.5. FACS analysis reportedly indicated that about 40% of the cells displayed detectable full-length antibodies on the cell surface. FACS screening of the library identified antibodies binding to HBsAg. Of a sample of 8 selected library members which bound to the antigen, six were found to have the same antibody, so in total three unique anti-HBsAg clones were identified.
[0018] The rather limited success of this work may be due to the fact that the Flp-In system is designed for accurate integration in a limited number of clones rather than large library construction. There is therefore a potential conflict between achieving fidelity of integration versus achieving maximal library size. The Flp-In system utilises a mutant Flp recombinase in the plasmid pOG44 which possesses only 10% of the activity at 37.degree. C. of the native Flp recombinase [19]. A variant of Flp recombinase (Flpe) with better thermostability and higher activity than wild type has been identified [19, 20]. This was further improved by codon optimization to create Flp.sub.o encoded within plasmid cCAGGS-Flp.sub.o (Genebridges Cat. A203) According to the Flp-In manual however: [0019] "When generating Flp-In.TM. expression cell lines, it is important to remember that you are selecting for a relatively rare recombination event since you want recombination and integration of your pcDNA.TM.5/FRT construct to occur only through the FRT site and for a limited time. In this case, using a highly inefficient Flp recombinase is beneficial and may decrease the occurrence of other undesirable recombination events . . . . [0020] . . . To increase the likelihood of obtaining single integrants, you will need to lower the transfection efficiency by limiting the amount of plasmid DNA that you transfect" This is echoed by Buchholz et al., 1996 [19]: [0021] "FLP may be particularly useful for applications that do not rely on efficiency but depend on tight regulation".
[0022] In model experiments and using "instructions described in the manual", Zhou et al. (2010) [17] indeed demonstrated that single copy insertions occurred in >90% of clones. In library construction however relatively high amounts of expression plasmid (2.5-3.2 .mu.g per 10.sup.6 cells) and a donor excess over pOG44 recombinase-encoding plasmid was used [17, 18]. The Flp-In system recommends using a ratio of at least 9:1 in favour of the recombinase encoding plasmid versus the expression plasmid. However, when seeking to increase library size by transfecting larger amounts of DNA there is the potential for random integration of the incoming plasmid [21]. In all studies the accuracy of integration and the number of integrants per cell under "library construction" conditions was not reported.
[0023] In nuclease-directed integration of genes a site-specific nuclease is used to cleave cellular DNA at a specific location. It has previously been shown that this enhances the rate of homologous recombination by at least 40,000 fold and also allows repair by non-homologous end-joining mechanisms. This enhancement of site-specific integration has not previously been used or contemplated to solve the problems associated with creating libraries of binders.
[0024] US20100212035 describes methods for generation of rodents capable of expressing exogenous antibody by targeting the immunoglobulin locus of a mammalian embryo with a meganuclease to direct integration of a donor DNA. The potential to create variant libraries of meganucleases to create new DNA cleavage specificities is described but it his does not contemplate the use of meganucleases towards the generation of libraries of binders.
[0025] WO 2013/190032 A1 describes integration of genes into a specific locus (FerlL4) previously modified with exogenous DNA ("a site specific integration" SSI host cell) to incorporate recombinase sites, such as loxP and FRT sites for recombinase-mediated site-specific gene introduction. Nuclease-directed library generation is not described.
[0026] WO 2012/167192 A2 describes targetting genes to a locus that can then be selected for amplification. Nuclease-directed methods are employed to target the locus. Nuclease-directed library generation is not described.
[0027] US 2009/0263900A1 describes DNA molecules comprising homology arms and their use in methods of homologous recombination. Nuclease-directed library generation is not described.
[0028] WO 2011/100058 describes methods for integration of nucleic acid into a genome that avoids the need for long homology arms and instead relies on microhomology or "sticky ends" on the genome and donor to help direct integration. Nuclease-directed library generation is not described.
[0029] WO 2011/090804 describes methods for integration of multiple genes or multiple copies of the same gene using different zinc finger nucleases (ZFNs) in sequential rounds. Nuclease-directed library generation is not described.
[0030] WO2014/039872 describes methods for engineering plant cells, incorporating a "landing site" into which donor DNA is integrated by homologous recombination or non-homologous end joining using site-directed nucleases. Bacterial artificial chromosome (BAC) libraries are used for initial cloning of donor DNA. Libraries are mentioned in relation to Illumina sequencing methods. Nuclease-directed library generation is not described.
[0031] WO2007/047859 A2 describes methods for engineering specificity of meganucleases and their used to target genomic loci. Libraries of mutant meganucleases that may contain meganucleases with new nuclease specificity are described. Nuclease-directed library generation is not described.
[0032] US2014/0113375 A1 describes a transient expression system for generation single-stranded DNA sequences homologous to a target genomic sequence, which can be transported to the nucleus to alter the genetic information of the target genomic sequence via DNA repair pathways or homologous recombination. It is suggested that a "library" of mutations could be created by low fidelity reverse transcription of the introduced (non-library) DNA. Mammalian display and selection of molecules with binding activity is not described.
[0033] US2012/0277120 describes methods and compositions for the simultaneous integration of a plurality of exogenous nucleic acids is in a single transformation reaction using the native homologous recombination machinery in yeast, which recombination may be further enhanced by inducing targeted double-strand breaks in the host cell's genome at the intended sites of integration. The methods are intended to overcome the need for multiple rounds of engineering to integrate multiple DNA assemblies, for example, for the construction of functional metabolic pathways in industrial microbes, such as yeast. The display or expression of libraries of binding molecules, the use of higher eukaryotes and the selection of molecules with binding activity is not described.
[0034] To fully realize the potential for antibody display on mammalian cells and other higher eukaryotes there is a need for a system to create large libraries which combine accurate integration into a pre-defined site with an efficiency that allows construction of large libraries.
SUMMARY OF THE INVENTION
[0035] We have overcome the problem of creating large libraries of binders encompassing one or two binder genes per cell by using nuclease-directed integration of populations of genes encoding binders. The invention thus allows preparation of populations of eukaryotic cells wherein a repertoire of binder-encoding is integrated into a fixed locus in the genome allowing expression of the encoded binding molecule, thereby creating a population of cells expressing different binders.
[0036] The present invention relates to methods of producing eukaryotic cell libraries encoding a repertoire of binding molecules ("binders"), wherein the methods use a site-specific nuclease for targeted cleavage of cellular DNA to enhance site-specific integration of binder genes through endogenous cellular repair mechanisms. Site-specific nucleases permit the accurate introduction of donor DNA encoding binder molecules into one or more defined loci within the eukaryotic genome or other eukaryotic cell DNA. The invention provides methods of preparing populations of eukaryotic cells in which a repertoire of genes encoding binders are integrated into a desired locus in cellular DNA (e.g., a genomic locus) allowing expression of the encoded binding molecule, thereby creating a population of cells expressing different binders.
[0037] Construction of libraries of binders within eukaryotic cells according to the present invention has advantages over recombinase-directed approaches for site-directed incorporation of expression constructs. The present invention uses cellular DNA cleavage by site-specific nucleases to solve problems previously associated with construction of large repertoires of binder genes in eukaryotic cells and particularly higher eukaryotes. This invention allows the efficient creation of large populations of cell clones each expressing individual binders integrated at a fixed locus in cellular DNA. From these libraries of cellular clones it becomes possible to isolate genes encoding novel binding or function-modifying proteins and peptides.
[0038] Rather than recombinase-directed exchange of DNA, the approach of the present invention utilises site-specific cleavage of cellular (e.g., genomic) DNA followed by the use of natural repair mechanisms to integrate binder-encoding donor DNA. Following cleavage of the cellular DNA at a sequence recognised by the site-specific nuclease ("recognition sequence"), breaks in the cellular DNA are repaired using mechanisms such as homologous recombination or non-homologous end joining (NHEJ). Creation of site-specific breaks in the cellular DNA enhances incorporation of exogenous donor DNA allowing the construction of large populations of cells with binder genes integrated at a fixed locus.
[0039] To date, site-specific nucleases such as meganucleases, ZFNs, TALE nucleases and CRISPR/Cas systems have been directed towards the efficient creation of cells with modifications to endogenous genes or for introduction of reporter genes for the study of cell function. There are also instances where nuclease-directed genomic targeting has been used to integrate genes encoding single secreted antibodies for antibody production (by purification from culture medium) [21, 22,].
[0040] The invention simplifies construction of large libraries while directing integration to a single or limited number of defined genetic loci. Integration of donor DNA at one or more fixed loci normalises transcription compared with random integration of variable numbers of transgenes, and allows selection of antibody clones on the basis of translational and stability properties of the binder itself. Faithful integration of donor DNA at a pre-determined location or locations in the cellular DNA results in relatively uniform levels of transcription of binders in the library, and high efficiency of donor DNA introduction, make cell populations created by the methods of the invention particularly useful as libraries for display and selection of binders. Methods of the invention thus produce high quality libraries of binders in eukaryotic cells, which can be screened to identify cells encoding and expressing a specific binder for a target of interest.
[0041] In various aspects the invention relates to new and improved methods of preparing eukaryotic cell libraries, the libraries themselves, isolation of desired binders, encoding nucleic acid and cells from the libraries, and uses of the libraries such as for expression and screening of binding molecules and for screening for the effects of binding molecules. Various methods will be described for producing libraries in vitro and using libraries in vitro or in vivo.
[0042] The invention provides a method of producing a library of eukaryotic cell clones containing DNA encoding a diverse repertoire of binders, the method comprising using a site-specific nuclease to target cleavage of eukaryotic cell DNA to enhance site-specific integration of binder genes into the cellular DNA through endogenous cellular DNA repair mechanisms.
[0043] A method of producing a library of eukaryotic cell clones containing DNA encoding a diverse repertoire of binders may comprise:
[0044] providing donor DNA molecules encoding the binders, and eukaryotic cells,
[0045] introducing the donor DNA into the cells and providing a site-specific nuclease within the cells, wherein the nuclease cleaves cellular DNA to create an integration site at which the donor DNA becomes integrated into the cellular DNA, integration occurring through DNA repair mechanisms endogenous to the cells.
[0046] For multimeric binders comprising at least a first and second subunit (i.e., separate polypeptide chains, such as antibody VH and VL domains presented within a Fab or IgG format), the multiple subunits may be encoded on the same molecule of donor DNA. However, it may be desirable to integrate the different subunits into separate loci, in which case the subunits can be provided on separate donor DNA molecules. These could be integrated within the same cycle of nuclease-directed integration or they may be integrated sequentially using nuclease-directed integration for one or both integration steps.
[0047] Methods of producing libraries of eukaryotic cell clones encoding multimeric binders may comprise:
[0048] providing eukaryotic cells containing DNA encoding the first subunit, and providing donor DNA molecules encoding the second binder subunit,
[0049] introducing the donor DNA into the cells and providing a site-specific nuclease within the cells, wherein the nuclease cleaves a recognition sequence in cellular DNA to create an integration site at which the donor DNA becomes integrated into the cellular DNA, integration occurring through DNA repair mechanisms endogenous to the cells, thereby creating recombinant cells which contain donor DNA integrated in the cellular DNA. These recombinant cells will contain DNA encoding the first and second subunits of the multimeric binder, and may be cultured to express both subunits. Multimeric binders are obtained by expression and assembly of the separately encoded subunits.
[0050] In the above example, nuclease-directed integration is used to integrate DNA encoding a second subunit into cells already containing DNA encoding a first subunit. The first subunit could be previously introduced using the techniques of the present invention or any other suitable DNA integration method. An alternative approach is to use nuclease-directed integration in a first cycle of introducing donor DNA, to integrate a first subunit, followed by introducing the second subunit either by the same approach or any other suitable method. If the nuclease-directed approach is used in multiple cycles of integration, different site-specific nucleases may optionally be used to drive nuclease-directed donor DNA integration at different recognition sites. A method of generating the library may comprise:
[0051] providing first donor DNA molecules encoding the first subunit, and providing eukaryotic cells,
[0052] introducing the first donor DNA into the cells and providing a site-specific nuclease within the cells, wherein the nuclease cleaves a recognition sequence in cellular DNA to create an integration site at which the donor DNA becomes integrated into the cellular DNA, integration occurring through DNA repair mechanisms endogenous to the cells, thereby creating a first set of recombinant cells containing first donor DNA integrated in the cellular DNA,
[0053] culturing the first set of recombinant cells to produce a first set of clones containing DNA encoding the first subunit,
[0054] introducing second donor DNA molecules encoding the second subunit into cells of the first set of clones, wherein the second donor DNA is integrated into cellular DNA of the first set of clones, thereby creating a second set of recombinant cells containing first and second donor DNA integrated into the cellular DNA, and
[0055] culturing the second set of recombinant cells to produce a second set of clones, these clones containing DNA encoding the first and second subunits of the multimeric binder,
[0056] thereby providing a library of eukaryotic cell clones containing donor DNA encoding the repertoire of multimeric binders.
[0057] Site-specific integration of donor DNA into cellular DNA creates recombinant cells, which can be cultured to produce clones. Individual recombinant cells into which the donor DNA has been integrated are thus replicated to generate clonal populations of cells--"clones"--each clone being derived from one original recombinant cell. Thus, the method generates a number of clones corresponding to the number of cells into which the donor DNA was successfully integrated. The collection of clones form a library encoding the repertoire of binders (or, at an intermediate stage where binder subunits are integrated in separate rounds, the clones may encode a set of binder subunits). Methods of the invention can thus provide a library of eukaryotic cell clones containing donor DNA encoding the repertoire of binders.
[0058] Methods of the invention can generate libraries of clones containing donor DNA integrated at a fixed locus, or at multiple fixed loci, in the cellular DNA. By "fixed" it is meant that the locus is the same between cells. Cells used for creation of the library may therefore contain a nuclease recognition sequence at a fixed locus, representing a universal landing site in the cellular DNA at which the donor DNA can integrate. The recognition sequence for the site-specific nuclease may be present at one or more than one position in the cellular DNA.
[0059] Libraries produced according to the present invention may be employed in a variety of ways. A library may be cultured to express the binders, thereby producing a diverse repertoire of binders. A library may be screened for a cell of a desired phenotype, wherein the phenotype results from expression of a binder by a cell. Phenotype screening is possible in which library cells are cultured to express the binders, followed by detecting whether the desired phenotype is exhibited in clones of the library. Cellular read-outs can be based on alteration in cell behaviour such as altered expression of endogenous or exogenous reporter genes, differentiation status, proliferation, survival, cell size, metabolism or altered interactions with other cells. When the desired phenotype is detected, cells of a clone that exhibits the desired phenotype may then be recovered. Optionally, DNA encoding the binder is then isolated from the recovered clone, providing DNA encoding a binder which produces the desired phenotype when expressed in the cell.
[0060] A key purpose for which eukaryotic cell libraries have been used is in methods of screening for binders that recognise a target of interest. In such methods a library is cultured to express the binders, and the binders are exposed to the target to allow recognition of the target by one or more cognate binders, if present, and detecting whether the target is recognised by a cognate binder. In such methods, binders may be displayed on the cell surface and those clones of the library that display binders with desired properties can be isolated. Thus cells incorporating genes encoding binders with desired functional or binding characteristics could be identified within the library. The genes can be recovered and used for production of the binder or used for further engineering to create derivative libraries of binders to yield binders with improved properties.
[0061] The present invention offers advantages over previous approaches for construction of libraries in higher eukaryotes. Some studies have used lentiviral infection to introduce antibody genes into mammalian reporter cells [106]. This has the advantage that large libraries can be generated but there is no control over the site of integration and copy number is controlled by using a low multiplicity of infection (as discussed above). In an alternative approach antibody genes were introduced via homologous recombination, without the benefit of nuclease-directed integration and using homology arms of 10 kb but the efficiency of targeting was relatively low meaning that the potential library size was limited [105]. In contrast the use of sequence-directed nucleases retains the benefits of targeted integration to one or a few loci of choice while allowing efficient construction of large libraries. Nuclease-directed integration has the advantage that transgenes are targeted to a fixed locus or fixed loci within the cellular DNA. This means that promoter activity driving transcription of binder genes in all clones will be the same and the functionality of each binder will be a reflection of its inherent potency, translational efficiency and stability rather than being due to variation related to the integration site. Targeting to a single or limited number of loci will also enable better control of expression if required e.g., using inducible promoters.
[0062] Various features of the invention are further described below. It is noted that headings used throughout this specification are to assist navigation only and should not be interpreted as definitive, and that embodiments described in different sections may be combined as appropriate.
DETAILED DESCRIPTION
Eukaryotic Cells
[0063] The potential of populations of eukaryotic cells expressing a diverse repertoire of binders is exemplified and discussed in the Examples herein in relation to expression of antibody repertoires on the surface of mammalian cells. The benefits of the invention are not limited to mammalian cells and include all eukaryotes.
[0064] Yeast (e.g., Saccharomyces cerevisiae) has a smaller genome than mammalian cells and homologous recombination directed by homology arms (in the absence of nuclease-directed cleavage) is an effective way of introducing foreign DNA compared to higher eukaryotes. Thus, a particular benefit of nuclease-directed integration of the present invention relates to integration of binder genes into higher eukaryotic cells with larger genomes where homologous recombination in the absence of nuclease cleavage is less effective. Nuclease-directed integration has been used in yeast cells to solve the problem of efficient integration of multiple genes into individual yeast cells, e.g., for engineering of metabolic pathways (US2012/0277120), but this work does not incorporate introduction of libraries of binders nor does it address the problems of library construction in higher eukaryotes.
[0065] Libraries of eukaryotic cells according to the present invention are preferably higher eukaryotic cells, defined here as cells with a genome greater than that of Saccharomyces cerevisiae which has a genome size of 12.times.10.sup.6 base pairs (bp). The higher eukaryotic cells may for example have a genome size of greater than 2.times.10.sup.7 base pairs. This includes, for example, mammalian, avian, insect or plant cells. Preferably the cells are mammalian cells, e.g., mouse or human. The cells may be primary cells or may be cell lines. Chinese hamster ovary (CHO) cells are commonly used for antibody and protein expression but any alternative stable cell line may be used in the invention. HEK293 cells are used in Examples herein. Methods are available for efficient introduction of foreign DNA into primary cells allowing these to be used (e.g., by electroporation where efficiencies and viabilities up to 95% have been achieved http://www.maxcyte. com/technology/primary-cells-stem-cells.php).
[0066] T lymphocyte lineage cells (e.g., primary T cells or a T cell line) or B lymphocyte lineage cells are among the preferred cell types. Of particular interest are primary T-cells or T cell derived cell lines for use in TCR libraries including cell lines which lack TCR expression [23, 24, 25]. Examples of B lymphocyte lineage cells include B cells, pre-B cells or pro-B cells and cell lines derived from any of these.
[0067] Construction of libraries in primary B cells or B cell lines would be of particular value for construction of antibody libraries. Breous-Nystrom et al. [15] have generated libraries in a murine pre-B cell line (1624-5). The chicken B cell derived cell line DT40 (ATCC CRL-2111) has particular promise for construction of libraries of binders. DT40 is a small cell line with a relatively rapid rate of cell division. Repertoires of binders could be targeted to specific loci using ZFNs, TALE nucleases or CRISPR/Cas9 targeted to endogenous sequences or by targeting pre-integrated heterologous sites which could include meganuclease recognition sites. DT40 cells express antibodies and so it will be advantageous to target antibody genes within the antibody locus either with or without disruption of the endogenous chicken antibody variable domains. DT40 cells have also been used as the basis of an in vitro system for generation of chicken IgMs termed the Autonomously Diversifying Library system (ADLib system) which takes advantage of intrinsic diversification occurring at the chicken antibody locus. As a result of this endogenous diversification it is possible to generate novel specificities. The nuclease-directed approach described here could be used in combination with ADLib to combine diverse libraries of binders from heterologous sources (e.g., human antibody variable region repertoires or synthetically derived alternative scaffolds) with the potential for further diversification with the chicken IgG locus. Similar benefits could apply to human B cell lines such as Nalm6 [26]. Other B lineage cell lines of interest include lines such as the murine pre-B cell line 1624-5 and the pro-B cell line Ba/F3. Ba/F3 is dependent on IL-3 [27] and its use is discussed elsewhere herein. Finally a number of human cell lines could be used including those listed in the "Cancer Cell Line Encyclopaedia" [28] or "COSMIC catalogue of somatic mutations in cancer" [29].
[0068] Typically the library will be composed of a single type of cells, produced by introduction of donor DNA into a population of clonal eukaryotic cells, for example by introduction of donor DNA into cells of a particular cell line. The main significant difference between the different library clones will then be due to integration of the donor DNA.
Eukaryotic Viral Systems
[0069] The advantages of the system in creation of libraries of binders in eukaryotic cells could be applied to viral display systems based around eukaryotic expression systems, e.g., baculoviral display or retroviral display [1, 2, 3, 4]. In this approach each cell will encode a binder capable of being incorporated into a viral particle. In the case of retroviral systems the encoding mRNA would be packaged and the encoded binder would be presented on the cell surface. In the case of baculoviral systems, genes encoding the binder would need to be encapsulated into the baculoviral particle to maintain an association between the gene and the encoded protein. This could be achieved using host cells carrying episomal copies of the baculoviral genome. Alternatively integrated copies could be liberated following the action of a specific nuclease (distinct from the one used to drive site-specific integration). In the case of multimeric binder molecules some partners could be encoded within the cellular DNA with the genes for one or more partners being packaged within the virus.
Site-Specific Nuclease
[0070] The invention involves use of a site-specific nuclease for targeted cleavage of cellular DNA in the construction of a library of eukaryotic cells containing DNA encoding a repertoire of binders, wherein nuclease-mediated DNA cleavage enhances site-specific integration of binder genes through endogenous cellular DNA repair mechanisms. The site-specific nuclease cleaves cellular DNA following specific binding to a recognition sequence, thereby creating an integration site for donor DNA. The nuclease may create a double strand break or a single strand break (a nick). Cells used for creation of the library may contain endogenous sequences recognised by the site-specific nuclease or the recognition sequence may be engineered into the cellular DNA.
[0071] The site-specific nuclease may be exogenous to the cells, i.e., not occurring naturally in cells of the chosen type.
[0072] The site-specific nuclease can be introduced before, after or simultaneously with introduction of the donor DNA encoding the binder. It may be convenient for the donor DNA to encode the nuclease in addition to the binder, or on separate nucleic acid which is co-transfected or otherwise introduced at the same time as the donor DNA. Clones of a library may optionally retain nucleic acid encoding the site-specific nuclease, or such nucleic acid may be only transiently transfected into the cells.
[0073] Any suitable site-specific nuclease may be used with the invention. It may be a naturally occurring enzyme or an engineered variant. There are a number of known nucleases that are especially suitable, such as those which recognise, or can be engineered to recognise, sequences that occur only rarely in cellular DNA. Nuclease cleavage at only one or two sites is advantageous since this should ensure that only one or two molecules of donor DNA are integrated per cell. Rarity of the sequence recognised by the site-specific nuclease is more likely if the recognition sequence is relatively long. The sequence specifically recognised by the nuclease may for example be a sequence of at least 10, 15, 20, 25 or 30 nucleotides.
[0074] Examples of suitable nucleases include meganucleases, zinc finger nucleases (ZFNs), TALE nucleases, and nucleic acid-guided (e.g., RNA-guided) nucleases such as the CRISPR/Cas system. Each of these produces double strand breaks although engineered forms are known which generate single strand breaks.
[0075] Meganucleases (also known as homing endonucleases) are nucleases which occur across all the kingdoms of life and recognise relatively long sequences (12-40 bp). Given the long recognition sequence they are either absent or occur relatively infrequently in eukaryotic genomes. Meganucleases are grouped into 5 families based on sequence/structure. (LAGLIDADG, GIY-YIG, HNH, His-Cys box and PD-(D/E)XK). The best studied family is the LAGLIDADG family which includes the well characterised I-SceI meganuclease from Saccharomyces cerevisiae. I-SceI recognises and cleaves an 18 bp recognition sequence (5' TAGGGATAACAGGGTAAT) leaving a 4 bp 3' overhang. Another commonly used example is I-Cre1 which originates from the chloroplast of the unicellular green algae of Chlamydomonas reinhardtii, and recognizes a 22 bp sequence [30]. A number of engineered variants have been created with altered recognition sequences [31]. Meganucleases represent the first example of the use of site-specific nucleases in genome engineering [49, 50]. As with recombinase-based approaches, use of I-SceI and other meganucleases requires prior insertion of an appropriate recognition site to be targeted within the genome or engineering of meganucleases to recognize endogenous sites [30]. By this approach targeting efficiency in HEK293 cells (as judged by homology-directed "repair" of an integrated defective GFP gene) was achieved in 10-20% of cells through the use of I-SceI [32].
[0076] A preferred class of meganucleases for use in the present invention is the LAGLIDADG endonucleases. These include I-Sce I, I-Chu I, I-Cre I, Csm I, Pl-Sce I, PI-Tli I, PI-Mtu I, I-Ceu I, I-Sce II, I-Sce III, HO, Pi-Civ I, PI-Ctr I, PI-Aae I, PI-Bsu I, PI-Dha I, PI-Dra I, PI-Mav I, PI-Mch I, PI-Mfu PI-Mfl I, PI-Mga I, PI-Mgo I, PI-Min I, PI-Mka I, PI-Mle I, PI-Mma I, PI-Msh I, PI-Msm I, PI-Mth I, PI-Mtu PI-Mxe I, PI-Npu I, PI-Pfu I, PI-Rma I, Pl-Spb I, PI-Ssp I, PI-Fac I, PI-Mja I, PI-Pho I, Pi-Tag I, PI-Thy I, PI-Tko I, I-Msol, and PI-Tsp I; preferably, I-Sce I, I-Cre I, I-Chu I, I-Dmo I, I-Csm I, Pl-Sce I, PI-Pfu I, PI-Tli I, PI-Mtu I, and I-Ceu I.
[0077] In recent years a number of methods have been developed which allow the design of novel sequence-specific nucleases by fusing sequence-specific DNA binding domains to non-specific nucleases to create designed sequence-specific nucleases directed through bespoke DNA binding domains. Binding specificity can be directed by engineered binding domains such as zinc finger domains. These are small modular domains, stabilized by Zinc ions, which are involved in molecular recognition and are used in nature to recognize DNA sequences. Arrays of zinc finger domains have been engineered for sequence specific binding and have been linked to the non-specific DNA cleavage domain of the type II restriction enzyme Fok1 to create zinc finger nucleases (ZFNs). ZFNs can be used to create double stranded break at specific sites within the genome. Fok1 is an obligate dimer and requires two ZFNs to bind in close proximity to effect cleavage. The specificity of engineered nucleases has been enhanced and their toxicity reduced by creating two different Fok1 variants which are engineering to only form heterodimers with each other [33]. Such obligate heterodimer ZFNs have been shown to achieve homology-directed integration in 5-18% of target cells without the need for drug selection [21, 34, 35]. Incorporation of inserts up to 8 kb with frequencies of >5% have been demonstrated in the absence of selection.
[0078] It has recently been shown that single-stranded 5' overhangs created by nucleases such as ZFNs help drive efficient integration of transgenes to the sites of cleavage [45]. This has been extended to show that in vivo cleavage of donor DNA (through inclusion of a specific nuclease recognition site within the donor plasmid) enhances the efficiency on non-homologous integration. The mechanism is not entirely clear but it is possible that reduced exposure to cellular nucleases through in vivo linearisation may have contributed to the enhancement [45]. It is also possible that matches in the 5' overhangs of donor and acceptor DNA, generated by the nucleases drive ligation. Examination of sequences at the junctions however showed the occurrence of deletions. It is possible that perfectly matched junctions continue to act as substrate for the site-directed nucleases until deletion of the recognition sequence occurs. To overcome this potential problem, Maresca et al. [36] have inverted the recognition sites of left and right ZFNs within the donor DNA such that ligation of donor DNA into the genomic locus will lead to duplication of two left hand ZFNs on one flank of the integration and duplication of two right hand ZFNs at the other flank. The use of obligate heterodimer nucleases (as described for Fok1) means that neither of these newly created flanking sequences can be cleaved by the targeted nuclease.
[0079] The ability to engineer DNA binding domains of defined specificity has been further simplified by the discovery in Xanathomonas bacteria of Transcription activator-like effectors (TALE) molecules. These TALE molecules consist of arrays of monomers of 33-35 amino acids with each monomer recognising a single base within a target sequence [37]. This modular 1:1 relationship has made it relatively easy to design engineered TALE molecules to bind any DNA target of interest. By coupling these designed TALEs to Fok1 it has been possible to create novel sequence-specific TALE-nucleases. TALE nucleases, also known as TALENs, have now been designed to a large number of sites and exhibit high success rate for efficient gene modification activity [38]. In examples herein we demonstrate the enhanced integration of donor DNA through the use of TALE nucleases. Other variations and enhancements of TALE nuclease technology have been developed and could be used for the generation of libraries of binders through nuclease-directed integration. These included "mega-TALENs" where a TALE nuclease binding domain is fused to a meganuclease [39] and "compact TALENs" where a single TALE nuclease recognition domain is used to effect cleavage [40].
[0080] In recent years another system for directing double- or single-stranded breaks to specific sequences in the genome has been described. This system called "Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) and CRISPR Associated (Cas)" system is based on a bacterial defence mechanism [41]. The CRISPR/Cas system targets DNA for cleavage via a short, complementary single-stranded RNA (CRISPR RNA or crRNA) adjoined to a short palindromic repeat. In the commonly used "Type II" system, the processing of the targeting RNA is dependent on the presence of a trans-activating crRNA (tracrRNA) that has sequence complementary to the palindromic repeat. Hybridization of the tracrRNA to the palindromic repeat sequence triggers processing. The processed RNA activates the Cas9 domain and directs its activity to the complementary sequence within DNA. The system has been simplified to direct Cas9 cleavage from a single RNA transcript and has been directed to many different sequences within the genome [42, 43]. This approach to genome cleavage has the advantage of being directed via a short RNA sequence making it relatively simple to engineer cleavage specificity. Thus there are a number of different ways to achieve site-specific cleavage of genomic DNA. As described above this enhances the rate of integration of a donor plasmid through endogenous cellular DNA repair mechanisms.
[0081] Use of meganucleases, ZFNs, TALE nuclease or nucleic acid guided systems such as the CRISPR/Cas9 systems will enable targeting of endogenous loci within the genome. In the Examples herein we have demonstrated targeting to the AAVS locus but alternative loci could be targeted. For example the Type I collagen gene locus has been used for efficient transgene expression [44].
[0082] Alternatively heterologous recognition sites for targeted nucleases, including meganucleases, ZFNs and TALE nucleases could be introduced in advance for subsequent library targeting. In Examples herein, we describe the use of a TALE nuclease recognising a sequence within the AAVS locus to introduce by homologous recombination, an I-Scel meganuclease recognition sequence and heterologous TALE nuclease recognition sites within the AAVS locus. Nuclease-directed targeting could be used to drive insertion of target sequences by homologous recombination or NHEJ using vector DNA or even double stranded oligonucleotides [45]. As an alternative, non-specific targeting methods could be used to introduce targeting sites through the use of transposon-directed integration [46] to introduce recognition sites for site-specific nucleases. Viral-based systems, such as lentivirus, applied at low titre could also be used to introduce targeting sites. Transfection of DNA coupled with screening for single copy insertion has also been used to identify unique integration sites [17]. Such non-specific approaches would be particularly useful in the case of cells which do not have an obvious site to target or for genomes which have not been sequenced or for genomes for which no existing TALE nucleases, ZFNs or Cas9/CRISPR systems are available. Once a cell line has been established following random insertion of a nuclease recognition site, the cell line can be used subsequently to create libraries of binders where all clones of the library contain the transgene at the fixed locus using nuclease-directed integration.
[0083] In the Examples presented, three different plasmids are used encompassing pairs of TALE nucleases or ZFNs on individual plasmids with a separate plasmid for donor DNA. In the case of meganuclease the site-specific nuclease is encoded by a single gene and this is introduced on one plasmid with the donor DNA present on a second plasmid. Of course, combinations could be used incorporating two or more of these elements on the same plasmid and this could enhance the efficiency of targeting by reducing the number of number of plasmids to be introduced. In addition it may be possible to pre-integrate the nuclease(s) which could also be inducible to allow temporal control of nuclease activity as has been demonstrated for transposases [46]. Finally the nuclease could be introduced as recombinant protein or protein:RNA complex (for example in the case of an RNA directed nuclease such as CRISPR:Cas9).
Locus
[0084] A recognition sequence for the site-specific nuclease may be present in genomic DNA, or episomal DNA which is stably inherited in the cells. Donor DNA may therefore be integrated at a genomic or episomal locus in the cellular DNA.
[0085] In its simplest form a single gene encoding a binder (binder gene) is targeted to a single site within the eukaryotic genome. Identification of a cell demonstrating a particular binding activity or cellular phenotype will allow direct isolation of the gene encoding the desired property (e.g., by PCR from mRNA or genomic DNA). This is facilitated by using a unique recognition sequence for the site-specific nuclease, occurring once in the cellular DNA. Cells used for creation of the library may thus contain a nuclease recognition sequence at a single fixed locus, i.e., one identical locus in all cells. Libraries produced from such cells will contain donor DNA integrated at the fixed locus, i.e., occurring at the same locus in cellular DNA of all clones in the library.
[0086] Optionally, recognition sequences may occur multiple times in cellular DNA, so that the cells have more than one potential integration site for donor DNA. This would be a typical situation for diploid or polyploid cells where the recognition sequence is present at corresponding positions in a pair of chromosomes, i.e., replicate loci. Libraries produced from such cells may contain donor DNA integrated at replicate fixed loci. For example libraries produced from diploid cells may have donor DNA integrated at duplicate fixed loci and libraries produced from triploid cells may have donor DNA integrated at triplicate fixed loci. Many suitable mammalian cells are diploid, and clones of mammalian cell libraries according to the invention may have donor DNA integrated at duplicate fixed loci.
[0087] The sequence recognised by the site-specific nuclease may occur at more than one independent locus in the cellular DNA. Donor DNA may therefore integrate at multiple independent loci. Libraries of diploid or polyploid cells may comprise donor DNA integrated at multiple independent fixed loci and/or at replicate fixed loci.
[0088] In cells containing recognition sequences at multiple loci (whether replicate or independent loci), each locus represents a potential integration site for a molecule of donor DNA. Introduction of donor DNA into the cells may result in integration at the full number of nuclease recognition sequences present in the cell, or the donor DNA may integrate at some but not all of these potential sites. For example, when producing a library from diploid cells containing recognition sequences at first and second fixed loci (e.g., duplicate fixed loci), the resulting library may comprise clones in which donor DNA is integrated at the first fixed locus, clones in which donor DNA is integrated at the second fixed locus, and clones in which donor DNA is integrated at both the first and second fixed loci.
[0089] Methods of producing libraries may therefore involve site-specific nuclease cleavage of multiple fixed loci in a cell, and integration of donor DNA at the multiple fixed loci. As noted above, in cases where there are multiple copies of the same recognition sequence (e.g., as occurs when targeting endogenous loci in diploid or polyploid cells) it is possible that two binder genes will be integrated, particularly when an efficient targeting mechanisms is used, with only one gene being specific to the target. This can be resolved during subsequent screening once binder genes have been isolated.
[0090] In some instances it may be desirable to introduce more than one binder per cell. For example bi-specific binders could be generated from two different antibodies coming together and these may have properties absent in the individual binders [47]. This could be achieved by introducing different antibody genes into both alleles at duplicate fixed loci or by targeting different antibody populations into independent fixed loci using the methods described herein. Furthermore a binder may itself be composed of multiple chains (e.g., antibody VH and VL domains presented within a Fab or IgG format). In this case it may be desirable to integrate the different sub-units into different loci. These could be integrated within the same cycle of nuclease-directed integration, they could be integrated sequentially using nuclease-directed integration for one or both integration steps.
Introduction of Donor DNA
[0091] Numerous methods have been described for introducing donor DNA into eukaryotic cells, including transfection, infection or electroporation. Transfection of large numbers of cells is possible by standard methods including polyethyleneimine-mediated transfection as described herein. In addition methods are available for highly efficient electroporation of 10.sup.10 cells in 5 minutes, e.g., http://www.maxcyte.com.
[0092] Combinatorial libraries could be created wherein members of multimeric binding pairs (e.g., VH and VL genes of antibody genes) or even different parts of the same binder molecule are introduced on different plasmids. Introduction of separate donor DNA molecules encoding separate binders or binder subunits may be done simultaneously or sequentially. For example an antibody light chain could be introduced by transfection or infection, the cells grown up and selected if necessary. Other components could then be introduced in a subsequent infection or transfection step. One or both steps could involve nuclease-directed integration to specific genomic loci.
Integration of Donor DNA
[0093] The donor DNA is integrated into the cellular DNA, forming recombinant DNA having a contiguous DNA sequence in which the donor DNA is inserted at the integration site. In the present invention, integration is mediated by the natural DNA repair mechanisms that are endogenous to the cell. Thus, integration can be allowed to occur simply by introducing the donor DNA into a cell, allowing the site-specific nuclease to create an integration site, and allowing the donor DNA to be integrated. Cells may be kept in culture for sufficient time for the DNA to be integrated. This will usually result in a mixed population of cells, including (i) recombinant cells into which the donor DNA has integrated at the integration site created by the site-specific nuclease, and optionally (ii) cells in which donor DNA has integrated at sites other than the desired integration site and/or optionally (iii) cells that into which donor DNA has not integrated. The desired recombinant cells and the resulting clones of the library may thus be provided in a mixed population of other eukaryotic cells. Selection methods described elsewhere herein may be used to enrich for cells of the library.
[0094] Endogenous DNA repair mechanisms in eukaryotic cells include homologous recombination, non-homologous end joining (NHEJ) and microhomology-directed end joining. The efficiency of DNA modification by such processes can be increased by the introduction of double stranded breaks (DSBs) in the DNA and efficiency gains of 40,000 fold have been reported using rare cutting endonucleases (meganucleases) such as I-SceI [48, 49, 50].
[0095] Unlike the site-specific recombination involved in systems such as the Flp-In system [16], the present invention does not require exogenous recombinases or engineered recombinase recognition sites. Therefore, optionally the present invention does not include a step of recombinase-mediated DNA integration in creating the library, and/or optionally the eukaryotic cells into which the donor DNA is introduced lack a recombination site for a site-specific recombinase. The mechanisms and practicalities of directed insertion of donor DNA into cellular DNA by recombinases and nucleases are very distinct. As discussed by Jasin 1996 [50]:
[0096] " . . . the reaction catalyzed by site-specific recombinases is quite distinct from cellular repair of DSBs. Site-specific recombinases, such as cre, synapse two recognition sites and create single-strand breaks within the sites, thus forming Holliday intermediates. The intermediates are resolved to produce deletions, inversions and insertions (cointegrants), all of which restore the two recognition sites. The reaction is absolutely precise and, hence, reversible. The breaks are never exposed to the cellular repair machinery."
[0097] In contrast site-specific nuclease act to create breaks or nicks within the cellular DNA (e.g., genomic or episomal), which are exposed to and repaired by endogenous cellular repair mechanisms such as homologous recombination or NHEJ. Recombinase-based approaches have an absolute requirement for pre-integration of their recognition sites, so such methods require engineering of the "hot spot" integration site into the cellular DNA as a preliminary step. With nuclease-directed integration it is possible to engineer nucleases or direct via guide RNA in the case of CRISPR:Cas9 to recognise endogenous loci, i.e., nucleic acid sequences occurring naturally in the cellular DNA. Finally, at a practical level nuclease-directed approaches are more efficient for direct integration of transgenes at the levels required to make large libraries of binders.
[0098] The DNA repair mechanism by which the donor DNA is integrated in methods of the invention can be pre-determined or biased to some extent by design of the donor DNA and/or choice of site-specific nuclease.
[0099] Homologous recombination is a natural mechanism used by cells to repair double stranded breaks using homologous sequence (e.g., from another allele) as a template for repair. Homologous recombination has been utilised in cellular engineering to introduce insertions (including transgenes), deletions and point mutations into the genome. Homologous recombination is promoted by providing homology arms on the donor DNA. The original approach to engineering higher eukaryotic cells typically used homology arms of 5-10 kb within a donor plasmid to increase efficiency of targeted integration into the site of interest. Despite this, homologous recombination driven purely by long homology arms, is less efficient than Flp and Cre directed recombination particularly in higher eukaryotes with large genomes. Homologous recombination is particularly suitable for eukaryotes such as yeast, which has a genome size of only 12.5.times.10.sup.6 bp, where it is more effective compared with higher eukaryotes with larger genomes e.g., mammalian cells with 3000.times.10.sup.6 bp.
[0100] Homologous recombination can also be directed through [52] nicks in genomic DNA and this could also serve as a route for nuclease-directed integration into genomic DNA. Two distinct pathways have been shown to promote homologous recombination at nicked DNA. One is essentially similar to repair at double strand breaks, utilizing Rad51/Brca2, while the other is inhibited by Rad51/Brca2 and preferentially uses single-stranded DNA or nicked double stranded donor DNA [51].
[0101] Non homologous end-joining (NHEJ) is an alternative mechanism to repair double stranded breaks in the genome where the ends of DNA are directly re-ligated without the need for a homologous template. Nuclease-directed cleavage of genomic DNA can also enhance transgene integration via non-homology based mechanisms. This approach to DNA repair is less accurate and can lead to insertions or deletions. NHEJ nonetheless provides a simple means of integrating in-frame exons into intron or allows integration of promoter:gene cassettes into the genome. Use of non-homologous methods allows the use of donor vectors which lack homology arms thereby simplifying the construction of donor DNA.
[0102] It has been pointed out that short regions of terminal homology are used to re-join DNA ends and it was hypothesized that 4 bp of microhomology might be utilized for directing repairing at double strand breaks, referred to as microhomology-directed end joining [50].
Donor DNA
[0103] The donor DNA will usually be circularised DNA, and may be provided as a plasmid or vector. Linear DNA is another possibility. Donor DNA molecules may comprise regions that do not integrate into the cellular DNA, in addition to one or more donor DNA sequences that integrate into the cellular DNA. The DNA is typically double-stranded, although single-stranded DNA may be used in some cases. The donor DNA contains one or more transgenes encoding a binder, for example it may comprise a promoter:gene cassette.
[0104] In the simplest format double-stranded, circular plasmid DNA can be used to drive homologous recombination. This requires regions of DNA flanking the transgenes which are homologous to DNA sequence flanking the cleavage site in genomic DNA. Linearised double-stranded plasmid DNA or PCR product or synthetic genes could be used to drive both homologous recombination and NHEJ repair pathways. As an alternative to double-stranded DNA it is possible to use single-stranded DNA to drive homologous recombination [52]. A common approach to generating single-stranded DNA is to include a single-stranded origin of replication from a filamentous bacteriophage into the plasmid.
[0105] Single-stranded DNA viruses such as adeno-associated virus (AAV) have been used to drive efficient homologous recombination where the efficiency has been shown to be improved by several orders of magnitude [53, 54]. Systems such as the AAV systems could be used in conjunction with nuclease-directed cleavage for the construction of large libraries of binders. The benefits of both systems could be applied to targeting of libraries of binders. The packaging limit of AAV vectors is 4.7 kb but the use of nuclease digestion of target genomic DNA will reduce this allowing larger transgene constructs to be incorporated.
[0106] A molecule of donor DNA may encode a single binder or multiple binders. Optionally, multiple subunits of a binder may be encoded per molecule of donor DNA. In some embodiments, donor DNA encodes a subunit of a multimeric binder.
Promoters and Genetic Elements for Selection
[0107] Transcription of the binder from the encoding donor DNA will usually be achieved by placing the sequence encoding the binder under control of a promoter and optionally one or more enhancer elements for transcription. A promoter (and optionally other genetic control elements) may be included in the donor DNA molecule itself. Alternatively, the sequence encoding the binder may lack a promoter on the donor DNA, and instead may be placed in operable linkage with a promoter on the cellular DNA, e.g., an endogenous promoter or a pre-integrated exogenous promoter, as a result of its insertion at the integration site created by the site-specific nuclease.
[0108] Donor DNA may further comprise one or more further coding sequences, such as genetic elements enabling selection of cells containing or expressing the donor DNA. As with the sequence encoding the binder, discussed above, such elements may be associated with a promoter on the donor DNA or may be placed under control of a promoter as a result of integration of the donor DNA at a fixed locus. The latter arrangement provides a convenient means of selecting specifically for those cells which have integrated the donor DNA at the desired site, since these cells should express the genetic element for selection. This may be, for example, a gene conferring resistance to a negative selection agent such as blasticidin or puromycin. One or more selection steps may be applied to remove unwanted cells, such as cells that lack the donor DNA or which have not integrated the donor DNA at the correct position. Alternatively these cells may be permitted to remain mixed with clones of the library.
[0109] The expression of a membrane anchored binder could itself be used as a form of selectable marker. For example if a library of antibody genes, formatted as IgG or scFv-Fc fusions are introduced, then cells which express the antibody can be selected using secondary reagents which recognise the surface expressed Fc using methods described herein. Upon initial transfection with donor DNA encoding the transgene under the control of an exogenous promoter, transient expression (and cell surface expression) of the binder will occur and it will be necessary to wait for transient expression to abate (to achieve targeted integration of e.g., 1-2 antibody genes/cell).
[0110] As an alternative a construct encoding a membrane tethering element (e.g., the Fc domain of the present example fused to the PDGF receptor transmembrane domain) could be pre-integrated before the library of binders is introduced. If this membrane-tethering element lacks a promoter or is encoded within an exon which is out of frame with the preceding exon then surface expression will be compromised. Targeted integration of an incoming donor molecule can then correct this defect (e.g., by targeting a promoter or an "in-frame" exon into the intron which is upstream of the defective tethering element). If the frame "correcting exon" also encodes a binder then a fusion will be produced between the binder and the membrane tethering element resulting in surface expression of both. Thus correctly targeted integration will result in-frame expression of the membrane tethering element alone or as part of a fusion with the incoming binder. Furthermore if the incoming library of binders lack a membrane tethering element and these are incorrectly integrated they will not be selected. Thus expression of the binder itself on the cell surface can be used to select the population of cells with correctly targeted integration.
Number of Clones and Library Diversity
[0111] Yeast display libraries of 10.sup.7-1010 have previously been constructed and demonstrated to yield binders in the absence of immunisation or pre-selection of the population [9, 55, 56, 57]. Many of the previously published mammalian display libraries used antibody genes derived from immunised donors or even enriched antigen-specific B lymphocytes, given the limitations of library size and variability when using cells from higher eukaryotes. Thanks to the efficiency of gene targeting described in the present invention large, naive libraries can be constructed in higher eukaryotes such as mammalian cells, which match those described for simpler eukaryotes such as yeast.
[0112] Following integration of donor DNA into the cellular DNA, the resulting recombinant cells are cultured to allow their replication, generating a clone of cells from each initially-produced recombinant cell. Each clone is thus derived from one original cell into which donor DNA was integrated at an integration site created by the site-specific nuclease. Methods according to the present invention are associated with a high efficiency and high fidelity of donor DNA integration, and a library according to the present invention may contain at least 100, 10.sup.3, 10.sup.4, 10.sup.5, 10.sup.6, 10.sup.7, 10.sup.8, 10.sup.9 or 10.sup.10 clones.
[0113] Using nuclease-directed integration it is possible to target 10% or more of transfected mammalian cells. It is also practical to grow and transform >10.sup.10 cells (e.g. from 5 litres of cells growing at 2.times.10.sup.6 cells/ml). Transfection of such large numbers of cells could be done using standard methods including polyethyleneimine-mediated transfection as described herein. In addition methods are available for highly efficient electroporation of 10.sup.10 cells in 5 minutes e.g. http://www.maxcyte.com. Thus using the approach of the present invention it is possible to create libraries in excess of 10.sup.9 clones.
[0114] When the population of donor DNA molecules that is used to create the library contains multiple copies of the same sequence, two or more clones may be obtained that contain DNA encoding the same binder. It can also be the case that a clone may contain donor DNA encoding more than one different binder, for example if there is more than one recognition sequence for the site-specific nuclease, as detailed elsewhere herein. Thus, the diversity of the library, in terms of the number of different binders encoded or expressed, may be different from the number of clones obtained.
[0115] Clones in the library preferably contain donor DNA encoding one or two members of the repertoire of binders and/or preferably express only one or two members of the repertoire of binders. A limited number of different binders per cell is an advantage when it comes to identifying the clone and/or DNA encoding a particular binder identified when screening the library against a given target. This is simplest when clones encode a single member of the repertoire of binders. However it is also straightforward to identify the relevant encoding DNA for a desired binder if a clone selected from a library encodes a small number of different binders, for example a clone may encode two members of the repertoire of binders. As discussed elsewhere herein, clones encoding one or two binders are particularly convenient to generate by selecting a recognition sequence for the site-specific nuclease that occurs once per chromosomal copy in a diploid genome, as diploid cells contain duplicate fixed loci, one on each chromosomal copy, and the donor DNA may integrate at one or both fixed loci. Thus, clones of the library may each express only one or two members of the repertoire of binders.
[0116] Binders displayed on the surface of cells of the library may be identical to (having the same amino acid sequence as) other binders displayed on the same cell. The library may consist of clones of cells which each display a single member of the repertoire of binders, or of clones displaying a plurality of members of the repertoire of binders per cell. Alternatively a library may comprise some clones that display a single member of the repertoire of binders, and some clones that display a plurality of members (e.g., two) of the repertoire of binders.
[0117] Accordingly, a library according to the present invention may comprise clones encoding more than one member of the repertoire of binders, wherein the donor DNA is integrated at duplicate fixed loci or multiple independent fixed loci.
[0118] As noted above, it is easiest to identify the corresponding encoding DNA for a binder if the corresponding clone expresses only one binder. Typically, a molecule of donor DNA will encode a single binder. The binder may be multimeric so that a molecule of donor DNA includes multiple genes or open reading frames corresponding to the various subunits of the multimeric binder.
[0119] A library according to the present invention may encode at least 100, 10.sup.3, 10.sup.4, 10.sup.5 or 10.sup.6, 10.sup.7, 10.sup.8, 10.sup.9 or 10.sup.10 different binders. Where the binders are multimeric, diversity may be provided by one or more subunits of the binder. Multimeric binders may combine one or more variable subunits with one or more constant subunits, where the constant subunits are the same (or of more limited diversity) across all clones of the library. In generating libraries of multimeric binders, combinatorial diversity is possible where a first repertoire of binder subunits may pair with any of a second repertoire of binder subunits.
Binders
[0120] A "binder" in accordance with the present invention is a binding molecule, representing a specific binding partner for another molecule. Typical examples of specific binding partners are antibody-antigen and receptor-ligand.
[0121] The repertoire of binders encoded by a library will usually share a common structure and have one or more regions of diversity. The library therefore enables selection of a member of a desired structural class of molecules, such as a peptide or a scFv antibody molecule. For example, the binders may be polypeptides sharing a common structure and having one or more regions of amino acid sequence diversity.
[0122] This can be illustrated by considering a repertoire of antibody molecules. These may be antibody molecules of a common structural class, e.g., IgG, Fab, scFv-Fc or scFv, differing in one or more regions of their sequence. Antibody molecules typically have sequence variability in their complementarity determining regions (CDRs), which are the regions primarily involved in antigen recognition. A repertoire of binders in the present invention may be a repertoire of antibody molecules which differ in one or more CDRs, for example there may be sequence diversity in all six CDRs, or in one or more particular CDRs such as the heavy chain CDR3 and/or light chain CDR3.
[0123] Antibody molecules and other binders are described in more detail elsewhere herein. The potential of the present invention however extends beyond antibody display to include display of libraries of peptides or engineered proteins, including receptors, ligands, individual protein domains and alternative protein scaffolds [58, 59]. Nuclease-directed site-specific integration can be used to make libraries of other types of binders previously engineered using other display systems. Many of these involve monomeric binding domains such as DARPins and lipocalins, affibodies and adhirons [58, 59, 152]. Display on eukaryotes, particularly mammalian cells, also opens up the possibility of isolating and engineering binders or targets involving more complex, multimeric targets. For example T cell receptors (TCRs) are expressed on T cells and have evolved to recognise peptide presented in complex with MHC molecules on antigen presenting cells. Libraries encoding and expressing a repertoire of TCRs may be generated, and may be screened to identify binding to MHC peptide complexes as further described elsewhere herein.
[0124] For multimeric binders, donor DNA encoding the binder may be provided as one or more DNA molecules. For example, where individual antibody VH and VL domains are to be separately expressed, these may be encoded on separate molecules of donor DNA. The donor DNA integrates into the cellular DNA at multiple integration sites, e.g., the binder gene for the VH at one locus and the binder gene for the VL at a second locus. Methods of introducing donor DNA encoding separate binder subunits are described in more detail elsewhere herein. Alternatively, both subunits or parts of a multimeric binder may be encoded on the same molecule of donor DNA which integrates at a fixed locus.
[0125] A binder may be an antibody molecule or a non-antibody protein that comprises an antigen-binding site. An antigen binding site may be provided by means of arrangement of peptide loops on non-antibody protein scaffolds such as fibronectin or cytochrome B etc., or by randomising or mutating amino acid residues of a loop within a protein scaffold to confer binding to a desired target [60, 61, 62]. Protein scaffolds for antibody mimics are disclosed in WO/0034784 in which the inventors describe proteins (antibody mimics) that include a fibronectin type III domain having at least one randomised loop. A suitable scaffold into which to graft one or more peptide loops, e.g., a set of antibody VH CDR loops, may be provided by any domain member of the immunoglobulin gene superfamily. The scaffold may be a human or non-human protein.
[0126] Use of antigen binding sites in non-antibody protein scaffolds has been reviewed previously [63]. Typical are proteins having a stable backbone and one or more variable loops, in which the amino acid sequence of the loop or loops is specifically or randomly mutated to create an antigen-binding site having for binding the target antigen. Such proteins include the IgG-binding domains of protein A from S. aureus, transferrin, tetranectin, fibronectin (e.g. 10th fibronectin type III domain) and lipocalins. Other approaches include small constrained peptide e.g., based on"knottin" and cyclotides scaffolds [64]. Given their small size and complexity particularly in relation to correct formation of disulphide bond, there may be advantages to the use of eukaryotic cells for the selection of novel binders based on these scaffolds. Given the common functions of these peptides in nature, libraries of binders based on these scaffolds may be advantageous in generating small high affinity binders with particular application in blocking ion channels and proteases.
[0127] In addition to antibody sequences and/or an antigen-binding site, a binder may comprise other amino acids, e.g., forming a peptide or polypeptide, such as a folded domain, or to impart to the molecule another functional characteristic in addition to ability to bind antigen. A binder may carry a detectable label, or may be conjugated to a toxin or a targeting moiety or enzyme (e.g., via a peptidyl bond or linker). For example, a binder may comprise a catalytic site (e.g., in an enzyme domain) as well as an antigen binding site, wherein the antigen binding site binds to the antigen and thus targets the catalytic site to the antigen. The catalytic site may inhibit biological function of the antigen, e.g., by cleavage.
Antibody Molecules
[0128] Antibody molecules are preferred binders. Antibody molecules may be whole antibodies or immunoglobulins (Ig), which have four polypeptide chains--two identical heavy chains and two identical light chains. The heavy and light chains form pairs, each having a VH-VL domain pair that contains an antigen binding site. The heavy and light chains also comprise constant regions: light chain CL, and heavy chain CH1, CH2, CH3 and sometimes CH4 (the fifth domain CH4 is present in human IgM and IgE). The two heavy chains are joined by disulphide bridges at a flexible hinge region. An antibody molecule may comprise a VH and/or a VL domain.
[0129] The most common native format of an antibody molecule is an IgG which is a heterotetramer consisting of two identical heavy chains and two identical light chains. The heavy and light chains are made up of modular domains with a conserved secondary structure consisting of a four-stranded antiparallel beta-sheet and a three-stranded anti-parallel beta-sheet, stabilised by a single disulphide bond. Antibody heavy chains each have an N terminal variable domain (VH) and 3 relatively conserved "constant" immunoglobulin domains (CH1, CH2, CH3) while the light chains have one N terminal variable domain (VL) and one constant domains (CL). Disulphide bonds stabilise individual domains and form covalent linkages to join the four chains in a stable complex. The VL and CL of the light chain associates with VH and CH1 of the heavy chain and these elements can be expressed alone to form a Fab fragment. The CH2 and CH3 domains (also called the "Fc domain") associate with another CH2:CH3 pair to give a tetrameric Y shaped molecule with the variable domains from the heavy and light chains at the tips of the "Y". The CH2 and CH3 domains are responsible for the interactions with effector cells and complement components within the immune system. Recombinant antibodies have previously been expressed in IgG format or as Fabs (consisting of a dimer of VH:CH1 and a light chain). In addition the artificial construct called a single chain Fv (scFv) could be used consisting of DNA encoding VH and VL fragments fused genetically with DNA encoding a flexible linker.
[0130] Binders may be human antibody molecules. Thus, where constant domains are present these are preferably human constant domains.
[0131] Binders may be antibody fragments or smaller antibody molecule formats, such as single chain antibody molecules. For example, the antibody molecules may be scFv molecules, consisting of a VH domain and a VL domain joined by a linker peptide. In the scFv molecule, the VH and VL domains form a VH-VL pair in which the complementarity determining regions of the VH and VL come together to form an antigen binding site.
[0132] Other antibody fragments that comprise an antibody antigen-binding site include, but are not limited to, (i) the Fab fragment consisting of VL, VH, CL and CH1 domains; (ii) the Fd fragment consisting of the VH and CH1 domains; (iii) the Fv fragment consisting of the VL and VH domains of a single antibody; (iv) the dAb fragment [65, 66, 67], which consists of a VH or a VL domain; (v) isolated CDR regions; (vi) F(ab')2 fragments, a bivalent fragment comprising two linked Fab fragments (vii) scFv, wherein a VH domain and a VL domain are linked by a peptide linker which allows the two domains to associate to form an antigen binding site [68, 69]; (viii) bispecific single chain Fv dimers (PCT/US92/09965) and (ix) "diabodies", multivalent or multispecific fragments constructed by gene fusion (WO94/13804; [70]). Fv, scFv or diabody molecules may be stabilised by the incorporation of disulphide bridges linking the VH and VL domains [71].
[0133] Various other antibody molecules including one or more antibody antigen-binding sites have been engineered, including for example Fab2, Fab3, diabodies, triabodies, tetrabodies and minibodies (small immune proteins). Antibody molecules and methods for their construction and use have been described [72].
[0134] Other examples of binding fragments are Fab', which differs from Fab fragments by the addition of a few residues at the carboxyl terminus of the heavy chain CH1 domain, including one or more cysteines from the antibody hinge region, and Fab'-SH, which is a Fab' fragment in which the cysteine residue(s) of the constant domains bear a free thiol group.
[0135] A dAb (domain antibody) is a small monomeric antigen-binding fragment of an antibody, namely the variable region of an antibody heavy or light chain. VH dAbs occur naturally in camelids (e.g., camel, llama) and may be produced by immunizing a camelid with a target antigen, isolating antigen-specific B cells and directly cloning dAb genes from individual B cells. dAbs are also producible in cell culture. Their small size, good solubility and temperature stability makes them particularly physiologically useful and suitable for selection and affinity maturation. Camelid VH dAbs are being developed for therapeutic use under the name "Nanobodies.TM.".
[0136] Synthetic antibody molecules may be created by expression from genes generated by means of oligonucleotides synthesized and assembled within suitable expression vectors, for example as described by Knappik et al. [73] or Krebs et al. [74].
[0137] Bispecific or bifunctional antibodies form a second generation of monoclonal antibodies in which two different variable regions are combined in the same molecule [75]. Their use has been demonstrated both in the diagnostic field and in the therapy field from their capacity to recruit new effector functions or to target several molecules on the surface of tumour cells. Where bispecific antibodies are to be used, these may be conventional bispecific antibodies, which can be manufactured in a variety of ways [76], e.g., prepared chemically or from hybrid hybridomas, or may be any of the bispecific antibody fragments mentioned above. These antibodies can be obtained by chemical methods [77, 78] or somatic methods [79, 80] but likewise and preferentially by genetic engineering techniques which allow the heterodimerisation to be forced and thus facilitate the process of purification of the antibody sought [81]. Examples of bispecific antibodies include those of the BiTE.TM. technology in which the binding domains of two antibodies with different specificity can be used and directly linked via short flexible peptides. This combines two antibodies on a short single polypeptide chain. Diabodies and scFv can be constructed without an Fc region, using only variable domains, potentially reducing the effects of anti-idiotypic reaction.
[0138] Bispecific antibodies can be constructed as entire IgG, as bispecific Fab'2, as Fab'PEG, as diabodies or else as bispecific scFv. Further, two bispecific antibodies can be linked using routine methods known in the art to form tetravalent antibodies.
[0139] Bispecific diabodies, as opposed to bispecific whole antibodies, may also be particularly useful. Diabodies (and many other polypeptides, such as antibody fragments) of appropriate binding specificities can be readily selected. If one arm of the diabody is to be kept constant, for instance, with a specificity directed against an antigen of interest, then a library can be made where the other arm is varied and an antibody of appropriate specificity selected. Bispecific whole antibodies may be made by alternative engineering methods as described in Ridgeway et al., 1996 [82].
[0140] A library according to the invention may be used to select an antibody molecule that binds one or more antigens of interest. Selection from libraries is described in detail below. Following selection, the antibody molecule may then be engineered into a different format and/or to contain additional features. For example, the selected antibody molecule may be converted to a different format, such as one of the antibody formats described above. The selected antibody molecules, and antibody molecules comprising the VH and/or VL CDRs of the selected antibody molecules, are an aspect of the present invention. Antibody molecules and their encoding nucleic acid may be provided in isolated form.
[0141] Antibody fragments can be obtained starting from an antibody molecule by methods such as digestion by enzymes e.g. pepsin or papain and/or by cleavage of the disulphide bridges by chemical reduction. In another manner, the antibody fragments can be obtained by techniques of genetic recombination well known to the person skilled in the art or else by peptide synthesis by means of, for example, automatic peptide synthesisers, or by nucleic acid synthesis and expression.
[0142] It is possible to take monoclonal and other antibodies and use techniques of recombinant DNA technology to produce other antibodies or chimaeric molecules that bind the target antigen. Such techniques may involve introducing DNA encoding the immunoglobulin variable region, or the CDRs, of an antibody to the constant regions, or constant regions plus framework regions, of a different immunoglobulin. See, for instance, EP-A-184187, GB 2188638A or EP-A-239400, and a large body of subsequent literature.
[0143] Antibody molecules may be selected from a library and then modified, for example the in vivo half-life of an antibody molecule can be increased by chemical modification, for example PEGylation, or by incorporation in a liposome.
Sources of Binder Genes
[0144] The traditional route for generation of monoclonal antibodies utilises the immune system of laboratory animals like mice and rabbits to generate a pool of high affinity antibodies which are then isolated by the use of hybridoma technology. The present invention provides an alternative route to identifying antibodies arising from immunisation. VH and VL genes could be amplified from the B cells of immunised animals and cloned into an appropriate vector for introduction into eukaryotic libraries followed by selection from these libraries. Phage display and ribosome display allows very large libraries (>10.sup.9 clones) to be constructed enabling isolation of human antibodies without immunisation. The present invention could also be used in conjunction with such methods. Following rounds of phage display selection, the selected population of binders could be introduced into eukaryotic cells by nuclease-directed integration as described herein. This would allow the initial use of very large libraries based in other systems (e.g., phage display) to enrich a population of binders while allowing their efficient screening using eukaryotic cells as described above. Thus the invention can combine the best features of both phage display and eukaryotic display to give a high throughput system with quantitative screening and sorting.
[0145] Using phage display and yeast display it has previously been demonstrated that it is also possible to generate binders without resorting to immunisation, provided display libraries of sufficient size are used. For example multiple binders were generated from a non-immune antibody library of >10.sup.7 clones [83]. This in turn allows generation of binders to targets which are difficult by traditional immunisation routes e.g., generation of antibodies to "self-antigens" or epitopes which are conserved between species. For example, human/mouse cross-reactive binders can be enriched by sequential selection on human and then mouse versions of the same target. Since it is not possible to specifically immunise humans to most targets of interest, this facility is particularly important in allowing the generation of human antibodies which are preferred for therapeutic approaches.
[0146] In examples of mammalian display to date, where library sizes and quality were limited, binders have only been generated using repertoires which were pre-enriched for binders, e.g., from immunisation or from engineering of pre-existing binders. The ability to make large libraries in eukaryotic cells and particularly higher eukaryotes creates the possibility of isolating binders direct from these libraries starting with non-immune binders or binders which have not previously been selected within another system. With the present invention it is possible to generate binders from non-immune sources. This in turn opens up the possibilities for using binder genes from multiple sources. Binder genes could come from PCR of natural sources such as antibody genes. Binder genes could also be re-cloned from existing libraries, such as antibody phage display libraries, and cloned into a suitable donor vector for nuclease-directed integration into target cells. Binders may be completely or partially synthetic in origin. Furthermore various types of binders are described elsewhere herein, for example binder genes could encode antibodies or could encode alternative scaffolds [58, 59], peptides or engineered proteins or protein domains.
Binder Display
[0147] To provide a repertoire of binders for screening against a target of interest, the library may be cultured to express the binders in either soluble secreted form or in transmembrane form. For cell surface display it is necessary to retain the expressed binder on the surface of the cell which encodes it. Binders may comprise or be linked to a membrane anchor, such as a transmembrane domain, for extracellular display of the binder at the cell surface. This may involve direct fusion of the binder to a membrane localisation signal such as a GPI recognition sequence or to a transmembrane domain such as the transmembrane domain of the PDGF receptor [84]. Retention of binders at the cell surface can also be done indirectly by association with another cell surface retained molecule expressed within the same cell. This associated molecule could itself be part of a heterodimeric binder, such as tethered antibody heavy chain in association with a light chain partner that is not directly tethered.
[0148] Although cell surface immobilisation facilitates selection of the binder, in many applications it is necessary to prepare cell-free, secreted binder. It will be possible to combine membrane tethering and soluble secretion using a recapture method of attaching the secreted binders to cell surface receptors. One approach is to format the library of binders as secreted molecules which can associate with a membrane anchored molecule expressed within the same cell which can function to capture a secreted binder. For example, in the case of antibodies or binder molecules fused to antibody Fc domains, a membrane tethered Fc can "sample" secreted binder molecules being expressed in the same cell resulting in display of a monomeric fraction of the binder molecules being expressed while the remainder is secreted in a bivalent form (U.S. Pat. No. 8,551,715). An alternative is to use a tethered IgG binding domain such as protein A.
[0149] Other methods for retaining secreted antibodies with the cells producing them are reviewed in Kumar et al. (2012) [85] and include encapsulation of cells within microdrops, matrix aided capture, affinity capture surface display (ACSD), secretion and capture technology (SECANT) and "cold capture" [85]. In examples given for ACSD and SECANT [85], biotinylation is used to facilitate immobilisation of streptavidin or a capture antibody on the cell surface. The captured molecule in turn captures secreted antibodies. In the example of SECANT in vivo biotinylated of the secreted molecule occurs. Using the "cold capture" technique secreted antibody can be detected on producer cells using antibodies directed to the secreted molecule. It has been proposed that this due to association of the secreted antibody with the glycocalyx of the cell [86]. Alternatively it has been suggested that the secreted product is trapped by staining antibodies on the cell surface before being endocytosed [87]. The above methods have been used to identify high expressing clones within a population but could potentially be adapted for identification of binding specificity, provided the association has sufficient longevity at the cell surface.
[0150] Even when the binder is directly tethered to the cell surface it is possible to generate a soluble product. For example the gene encoding the selected binder can be recovered and cloned into an expression vector lacking the membrane anchored sequence. Alternatively, and as demonstrated in the Examples, an expression construction can be used in which the transmembrane domain is encoded within an exon flanked by recombination sites, e.g., ROX recognition sites for Dre recombinase [88]. In this example the exon encoding the transmembrane domain can be removed by transfection with a gene encoding Dre recombinase to switch expression to a secreted form. As demonstrated herein, secreted antibody was produced by this method without the need for recombinase action. This is presumably as a result of alternative splicing [89].
[0151] Any of the above methods or other suitable approaches can be used to ensure that binders expressed by clones of a library are displayed on the surface of their expressing cells.
Screening to Identify Binders to a Target of Interest
[0152] As noted, the eukaryotic cell library may be used in a method of screening for a binder that recognises a target. Such a method may comprise:
[0153] providing a library as described herein,
[0154] culturing cells of the library to express the binders,
[0155] exposing the binders to the target, allowing recognition of the target by one or more cognate binders, if present, and
[0156] detecting whether the target is recognised by a cognate binder.
[0157] Selections could be carried using a range of target molecule classes, e.g., protein, nucleic acid, carbohydrate, lipid, small molecules. The target may be provided in soluble form. The target may be labelled to facilitate detection, e.g., it may carry a fluorescent label or it may be biotinylated. Cells expressing a target-specific binder may be isolated using a directly or indirectly labelled target molecule, where the binder captures the labelled molecule. For example, cells that are bound, via the binder:target interaction, to a fluorescently labelled target can be detected and sorted by flow cytometry or FACS to isolate the desired cells. Selections involving cytometry require target molecules which are directly fluorescently labelled or are labelled with molecules which can be detected with secondary reagents, e.g., biotinylated target can be added to cells and binding to the cell surface can be detected with fluorescently labelled streptavidin such as streptavidin-phycoerythrin. A further possibility is to immobilise the target molecule or secondary reagents which bind to the target on a solid surface, such as magnetic beads or agarose beads, to allow enrichment of cells which bind the target. For example cells that bind, via the binder: target interaction, to a biotinylated target can be isolated on a substrate coated with streptavidin, e.g., streptavidin-coated beads.
[0158] In screening libraries it is preferable to over-sample, i.e., screen more clones than the number of independent clones present within the library to ensure effective representation of the library. Identifying binders from very large libraries provided by the present invention could be done by flow sorting but this would take several days, particularly if over-sampling the library. As an alternative initial selections could be based on the use of recoverable antigen, e.g., biotinylated antigen recovered on streptavidin-coated magnetic beads. Thus streptavidin-coated magnetic beads could be used to capture cells which have bound to biotinylated antigen. Selection with magnetic beads could be used as the only selection method or this could be done in conjunction with flow cytometry where better resolution can be achieved, e.g., differentiating between a clone with higher expression levels and one with a higher affinity [56, 57].
[0159] The in vitro nature of display technology approaches makes it is possible to control selection in a way that is not possible by immunisation, e.g., selecting on a particular conformational state of a target [90, 91]. Targets could be tagged through chemical modification (fluorescein, biotin) or by genetic fusion (e.g. protein fused to an epitope tag such as a FLAG tag or another protein domain or a whole protein). The tag could be nucleic acid (e.g., DNA, RNA or non-biological nucleic acids) where the tag is part fused to target nucleic acid or could be chemically attached to another type of molecule such as a protein. This could be through chemical conjugation or through enzymatic attachment [92]. Nucleic acid could be also fused to a target through a translational process such as ribosome display. The "tag" may be another modification occurring within the cell (e.g., glycosylation, phosphorylation, ubiqitinylation, alkylation, PASylation, SUMO-lation and others described at the Post-translational Database (db-PTM) at http://dbptm.mbc.nctu.edu.tw/statistics.php) which can be detected via secondary reagents. This would yield binders which bind an unknown target protein on the basis of a particular modification.
[0160] Targets could be detected using existing binders which bind to that target molecule, e.g., target specific antibodies. Use of existing binders for detection will have the added advantage of identifying binders within the library of binders which recognise an epitope distinct from the binder used for detection. In this way pairs of binders could be identified for use in applications such as sandwich ELISA. Where possible a purified target molecule would be preferred. Alternatively the target may be displayed on the surface of a population of target cells and the binders are displayed on the surface of the library cells, the method comprising exposing the binders to the target by bringing the library cells into contact with the target cells. Recovery of the cells expressing the target (e.g., using biotinylated cells expressing target) will allow enrichment of cells which express binders to them. This approach would be useful where low affinity interactions are involved since there is the potential for a strong avidity effect.
[0161] The target molecule could also be unpurified recombinant or unpurified native targets provided a detection molecule is available to identify cell binding (as described above). In addition binding of target molecules to the cell expressing the binder could be detected indirectly through the association of target molecule to another molecule which is being detected, e.g., a cell lysate containing a tagged molecule could be incubated with a library of binders to identify binders not only to the tagged molecule but also binders to its associated partner proteins. This would result in a panel of antibodies to these partners which could be used to detect or identify the partner (e.g., using mass spectrometry). Cellular fractionation could be used to enrich targets from particular sub-cellular locations. Alternatively differential biotinylation of surface or cytoplasmic fractions could be used in conjunction with streptavidin detection reagents for eukaryotic display [93,94]. The use of detergent solubilised target preparations is a particularly useful approach for intact membrane proteins such as GPCRs and ion channels which are otherwise difficult to prepare. The presence of detergents may have a detrimental effect on the eukaryotic cells displaying the binders requiring recovery of binder genes without additional growth of the selected cells.
[0162] Following detection of target recognition by a cognate binder, cells of a clone containing DNA encoding the cognate binder may be recovered. DNA encoding the binder may then be isolated (e.g., identified or amplified) from the recovered clone, thereby obtaining DNA encoding a binder that recognises the target.
[0163] Exemplary binders and targets are detailed elsewhere herein. A classic example is a library of antibody molecules, which may be screened for binding to a target antigen of interest. Other examples include screening a library of TCRs against a target MHC:peptide complex or screening a library of MHC:peptide complexes against a target TCR.
TCR:MHC and Other Receptor Interactions
[0164] T cell receptors (TCRs) are expressed on T cells and have evolved to recognise peptide presented in complex with MHC molecules on antigen presenting cells. TCRs are heterodimers consisting in 95% of cases of alpha and beta heterodimers and in 5% of cases of gamma and delta heterodimers. Both monomer units have an N terminal immunoglobulin domain which has 3 variable complementarity determining regions (CDRs) involved in driving interaction with target. The functional TCR is present within a complex of other sub-units and signalling is enhanced by co-stimulation with CD4 and CD8 molecules (specific for class I and class II MHC molecules respectively). On antigen presenting cells, proteins are processed, and presented on the cell surface in complex with MHC molecules which are themselves part of a multimeric protein complex. TCRs recognizing peptides originating from "self" are removed during development and the system is poised for recognition of foreign peptides presented on antigen presenting cells to effect an immune response. The outcome of recognition of a peptide:MHC complex depends on the identity of the T cell and the affinity of that interaction.
[0165] It would be valuable to identify the genes encoding TCRs or MHC:peptide complexes which drive interactions involved in pathological conditions, e.g., as occurs in autoimmune disease. In the case of autoimmune disease, identification of interacting partners, e.g., a TCR driving a pathogenic condition could pave the way to either specifically blocking such interactions or removing offending cytotoxic cells. It would be desirable to engineer TCRs for altered binding e.g. higher affinity to targets of interest, e.g., in re-targeting T cells in cancer or enhancing the effect of existing T cells [95]. Alternatively the behaviour of regulatory or suppressive T cells might be altered as a therapeutic modality, e.g., for directing or enhancing immunotherapy of cancer by introducing specific TCRs into T cells or by using expressed TCR protein as therapeutic entities [96].
[0166] Display of libraries of TCRs on surface of yeast cells and mammalian cells has previously been demonstrated. In the case of yeast cells it was necessary to engineer the TCR and present it in a single chain format. Since the affinity of interaction between TCR and peptide:MHC complex is low, the soluble component (e.g., peptide:MHC in this case) is usually presented in a multimeric format. TCR specificity has been engineered for peptides in complex with MHC class I [97] and MHC class II [98]. TCRs have also been expressed on the surface of a mutant mouse T cells (lacking TCR alpha and beta chains) and variant TCRs with improved binding properties have been isolated [99]. For example Chervin et al. introduced TCRs by retroviral infection and an effective library size of 10.sup.4 clones was generated [100]. Using nuclease-directed integration of binders as proposed here, a similar approach could be taken to engineering T cells. As well as selecting TCRs with altered recognition properties, display libraries could be used to screen libraries of peptide or of MHC variants for recognition by TCRs. For example peptide:MHC complexes have been displayed on insect cells and used to epitope map TCRs presented in a multimeric format [101].
[0167] As noted, screening methods may involve displaying the repertoire of binders on the cell surface and probing with a target presented as a soluble molecule, which may be a multimeric target. An alternative, which can be especially useful with multimeric targets, is to screen directly for cell:cell interactions, where binder and target are presented on the surface of different cells. For example if activation of a TCR of interest led to expression of a reporter gene this could be used to identify activating peptides or activating MHC molecules presented within a peptide:MHC library. In this particular example the reporter cell does not encode the library member but could be used to identify the cell which does encode it. The approach could potentially extend to a "library versus library" approach. For example extending the example described above, a TCR library could be screened against a peptide:MHC library. More broadly the example of screening a library of binders presented on one cell surface using a binding partner on another cell could be extended to other types of cell:cell interactions e.g., identification of binders which inhibit or activate signalling within the Notch or Wnt pathways. Thus the present invention could be used in alternative cell based screening system including recognition systems based on cell:cell interactions.
[0168] As an example, chimeric antigen receptors ("CARS") represent a fusion between an antibody binding domain (usually formatted as a scFv) and a signalling domain. These have been introduced into T cells with the aim of re-directing the T cell in vivo to attack tumour cells through antibody recognition and binding to tumour-specific antigens. A number of different factors could affect the success of this strategy including the combination of antibody specificity, format, antibody affinity, linker length, fused signalling module, expression level in T cells, T cell sub-type and interaction of the CAR with other signalling molecules [102, 103]. The ability to create large libraries of CARs in primary T cells incorporating individual or combinations of the above variables would allow a functional search for effective and optimal CAR construction. This functional "search" could be carried out in vitro or in vivo. For example Alonso-Camino (2009) have fused a scFv recognizing CEA to the .xi. chain of the TCR:CD3 complex and introduced this genetic construct into a human Jurkat cell line [104]. Upon interaction with CEA present on either HeLa cells or tumour cells they showed upregulation of the early T cell activation marker CD69. This approach could be used to identify CAR fusion constructs with appropriate activation or inhibitory properties using cultured or primary cells.
[0169] Going one step further functionality of CAR constructs could be assessed in vivo. For example a library of CARs constructed in primary mouse T cells could be introduced into tumour bearing mice to identify T cell clones stimulated to proliferate through encounter with tumour. If necessary this T cell library could be pre-selected based on antigen binding specificity using the methods described above. In either case the incoming library of binders could be used to replace an existing binder molecule (e.g., MHC or TCR or antibody variable domain).
Phenotype Screens
[0170] Described here are various methods for selection of binders which modify cell signalling and cellular behaviour.
[0171] A library may be screened for altered cellular phenotype as a result of the action of the binder on its target.
[0172] Antibodies which modify cell signalling by binding to ligands or receptors have a proven track record in drug development and the demand for such therapeutic antibodies continues to grow. Such antibodies and other classes of functional binders also have potential in controlling cell behaviour in vivo and in vitro. The ability to control and direct cellular behaviour however relies on the availability of natural ligands which control specific signalling pathways. Unfortunately many natural ligands such as those controlling stem cell differentiation (e.g., members of FGF, TGF-beta, Wnt and Notch super-families) often exhibit promiscuous interactions and have limited availability due to their poor expression/stability profiles. With their exquisite specificity, antibodies have great potential in controlling cellular behaviour.
[0173] The identification of functional antibodies that modify cell signalling has historically been relatively laborious involving picking clones, expressing antibody, characterising according to sequence and binding properties, conversion to mammalian expression systems and addition to functional cell based assays. The eukaryotic display approach described herein will reduce this effort but there is still a requirement for production of antibody and addition to a separate reporter cell culture. Therefore, a preferred alternative may be to directly screen libraries of binders expressed in eukaryotic cells for the effect of binding on cell signalling or cell behaviour by using the production cell itself as a reporter cell. Following introduction of antibody genes, clones within the resulting population of cells showing alteration in reporter gene expression or altered phenotypes can be identified.
[0174] A number of recent publications have described the construction of antibody libraries by cloning repertoires of antibody genes into reporter cells [47, 105, 106]. These systems combine expression and reporting within one cell, and typically introduce a population of antibodies selected against a pre-defined target (e.g., using phage display).
[0175] A population of antibody genes may be introduced into reporter cells to produce a library by methods described herein, and clones within the population with an antibody-directed alteration in phenotype (e.g., altered gene expression or survival) can be identified. For this phenotypic-directed selection to work there is a requirement to retain a linkage between the antibody gene present within the expressing cell (genotype) and the consequence of antibody expression (phenotype). This has been achieved previously either through tethering the antibody to the cell surface [47] as described for antibody display or through the use of semi-solid medium to retain secreted antibodies in the vicinity of producing cells [105]. Alternatively antibodies and other binders can be retained inside the cell [107]. Binders retained on the cell surface or in the surrounding medium can interact with an endogenous or exogenous receptor on the cell surface causing activation of the receptor. This in turn can cause a change in expression of a reporter gene or a change in the phenotype of the cell. As an alternative the antibody can block the receptor or ligand to reduce receptor activation. The gene encoding the binder which causes the modified cellular behaviour can then be recovered for production or further engineering.
[0176] An alternative to this "target-directed" approach, it is possible to introduce a "naive" antibody population which has not been pre-selected to a particular target [108]. The cellular reporting system is used to identify members of the population with altered behaviour. Since there is no prior knowledge of the target, this non-targeted approach has a particular requirement for a large antibody repertoire, since pre-enrichment of the antibody population to the target is not possible. This approach will benefit from using nuclease-directed transgene integration as described in the present invention.
[0177] The "functional selection" approach could be used on other applications involving libraries in eukaryotic cells, particularly higher eukaryotes such as mammalian cells. The antibody could be fused to a signalling domain such that binding to target causes activation of the receptor. Kawahara et al. have constructed chimeric receptors where an extra-cellular scFv targeting fluorescein was fused to a spacer domain (the D2 domain of the Epo receptor) and various intracellular cytokine receptor domain including the thrombopoeitin (Tpo) receptor, erythropoietin (Epo) receptor, gp130, IL-2 receptor and the EGF receptor [109, 110, 111]. These were introduced into an IL-3 dependent proB cell line (BaF3) [27], where chimaeric receptors were shown to exhibit antigen-dependent activation of the chimaeric receptor leading to IL-3 independent growth. This same approach was used in model experiments to demonstrate antigen mediated chemoattraction of BaF3 cells [110]. The approach was extended beyond stable culture cells to primary cells exemplified by the survival and growth of Tpo-responsive haematopoeitic stem cells [112] or IL2 dependant primary T cells where normal stimulation by Tpo and IL-2 respectively was replaced by fluorescein directed stimulation of scFv chimaeric receptors. Thus a system based on chimaeric antibody-receptor chimaeras can be used to drive target dependent gene expression or phenotypic changes in primary or stable reporter cells. This capacity could be used to identify fused binders which drive a signalling response or binders which inhibit the response.
[0178] In a modification of the above approach separate VH and VL domains from an anti-lysozyme antibody were fused to the Epo intracellular domain [113]. Cells grew in response to addition of lysozyme indicating an antigen induced dimerisation or stabilisation of the separate VH and VL fusion partners. Thus three interacting components come together for an optimal response in this system.
[0179] Although described here with reference to antibody molecules, the above methods may also be adapted and performed with libraries of other binders.
[0180] Protein fragment complementation represents an alternative system for studying and for selecting protein:protein interactions in mammalian cells [114, 115]. This involves restoring function of split reporter proteins through protein:protein interactions. Reporter proteins which have been used include ubiquitin, DNAE intein, beta-galactosidase, dihydrofolate reductase, GFP, firefly luciferase, beta-lactamase, TEV protease. For example a recent example of this approach is the mammalian membrane 2 hybrid (MaMTH) approach where association of a bait protein:split ubiquitin:transcription factor fusion with a partner protein:split ubiquitin restores ubiquitin recognition and liberates the transcription factor to effect reporter gene expression [116]. Again binders which interfere with or enhance this interaction could be identified through perturbed signalling.
Recovery and Reformatting of Binders and Encoding DNA
[0181] Following selection of a binder or clone of interest from the library, a common next step will be to isolate (e.g., identify or amplify) the DNA encoding the binder. Optionally, it may be desired to modify the nucleic acid encoding the binder, for example to restructure the binder and/or to insert the encoding sequence into a different vector.
[0182] Where the binder is an antibody molecule, a method may comprise isolating DNA encoding the antibody molecule from cells of a clone, amplifying DNA encoding at least one antibody variable region, preferably both the VH and VL domain, and inserting DNA into a vector to provide a vector encoding the antibody molecule. A multimeric antibody molecule bearing a constant domain may be converted to a single chain antibody molecule for expression in a soluble secreted form.
[0183] Antibodies may be presented in different formats but whatever format an antibody is selected in, once the antibody gene is isolated it is possible to reconfigure it in a number of different formats. Once VH or VL domains are isolated, they can be re-cloned into expression vectors encompassing the required partner domains (see Example 1 showing a dual promoter IgG expression cassette).
[0184] A reformatting step may comprise reformatting of binders composed of a pair of subunits (e.g., scFv molecules), to a different molecular binder format (e.g., Ig or Fab) in which the original pairing of the subunits is maintained. Such methods are described in more detail elsewhere herein and can be used for monoclonal, oligoclonal or polyclonal clone reformatting. The method can be used to convert "en masse" an entire output population from any of the commonly used display technologies including phage, yeast or ribosome display.
Display of scFvs on the Surface of Mammalian Cells Fused to Fc Domains.
[0185] Although many antibody phage display libraries are formatted to display scFvs, eukaryotic display systems will allow presentation in Fab or IgG format. To take full advantage of the potential for IgG/Fab expression, particularly when using scFvs from other display systems will be necessary to take selected linked VH and VL domains within a bacterial expression system and express them within a eukaryotic system fused to appropriate constant domains. Described here is a method to convert scFv populations to immunoglobulin (Ig) or fragment, antigen binding (Fab) format in such a way that original VH and VL chain pairings are maintained. In the present invention, conversion is possible using individual clones, oligoclonal mixes or whole populations formatted as scFv while retaining the original pairing of VH and VLs chains. The method proceeds via the generation of an intermediate non-replicative "mini-circle" DNA which brings in a new "stuffer" DNA fragment. The circular DNA is linearised (e.g., by restriction digestion or PCR) which alters the relative position of the original VH and VL fragments and places the "stuffer" DNA between them. Following linearization the product can be cloned into a vector of choice, e.g., a mammalian expression vector. In this way all of the elements apart from the VH and VL can be replaced. Elements for bacterial expression can be replaced with elements for mammalian expression and fusion to alternative partners. The complete conversion process only requires a single transformation step of E. coli bacteria to generate a population of bacterial colonies each harbouring a plasmid encoding a unique Ig or Fab formatted recombinant antibody. Extending beyond conversion of scFv to IgG/Fab, the method can be employed to reformat any two joined DNA elements to clone into a vector such that after re-formatting each DNA element is surrounded by different DNA control features whilst maintaining the original pairing. A previous method has been described wherein 2 sequential cloning steps are used [117] to replace these elements in contrast to the present method which proceeds via an intermediate non-replicative circular intermediate.
[0186] A method of restructuring a binder, or population of binders, may comprise converting scFv to Ig or a fragment thereof, e.g., Fab. The method may comprise converting nucleic acid encoding scFv to DNA encoding an immunoglobulin (Ig) or fragment thereof such as Fab format, in such a way that the original variable VH and VL chain pairings are maintained. Preferably the conversion proceeds via circular DNA intermediate which may be a non-replicative "mini-circle" DNA. The method requires a single transformation of E. coli for the direct generation of bacterial transformants harbouring plasmids encoding Ig or Fab DNA.
[0187] The method may be used for monoclonal, oligoclonal or polyclonal clone reformatting. The method may be used to convert "en masse" an entire output population from any of the commonly used display technologies including phage, yeast or ribosome display.
[0188] More generally, this aspect of the invention relates to a method that allows the reformatting of any two joined DNA elements into a vector where the DNA elements are cloned under the control of separate promoters, or separated by alternative control elements, but maintaining the original DNA pairing.
[0189] Examples 7 and 14 describe such methods in further detail.
[0190] Isolation, and optional restructuring, of DNA encoding binders may be followed by introduction of that DNA into further cells to create a derivative library as described elsewhere herein, or DNA encoding one or more particular binders of interest may be introduced into a host cell for expression. The host cell may be of a different type compared with the cells of the library from which it was obtained. Generally the DNA will be provided in a vector. DNA introduced into the host cell may integrate into cellular DNA of the host cell. Host cells expressing the secreted soluble antibody molecule can then be selected.
[0191] Host cells encoding one or more binders may be provided in culture medium and cultured to express the one or more binders.
Derivative Libraries
[0192] Following production of a library by the method of the invention, one or more library clones may be selected and used to produce a further, second generation library. When a library has been generated by introducing DNA into eukaryotic cells as described herein, the library may be cultured to express the binders, and one or more clones expressing binders of interest may be recovered, for example by selecting binders against a target as described elsewhere herein. These clones may subsequently be used to generate a derivative library containing DNA encoding a second repertoire of binders.
[0193] To generate the derivative library, donor DNA of the one or more recovered clones is mutated to provide the second repertoire of binders. Mutations may be addition, substitution or deletion of one or more nucleotides. Where the binder is a polypeptide, mutation will be to change the sequence of the encoded binder by addition, substitution or deletion of one or more amino acids. Mutation may be focussed on one or more regions, such as one or more CDRs of an antibody molecule, providing a repertoire of binders of a common structural class which differ in one or more regions of diversity, as described elsewhere herein.
[0194] Generating the derivative library may comprise isolating donor DNA from the one or more recovered clones, introducing mutation into the DNA to provide a derivative population of donor DNA molecules encoding a second repertoire of binders, and introducing the derivative population of donor DNA molecules into cells to create a derivative library of cells containing DNA encoding the second repertoire of binders.
[0195] Isolation of the donor DNA may involve obtaining and/or identifying the DNA from the clone. Such methods may encompass amplifying the DNA encoding a binder from a recovered clone, e.g., by PCR and introducing mutations. DNA may be sequenced and mutated DNA synthesised.
[0196] Mutation may alternatively be introduced into the donor DNA in the one or more recovered clones by inducing mutation of the DNA within the clones. The derivative library may thus be created from one or more clones without requiring isolation of the DNA, e.g., through endogenous mutation in avian DT40 cells.
[0197] Antibody display lends itself especially well to the creation of derivative libraries. Once antibody genes are isolated, it is possible to use a variety of mutagenesis approaches (e.g., error prone PCR, oligonucleotide-directed mutagenesis, chain shuffling) to create display libraries of related clones from which improved variants can be selected. For example, with chain-shuffling the DNA encoding the population of selected VH clone, oligoclonal mix or population can be sub-cloned into a vector encoding a suitable antibody format and encoding a suitably formatted repertoire of VL chains [118]. Alternatively and again using the example of VHs, the VH clone, oligomix or population could be introduced into a population of eukaryotic cells which encode and express a population of appropriately formatted light chain partners (e.g., a VL-CL chain for association with an IgG or Fab formatted heavy chain). The VH population could arise from any of the sources discussed above including B cells of immunised animals or scFv genes from selected phage populations. In the latter example cloning of selected VHs into a repertoire of light chains could combine chain shuffling and re-formatting (e.g., into IgG format) in one step.
[0198] A particular advantage of display on eukaryotic cells is the ability to control the stringency of the selection/screening step. By reducing antigen concentration, cells expressing the highest affinity binders can be distinguished from lower affinity clones within the population. The visualisation and quantification of the affinity maturation process using flow cytometry is a maj or benefit of eukaryotic display as it gives an early indication of percentage positives in naive library and allows a direct comparison between the affinity of the selected clones and the parental population during sorting. Following sorting, the affinity of individual clones can be determined by pre-incubating with a range of antigen concentrations and analysis in flow cytometry or with a homogenous Time Resolved Fluorescence (TRF) assay or using surface plasmon resonance (SPR) (Biacore).
Characteristics and Form of the Library
[0199] The present invention enables construction of eukaryotic cell libraries having many advantageous characteristics. The invention provides libraries having any one or more of the following features:
[0200] Diversity. A library may encode and/or express at least 100, 10.sup.3, 10.sup.4, 10.sup.5 10.sup.6, 10.sup.7, 10.sup.8 or 10.sup.9 different binders.
[0201] Uniform integration. A library may consist of clones containing donor DNA integrated at a fixed locus, or at a limited number of fixed loci in the cellular DNA. Each clone in the library therefore contains donor DNA at the fixed locus or at least one of the fixed loci. Preferably clones contain donor DNA integrated at one or two fixed loci in the cellular DNA. As explained elsewhere herein, the integration site is at a recognition sequence for a site-specific nuclease. Integration of donor DNA to produce recombinant DNA is described in detail elsewhere herein and can generate different results depending on the number of integration sites. Where there is a single potential integration site in cells used to generate the library, the library will be a library of clones containing donor DNA integrated at the single fixed locus. All clones of the library therefore contain the binder genes at the same position in the cellular DNA. Alternatively where there are multiple potential integration sites, the library may be a library of clones containing donor DNA integrated at multiple and/or different fixed loci. Preferably, each clone of a library contains donor DNA integrated at a first and/or a second fixed locus. For example a library may comprise clones in which donor DNA is integrated at a first fixed locus, clones in which donor DNA is integrated at a second fixed locus, and clones in which donor DNA is integrated at both the first and second fixed loci. In preferred embodiments there are only one or two fixed loci in the clones in a library, although it is possible to integrate donor DNA at multiple loci if desired for particular applications. Therefore in some libraries each clone may contain donor DNA integrated at any one or more of several fixed loci, e.g., three, four, five or six fixed loci.
[0202] For libraries containing binder subunits integrated at separate sites, clones of the library may contain DNA encoding a first binder subunit integrated at a first fixed locus and DNA encoding a second binder subunit integrated at a second fixed locus, wherein the clones express multimeric binders comprising the first and second subunits.
[0203] Uniform transcription. Relative levels of transcription of the binders between different clones of the library is kept within controlled limits due to donor DNA being integrated at a controlled number of loci, and at the same locus in the different clones (fixed locus). Relatively uniform transcription of binder genes leads to comparable levels of expression of binders on or from clones in a library. Binders displayed on the surface of cells of the library may be identical to (having the same amino acid sequence as) other binders displayed on the same cell. The library may consist of clones of cells which each display a single member of the repertoire of binders, or of clones displaying a plurality of members of the repertoire of binders per cell. Alternatively a library may comprise some clones that display a single member of the repertoire of binders, and some clones that display a plurality of members (e.g., two) of the repertoire of binders. Preferably clones of a library express one or two members of the repertoire of binders.
[0204] For example, a library of eukaryotic cell clones according to the present invention may express a repertoire of at least 10.sup.3, 10.sup.4, 10.sup.5 10.sup.6, 10.sup.7, 10.sup.8 or 10.sup.9 different binders, e.g., IgG, Fab, scFv or scFv-Fc antibody fragments, each cell containing donor DNA integrated at a fixed locus in the cellular DNA. The donor DNA encodes the binder and may further comprise a genetic element for selection of cells into which the donor DNA is integrated at the fixed locus. Cells of the library may contain DNA encoding an exogenous site-specific nuclease.
[0205] These and other features of libraries according to the present invention are further described elsewhere herein.
[0206] The present invention extends to the library either in pure form, as a population of library clones in the absence of other eukaryotic cells, or mixed with other eukaryotic cells. Other cells may be eukaryotic cells of the same type (e.g., the same cell line) or different cells. Further advantages may be obtained by combining two or more libraries according to the present invention, or combining a library according to the invention with a second library or second population of cells, either to facilitate or broaden screening or for other uses as are described herein or which will be apparent to the skilled person.
[0207] A library according to the invention, one or more clones obtained from the library, or host cells into which DNA encoding a binder from the library has been introduced, may be provided in a cell culture medium. The cells may be cultured and then concentrated to form a cell pellet for convenient transport or storage.
[0208] Libraries will usually be provided in vitro. The library may be in a container such as a cell culture flask containing cells of the library suspended in a culture medium, or a container comprising a pellet or concentrated suspension of eukaryotic cells comprising the library. The library may constitute at least 75%, 80%, 85% or 90% of the eukaryotic cells in the container.
BRIEF DESCRIPTION OF THE DRAWINGS
[0209] Embodiments of the invention will now be described in more detail, with reference to the accompanying drawings, which are as follows:
[0210] FIG. 1. Vector for expression of IgG formatted antibodies [0211] a. pDUAL D1.3, a dual promoter expression vector for IgG secretion (in in "pCMV/myc/ER" vector backbone) [0212] b. pINT3-D1.3, a dual promoter expression vector for IgG secretion (in "pSF-CMV-f1-Pac1" vector backbone) [0213] c. pCMV/myc/ER vector backbone. ECoR1 site precedes CMV promoter. BstB1 and BstZ171 sites flank the SV40 poly A sites. [0214] d. pSF-CMV-f1-Pac1 vector backbone (Oxford Genetics) [0215] e. synthetic gene with exon encoding PDGFR transmembrane Region.TM. and exon causing secretion (sec). Solid arrows represent Rox recombination sites.
[0216] FIG. 2. Sequence of pD1 (SEQ ID NO: 1, 2, 3, 4 & 5): a dual promoter antibody expression cassette for surface expression.
Features:
[0217] pEF promoter 13-1180 BM40 leader 1193-1249
Humanised D1.3 VL 1250-1578
[0218] Human C kappa 1577-1891BGH poly A 1916-2130 CMV promoter 2146-2734 Mouse VH leader with intron 32832-3414
Humanised D1.3 VH 3419-3769
[0219] Optimised human IgG2 CH1-CH3
[0220] FIG. 3. Construction of AAVS Donor plasmid (pD2)
a. Representation of the human AAVS locus. Exon 1 and Exon 2 of the AAVS locus (encoding protein phosphatase 1, regulatory subunit 12C, PPP1R12C) are separated by an intron of 4428 bp. Splicing is in frame "0" i.e. splicing occurs between 2 intact codons from each exon. TALENs and CRISPR/Cas9 constructs are available to cleave within this intron. Hatched blocks represent the regions to the left and right of this cleavage site that are used within vector constructs to drive homologous recombination into this locus (AAVS homology arm left "Left HA" and AAVS homology arm right ("right HA"). b. Representation of the antibody encoding donor plasmid pD2. Left and right homology arms are shown at the ends of the construct representation. A synthetic Blasticidin gene is preceded by a splice acceptor which creates an "in-frame" fusion with AAVS exon 1. Also shown is the antibody expression cassette consisting of a D1.3 light chain and a D1.3 IgG2 heavy chain driven by pEF and CMV promoters respectively. c. Sequence of donor construct pD1-huD1.3 (SEQ ID NO: 6, 7 & 8). AAVS homology arms are shown underlined and emboldened. For brevity antibody cassette (previously shown in FIG. 2) is not shown in detail. Restriction sites used in clone are shown emboldened. The sequence of the plasmid backbone is shown up to the ampicillin resistance gene.
[0221] FIG. 4. Expression of IgG on cell surface
a, c. Analysis was focused on viable cells using forward scatter and staining in the FL3 channel (a, c). Cells positive for staining in the FL3 channel (representing non-viable cells which took up 7-AAD) were excluded. All cells were transfected with pD2-D1.3 in absence (a, b) or presence (c.d) of the AAVS TALENs and were stained with anti-Fc antibody.
[0222] FIG. 5. Antibody binding of antigen on cell surface. Viable cells were selected on the basis of Forward Scatter and occlusion of 7AAD (a). Cells were incubated with fluorescently labelled hen egg lysozyme (b).
[0223] FIG. 6. Effect of TALEN-directed genomic cleavage on integration of blasticidin resistance gene. Figure shows number of colonies under the conditions described.
[0224] FIG. 7. Analysis of integration of pD2-D1.3 donor plasmid into AAVS locus. Following transfection cells were selected in Blasticidin and genomic DNA was prepared. Samples 1-9 benefitted from addition of AAVS-directed TALE nucleases, samples 10-11 were from clones arising from cell transfected in the absence of TALE nuclease. Genomic DNA analysis carried out by PCR as described in text. [0225] a. Verification from 5' end of integration site. [0226] b. Verification from 3' end of integration site.
[0227] FIG. 8. Construction of the scFv-Fc expression vector pD6 [0228] a. Antibodies formatted as scFv are cloned into the Nco1/Not1 sites to create an "in-frame" fusion with the human Fc region of IgG2. [0229] b. Sequence of pD6 from the Nco1 to Pme1 sites (SEQ ID NO: 9, 10 &11).
[0230] FIG. 9. Selection of binders from cell surface scFv-Fc library (from selected phage populations). Flow cytometry analysis is shown of cells with:
a-c. integrated anti-CD229 sFv-Fc population from 2 rounds of phage display selection on CD229 d. f integrated anti-.beta.-galactosidase sFv-Fc population from 1 round of phage display selection on .beta.-galactosidase .beta.-galR1 cells) e. integrated anti-.beta.-galactosidase sFv-Fc population from 2 rounds of phage display selection on .beta.-galactosidase Sample (a) shows unstained cells and the rest were stained with human anti-Fc-phycoerythrin (in FL2) and 100 nM appropriate biotinylated antigen/streptavidin FITC (in FL1). Cells were analysed after 13 days (a, b, d, e). Examples c and f show cells stained after 20 days and the marked region shows cells collected by flow cytometry h. .beta.-galR1 cells selected by flow cytometry (FIG. 6f) were grown for 22 days and re-analysed for scFv-Fc expression and antigen binding (using 100 nM antigen). g. show the unstained equivalent. j. shows unsorted .beta.-galR1 cells from the original population (as in d) which had been grown for 42 days after transfection (j). Unlabelled cells of each population are shown for comparison (g, i)
[0231] FIG. 10. Mammalian display and sorting of IgG formatted library.
A population of antibodies were selected on .beta.-galactosidase using 1 or 2 rounds phage display, reformatted as IgG and targeted via nuclease-directed integration into the AAVS locus of HEK293 cells. Panels a, b show cells derived from the round 1 phage population either a, unsorted (after 38 days growth) or b, sorted by flow cytometry and grown for 19 days. Panels c, d show cells derived from round 2 phage population either c, unsorted (after 38 days growth or d, sorted and grown for 19 days.
[0232] FIG. 11. Construction of large naive scFv-Fc library and selection of binders Cells from the naive scFv-Fc library were stained with 500 nM biotinylated antigen and streptavidin-FITC along with phycoerythrin-labelled anti-Fc antibody as before. Region shows cells which were selected by flow sorting. Samples were labelled with biotinylated: [0233] a. CD28 [0234] b. .beta.-galactosidase [0235] c. Thyroglobulin [0236] d. EphB4
[0237] FIG. 12. Targeting vector to introduce intron containing "multiple landing" sites for comparison of integration methods. [0238] a. Intermediate GFP expression plasmid (pD3) [0239] b. AAVS1_directed targeting vector (pD4). "Landing site" incorporates elements for directing integration which are FRT, lox 2272, I-SceI meganuclease and GFP TALEN Following integration of pD4 into the AAVS locus, multiple recombination or nuclease cleavage sites are present within the genome. The incoming pD5 plasmid (FIG. 15) has left and right homology arms equivalent to the sequence present on either side of the "landing site" to drive antibody insertion by homologous recombination.
[0240] FIG. 13. Sequence of pD4 (SEQ ID NO: 12, 13, 14, 15, 16, 17 & 18.).
5 Sequence features include: AAVS Left homology 19-822 FRT site 832-879, Lox 2272 site 884-917, I-Sce1meganuclease site 933-950 GFP left TALEN binding 954-968, GFP right TALEN binding 984-997
T2A 1041-1103
GFP 1104-1949
[0241] PGK promoter 2178-2691 Puromycin delta thymidine kinase 2706-4307 loxP 4634-4667 AAVS right homology 4692-5528
[0242] FIG. 14. Verification of integration of "multiple landing site" intron in clone 6F. Following transfection cells were selected in puromycin and genomic DNA was prepared. Samples 1 represents the whole selected population. Sample 2 represents clone 6F, sample 3 is a clone transfected in the absence of TALENs and sample 4 is wild-type HEK293 cells. Primers and conditions described in text were used to verify by PCR the correct integration at the 5' and 3' ends of the genomic insertion. The major (correct sized) band is seen for the selected clone (6F) as well as the selected population.
[0243] FIG. 15. Sequence of donor plasmid for integration into Flp/GFP TALEN sites (pD5) (SEQ ID NO: 19, 20 & 21). Features include:
AAVS HA 13-233
[0244] FRT site 243-290
Lox2272 295-328
I-SceI 344-361
[0245] Blasticidin resistance 417-818
Poly A 832-1070
[0246] FIG. 16. Sequence of I-SceI meganuclease construct (SEQ ID NO: 22 & 23)
[0247] FIG. 17. Flow cytometry analysis comparing nuclease-directed integration using I-Scel meganuclease with recombinases.
Clone 6F cells were co-transfected with pD5-D1.3 and plasmids encoding the indicated nuclease/recombinase. Cells were selected with blasticidin and analysed 13 days after transfection using biotinylated anti-human Fc antibodies and streptavidin phycoerythrin. Percentage positive cells are indicated (also summarized in Table 5) a. Non-transfected, b Donor only c. I-Scel, d, eGFP TALEN, e. Cre, f. Flp recombinase (encoded by pOG44 plasmid).
[0248] FIG. 18. Nuclease-directed integration drives homologous recombination and "non-homologous end joining" (NHEJ).
a. Representation of structure of plasmid pD5 used to target the multiple "landing site" within the intron of "clone 6F cells showing position of primer J48. The "landing site" in this plasmid incorporates a FRT site, a lox2272 site and an I-SceI meganuclease site (but no GFP TALEN site). b. Representation of integration site within clone 6F (derived from pD4) showing position of primer J44. "Landing site" incorporates elements for directing integration which are FRT, lox 2272, I-SceI meganuclease and GFP TALEN. c. Representation of clone 6F integration site after homologous recombination of pD5, showing position of primers J44 and J48. d. Representation of integration site of clone 6F after NHEJ or Flp recombination of pD5, showing position of primers J44, J46 and reverse primer J44. The double headed arrow indicates the "extra" plasmid derived DNA incorporated by NHEJ or Flp-directed integration. Note in this example the incoming plasmid DNA (pD5) has homology arms (which direct homologous recombination but are not required for NHEJ). These sequences are retained after integration by NHEJ, causing a duplication of the sequence represented within the homology arms with one pair coming from the plasmid in this case and the other pair representing the endogenous genomic sequences. For simplicity the plasmid encoded homology arms are not shown, just their equivalent sequence within the genome. e. Primers 44 and 48 were used as PCR primers for samples i-iv where genomic DNA from cells transfected with the following nucleases/integrases were used: i. Scel, ii. TALEN (GFP), iii. Flp (pOG44), iv. Donor only. Molecular weight markers were "GeneRuler 1 kb ladder (New England Biolabs). Primers J44 and J48 reveal homologous recombination has occurred producing a band of 1928 bp (indicated by arrow) in nuclease cleaved samples i and ii. Primers 44 and 46 were used for samples v-viii where genomic DNA from the following samples was used. v. Scel, vi. TALEN (GFP), vii. Flp (pOG44), viii. Donor only. Primers J44 and J46 reveal that cleavage of donor and genomic DNA by I-SceI meganuclease has resulted in NHEJ (sample v.) producing a band of 1800 bp (indicated by arrow). As expected a similar sized band was achieved by Flp mediated integration (vii). NHEJ has not occurred with GFP TALEN since there was no cleavage site in the incoming plasmid.
[0249] FIG. 19. Secretion of IgG antibodies into culture supernatant.
a. Coomassie stained gel of protein A purified IgG from culture supernatants. i. IgG purified from supernatant of pD2-D 1.3 cells without transfection of Dre recombinase gene. ii. IgG purified from supernatant of pD2-D 1.3 cells transfected with Dre recombinase gene. b. Polyclonal ELISA of secreted antibodies. Sorted cells from the experiment shown in FIG. 9H (originally from antibody population cells selected by 1 round of phage display) were grown for 7 days post sorting and the culture supernatant collected. ELISA plates were coated with either .beta.-galactosidase (10 ug/ml) or BSA (10 ug/ml) overnight. Culture supernatants were diluted down to 66% after mixing with a 33% volume of 6% Marvel-PBS (this is described above as the `neat` sample). Supernatant was also diluted 1/10 in PBS and mixed with 6% MPBS in the same manner. Detection of bound scFv-Fc fusion was performed using anti-Human IgG-Eu (Perkin Elmer Cat 1244-330).
[0250] FIG. 20. Preparation of DNA fragments for the conversion of selected populations of scFv to IgG format. (a) Generation of CL-pA-CMV-Sigp DNA insert (as depicted in FIG. 21b). PCR amplification from plasmid pD2 with primers 2595 and 2597 and gel purified. Lane m, Generuler 1 kb ladder (Thermo, SM031D), lane 1 C.sub.L-pA-CMV-Sigp DNA insert. (b) Generation of scFv DNA insert was as described in Example 6. Lane m, Generuler 1 kb ladder (Thermo, SM031D), lane 1 blank, lane 2 purified scFv, lane 3 .beta.-galactosidase round 1 output scFv population, lane 4 .beta.-galactosidase round 2 output scFv population, lane 5 CD229 round 2 output scFv population. (c) Purification of NheI and XhoI digested "mini-circle" DNA. Ligations between NcoI/NotI digested DNA encoding scFv (FIG. 21, insert a) and DNA encoding constant light (CL) chain, poly A (pA), CMV promoter and signal peptide (FIG. 21, insert b) to form "mini-circle" DNA (FIG. 21c) were spin column purified, digested with NheI and XhoI and purified by 1% agarose gel. Lanes are m, Generuler 1 kb ladder (Thermo, SM031D), lane 1 .beta.-galactosidase round 1 output, lane 2 .beta.-galactosidase round 2 output, lane 3 CD229 round 2 output. Linearised product at 2.6 kb, indicated by arrow, was excised and purified.
[0251] FIG. 21. Schematic representation of the conversion process from scFv to IgG format. A DNA insert (a) encoding the antibody VH and VL domains is ligated with DNA fragment (b) encoding a constant light (CL) chain, a polyadenylation sequence (pA) a cytomegalovirus (CMV) promoter and a signal peptide (SigP). The joining of DNA molecules a and b to create a non-replicative DNA "mini-circle" c is facilitated by a "sticky-end" ligation. After ligation, the "mini-circle" c is linearized with restriction enzymes NheI and XhoI. Linearized product d is then purified and ligated with the digested vector e. The vector e includes a pEF promoter and SigP sequence upstream of the NheI site and encodes the antibody constant heavy (CH) domains 1 to 3 downstream of the XhoI site. The product of ligation of insert d with vector e would result in plasmid f, which can be used to transform bacteria and growth with a suitable selectable marker would allow the production and purification of plasmid DNA by standard methods. Purified plasmid f can be introduced into mammalian cells [134] for heterologous Ig antibody expression. Alternatively DNA encoding CH1-3 in vector e, could be replaced with DNA encoding a single CH1 domain for Fab expression. V.sub.H and V.sub.L are antibody variable heavy and light chain respectively. DNA encoding an elongation factor promoter (pEF) an antibody constant light chain (CL) and constant heavy domains 1 to 3 (CH1-3), a polyadenylation sequence (pA) a cytomegalovirus (CMV) promoter and a signal peptide (SigP) are depicted.
[0252] FIG. 22. Additional example of preparation of DNA fragments required for the conversion of scFv to IgG (a) scFv inserts generated as described in Example 14 were separated on a 1% agarose TBE gel. Lanes 1 and 14 is a 500 bp DNA ladder starting at 500 bp. Lanes 2 to 13 are scFv PCRs. (b) The purification of the linearised "mini-circle" d (FIG. 21) was performed by separation on a 1% agarose TBE gel. From left to right, the first lane is a DNA ladder (1 kb ladder, Lifetech, 15615-024) and remaining lanes linearised "mini-circle" d. (c) As (b) except the CMV promoter is replaced by a P2A sequence and the DNA ladder employed was Generuler 1 kb ladder (Thermo, SM031D).
[0253] FIG. 23. Nuclease-directed integration of binder genes using flow electroporation systems. A 50:50 mix pD6 plasmids encoding either an anti-FGFR1 or an FGFR2 antibody was electroporated using a Flow electroporation system. After 13 days blasticidin selection cells were labelled with FGFR1-Fc labelled with Dyelight-633 (FGFR1-Dy633) or FGFR2-Fc labelled with Dyelight 488 (FGFR2-Dy488). Dot blots represent: [0254] a. Single staining FGFR2-488 (sample 1b from Table 7) [0255] b. single staining FGFR1-633 (sample 1b from Table 7) [0256] c. Dual staining FGFR1-633/FGFR2-488 (sample 1b from Table 7) [0257] d. Dual staining FGFR1-633/FGFR2-488 (sample 3 from Table 7)
[0258] FIG. 24. Recovery of antibody genes after flow sorting.
A population of antibodies was selected by one round of phage display and a mammalian display library was created by Flow electroporation using the Maxcyte system. Cells were sorted using either 1 nM or 10 nM antigen and mRNA isolated directly. The antibody genes were recovered by PCR, cloned into a bacterial expression vector and the proportion of ELISA positives determined ("1 nM output", "10 nM output"). This was compared with the original round 1 output ("R1 phage output"). Plot shows the profile of ELISA signals obtained with each population.
[0259] FIG. 25. pINT20 vector for expression of T Cell Receptors. [0260] a. Representation of the dual promoter plasmid pINT20 showing AAVS homology arms, puromycin selectable gene (with region around splice acceptor site shown below). Alpha chain (encompassing variable alpha, mouse alpha constant-CD3) is flanked by Nhe1, Not1 and Acc65I restriction sites and is under the control of pEF promoter. The beta chain (encompassing variable beta, mouse beta constant-CD3) is flanked by Nco1, XhoI and hind3 sites and is under the control of the CMV promoter. [0261] b. Sequence at the splice acceptor and beginning of puromycin gene (SEQ ID NO: 24 & 25) [0262] c. Sequence of T cell receptor clone c12/c2 alpha chain construct showing Nhe1, Not1 and Acc65I restriction sites (SEQ ID NO: 26 & 27). [0263] d. Sequence of T cell receptor clone c12/c2 beta chain construct showing Nco1. XhoI and Hind 3 restriction sites (SEQ ID NO: 28 & 29). [0264] e. Sequence of T cell receptor clone 4JFH alpha chain construct showing Nhe1/Not1 restriction sites (SEQ ID NO: 30 & 31). [0265] f. Sequence of T cell receptor clone 4JFH beta chain construct showing Nco1/XhoIrestriction sites (SEQ ID NO: 32 & 33). [0266] g. Strategy and primer used to mutate CDR3 of c12/c2 TCR alpha chain (SEQ ID NO: 34, 35, 36 & 37). [0267] h. Strategy and primer used to mutate CDR3 of c12/c2 TCR beta chain (SEQ ID NO: 38, 39, 40 & 41). (N=A, C, G, T; S'.sup.2 C OR G; W=A OR T)
[0268] FIG. 26. Recognition of peptide; MHC complexes by T cell receptors introduced into mammalian cells by nuclease-directed integration.
TCR1 is TCR c12/c2 recognising peptide 1 (SLLMWITQV) in the form of complex with phycoerythrin-labelled HLA-A2 (peptide 1). TCR2 is TCR 4JFH recognizing peptide 2 (ELAGIGILTV) in the form of complex with phycoerythrin-labelled HLA-A2 (peptide 2). Sample a and c show cells expressing TCR1 exposed to peptide 1 (a) or peptide 2 (c). Sample b and d show cells expressing TCR2 exposed to peptide 1 (a) or peptide 2 (c). Samples e and f show non-transfected HEK293 cells labelled with peptide 1 (e) or peptide 2 (f). g. Plasmid encoding TCR1 was mixed with 100 fold excess of TCR2 plasmid, introduced by nuclease-directed integration into HEK cells. 1.15% of cells and was labelled with peptide 1. 1.15% of cells were positive. h. Plasmid encoding TCR2 was mixed with 100 fold excess of TCR1 plasmid, introduced by nuclease-directed integration and was labelled with peptide 2. 0.62% of cells were labelled. Positive cells collected by flow sorting and mRNA recovered for analysis of specific TCR enrichment. Samples i-1 illustrate expression of a T cell library in HEK293 cells. TCR library was introduced by Maxcyte electroporation and selected for 11 days in puromycin I. Shows cells labelled with an APC labeled anti-TCR antibody (y axis). j. Shows cells labelled with phycoerythrin-labelled peptide 1:MHC (x axis) k. Shows untransfected cells labelled with both anti-TCR antibody and peptide 1:MHC l. Shows TCR1 library transfected cells labelled with both anti-TCR antibody and peptide 1:MHC Samples m-n illustrate expression of TCRs in Jurkat cells. TCR1 was delivered by Amaxa electroporation and selected for 25 days in puromycin. Plasmid was transfected in presence (m) or absence (n) of TALE nuclease and was incubated with an APC labelled anti-TCR .beta. chain antibody. Samples o-r illustrates T cell receptor activation of the same TCR1-transfected Jurkat cells. All cells are labelled with anti-CD69 antibody (y axis). Sample o was unstimulated and p was stimulated for 24 hours with an anti-CD3 antibody. Samples q and r were incubated for 24 hours with 2 ul and 6 ul respectively of PE labeled MHC:peptide 1. All cells were also exposed to CD28 antibody.
[0269] FIG. 27. pINT21 CAR1 and pINT21 CAR2 vectors for introduction of Chimeric Antigen Receptor (CAR) libraries into human cells.
Representation of the single promoter plasmid pINT21 showing AAVS homology arms, puromycin selectable gene, CMV promoter driving fusion of binder to CD3 signalling domain. Nco1 and Not 1 sites are sued for cloning the binder. [0270] a. pINT21 CAR1 fuses the binder to the juxtamembrane, transmembrane and signalling domain of CD3. [0271] b. pINT21 CAR2 fuses the binder to CD8 hinge and transmembrane domain, 4-1BB and CD3.xi. activation domains [0272] c. Sequence of CD3.xi. in pINT 21_CAR1 (SEQ ID NO: 42, 43 & 44) [0273] d. Sequence of CD8, 4-1BB and CD3.xi. in pINT 21_CAR2 (SEQ ID NO: 45 & 46) [0274] e. Sequence of FMC63 H-L (anti CD19 antibody) (SEQ ID NO: 47 & 48)
[0275] FIG. 28. Expression of scFv and alternative scaffold within chimeric antigen receptor construct introduced into human cells by nuclease-mediated integration.
HEK cells were transfected with anti-FGFR1 antibodies (b) or lox1 adhiron (d) and labelled with labelled FGFR1 and lox1 respectively. As control the same antigens were incubated with non-transfected HEK293 cells (a and c respectively). Populations from phage display libraries selected on mesothelin and CD229 were introduced into HEK cells by nuclease-mediated integration (f and h respectively) and were selected in puromycin for 11 days. These cells or untransfected HEK293 cells were incubated with labelled mesothelin (e, f) or CD229 (g, h). e and h represent untransfected HEK293 cells.
[0276] FIG. 29. Sequence of alternative binder scaffolds for mammalian display Libraries of different binder formats can easily be introduced by nuclease-directed integration using vector described herein. By example Adhiron constructs were prepared with flanking Nco1 and Not 1 sites for introduction into CAR constructs or Fc fusion constructs. [0277] a. Sequence of lox1 binding Adhiron_lox1A (SEQ ID NO: 49 & 50) [0278] b. Sequence of lox1 binding Adhiron_lox1B (SEQ ID NO: 51 & 52) (Variable loops are shown emboldened and underlined on the protein sequence). [0279] c. Potential mutagenic primers for construction of library of binders within loop 1 (adhiron mut1) (SEQ ID NO: 53, 55 & 56) or loop 2 (adhiron mut2) (SEQ ID NO: 54, 57 & 58), Below is a representation of the region covered by the primers (lower strand) showing protein translation. n represents variable number of NNS codons giving rise to different loop lengths. [0280] d. Sequence of trypsin binding knottin MCoTI-II with flanking Nco1 and Not1 sites allowing knottin expression within vectors described herein. Sequence of first loop is underlined SEQ ID NO: 59 & 60). [0281] e. Strategy for creation of library of knottin mutants. In this example loop 1 is replaced by 10 randomised amino acids. In this example VNS codons are introduced (V=A, C or G) providing 24 codons encoding 17 amino acids. This sequence can be introduced into a clone encoding MCoTI-II using standard methods (SEQ ID NO: 61, 62 & 63).
[0282] FIG. 30. Example sequences for Nuclease mediated antibody gene insertion by ligation or microhomology-mediated end-joining (MMEJ).
a. Sequence of pD7-SceI (nucleotides 1-120) (SEQ ID NO: 64, 65). The sequence is as pD6 (see FIG. 3c, 8) except the AAVS left arm between EcoR1 and Nsi1 has been replaced by the I-SceI meganuclease recognition (bold). Also the AAVS right arm between Asc1 and Mlu1 has been replaced by an insert encoded by primers 2723 and 2734 (not shown). b. Sequence of pD7-ObLiGaRe (nucleotides 1-120) (SEQ ID NO: 66, 67). The sequence is as pD6 (see FIG. 3c, 8) except the AAVS left arm between EcoR1 and Nsi1 has been replaced by the AAVS TALE right and left arm recognitions sites. Also the AAVS right arm between Asc1 and Mlu1 has been replaced by an insert encoded by primers 2723 and 2734 (not shown).
[0283] FIG. 31. Nuclease directed integration of binders into the ROSA 26 locus. [0284] a. Shows sequence of left homology arm up to the beginning of the puromycin gene showing primers and restriction sites mentioned in example 22 (SEQ ID NO: 68). [0285] b. Sequence of right homology arm for nuclease directed integration into the ROSA 26 locus showing primers and restriction sites mentioned in example 22 (SEQ ID NO: 69).
EXAMPLES
Example 1. Construction of Vectors for Expression of IgG Formatted Antibodies
[0286] To effect genetic selections of binders (e.g. antibody, protein or peptide) it is necessary to introduce a gene encoding this binder and to drive expression of this gene from an exogenous promoter, or by directing integration of the transgene downstream of a promoter pre-existing in the cellular DNA, e.g., an endogenous promoter. Antibodies represent the most commonly used class of binders and they can be formatted for expression in different forms. In examples below, we describe expression of a single gene format where a scFv is fused to a Fc domain (scFv-Fc). We also exemplify expression of antibodies formatted as human IgG2 molecules. To express IgG or FAb formatted antibodies in producer cells such as higher eukaryotes, it is necessary to express the separate heavy and light chains. This can be done by introducing separate plasmids encoding each chain or by introducing them on a single plasmid. Within a single plasmid the 2 chains can be expressed from a multi-cistronic single mRNA. Expression of distinct proteins from a single message requires elements such as an Internal Ribosome Entry (IRE) sequences which enables translation to initiate at a secondary downstream location. Alternatively, sequence elements promoting stalling/re-initation of translation such as viral 2A sequences could be used [119].
[0287] Alternatively, multiple distinct proteins can be expressed from a single plasmid using multiple promoters. FIGS. 1a and 1b show the organisation of 2 similar expression cassettes within different vector backbones (pDUAL and pINT3) which were developed for expression of secreted IgG formatted antibodies. These expression cassettes were created using a combination of gene synthesis and polymerase chain reaction amplification of standard elements such as promoters and poly A sequences. First separate plasmids were created within pCMV/myc/ER (FIG. 1c, Life Technologies) for expression of antibody heavy chain (pBIOCAM1-NewNot) and light chain (pBIOCAM2-pEF). The elements from pBIOCAM2-pEF (including pEF promoter, light chain gene and poly A site) were cloned into pBIOCAM1-NewNot) to create pDUAL. The examples shown include VH and VL domains from a humanised anti-lysozyme antibody called D1.3 [120] and are referred to as pDUAL-D1.3 and pINT3-D1.3. The elements of pDUAL D1.3 represented in FIG. 1a are present between the EcoR1 and BGH polyA site of the plasmid backbone from pCMV/myc/ER (Life Technologies Cat V82320 FIG. 1c).
[0288] In a similar way separate light chain and heavy chain cassettes were introduced into pSF-pEF (Oxford Genetics OG43) and pSF-CMV--F1-Pac1 (Oxford Genetics OG111) respectively to create pINT1 and pINT2. These were combined by cloning the light chain cassette (including pEF promoter, light chain gene and poly A site) upstream of the CMV promoter in pINT2, to create pINT3. The elements of pINT3-D1.3 represented in FIG. 1b are cloned between the first Bgl2 and the Sbf1 represented within the plasmid pSF-CMV--F1-Pac1 (FIG. 1d, Oxford Genetics OG111).
[0289] The immediate early promoter of cytomegalovirus (CMV promoter) is a powerful promoter and was used to drive expression of heavy chains. pDUAL D1.3 also incorporates an adenovirus 2 tripartite leader (TPL) and enhanced major late promoter (enh MLP) immediately downstream of the CMV promoter [121]. Elongation factor-1 alpha protein is ubiquitously and abundantly expressed in most eukaryotic cells and its promoter (pEF promoter) is commonly used for driving transgene expression [122]. In pDUAL-D 1.3 and pINT3-D1.3 the pEF promoter is used to drive antibody light chain expression. The polyadenylation sites originating in bovine growth hormone (BGH polyA) is present at the end of each expression cassette.
[0290] Secretion of the separate heavy and light chains in the endoplasmic reticulum (and ultimately culture supernatant) is directed by 2 different leader sequences. Light chain secretion is directed by a BM40 leader sequence [123]. This is followed by Nhe1 and Not1 cloning sites which allow in-frame cloning of VL genes which are in turn fused to a human C kappa gene. Secretion of the heavy chain is directed by a leader split by an intron originating from a mouse VH gene (as found in pCMV/myc/ER). The leader is followed by Nco1 and XhoI sites allowing in frame cloning of antibody VH genes followed by a codon optimised IgG2 gene. The VL and VH genes of the humanised D1.3 antibody [120] were cloned into the Nhe1/Not1 and Nco1/XhoI sites respectively within pDUAL-D1.3 and pINT3-D1.3.
[0291] Membrane anchored versions of these plasmids were created for mammalian display. Plasmid pD1 was created by digesting pDUAL-D1.3 with Bsu36I (which cuts in CH3 domain of the IgG2 heavy chain gene) and with BstZ171, which cuts in the backbone after the SV40 poly A region of the neomycin resistance cassette (FIG. 1c). This therefore removes most of the CH3 domain and the entire neomycin expression cassette. The CH3 domain is replaced by a synthetic insert with compatible Bsu36I and BstZ171 ends (represented in FIG. 1e). The synthetic insert was designed to replace the stop codon at the end of the antibody CH3 domain with a splice donor and intron which causes splicing of the CH3 terminus to an exon encoding the human PDGF receptor transmembrane domain [84] the first 5 intracellular residues, a stop codon and an additional splice donor. This is followed by an additional intron and splice acceptor followed by a codon for single amino acid then a stop codon (FIG. 1e). The 2 synthetic introns which flank the exon encoding the transmembrane domain were designed with ROX recognition sites located within them. ROX sites are recognized by Dre recombinase causing recombination between DNA containing these sites [88]. Inclusion of 2 ROX sites flanking the transmembrane domain-encoding exon creates the potential to remove this exon by the transfection of a gene encoding Rox recombinase. This would be anticipated to create a secreted antibody product.
[0292] FIG. 2 shows the sequence of the resulting dual promoter antibody expression plasmid expressing a humanised D1.3 anti-lysozyme antibody (hereafter referred to as pD1-D1.3 (SEQ ID NO: 1). Anti-lysozyme binding specificity is incorporated through inclusion of VH and VL sequences from D1.3 [120] between Nco/XhoI and Nhe1/Not1 restriction sites respectively. The sequence is shown from the ECoR1 site to the BstZ171. The sequences beyond the ECoR1 and BstZ171 sites are from the vector backbone as represented in FIG. 1c.
Example 2. Construction of Vector (pD2) for Targeting an Antibody Cassette to the AAVS Locus
[0293] Cleavage within the genome using site-specific nucleases facilitates the insertion of heterologous DNA through homologous recombination or non-homologous end joining (NHEJ). Human HEK293 cells were cleaved with nucleases targeting the first intron of the protein phosphatase 1, regulatory subunit 12C (PPP1R12C) gene. This locus was identified as a common integration site of adeno-associated virus and is referred to as the AAVS site (FIG. 3a). The AAVS site is considered a "safe harbour" locus for insertion and expression of heterologous genes in human cells [124].
[0294] Following site-specific cleavage within the genome it is possible to promote integration of a protein expressing cassette using homologous recombination. To do this it is necessary to flank the expression cassette with regions homologous to the sequences found on either side of the genomic cleavage site. To direct integration into the AAVS locus, an 804 bp section of the AAVS locus 5' to the intended cleavage site, was PCR amplified to create an EcoR1 and an Mfe 1 site at the 5' and 3' end respectively. This product, representing the left homology arm for targeting the antibody cassette, was cloned into the EcoR1 site of pD1 recreating the EcoR1 site at the 5' end. For the right homology arm an 836 bp section of the AAVS locus, 3' of the cleavage site, was PCR amplified to create Bstz171 sites at each terminus and this was cloned into the Bstz171 of pD1. The construct is represented in FIG. 3b and the sequence of the resulting construct (pD2) is shown in FIG. 3c.
[0295] During cloning of the AAVS left homology arm Nsi1 and Pac1 restriction sites were also inserted at the 3' end. These sites were subsequently used to clone a synthetic intron followed by a blasticidin gene with an accompanying poly A site. The blasticidin gene lacks a promoter but is preceded by a splice acceptor site that creates an in-frame fusion with the upstream exon from the AAVS locus (FIG. 3a, b). Integration into the AAVS locus causes expression of the promoter-less blasticidin gene. The sequence of this final construct, called pD2, is shown in FIG. 3c.
[0296] The sequence of the antibody cassette, encompassing the pEF promoter, D1.3 light chain, poly A region, CMV promoter, D1.3 heavy chain, alternative splice sites and poly A site, is shown in FIG. 2. To avoid duplication this sequence is represented in FIG. 3c as a block labelled "D1.3 ANTIBODY EXPRESSION CASSETTE".
Example 3. AAVS TALEN-Directed Integration of IgG Construct for Cell Surface Antibody Expression and Antigen Binding
[0297] HEK293F cells (Life Technologies), grown in Freestyle medium were transfected with pD2-D1.3 DNA in the presence or absence of an AAVS directed TALEN vector pair. An AAVS TALEN pair ("AAVS original") was previously described [125] and recognises the sequence:
TABLE-US-00001 LEFT TALEN: (SEQ ID NO: 70) 5' (T)CCCCTCCACCCCACAGT (SEQ ID NO: 71) Spacer 5' GGGGCCACTAGGGAC Right TALEN: (SEQ ID NO: 72) complement of 5' AGGATTGGTGACAGAAAA (SEQ ID NO: 73) (i.e. 5' TTTTCTGTCACCAATCCT
[0298] An alternative, more efficient AAVS targeted TALEN pair was identified and used in later experiments (pZT-AAVS1 L1 TALE-N and pZT-AAVS1 R1 TALE, Cat No GE601A-1 System Biosciences). This pair, which recognises the same site (but not the first "T" residue shown in brackets above), are referred to as the "AAVS-SBI" TALEN pair.
[0299] Cells were seeded at 0.5.times.10.sup.6 cells/ml and transfected next day at 10.sup.6 cells/ml using DNA:polyethylene imine (PolyPlus) added at a ratio of 1:2 (w/w). Cells were transfected with 0.6 .mu.g/ml of pD2 and were co-transfected with either pcDNA3.0 as a control (0.6 .mu.g/ml) or the combined left and right "original AAVS" TALEN plasmids (0.3 .mu.g each/ml). pD3 which expresses EGFP from the CMV promoter (see below) was included in the experiment as a transfection control and showed 35% transfection efficiency. Cells were selected in suspension culture using Freestyle medium (Life Technology) supplemented with 5 ug/ml blasticidin.
[0300] To determine whether antibody expression had occurred on the cell surface, cells were stained with an anti-human Fc antibody according to the following protocol:
1. 16 days after transfection, 0.5-1.times.10.sup.6 cells from the populations selected with Blasticidin were centrifuged for 2 minutes (200-300.times.g) at 4.degree. C. 2. Wash cells with 1 ml wash buffer (0.1% BSA in PBS Gibco #10010) and spin cells for 2 minutes (200-300.times.g) at 4.degree. C. 3. Resuspend cells in 100 .mu.l staining buffer (1% BSA in PBS) and add 5-10 ul of fluorochrome--conjugated antibodies. Antibodies were phycoerythrin-labelled anti-human IgG Fc (clone HP6017, Cat. No. 409304, Biolegend) or phycoerythrin-labelled mouse IgG2a, K isotype control (Cat. No. 400214, Biolegend). Incubate for >30 min at 4.degree. C. in the dark. 4. Wash twice with lml wash buffer and resuspend in 500 ul wash buffer. 5. Add 5 ul of cell viability staining solution (#00-6993-50 eBioscience) containing 50 ug/ml 7-amino-actinomycin D (7-AAD) to identify dead cells. 6. Cells were analysed on a (Beckton Dickinson FACS II) flow cytometer.
[0301] FIG. 4 shows that there was a significantly higher population of antibody expressing cells when pD2-D1.3 is transfected in the presence of the AAVS targeted TALEN with 86% positive compared with pD2-D1.3 alone with 1.5% positive.
[0302] The functionality of the surface expressed anti-lysozyme antibody was determined by assessing binding to labelled antigen. Hen egg lysozyme (Sigma: L6876) was labelled using Lightning-Link Rapid conjugation system (Dylight 488, Innova Biosciences: 322-0010) as follows:
1. Add 10 ul LL-Rapid Modifier reagent to 100 ul lysozyme (200 ug dissolved in 100 ul PBS) and mix gently. 2. Add the mix to Lightning-Link.RTM. Rapid mix and resuspend gently by pipetting up and down. 3. Incubate the mix for 15-30 minutes in the dark at room temperature. 4. Add 10 ul LL-Rapid Quencher reagent to the reaction and mix gently. 5. Store at 4.degree. C. Final concentration of lysosyme-Dy488 is 1.6 .mu.g/l. 6. Use 6 ul lysosyme-Dy488 (.about.10 ug) per staining. 7. Staining, washing and flow cytometry was as described above.
[0303] Analysis shows that 86% of cells transfected with pD2-huD1.3 bound labelled HEL (as judged by the M1 gate) compared with 0.29% for un-transfected cells (FIG. 5).
Example 4. Site-Specific Nucleases (AAVS Directed TALENs) Enhance Donor DNA Integration
[0304] Transfected cells were also plated out and selected with blasticidin to determine the number of cells in which expression of the promoterless blasticidin gene was activated. 24 hours after transfection cells were plated at 0.25.times.10.sup.6 cells/10 cm petri dish (tissue culture treated) and were grown in 10% foetal bovine serum (10270-10.sup.6, Gibco) and 1% Minimal Essential medium non-essential amino acid (MEM_NEAA #11140-035 Life Technologies). 5 ug/ml blasticidin was added after another 24 hours and medium was changed every 2 days. After 9 days cells which did not receive pD2 plasmid were all dead. After 12 days plates were stained with 2% methylene blue (in 50% methanol). Colony density was too high for accurate quantitation but showed an increased number of blasticidin resistant colonies in the presence of the AAVS TALENs suggesting targeted integration into the AAVS locus. A reduced amount of DNA was introduced for more accurate quantitation.
[0305] Transfections were carried out as described earlier using either 50, 200 or 400 ng pD2-D1.3/10.sup.6 cells in presence or absence of the AAVS TALENs (0.3 ug/ml of each TALEN where present, Table 1A). The total DNA input was adjusted to 1.2 ug DNA per 10.sup.6 cells with control plasmid pcDNA3.0. After 24 hrs of transfection, 0.25.times.10.sup.6 cells were plated in a 10 cm dish and 7.5 ug/ml blasticidin was added after 24 hrs of plating. 10 days after blasticidin selection the colonies are stained with 2% methylene blue (in 50% methanol). Results are shown in FIG. 6 and summarised in Table 1A. This shows that co-transfection of DNA encoding AAVS-directed TALENs increases the number of blasticidin resistant colonies achieved by approximately 10 fold.
[0306] A comparison was carried out between "AAVS original" and the "AAVS SBI" TALEN pairs targeting the AAVS locus. Table 1B shows an increased number of blasticidin resistant colonies using the "AAVS SBI" TALEN pair.
TABLE-US-00002 TABLE 1 Quantitation of blasticidin-resistant colonies from transfection of pD2-D1.3 A. pD2-D1.3 donor With AAVS Without AAVS Enzyme plasmid (ng/10.sup.6 cells) TALEN TALEN AAVS original 50 319 32 AAVS original 200 526 41 AAVS original 400 686 75 B. Without AAVS TALEN pD2-D1.3 donor With AAVS (Control Enzyme plasmid (ng/10.sup.6 cells) TALEN pcDNA3.0) AAVS original 300 1420 111 AAVS original 1000 1080 127 AAVS original 3000 560 70 AAVS-SBI 300 2800 111 AAVS-SBI 1000 1630 127 AAVS-SBI 3000 870 70
[0307] Here we have compared the effect of TALEN nuclease addition using either cell surface antibody expression (Example 3) or activation of a promoter-less blasticidin gene (Example 4). The benefit of nuclease-directed integration is more obvious when measuring antibody expression compared to effect on blasticidin-resistant colonies. One likely explanation is that the levels of expression required to effect survival in the presence of blasticidin may be significantly less than the expression levels required to detect IgG2 expression on the surface. Thus misincorporation/splicing of the promoter-less blasticidin gene could lead to a low level expression of the blasticidin resistance gene causing a higher background of blasticidin resistant colonies in the absence of significant antibody expression.
Example 5. Determination of Accuracy of Integration Using AAVS TALEN
[0308] To investigate the accuracy of integration, colonies were picked from the experiment in Example 4/Table 1A (from duplicate, unstained plates), expanded and genomic DNA from these cells was used as template in PCR. For preparation of genomic DNA, cells were harvested and were re-suspended in 700 .mu.L of lysis buffer (10 mM Tris.Cl, pH=8.0, 50 mM EDTA, 200 mM NaCl, 0.5% SDS, supplemented with 0.5 mg/mL of Proteinase K (added just before lysis). The cell re-suspension in lysis buffer was then transferred to a microfuge tube and kept at 60.degree. C. for about 18 hours. Next day, 700 .mu.L of isopropanol was added to the lysate in order to precipitate genomic DNA. The microfuge tube was spun at 13,000 rpm for 20 minutes. The genomic DNA pellet was then washed with 70% ethanol, and spun at 13,000 rpm for another 10 minutes. After spinning, the supernatant was carefully separated taking care not to touch the genomic DNA pellet. The genomic DNA pellet was then re-suspended in 100 .mu.L buffer containing 10 mM Tris (pH 8.0), and 1 mM EDTA and kept at 60.degree. C. for 30 minutes keeping the lid open in order to get rid of traces of ethanol. To this 100 .mu.L solution, RNAse A was added (final concentration of 20 g/mL), and incubated at 60.degree. C. for about one hour. Genomic DNA concentration was measured using nanodrop spectrophotometer (Nanodrop).
[0309] To identify correct integration, PCR primers were designed which hybridise in the AAVS genomic locus beyond the left and right homology arms. These were paired up with insert specific primers. At the 5' end the primers were:
TABLE-US-00003 AAVS-Left-arm-junction-PCR-Forw (9625) (SEQ ID NO: 74) 5' CCGGAACTCTGCCCTCTAAC BSD_Junction PCR-rev (9626): (SEQ ID NO: 75) 5' TAGCCACAGAATAGTCTTCGGAG
[0310] These give a product of 1.1 kb where correct integration occurs. 8/9 clones arising from AAVS directed integration gave a band of correct size (FIG. 7a, b). 2 blasticidin resistant clones derived without TALENs did not give a product (FIG. 11a) indicative of random integration. At the 3' end primers were:
TABLE-US-00004 Donor_plasmid_seq_PDGFRTM-2 Forw (SEQ ID NO: 76) 5' ACACGCAGGAGGCCATCGTGG AAVS1_right arm_junction_PCR_rev (SEQ ID NO: 77) 5' TCCTGGGATACCCCGAAGAG
[0311] These give a product of 1.5 kb with correct integration. 7/9 clones arising from AAVS directed integration gave a band of correct size. 2 blasticidin resistant clones derived without TALENs did not give a product (FIG. 11b). Thus the majority of blasticidin resistant cells arise from correct integration into the AAVS locus whereas blasticidin resistant colonies arising in the absence of TALENs are not correctly integrated.
Example 6. Construction of an scFv Display Library from a Selected Population from Phage Display and Selection Via Mammalian Display
[0312] scFv formatted soluble antibodies have previously been expressed from the vector pBIOCAM5-3F where expression is driven by the CMV promoter and the vector provides a C-terminal fusion partner, consisting of human Fc, His6 and 3.times.FLAG, to the antibody gene [105, 126]. This was modified to create the vector pBIOCAM5newNot where the Not1 site was embedded within the Fc region of the antibody (as shown in FIG. 8). This was used as a starting point to create the vector pD6 (FIG. 8) for expression of scFv-Fc fusions tethered to the cell surface. Primers (2598 and 2619) were designed to allow amplification of the CMV promoter-scFv-Fc expression cassette from pBIOCAM5newNotPrimer 2598 hybridises upstream of the CMV promoter and places a Pac1 site (bold text) at the end.
TABLE-US-00005 2598: (SEQ ID NO: 78) TTTTTTTTAATTAAGATTATTGACTAGTTATTAATAGTAATCAATTACGG GGTC
[0313] Primer 2619 hybridises near the end of the Fc domain and introduces a slice donor site and Pme1 site (bold text) at the beginning of the intron.
TABLE-US-00006 2619: (SEQ ID NO: 79) TTTTTTGTTTAAACTTACCTTGGATCCCTTGCCGGGGCTCAGGCTCAGGG AC
[0314] The resulting PCR product is compatible with the Pac1 and Pme1 sites of pD2 (FIG. 3).
Digestion of pD2 with Pac1 and Pme1 removes: pEF promoter-leader--light chain-CMV promoter-leader-heavy chain Cloning of the Pac1/Pme1 cut PCR product insets: CMV promoter-leader-Nco1/Not1 sites-human Fc.
[0315] Cloning in this way positions the scFv-Fc cassette appropriately for splicing to the downstream trans-membrane domain previously described for IgG presentation on the cell surface in pD2. The final vector pD6 is shown in FIG. 8, the sequence ofD6 from Nco1 to Pme1 sites is shown.
[0316] Phage display selections were carried out using the McCafferty phage display library [7] using beta-galactosidase (Rockland, Cat B000-17) and CD229 (R and D Systems, Cat 898-CD-050) as antigens. Methods for selection and sub-cloning were essentially as described previously [6, 7, 118, 127]. scFv genes from populations arising from one or two rounds of selection on beta-galactosidase and two rounds of selection on CD229 were recovered by PCR. Primers M13Leadseq hybridises within the bacterial leader sequence preceding the scFv gene and Notmycseq hybridises in the myc tag following the scFv gene in the phage display vector [127].
TABLE-US-00007 M13Leadseq (SEQ ID NO: 80) AAA TTA TTA TTC GCA ATT CCT TTG GTT GTT CCT Notmycseq (SEQ ID NO: 81) GGC CCC ATT CAG ATC CTC TTC TGA GAT GAG
[0317] PCR product was digested with Nco1 and Not1, the digested insert was gel purified. The digested product was ligated into the Nco1 and Not 1 sites of the bacterial expression plasmid pSANG10-3F and antibody expressed and screened as described [127]. After 2 rounds of selection on beta-galactosidase and CD229, 40/190 (21%) and 35/190 (18%) clones were found to be positive by ELISA.
[0318] 550 ng of Nco/Not cut insert was also ligated into the Nco1 and Not 1 sites of pD6 (2.4 .mu.g) to create a construct expressing a fusion between the scFv and the Fc region of human IgG2. Ligated DNA was transformed into electro-competent NEB5alpha cells (New England Biolabs, Cat C2989) which generated a library size of 2-3.times.10.sup.7 clones for each population. DNA was prepared and was co-transfected into 100 mls HEK293 cells grown in Freestyle medium as described above using 0.3 .mu.g donor DNA (pD6-library) per 10.sup.6 cells. Cells were co-transfected with 0.5 .mu.g each of "AAVS-SBI" TALENs (pZT-AAVS1 L1 TALE-N and pZT-AAVS1 R1 TALE, Cat No GE601A-1 System Biosciences).
[0319] 24 hours after transfection the volume of the bulk culture was doubled and 24 hours later blasticidin (10 .mu.g/ml) was added. Medium was refreshed every 3-4 days and after 6 days blasticidin concentration was increased to 20 .mu.g/ml.
[0320] In order to determine the library size, 20,000 cells were plated in a 10 cm petri dish (tissue culture treated) 24 hours after transfection and were grown in 10% foetal bovine serum (10270-106, Gibco) and 1% Minimal Essential medium non-essential amino acid (MEM_NEAA #11140-035 Life Technologies). 10 .mu.g/ml blasticidin was added after another 24 hours and medium was changed every 2 days. After 8 days plates were stained with 2% methylene blue (in 50% methanol). Results are shown in Table 2. This shows that libraries of around 3.times.10.sup.6 clones (representing 3% of transfected cells) were obtained for the 3 populations.
TABLE-US-00008 TABLE 2 Determination of scFv-Fc library size. No colonies/ No colonies/ Sample 20,000 cells 10.sup.6 cells Library size .beta.-galactosidase Rd1 546 27,300 2.7 .times. 10.sup.6 .beta.-galactosidase Rd2 654 32,700 3.2 .times. 10.sup.6 CD229 Rd2 556 27,800 2.8 .times. 10.sup.6
[0321] The protocol for labelling and flow sorting 10-20.times.10.sup.6 cells is shown below. Initial analysis was carried out 13 days post-transfection using only 10.sup.6 cells/sample and with reduced incubation volumes (reagent volumes that are 1/10th of those shown).
[0322] FIG. 9 shows that at 13 days post-transfection at least 43-46% of cells express scFv-Fc fusion on the cell surface and this can be detected using either FITC or phycoerythrin-labelled anti-Fc antibodies. Binding of biotinylated beta-galactosidase is also detected within this population using either FITC or phycoerythrin-labelled streptavidin. Using streptavidin-FITC 11.8% and 39% of the cell were positive for both antibody expression and antigen binding using libraries derived from output populations arising from 1 or 2 rounds of phage display selection respectively. For CD229 derived from 2 rounds of phage display, 66% of cells were positive for scFv-Fc and 24% of these were positive for CD229 binding (15% of the total population).
[0323] At 20 days after transfection cells were labelled according to the protocol below (using biotinylated antigen/phycoerythrin-labelled streptavidin and FITC-labelled anti human Fc).
1. Harvest, wash and adjust cells in 15-20.times.10.sup.6 cells per sample. Spin down cells at 250 g for 4', RT, wash cells with 1 ml PBS+0.1% BSA (4.degree. C.), spin down cells at 250 g for 4', RT, resuspend in 1 ml PBS+1% BSA 2. Add biotinylated antigen to a final conc. 100 nM and incubate 30' at 4.degree. C. 3. Wash the cells 2 times lml of 0.1% BSA by centrifugation at 1500 rpm for 5 minutes 4. Add either:
[0324] 10 .mu.l of FITC-labelled streptavidin (1 .mu.g/ml, Sigma Cat S3762) and 20 .mu.l of phycoerythrin-labelled anti human Fc (200 .mu.g/ml, BioLegend Cat. 409304), or:
[0325] 20 .mu.l phycoerythrin-labelled streptavidin (200 .mu.g/ml, Biolegend Cat 405203) and 20 .mu.l of FITC-labelled anti human Fc (200 .mu.g/ml, Biolegend Cat 409310) PBS+1% BSA, for 15 at 4.degree. C. in the dark
5. Wash the cells 2 times 1 ml of 0.1% BSA by centrifugation at 1500 rpm for 5 minutes 6. resuspend them in 500 .mu.l ice cold PBS+1% BSA 7. Add 20 ul of 7AAD/vial for viability staining
[0326] For sorting cells were gated on the basis of cell size, granularity, pulse width and viability (via 7-AAD staining, forward scatter and side scatter. Results are shown in FIGS. 9c and f In total 10 million cells were sorted and 3.1% and 7% of doubly positive cells were collected for libraries derived from output populations arising from 2 rounds of CD229 (CD229 R2) selection and 1 round of .beta.-galactosidase selection (.beta.-galR1) respectively.
[0327] Selected cells from the .beta.-galR1-derived cells were grown for a further 20 days and re-analysed (FIG. 9h). This shows that the majority of cells now express scFv-Fc and bind .beta.-galactosidase. This figure also shows that the proportion of double positive cells within the unselected population has not diminished 42 days after transfection (FIG. 9k).
[0328] Genomic DNA was prepared from 150,000-10.sup.6 sorted cells. Genomic DNA was prepared using method described earlier or using a GenElute mammalian genomic DNA miniprep kit (Sigma G1N10).
scFv genes were PCR amplified from genomic DNA using the following primers:
TABLE-US-00009 2623 (SEQ ID NO: 82) TAAAGTAGGCGGTCTTGAGACG 2624 (SEQ ID NO: 83) GAAGGTGCTGTTGAACTGTTCC
[0329] PCR reactions were carried out using Phusion polymerase (NEB Cat M0532S) in manufacturer's buffer containing 0.3 uM of each primer and 3% DMSO. 100-1000 ng of genomic DNA was used as template in a 50 ul reaction. 30 cycles were carried out at 98.degree. C. for 10 secs, 55.degree. C. for 25 secs, 72.degree. C. for 45 secs. This gave a product of 1.4 kB which was digested with Nco1 and Not1. A band of approximately 750-800 bp was generated and gel purified before cloning into pSANG10. Ligated DNA was transformed into BL21 cells (Edge Bio Ultra BL21 (DE3) competent cells, Cat. 45363). In this way scFv fragments derived from the sorted population can be expressed in bacteria as described previously [7, 127].
[0330] As an alternative to isolating the antibody gene and expressing in an alternative vector/host combination, it is possible to derive secreted antibody directly from the selected cells either following single cell cloning or using a sorted population to generate a polyclonal antibody mix. To exemplify this culture supernatant was taken from sorted cells (from .beta.-galR1 cells) after 7 days in culture. This was shown to be positive in ELISA using plates coated with (3-galactosidase (see Example 13 and FIG. 19b).
Example 7. Construction and Selection from an IgG Display Library from a Selected Population from Phage Display
[0331] DNA fragments encoding scFv, representing the round 1 and 2 antibody phage display outputs of selections against .beta.-galactosidase and CD229, were generated as described in Example 6. The scFv populations were converted to IgG format according to Example 14 and as detailed in the method below.
[0332] A DNA insert encoding the human kappa light chain constant domain (CL), polyadenylation sequence (pA), CMV promoter and signal peptide from murine V.sub.H chain (represented between the Not1 and Nco1 sites of pD2 shown in FIG. 21b) was PCR amplified from plasmid pD2 with primers 2595 (GAGGGCTCTGGCAGCTAGC) (SEQ ID NO: 84) and 2597 (TCGAGACTGTGACGAGGCTG) (SEQ ID NO: 85). PCR reactions were carried out using KOD hot start polymerase (Novagen Cat 71086-4) in manufacturer's buffer containing 0.25 .mu.M of each primer. 10 ng of pD2 plasmid DNA was used as template in a 50 ul reaction. 25 cycles were carried out at 98.degree. C. for 10 secs, 55.degree. C. for 25 secs, 72.degree. C. for 40 secs. This gave a product of 1.8 kB which was digested with Nco1 and Not1 and gel purified (FIG. 20a represented as CL-pA-CMV-SigP insert in FIG. 21b).
[0333] DNA fragments encoding scFv, representing the round 1 and 2 antibody phage display outputs of selections against .beta.-galactosidase and CD229, were generated as described in Example 6. FIG. 20b shows scFv populations selected against .beta.-galactosidase and CD229 separated by 1% agarose gel electrophoresis.
[0334] Ligations between scFv insert and CL-pA-CMV-SigP insert were performed by incubating NcoI/NotI digested scFv insert (1 .mu.g) with NcoI/NotI digested CL-pA-CMV-SigP insert (1 .mu.g) with T4 DNA ligase (1.5 al, Roche, 10-481-220-001) in manufacturer's buffer in a total volume of 40 .mu.l to form the "mini-circle" depicted in FIG. 21c. Ligations were incubated at 16.degree. C. for 16 hours, purified by spin column, digested with NheI and XhoI and the 2.6 kb product (depicted in FIG. 21d) purified by electrophoretic separation on 1% agarose gel (FIG. 20c).
[0335] The DNA insert depicted in FIG. 21d encoding V.sub.L-C.sub.L-pA-CMV-SigP-V.sub.H (0.5 .mu.g) was ligated with NheI/XhoI digested, gel purified vector pD2 (0.7 .mu.g) (FIG. 21e) with T4 DNA ligase (1.5 .mu.l, Roche, 10-481-220-001) in manufacturer's buffer in a total volume of 40 .mu.l to produce the targeting vector depicted in FIG. 21f. This encodes populations of antibodies formatted as IgGs, originating from first or second round antibody phage display selections to (3-galactosidase or CD229. Ligations were incubated at 16.degree. C. for 16 hours, purified by spin column and eluted with HPLC grade water.
[0336] Ligated DNA was transformed into electro-competent NEB5alpha cells (New England Biolabs, Cat C2989) which generated a library size of 1-4.times.10.sup.5 clones for each population. DNA was prepared and was co-transfected into 100 mls HEK293 cells grown in Freestyle medium as described above using 0.3 Lg donor DNA (pD6-library) per 10.sup.6 cells. Cells were co-transfected with 0.5 .mu.g each of"AAVS-SBI" TALENs (pZT-AAVS1 L1 TALE-N and pZT-AAVS1 R1 TALE, Cat No GE601A-1 System Biosciences).
[0337] 24 hours after transfection the volume of the bulk culture was doubled and 24 hours later blasticidin (10 .mu.g/ml) was added. Medium was refreshed every 3-4 days and after 6 days blasticidin concentration was increased to 20 ag/ml.
[0338] In order to determine the library size, 250,000 cells were plated in a 10 cm petri dish (tissue culture treated) 24 hours after transfection and were grown in 10% foetal bovine serum (10270-106, Gibco) and 1% Minimal Essential medium non-essential amino acid (MEM_NEAA #11140-035 Life Technologies). 10 .mu.g/ml blasticidin was added after another 24 hours and medium was changed every 2 days. After 8 days plates were stained with 2% methylene blue (in 50% methanol). Results are shown in Table 3. This shows that libraries of between 5.times.10.sup.5 and 9.times.10.sup.5 clones (representing 0.5% to 0.9% of transfected cells) were obtained for the 3 populations.
TABLE-US-00010 TABLE 3 Determination of size of mammalian display libraries formatted as IgG. No. colonies/ No. colonies/ Library Sample 0.25 .times. 10.sup.6 cells 10.sup.6 cells size (.times.10.sup.5) .beta.-galactosidase Rd1 1337 5348 5.3 .beta.-galactosidase Rd2 1972 7888 7.9 CD229 Rd2 2175 8700 8.7
[0339] The bulk of the population of cells transfected with the outputs from 1 or 2 rounds of selection on .beta.-galactosidase were selected in blasticidin containing medium as described earlier. After 19 days 10-20.times.10.sup.6 cells were labelled and flow sorting carried out as described in example 6. Sorted cells were grown for 17 days and re-analysed by flow cytometry (FIG. 10). This showed that the majority of cells were now double positive for IgG expression and binding to .beta.-galactosidase.
[0340] Genomic DNA was prepared from the sorted cells and DNA encoding the IgG insert was isolated by PCR. The IgG-encoding insert was amplified using KOD polymerase (Merck, cat. no. 71086-3), with annealing temperature of 60.degree. C. and employing 30 cycles. Manufacturer provided buffer with 5% DMSO was used with 0.3 .mu.M of primers 2597 (SEQ ID NO: 54) and 2598 (SEQ ID NO: 47). The product of desired size was gel purified. The gel purified product was then used for nested PCR using KOD polymerase (Merck, cat. no. 71086-3) in manufacturer's buffer with 5% DMSO using 0.3 .mu.M of primer 2625 (SEQ ID NO: 55) in combination with either primer 1999 (SEQ ID NO: 56) (for R1 sample), or 2595 (SEQ ID NO: 53) (for 4R1 and 5R1), using annealing temperature of 60.degree. C. employing 30 cycles. These nested PCR products were gel purified and subjected to double digestion with NheI-HF (NEB, cat. no. R3131S) and XhoI (NEB, cat. no. R0146S) in order to ligate them with similarly double digested pINT3 (FIG. 1) for expression of soluble IgG formatted binders. Primer sequences are:
TABLE-US-00011 2597: (SEQ ID NO: 85) AGGGGTTTTATGCGATGGAGTT 2598: (SEQ ID NO: 78) GTTACAGGTGTAGGTCTGGGTG 2625: (SEQ ID NO: 86) CCTTGGTGCTGGCACTCGA 1999: (SEQ ID NO: 87) AAAAAGCAGGCTACCATGAGGGCCTGGATCTTCTTTCTCC 2595: (SEQ ID NO: 84) GAGGGCTCTGGCAGCTAGC
Example 8. Construction and Selection from a Naive scFv Library
[0341] Schofield et al. [7] describe the construction of a phage display library (McCafferty library") wherein antibody genes from the B-lymphocytes of a number of human donors were first cloned into an "intermediate library" before re-cloning into the final functional phage display library. This same intermediated library and the same methodology was used to generate a new library (IONTAS library) of 4.times.10.sup.10 clones. Plasmid DNA was prepared from this library taking care to ensure sufficient representation of the library within the bacterial inoculation. A number of PCR reactions were set up using a total of 2 ug of DNA template. The PCR product was digested with Nco1 and Not 1 gel purified and ligated as described in Example 6. 9.3 ug of pD6 and 0.93 ug of PCR insert were ligated overnight, the ligation reaction cleaned up using phenol chloroform extraction and the DNA electroporated into DHSalpha cells as described previously [7]. As a result a library of 2.4.times.10.sup.8 clones was created within the scFv-Fc display vector. DNA was prepared from this "naive library" cloned in pD6 and transfected into 1 litre of HEK293F cells (Life Technologies) grown in Freestyle medium (as described above). 0.3 ug pD6-library DNA, 0.5 ug of each "AAVS-SBI" TALEN pair. 24 hours after transfection the culture volume was doubled and 48 hour after transfection Blasticidin selection was commenced as described above. The library size was determined by plating aliquots of the culture 24 hours after transformation and selecting on blasticidin as described above. A library of 0.9.times.10.sup.7 clones was created.
[0342] A number of antigens were biotinylated using EZ-Link Sulfo-NHS-LC-Biotin kit (Pierce Cat. No. 21327) according to manufacturer's instructions. Antigens were bovine thyroglobulin (Calbiochem Cat. 609310), human CD28-Fc chimera (R and D Systems, Cat 342-CD-200) and mouse EphB4-Fc chimera (R and D Systems, Cat. 446-B4-200). Biotinylated .beta.-galactosidase (Rockland Cat. B000-17) was also used.
[0343] Transfected cells were selected in blasticidin in liquid culture as described above for 17 days. Cells were harvested, washed and adjusted to 15-20.times.10.sup.6 cells per sample. Cells were prepared as described and biotinylated antigen added to a concentration of 500 nM. Labelling and flow sorting a as described above. Using control cells incubated with only the phycoerythrin-labelled anti-Fc antibody a "gate" was created which included 0.05% of these cells. Using the same gate for labelled cells between 0.28-0.51% of cells were included (FIG. 11). These were collected and grown to allow additional rounds of sorting and amplification of scFv genes from the naive library.
Example 9. Creation of a Cell Line with Multiple "Landing Sites" to Compare Nuclease and Recombinase-Directed Approaches to Genomic Integration
[0344] To allow comparison of integration methods based on either genomic cleavage or recombinase-mediated integration an AAVS-directed targeting vector (pD4) was constructed which introduces an intron with multiple "landing sites" (Example 3). These include an FRT site recognized by Flp recombinase and a pair of a lox2272/loxP sites recognized by Cre recombinase. To allow targeted cleavage, pD4 also includes a sequence from GFP for which a TALEN pair have been designed [128] and an I-Sce 1 meganuclease site to allow endonuclease-directed integration. A compatible incoming donor plasmid was constructed (pD5) with appropriate recognition sites such that nuclease or recombinase directed integration causes activation of a promoter-less blasticidin gene and integration an antibody expression cassette.
[0345] The organisation of the plasmid and the sequence of pD4 is shown in FIG. 12b. An intermediate plasmid pD3 was first created which encompasses a GFP gene under the control of the CMV promoter followed by a puromycin/Thymidine kinase gene fusion under the control of the PGK promoter (FIG. 12a). This was created by digesting pBIOCAM1-newNot with Sac1 (at the end of the CMV promoter) and BstB1 (between the Neo gene and the poly A site, FIG. 1a). This removed the neomycin expression cassette and allows replacement with a synthetic insert encompassing an enhanced Green Fluorescent Protein (EGFP) gene under the control of the CMV promoter. This GFP construct was fused at the C terminus to residues 422-461 of a mutated mouse ornithine decarboxylase PEST sequence. This PEST sequence is incorporated in plasmid pZsGreenl-DR (Clontech) and has been shown to reduce the half-life of the fused GFP to 1 hour. A cassette encoding a PGK promoter, Puromycin/Thymidine kinase gene fusion (Puro deltaTK) and polyA cassette was excised from the plasmid pFLEXIBLE [129] using Xmnl and Fsel and was cloned into Smal and Fsel sites present in the original synthetic insert. The resultant plasmid (called pD3) encodes a CMV-driven GFP gene and a puromycin resistance gene driven from a PGK promoter.
[0346] To create the final targeting vector pD4, the CMV promoter was removed and AAVS homology arms were inserted. An 850 bp section of the AAVS locus was PCR amplified to create an AAVS left homology arm flanked by an EcoR1 at the 5' end and an Mre1 at the 3' end. This was cloned into the ECoR1/Mre1 site of pD3 thereby removing the CMV promoter. An Nsi1 site was also incorporated at the 3' end of this AAVS left homology arm. The neighbouring Mre1 and Nsi1 sites were used to introduce a synthetic fragment fusing an intron to the EGFP gene as shown in FIG. 13. The synthetic intron preceding the EGFP gene incorporates:
a FRT recognition site for Flp recombinase a lox 2272 recombination site an I-SceI meganuclease site GFP TALEN recognition site a T2A ribosomal stalling sequence [130]
[0347] The AAVS right homology arm was generated by PCR to create a Hpa1 and BstZ171 sites at 5' and 3' ends. This fragment was cloned into the Hpa1 and BstZ171 sites of pD3. The resulting plasmid pD4, encodes a puromycin resistance cassette ("Puro deltaTK") and can be used to introduce a "landing sites" into the AAVS locus incorporating various nuclease and recombinase sites for comparison. The sequence of pD4 is shown in FIG. 13 (SEQ ID NO: 5, SEQ ID NO: 6 and SEQ ID NO: 7). The AAVS left and right targeting arms are shown in detail in FIG. 3 and are therefore abbreviated in FIG. 13.
[0348] The intron introduced into pD4 contains a TALEN recognition sites originating from GFP [128]. The eGFP directed TALEN pair (eGFP-TALEN-18-Left and eGFP-TALEN-18-right) recognise the sequence shown below (all sequences and primers are presented in a 5' to 3' direction) where capitals represent the recognition site of left and right TALENs and the lower case shows the spacer sequence. The right hand TALEN recognises the complement of the sequence shown. The first base pair is equivalent of the minus16 position of the sequence relative to the initiating ATG sequence of GFP (shown bold in the spacer).
TABLE-US-00012 (SEQ ID NO: 88) TCCACCGGTCGCCAccatggtgagcaagggCGAGGAGCTGTTCA
[0349] The plasmid pD4 also incorporates an I-SceI meganuclease site and an FRT site recognised by Flp recombinase. Finally pD4 incorporates lox 2272 and loxP (which are mutually incompatible) flanking the GFP and puromycin expression cassettes. Incorporation of these same 2 loxP sites flanking the donor plasmid (pD5 below) affords an opportunity to substitute the integrated cassette (including the PGK puro delta TK cassette) replacing it with an incoming cassette driving expression of blasticidin and antibodies via recombinase mediated cassette exchange.
Creation of a Cell Line by Transfection of pD4.
[0350] HEK293F cells were resuspended at 10.sup.6 cells/ml and DNA:polyethylene imine (PolyPlus) added at a ratio of 1:2 (w/w). Cells were transfected with 0.6 .mu.g/ml of pD4 and were co-transfected with either the "original AAVS" TALEN pair or pcDNA3.0 as a control (0.6 g/ml). pD3 which expresses EGFP from the CMV promoter was included in the experiment as a transfection control and showed 35% transfection efficiency. After 24 hours transfected cells were plated at 0.5.times.10.sup.6 cells/10 cm petri dish (tissue culture treated) and were grown in 10% foetal bovine serum (10270-106, Gibco) and 1% Minimal Essential medium non-essential amino acid (MEM_NEAA #11140-035 Life Technologies). 5 .mu.g/ml puromycin was added after another 24 hours and medium was changed every 2 days. After 5 days untransfected cells or cells transfected with pD3 only were dead. After 12 days there were approximately 200 colonies on cells transfected with pD4 only and approximately 400 colonies on cells transfected with pD4 and the AAVS TALEN pair.
[0351] The puromycin resistant population arising from transfection with pD4 and the AAVS TALEN pair was analysed for correct integration. In addition a single colony was picked from this population (clone 6F) and compared with a colony from the puromycin resistant population arising from pD4 transfection in the absence of the AAVS TALEN pair. To identify correct integration, PCR primers were designed which hybridise in the AAVS genomic locus beyond the left and right homology arms. These were paired up with insert specific primers. At the 5' end the primers were:
TABLE-US-00013 AAVS1_HA-L_Nested_Forw1 (SEQ ID NO: 89) GTGCCCTTGCTGTGCCGCCGGAACTCTGCCCTC EGFP_Synthetic_gene_Rev_Assembly (SEQ ID NO: 90) TTCACGTCGCCGTCCAGCTCGAC Purotk_seq_fow2 (SEQ ID NO: 91) TCCATACCGACGATCTGCGAC AAVS1_Right_arm_Junction_PCR_Rev (SEQ ID NO: 77) TCCTGGGATACCCCGAAGAG
[0352] FIG. 14 shows that clone 6F and the population are correct at both left and right ends but the clone picked from the non-AAVS directed population is negative. Thus PCR analysis indicates that the accuracy of integration of the donor cassette is greater when directed by AAVS TALEN cleavage of genomic DNA.
[0353] pD4 introduces a promoter-less, in-frame GFP gene driven from the AAVS promoter. Flow cytometry of the puromycin resistant population showed an absence of GFP expression. This failure to express could be due to the combination of a short half-life (from the murine ornithine decarboxylase PEST sequence element) combined with reduced expression arising from the use of the T2A promoter. In fact it was found that addition of the T2A element in front of a promoterless blasticidin element (as described for pD2) reduced the number of blasticidin resistant colonies by 4 fold. Despite the absence of GFP expression, the integration of multiple landing sites still affords an opportunity for comparison of recombinase-directed versus DNA cleavage directed genomic integration.
Example 10. Construction of a Vector for Inserting an Antibody Cassette (pD5) into the "Multiple Landing" Site
[0354] Following introduction of the "multiple landing site" intron into the AAVS locus, it is possible to introduce an antibody cassette via nuclease-directed or recombinase-directed means. To do this a donor plasmid pD5 was created where the expression cassette is flanked by left and right homology arms which are equivalent to the sequences flanking the GFP TALEN cleavage site introduced into pD4. pD5 does not itself incorporate an intact GFP TALEN recognition site and integration is driven by homologous recombination. Homology-directed integration of the donor plasmid will lead to introduction of a blasticidin gene which lacks a promoter but is preceded by a splice acceptor site that creates an in-frame fusion with the upstream exon from the AAVS locus as described earlier. Integration into the AAVS locus will cause expression of the promoter-less blasticidin gene. The inserted cassette also encodes an IgG formatted antibody heavy and light chains under the control of pEF and CMV promoters respectively as described above. pD5 incorporates an I-SceI meganuclease site which can lead to cleavage of the incoming donor affording an opportunity for NHEJ (see Example 12). An FRT site is also incorporated into the donor plasmid pD5 allowing recombinase-directed incorporation of the promoter-less blasticidin gene and antibody expression cassettes at the same locus. As discussed above Cre recombinase will act on the loxP sites in donor and genomic DNA to direct recombinase-mediated cassette exchange.
[0355] The sequence of pD5 is shown in FIG. 15. The sequence at the 5' end of the GFP TALEN site is from the AAVS locus. A 267 bp section of the AAVS locus upstream of the TALEN cleavage site was generated by PCR. Primers were used which created an EcoR1 and an Mfe 1 site at the 5' and 3' end and the product was cloned into the ECoR1 site of pD1-D1.3. The EcoR1 site is re-created at the 5' end. During cloning of the left homology arm an Nsi1 and Pac1 was also inserted at the 3' end. A right homology arm, incorporating approximately 700 bp equivalent to the sequence 3' of the GFP TALEN was created by PCR assembly. PCR primers introduce BstZ171 sites at 5' and at the 3' end of the assembled fragment and this was cloned into the BstZ171 site of pD1-D1.3. The PCR primers also introduced a Hpa1 site at the 5' end.
[0356] A PCR fragment encompassing the intron (which incorporates recognition sites for GFP TALEN, I-Scelendonuclease, Flp recombinase and Cre recombinase), a splice acceptor region, a Blasticidin resistance gene and poly A site (described above) was created with Nsi1 site at the 5' end and a Pac1 site at the 3' end. This was cloned into the Nsi1 and Pac1 site of the plasmid described above to create pD5-D1.3 (sequence shown in FIG. 15 and plasmid structure shown in FIG. 18a).
Example 11 Comparison of Nuclease-Directed and Flp-Directed Integration of an Antibody Expression Cassette
[0357] The Flp-In system which has previously been used for recombinase-mediated integration of antibody expression cassettes [18] uses a mutant Flp recombinase (in the plasmid pOG44) which possesses only 10% of the activity at 37.degree. C. of the native Flp recombinase [19]. A variant of Flp recombinase (Flpe) with better thermostability and activity at 37.degree. C. than wild type has been identified [19, 20]. This was further improved by codon optimization to create Flpo [131] encoded within plasmid cCAGGS-Flpo (Genebridges Cat. A203). The effect of both variants of Flp recombinase (encoded within pOG44 and cCAGGS-Flpo) was compared. Recombination directed by Cre recombinase was also examined by co-transfecting cells with a plasmid which encodes Cre recombinase [132] (pCAGGS-Cre, Genebridges Cat. A204). In each vector the recombinase is expressed under the control of the chicken-1-actin promoter and a CMV immediate early enhancer. An SV40 Large T nuclear localization sequence is used for nuclear localisation [20]. In the original vectors (cCAGGS-Flpo and pCAGGS-Cre) recombinase expression was linked to a puromycin resistance gene by an internal ribosomal entry site (IRES) which was removed using standard molecular biology techniques.
[0358] An experiment was carried out to compare the efficiencies of genomic cleavage-directed versus recombinase directed integration of an antibody cassette. The outcome was assessed in 2 ways:
1. Measuring the number of blasticidin-resistant colonies arising from integration of a promoter-less blasticidin gene 2. Assessing the extent of antibody expression achieved by the different approaches.
[0359] As described in Example 9, the recognition sites for Cre recombinase (lox2272 and loxP) and Flp recombinase (FRT) were previously integrated into the AAVS locus within clone 6F. In addition recognition sites for a GFP TALEN pair and for the meganuclease I-SceI are also present within the same intron. The donor plasmid pD5-D 1.3 carries the same recognition sites (apart from GFP TALEN) within an intron upstream of a promoter-less blasticidin gene. Correct integration will lead to activation of the blasticidin gene. pD5-D1.3 also encodes an IgG formatted D1.3 antibody gene which will be expressed on the cell surface.
[0360] Co-transfection of pD5-D1.3 with pOG44 or pCAGGS-Flpo (encoding 2 variants of Flp recombinase) should result in integration of the entire pD5 plasmid into the FRT site of clone 6F. The donor plasmid pD5-D1.3 also has lox2272 site within the synthetic intron upstream of the blasticidin gene and a loxP site at the end of the antibody expression cassette. Under the action of Cre recombinase expressed from pCAGGS-Cre, recombinase-mediated cassette exchange should result in the integration of the blasticidin and antibody expression cassettes into the lox2722 and loxP sites within clone 6F.
[0361] The efficiency of vector integration using recombinase-directed approaches with genomic cleavage-directed approaches was compared using a pair of TALENs (eGFP-TALEN-18-Left and eGFP-TALEN-18-right) directed towards a sequence from GFP (Reyon et al., 2012). In the case of GFP TALENs the element between the left and right homology arms will be integrated following genomic cleavage by TALENs.
[0362] To allow comparison with I-SceI meganuclease a codon optimised gene encoding I-Sce1 was constructed (FIG. 16). This gene has an N terminal HA epitope tag/SV40 nuclear localisation signal (NLS) at the N terminus and is flanked by Nco1 and Xba1 sites at the 5' and 3' termini. The gene was cloned into the vector pSF-CMV-F1-Pac1 (Oxford Genetics OG111) where expression is driven from the CMV promoter.
[0363] Transfections were carried out using the clone 6F with a correctly integrated "multiple landing site". Cells were suspended at 10.sup.6/ml and transfected with 50 ng of pD5-D1.3 donor plasmid/10.sup.6 cells along with enzyme encoding plasmids (Table 4A).
[0364] After 24 hours transfected cells were plated at 0.5.times.10.sup.6 cells/10 cm petri dish (tissue culture treated) and were grown in 10% foetal bovine serum (10270-106, Gibco) and 1% Minimal Essential medium non-essential amino acid (MEM_NEAA #11140-035 Life Technologies). 5 ug/ml blasticidin was added after another 24 hours and medium was changed every 2 days. After 12 days plates were stained with 2% methylene blue (in 50% methanol) and colony numbers counted (Table 2). In a direct comparison between Flp recombinase, Cre recombinase and TALEN, the greatest number of colonies were obtained through use of the GFP TALEN where there was a 9-fold increase compared with "donor only" (Table 4A). It also appears that the use of the optimised Flpo gene actually resulted in a reduction of the number of blasticidin resistant colonies compared to "donor only" control, presumably through toxicity of the enhanced activity Flp recombinase. There was also an increase in colony number using pCAGGS-Cre compared to the donor only control.
[0365] A second experiment was carried out comparing GFP TALEN with both enhanced Flp (from cCAGGS-Flpo) as well as the low activity Flp enzyme encoded within pOG44 from the Flp-In system (as used by Zhou et al. [17, 18, U.S. Pat. No. 7,884,054]. These were compared with GFP TALEN and Cre recombinase (Table 4B). Cells were transfected with the amounts DNA shown per million cells. 0.25.times.10.sup.6 cells were plated out and the number of blasticidin resistant colonies determined as described above. Cells were also selected for blasticidin resistance in liquid culture for 30 days before determining the proportion of cells which expressed surface IgG (as described above). Table 4B shows that the TALEN was superior to the other approaches in terms of number of resistant colonies. Again the use of optimised Flp within cCAGGS-Flpo actually caused a reduction in number of blasticidin resistant colonies compared to "donor only" controls. Cre recombinase again led to an increase in blasticidin colony count compared with control whereas the Flp gene within pOG44 showed only a marginal increase compared with control.
TABLE-US-00014 TABLE 4 Comparison of TALE nuclease-directed and recombinase- directed integration approaches. A. pD5-D1.3 donor Blasticidin (ng/10.sup.6 resistant colonies Enzyme plasmid Amount cells) (colonies/10.sup.6 cells) GFP TALEN pair 0.575 .mu.g each 50 152 (304) pCAGGS-Flp.sub.o 1.15 .mu.g 50 1 (2) pCAGGS-Cre 1.15 .mu.g 50 57 (114) control 1.15 .mu.g 50 17 (34) (pCDNA3.0) B. Blasticidin colonies per 10.sup.6 cells Sample pD5-D1.3 (percentage Percentage 2 .mu.g each per Donor Blasticidin blasticidin positive 10.sup.6 cells .mu.g/10.sup.6 cells colonies resistant) in flow GFP TALEN pair 0.6 270 1080 (1.1%) 95.6 cCAGGS-Flp.sub.o 0.6 35 140 (0.14% Too few cells pOG44 0.6 96 384 (0.38%) 6.4 pCAGGS-Cre 0.6 180 720 (0.72%) 4 Control 0.6 81 324 (0.32%) 37.3 (PCDNA3.0) C. Sample 2 .mu.g each pD5-DL3 Donor Blasticidin Percentage per 10.sup.6 cells .mu.g/10.sup.6 cells colonies positive in flow GFP TALEN 2 210 95.3 GFP TALEN 6 120 73.6 cCAGGS-F1p.sub.o 2 62 Too few cells cCAGGS-F1p.sub.o 6 41 Too few cells pOG44 7 178 35.6 pOG44 6 63 58.7 pCAGGS-Cre 2 84 40.9 pCAGGS-Cre 6 52 Too few cells Control 2 340 65.6 (PCDNA3.0) Control 6 82 55.5 (PGDN A3.0)
[0366] With the addition of more donor DNA (Table 4C) there was an increase in colony numbers at intermediate levels (2 .mu.g/million cells) and a decline at higher levels across the board (6 .mu.g/million cells). None of the other samples achieved levels of antibody display seen with the GFP TALEN directed integration.
[0367] Cells were also selected for blasticidin resistance in liquid culture and the cells stained for antibody expression as described above. TALEN-directed integration gave a significantly higher proportion of antibody-positive cells compared with the other approaches. Cells transfected with cCAGGS-Flpo and with high concentrations of pCAGGS-Cre were not healthy and there were insufficient numbers to carry out flow cytometry.
[0368] The comparison was extended to included I-Sce endonuclease. A synthetic gene encoding I-SceI was synthesised (FIG. 16) and cloned into the Nco1/Xba 1 site of pSF-CMV-f1-Pac1 (Oxford Genetics). Cells were suspended at 10.sup.6/ml and transfected for each ml of cells (10.sup.6 cells/ml) with 300 ng of pD5-D1.3 donor plasmid along with plasmids encoding enzymes (1 ug/10.sup.6 cells). Next day 0.05 ml of cells were plated and selected in blasticidin and stained after 14 days as described. Table 5 shows that the highest number of blasticidin resistant colonies came from I-SceI meganuclease followed by the eGFP TALEN pair. Both Cre and Flp recombinase (encoded within pOG44) gave numbers slightly higher than the "donor only" control. As before transfection with the Flpe encoding plasmids actually reduced colony numbers compared to "donor only".
TABLE-US-00015 TABLE 5 Comparison of meganuclease-directed and recombinase- directed integration approaches Blasticidin colonies per Sample pD5-D1.3 10.sup.6 cells Percentage Percentage 2 .mu.g each Donor (percentage positive positive per 10.sup.6 .mu.g/10.sup.6 Blasticidin blasticidin in flow in flow cells cells colonies resistant) d 7 d 13 GFP TALEN pair 0.6 90 1800 27.6 55% I-Scel 150 3000 29.9 47% meganuclease pOG44 0.6 60 1200 2.79 6.5% pCAGGS-Cre 0.6 56 1120 2.95 6.6% cCAGGS-Flp.sub.o 0.6 4 80 5.9 Too (low few cell nos) cells Control 40 800 3.3 4.9% (PCDNA3.0) (SBI AAVS 251 (x2) 10040 (1%) ND ND into WT HEKs
[0369] After transfection the bulk of cells were selected for blasticidin resistance in liquid culture and after 7 and 13 days were stained with an anti-Fc phycoerythrin labelled antibody as described above. FIG. 17 (summarised in Table 5) shows significantly higher antibody expression was achieved for cells transfected with I-SceI endonuclease and eGFP TALEN (47% and 55% respectively) compared to "donor only" (4.9%). In contrast the percentage of antibody positive cells when cells were co-transfected with plasmids encoding Flp recombinase (pOG44) or Cre recombinase were 6.6 and 6.5% respectively. The proportion of antibody positive cells continues to increase with continued selection in blasticidin and achieves 85-90% antibody positive in the case of the I-SceI and EGFP TALEN transfected samples when assayed on day 19. Thus meganucleases provide an alternative approach to effect nuclease-directed integration of antibody-encoding transgenes.
Example 12. Nuclease-Directed Integration of an Antibody Cassette can Occur by Both Homologous Recombination and NHEJ
[0370] The efficiency of integration of transgenes into cellular DNA can be enhanced by the introduction of double stranded breaks (DSBs). Endogenous DNA repair mechanisms in eukaryotic cells include homologous recombination non-homologous end joining (NHEJ) and variants of these. All provide a means for introducing genes encoding binders within a library. Homologous recombination provides a precise join between the regions of homology and the inserted transgene but require the provision of regions of homology in the donor plasmid. DNA for homologous recombination can be provided as linear or circular DNA. With NHEJ the ends of DNA are directly re-ligated without the need for a homologous template. This approach to DNA repair is less accurate and can lead to insertions or deletions. NHEJ nonetheless provides a simple means of integrating in-frame exons into intron or allows integration of promoter:gene cassettes into the genome. Use of non-homologous methods allows the use of donor vectors which lack homology arms thereby simplifying the construction of donor DNA.
[0371] Clone 6F has GFP TALEN and I-SceI nuclease recognition sites integrated into the genome and these will be cleaved when these nucleases are provided. The donor vector pD5 does not have a GFP TALE nuclease recognition site but has homology arms flanking the cleavage site and so is expected to integrate by homologous recombination only. Cleavage of genomic DNA at the neighbouring I-SceI meganuclease will also lead to integration of the pD5 elements by homologous recombination. pD5 however also has an I-SceI meganuclease site which can be cleaved in vivo when I-SceI is provided. This will create a linear DNA product which can potentially be integrated by NHEJ. As described earlier there may even be efficiency advantages using in vivo cleavage of donor DNA when NHEJ is used.
[0372] FIG. 18a represents the incoming pD5-D1.3 donor DNA and FIG. 18b represents the genomic locus of clone 6F cells incorporating the "multiple landing" site. FIG. 18c represents the consequence of homologous recombination between pD5-D1.3 (FIG. 18a) and the multiple landing site of clone 6F (FIG. 18b). FIG. 18d in contrast represents the consequence of NHEJ. In this case extra DNA from the backbone of the incoming plasmid is incorporated (represented by a double arrow). Flp mediated recombination at the "multiple landing" site will lead to a similar product. In order to determine which route is being used with the samples described in Example 11 (shown in FIG. 17) genomic DNA was prepared from the blasticidin selected population as described before. A reverse PCR primer (J44) was designed which hybridizes to the integrated PGK promoter. This was used in conjunction with either J48 which hybridises at the end of the IgG protein. Primers J44 and J48 were designed to reveal homologous recombination producing a band of 1928 bp when I-SceI is responsible for integration (indicated by arrow in FIG. 18e). (Potentially a band of 5131 bp could be produced by this primer pair when NHEJ has occurred but this longer product was not visible in the genomic PCRs of this experiment.)
[0373] Primer J46 was designed to hybridise within the .beta.-lactamase gene within the vector backbone. Primers J44 and J46 are anticipated to produce a band of 1800 bp when NHEJ has occurred. A similar sized band is expected where Flp recombinase has led to recombinase-mediated integration.
TABLE-US-00016 J44: (SEQ ID NO: 92) AAAAGCGCCTCCCCTACCCGGTAGAAT J46: (SEQ ID NO: 93) GGCGACACGGAAATGTTGAATACTCAT J48: (SEQ ID NO: 94) CACTACACCCAGAAGTCCCTGAGCCTG
[0374] FIG. 18e clearly reveals that homologous recombination occurs only with the samples treated with GFP TALEN and I-SceI meganuclease ((i and ii compared with iii and iv). In contrast NHEJ only occurs when cleavage is effected by I-SceI meganuclease (FIG. 18e v.) but not GFP TALEN (FIG. 18e vi). As expected a similar size band is found in the sample treated with Flp recombinase (FIG. 18e vii). Thus this experiment reveals nuclease-directed integration of an antibody cassette can occur by both homologous recombination and NHEJ.
Example 13. Generation of Secreted and Membrane Bound Antibody Fragments from the Same Cell
[0375] As described above, mammalian display vectors pD2 and pD5 were constructed with an exon encoding a transmembrane domain flanked by two ROX recognition sites recognised by Dre recombinase [88]. In order to determine whether it was possible to convert from a membrane bound form to a secreted form, the blasticidin resistant population arising from transfection with pD2-D1.3/AAVS TALEN pair was re-transfected with the plasmid encoding Dre recombinase (pCAGGs-Dre). This was based on the plasmid pCAGGs-Dre-IRES puro [88] which drives the Dre recombinase gene from a CAGGs promoter (GeneBridges A205). The puromycin resistance gene was removed using standard molecular biology techniques. After 22 days of blasticidin selection, cells were set up at 0.5.times.10.sup.6 cells/ml and transfected as described earlier with 0.5 .mu.g pCAGGs-Dre per 10.sup.6 cells. After 6 days supernatants were collected, antibody purified using protein A and samples run on an SDS-PAGE gel and stained with Coomassie blue. FIG. 19a shows that secreted antibody was found in the supernatant even without transfection with the Dre recombinase gene. This may arise from alternative splicing where the exon encoding the transmembrane domain is skipped. Alternatively antibody in the culture supernatant could arise from cleavage of the membrane bound antibody. Transfection of Dre recombinase increased the level of secreted antibody (FIG. 19a).
[0376] Production of secreted scFv-Fc fusion was also demonstrated in the experiment describe in example 7 (FIG. 9h). Antibody scFv populations selected by 1 round of phage display on 3-galactosidase were introduced into the pD6 vector and integrate into the AAVS locus of HEK293 cells using the AAVS TALEN. Antigen binding cells were sorted by flow sorting and selected cells were grown for 7 days post-sorting without a change of medium to allow antibody to accumulate. ELISA plates were coated with either .beta.-galactosidase (10 ug/ml) or BSA (10 ug/ml) overnight. Culture supernatants from 7 day cultures were mixed with a 50% volume of 6% Marvel-PBS and the sample tested in triplicate. A 1/10 dilution was also tested. Detection of bound scFv-Fc fusion was performed using anti-Human IgG-Eu (Perkin Elmer Cat 1244-330). FIG. 19b shows that antibody binding can be detected directly from culture supernatants either neat or with a 1/10 dilution. This illustrates that both surface display and antibody secretion can be achieved within the same cells without additional steps. It will be possible to derive secreted antibody directly from the selected cells either following single cell cloning or using a sorted population as shown here generate a polyclonal antibody mix.
Example 14. A Simple Method for Conversion of scFvs to IgG or Fab Format
[0377] A novel method was invented to effect the conversion of antibodies formatted as scFv to an IgG format as described in Example 7. This conversion is a necessary process during antibody drug discovery projects employing scFv antibody phage display libraries where an IgG or Fab formatted antibody is required as the final format. Current methods are tedious and involve individual cloning of the variable heavy (V.sub.H) and variable light (V.sub.L) chains into suitable expression vectors. Furthermore conversion of a population of scFvs "en masse" is not possible because the link between the V.sub.H and V.sub.L chains is lost. This is a problem because both the V.sub.H and V.sub.L chains contribute to antigen binding specificity. The current inability to easily convert populations of scFv to Ig or Fab format limits the ability to screen large numbers of antibodies in the final format they will be used in the clinic. The ability to screen recombinant antibodies in Ig or Fab format for target binding, cell reporter screens and biophysical properties and function including aggregation state is a necessary step to choose candidate antibody drugs as clinical candidates. The greater number of antibodies tested at this stage, in IgG or Fab format, the greater the chance of selecting the best antibody drug candidate.
[0378] Described here is a method to convert single chain antibody (scFv) populations to immunoglobulin (Ig) or Fab format in such a way that the original variable heavy (V.sub.H) and variable light (V.sub.L) chain pairings are maintained. The method allows one to convert monoclonal, oligoclonal or polyclonal scFvs simultaneously to Ig or Fab format. Preferably, the method proceeds via the generation of a non-replicative "mini-circle" DNA. Preferably the complete conversion process entails a single transformation of bacteria such as E. coli to generate a population of bacterial colonies each harbouring a plasmid encoding a unique Ig or Fab formatted recombinant antibody. This is distinct from alternative methods requiring two separate cloning and transformation steps [117].
[0379] More broadly this aspect of the invention relates to a method of converting a genetic construct with 3 linked genetic elements A, B and C (represented by the V.sub.H, linker and V.sub.L respectively in the case of an scFv) to a format where the order of the flanking elements (A and C) are reversed, in a single cloning step. The intermediate element could be retained but most usefully the method permits the replacement of this intermediate element by a new element D (to give C-D-A). In the example of conversion of a scFv to an IgG or Fab then C is an antibody V.sub.L domain and A is a V.sub.H domain. In this example element D encapsulate a light chain constant domain, poly A site, promoter and leader sequence fused to the V.sub.H (element A). In the process the product (C-D-A) is re-cloned allowing the flanking sequences to also be changed. In the scFv to IgG conversion example the V.sub.L element is preceded by a promoter and leader sequence and the V.sub.H is followed by a C.sub.H1-C.sub.H2-C.sub.H3 domain in the case of IgG formatted antibodies and by a CH1 domain in the case of Fab formatted antibodies. The method could be applied more broadly where elements A and C (using above nomenclature) could represent other genetic elements, e.g., in construction of proteins with circularly permutation where the original N and C termini are fused and novel internal termini are engineered.
[0380] FIG. 21 illustrates the conversion process schematically using scFv to IgG conversion as an example. A DNA insert (a) encoding the antibody VH and VL domains is ligated with DNA fragment (b) encoding a constant light (CL) chain, a polyadenylation sequence (pA), a cytomegalovirus (CMV) promoter and a signal peptide (SigP). DNA fragment (b) could also encode any promoter in place of the CMV promoter. Also the pA-CMV cassette could be replaced by an internal ribosomal entry sites (IRES) [119] or a 2A type "self-cleaving" small peptide [130, 133]. The joining of DNA molecules (a) and (b) to create a non-replicative DNA "mini-circle" (c) is facilitated by a "sticky-end" ligation. In FIG. 21, Nco1 and NotI sites are employed because these were used in the creation of the McCafferty phage display library [7] however any suitable restriction sites could be used to create the non-replicative "mini-circle" c. After ligation, the "mini-circle" c is linearized with restriction enzymes NheI and XhoI, the recognition sites of which flank the linker between the V.sub.H and V.sub.L domains. NheI and XhoI were chosen to illustrate this invention because they were used in the creation of the McCafferty phage display library [7], however any suitable restriction sites could be used.
[0381] Linearised product d is then purified and ligated with the digested vector (e). The vector (e) includes a CMV or pEF promoter and signal sequence upstream of the NheI site and encodes the antibody constant heavy (C.sub.H) domains 1 to 3 downstream of the XhoI site. The vector would also encode a bacterial origin or replication and antibiotic resistance marker (not shown) to enable selection and replication of the resultant plasmid DNA in bacteria. The product of ligation of insert (d) with vector (e) would result in plasmid f, which can be used to transform bacteria and growth with a suitable selectable marker would allow the production and purification of plasmid DNA by standard methods. Purified plasmid f can be introduced into mammalian cells [134] for heterologous Ig antibody expression. Alternatively DNA encoding CH.sub.1-3 in vector (e), could be replaced with DNA encoding a single C.sub.H1 domain for Fab expression.
[0382] In the detailed method description below used to illustrate this invention, the insert b contains either a CMV promoter or a P2A peptide which enables expression of the separate antibody light and heavy chains from a single messenger RNA (mRNA). The method is non-obvious and was refined after several experimental attempts. For example, initially linearisation of the DNA "mini-circle" (c) was attempted by PCR. However this resulted in the amplification of homo-dimer side-products, resulting in a low yield of the desired product (d). In contrast, direct digestion of the DNA "mini-circle" (c) provided sufficient material (d) to allow the method to be successfully implemented. Secondly, in an attempt to prevent undesired homo-dimer product, insert (a) was initially dephosphorylated. However, this required careful control to prevent "end" digestion resulting in product lacking the desired "sticky-ends" for ligation. The optimal method does not include dephosphorylation to maximise the proportion of ligation competent product. Lastly, careful control of the ratios used in the ligation of DNA inserts (a) and (b) was required to maximise the yield of the DNA "mini-circle" (c).
1. Preparation of PCR scFv Inserts From a bacterial glycerol stock, harbouring plasmid DNA encoding scFv scrape into 50 ul of water. Dilute this 1 in 10. Use 5 ul of this for PCR reaction containing forward primer pSANG10pelB (CGCTGCCCAGCCGGCCATGG SEQ ID NO: 95) (2.5 .mu.l, 5 M), reverse primer 2097 (GATGGTGATGATGATGTGCGGATGCG SEQ ID NO: 96), (2.5 .mu.l, 5 M), 10.times.KOD buffer (KOD hot start kit from Merck, 71086-4), dNTPs (5 .mu.l, 2 mM), MgSO4 (2 .mu.l, 25 mM), KOD hot start polymerase (2.5 units) in a total volume of 50 .mu.l. Cycling conditions were 94.degree. C. for 2 min then 25 cycles of 94.degree. C. 30 sec, 54.degree. C. for 30 sec then 72.degree. C. for 1 min. PCR clean up was performed by spin column (Qiagen or Fermentas) and the PCR reactions eluted in 90 .mu.l. FIG. 22a shows 1 .mu.l of PCR reaction loaded on a 1% agarose TBE gel. Purified scFv DNA (80 .mu.l, 8 .mu.g) was digested by the addition of buffer 4 (New England Biolabs), BSA (0.1 mg/ml) and 40 units of NcoI-HF and NotI-HF in a total volume of 100 .mu.l and incubated for 2 hours at 37.degree. C. Inserts were purified with a Qiagen PCR cleanup kit, eluted in 30 .mu.l and the DNA concentration measured by measuring the absorbance at 260 nM with a nanodrop spectrophotometer (Thermo).
2. Ligation of DNA Inserts (FIGS. 21a and b)
[0383] A ligation reaction is performed to produce the DNA "mini-circle" (FIG. 21c). The ligation reaction contains insert b (125 ng), scFv insert a (FIG. 21) (125 ng), 10.times. ligation buffer (Roche T4 DNA ligase kit, 1.5 ul), T4 DNA ligase (1 unit) in a total volume of 15 .mu.l. Incubate 1-2 hr 21.degree. C. Water (35 .mu.l) was added to the ligation mix and purified with a Qiagen PCR cleanup kit and eluted in 30 .mu.l.
3. Digestion of DNA "Mini-Circle" (FIG. 21c) with Xho1/Nhe1
[0384] Purified ligation reaction (28 .mu.l) is digested by the addition of buffer 4 (New England Biolabs, 3.5 .mu.l), BSA (0.1 mg/ml) and 10 units of NcoI-HF and NotI-HF in a total volume of 35 .mu.l and incubated for 2 hours at 37.degree. C. This is then purified by separation on a 1% agarose TBE (FIG. 22b). Alternatively FIG. 22c shows a linearised "mini-circle" containing a P2A sequence in place of a CMV promoter. DNA band at 2.6 kb (FIG. 22b) is excised and purified with Qiagen gel extraction kit and eluted in 30 .mu.l.
4. Ligation of Linearised DNA "Mini-Circle" d with pINT3 (XhoI/NheI Cut) Vector and Transformation of E. coli DH5a.
[0385] A standard ligation was set-up with pINT3 cut vector (50 ng), linearised "mini-circle" d (20 ng), 10.times. ligase buffer (Roche, 1.5 .mu.l) and 1 unit of T4 DNA ligase (NEB) in a final volume of 15 .mu.l. Incubation was at 21.degree. C. for 2 hrs. Transformation of E. coli DH5alpha chemically competent cells, subcloning efficiency, (Invitrogen, cat. 18265017) was according to the manufacturer's instructions. 80 .mu.l of chemically competent DH5a cells were added to 6 .mu.l ligation mix, placed on ice for 1 hour and heat shocked at 42.degree. C. 1 min, ice for 2 min and then transferred to a 14 ml polypropylene tube containing 900 .mu.l SOC media and incubated at 37.degree. C. for 1 hour and plated on LB amp plates.
Example 15: Construction of Large Display Libraries in Mammalian Cells by Nuclease-Directed Integration Using Flow Electroporation
[0386] Electroporation is an efficient way of introducing DNA, RNA and protein into cells and electroporation flow systems allow for efficient introduction of DNA into large numbers of mammalian cells. For example the "MaxCyte STX Scalable Transfection System" (Maxcyte) permits the electroporation of 10.sup.10 cells within 30 mins, creating the potential for transfecting up to 10.sup.11 cells in a day. Cells and DNA are mixed and passed from a reservoir to an electroporation chamber, electroporated, pumped out and the process repeated with a fresh aliquot of cells and DNA. The same method can be applied for introduction of DNA, RNA, protein or mixtures thereof into cultured cells (e.g., human HEK293 cells or Jurkat cells) or primary cells e.g. human lymphocytes [135]. Flow electroporation has been used to efficiently introduce DNA, RNA and protein into a large number of primary and cultured cells.
[0387] Here we exemplify the use of such a system to introduce donor DNA, encoding antibody genes, by co-transfecting with DNA encoding a pair of TALE nucleases targetted to the human AAVS locus of human HEK293 cells and Jurkat cells.
[0388] The distribution of the 2 different antibody specificities was determined by flow cytometry using fluorescently-labelled antigen. The generation of antibodies recognising the FGF receptors FGFR1 or FGFR2 has been described previously [105]. Clones .alpha.-FGFR1_A and .alpha.-FGFR2_A (described therein) were cloned into pD6 as described in example 6. In addition a population of scFv antibodies selected from the "McCafferty" phage display library [7] using one round of phage display on .beta.-galactosidase (.beta.-gal) were also cloned into this vector (as described in example 6).
[0389] HEK293 cells were centrifuged and re-suspended in a final volume of 10.sup.8 cells/ml in the manufacturer's electroporation buffer (Maxcyte Electroporation buffer, Thermo Fisher Scientific Cat. No. NC0856428)). An aliquot of 4.times.10.sup.7 cells (0.4 ml) was added to the electroporation cuvette with 100 .mu.g DNA (i.e., 2.5 .mu.g/10.sup.6 cells). The amounts of the different components used are shown below. Donor DNA encoding antibodies .alpha.-FGFR1_A and .alpha.-FGFR2_A was provided as an equimolar mix with the total amount per 10.sup.6 cells shown in Table 6 below. DNA encoding AAVS-SBI TALENs (pZT-AAVS1 L1 and pZT-AAVS R1 Systems Bioscience Cat. No. GE601A-1) was used as an equimolar mix with the total amount per 10.sup.6 cells shown in Table 6 below. In samples without added TALENs, the input DNA was brought to 2.5 .mu.g/10.sup.6 cells using control plasmid pcDNA3.0.
[0390] The percentage transfection efficiency was calculated by counting the number of blasticidin colonies achieved for a given input of total cells. The fold difference compared to negative controls (i.e., no TALEN DNA added) is shown in brackets. Finally, the number of transformed colonies achievable by running a full cycle of the Maxcyte system, involving electroporation of 10.sup.10 cells is calculated in the last column. This represents a single cycle of approximately 30 minutes, giving the potential to run multiple cycles in a day. Thus, the daily output could be 5-10-fold higher. Large scale fermentation and culture systems such as Wavebag system (GE Healthcare) or the Celltainer system (Celltainer Biotech) can be used to generate cells for transfection and can be used to cultivate the resulting libraries.
TABLE-US-00017 TABLE 6 Electroporation of HEK293 cells .alpha.FGFR1_A/ .alpha.FGFR2_A ng TALEN ng donor DNA/10.sup.6 No clones per Sample DNA/10.sup.6 cells cells % transfection 10.sup.10 cells 1 580 1920 5.1 (51x) 5.1 .times. 10.sup.8 1b 580 640 3.3 (33x) 3.3 .times. 10.sup.8 2 580 -- 0.1 -- 0.1 .times. 10.sup.8 3 194 1920 2.7 (89x) 2.7 .times. 10.sup.8 4 194 -- 0.03 0.03 .times. 10.sup.8 5 1185 1315 5.8 (25x) 5.8 .times. 10.sup.8 6 1185 -- 0.23 0.23 .times. 10.sup.8 7 1825 675 6.1 (11x) 6.1 .times. 10.sup.8 8 1825 -- 0.57 0.57 .times. 10.sup.8 9 580 (FGFR1 1920 5.3 5.3 .times. 10.sup.8 alone) 10 580 (FGFR2 1920 5.3 5.3 .times. 10.sup.8 alone) .beta.-galactosidase ng TALEN donor DNA/ DNA/10.sup.6 No clones per Sample 10.sup.6 cells cells % transfection 10.sup.10 cells 11 580 1920 4.5 4.5 .times. 10.sup.8 12 580 -- 0.21 0.21 .times. 10.sup.8 13 1185 1315 5.5 5.5 .times. 10.sup.8 14 1185 -- 0.21 0.21 .times. 10.sup.8
[0391] This example demonstrates that it is possible to make very large libraries of cells with integrated antibody cassettes. The transfection efficiency ranged from 2.7 to 6.1%. In the case of the .beta.-galactosidase selected population (sample 13) a library of 5.5.times.10.sup.8 clones can be created in a single flow electroporation session. With more than one session in a day, a library of 2-5.times.10.sup.9 clones can be generated.
[0392] After 13 days of blasticidin selection (10 .mu.g/ml), cells were labelled with phycoerythrin labelled anti-Fc antibody (Biolegend, Cat. No. 409304) as described earlier. Of the antibody population selected on .beta.-galactosidase, 34-36% of cells were positive for Fc expression and 11-13% were positive for binding of Dyelight-633-labelled antigen at 10 nM concentration.
[0393] Where FGFR binding clones were used, 98-99% of cells were positive for Fc expression. Use of a mixture of .alpha.-FGFR1_A and .alpha.-FGFR2_A antibodies affords an opportunity to examine the proportion of cells containing multiple integration events. For an individual cell with a correctly-integrated cassette (e.g., .alpha.-FGFR1_A) there is approximately a 50:50 chance that a second integration will be of the alternative specificity (i.e., .alpha.-FGFR2_A). If there are frequent multiple integrations, then the proportion of double-positive clones will be high, however, the proportion of double-positive clones was not found to be high, illustrating the fidelity of the nuclease-directed library integration system in generating one antibody gene/per cell. The ability of the surface displayed anti-FGFR antibodies to specifically bind their appropriate antigen was confirmed. Expression of antigen was from a plasmid pTT3DestrCD4(d3+4)-His10 [134] encoding mouse Fgfrl ectodomain (ENSMUSP00000063808). This was used to transfect HEK293 suspension cells and secreted Fgfrl-rCd4-His10 purified by immobilised metal affinity chromatography as described previously [134]. Mouse Fgfr2 ectodomain was PCR amplified from IMAGE clone 9088089 using primers:
TABLE-US-00018 2423 (SEQ ID NO: 97) (TTTTTTCCATGGGCCGGCCCTCCTTCAGTTTAGTTGAG) and 2437 (SEQ ID NO: 98) (TTTTTTGCGGCCGCGGAAGCCGTGATCTCCTTCTCTCTC),
digested with Nco1iNotI and cloned into expression plasmid pBIOCAM5 [126]. Fgfr2-Fc was expressed by transient transfection of HEK293 cells as described previously [134] and purified by affinity chromatography.
[0394] Transfected populations were probed for dual binding using both of the labelled antigens and the proportion of double positives was low. In this experiment the optimal balance of library size (2.7.times.10.sup.8 clones/per Maxcyte session) with low percentage of double positives (3.5%) was found using 197 ng donor DNA per 10.sup.6 cells (FIG. 23).
[0395] This proportion of double positives could represent mis-incorporation of a second antibody cassette but given the efficiency of nuclease-directed integration of the library it is also possible that both alleles (the AAVS locus in this example) could be targeted with incoming binders in a proportion of cells. The presence of two different antibody genes within a cell in itself does not prevent the isolation of binders or their encoding genes but this can be circumvented by first modifying the target cell at a single locus to introduce a single nuclease targeting site, e.g., a pre-integrated SceI meganuclease site as demonstrated in example 9.
Example 16. Recovery of Genes Encoding Binders from Sorted Library Populations
[0396] Phage display selections were carried on .beta.-galactosidase (as described in example 6) and antibody populations from 1-2 rounds of selection were cloned into vector pD6 and introduced into the AAVS locus of HEK293 cells as described in example 6. .beta.-galactosidase was labelled using Lightning Link Dyelight-633 (Innova Bioscience Cat. No 325-0010) according to manufacturer's instructions. Transfected cell populations were selected for 25 days in blasticidin (10 .mu.g/ml) and were labelled with 10 nM Dyelight-633 labelled .beta.-galactosidase and phycoerythrin-labelled anti-Fc (Biolegend Cat. No. 409304). Cells were incubated with antibodies for 30 minutes at 4.degree. C., washed twice in PBS/0.1% BSA, re-suspended in PBS/0.1% BSA and double-positive cells sorted using a flow sorter.
[0397] Sorted cells were expanded and a second round of sorting was carried out using 10 nM antigen. Cells were grown and either genomic DNA or mRNA was isolated from the sorted, selected populations.
[0398] Where binders encompassing different chains (e.g., IgG formatted antibodies) are present on the same genomic sequence (e.g., by introduction on the same plasmid) but are transcribed into different mRNAs it may be optimal to recover the separate genes encoding the multimeric binder by amplification from genomic DNA. As an alternative, binders encompassing multiple protein chains can be encoded on the same mRNA through the use of "internal ribosome entry sequence" (IRES) elements or sequences such as viral P2A or T2A sequences that promote translational stalling/protein cleavage [133, 136]. In this case and in the case of binders encoded on a single protein chain it will also be possible to isolate the encoded genes from mRNA.
[0399] Genomic DNA was prepared using the "DNeasy blood and tissue kit" (QIAGEN Cat. No. 69504). mRNA was prepared using an "Isolate II RNA mini kit" (Bioline Bio-52072). For amplification from genomic DNA a PCR reaction was set up using Phusion polymerase with the "2.times.Phusion GC" mix according to manufacturer's instructions. Primers which flank the Nco1 and Not1 cloning sites, e.g., primers 2622 (GAACAGGAACACGGAAGGTC) (SEQ ID NO: 99) and 2623 (TAAAGTAGGCGGTCTTGAGACG) (SEQ ID NO: 82) were used to amplify the antibody cassette (98.degree. C. 10 secs, 58.degree. C. 20 secs, 72.degree. C. 90 secs for 35 cycles.
[0400] Genes encoding selected scFv genes were amplified from mRNA. Total RNA was isolated from the sorted cells using the "Isolate II RNA mini kit" (Bioline Cat No Bio-52072). cDNA was synthesized from 2 g RNA using Superscript II reverse transcriptase (Life Technologies, Cat no 180064-022). The selected scFv genes were then amplified from the cDNA by PCR using KOD Hot Start DNA polymerase (Merck Millipore Cat. No. 71086-3) using primers which flank the Nco1 and Not1 cloning sites. In this case, primers:
TABLE-US-00019 41679 (SEQ ID NO: 100) ATGAGTTGGAGCTGTATCATCC and 2621 (SEQ ID NO: 101) GCATTCCACGGCGGCCGC
were used to amplify the antibody cassette (95.degree. C. for 20 seconds, 60.degree. C. for 10 seconds, 70.degree. C. for 15 seconds, for 25 cycles). PCR products were digested with Nco1 and Not 1 before cloning into the Nco/Not1 site of bacterial antibody expression vector pSANG10. Construction of pSANG10, methods for bacterial expression and screening by ELISA are described in Martin et al 2006 [127].
[0401] ELISA screening of the population from 1 round of selection by phage display revealed that 0/90 of the clones were positive. In contrast, when this same population was further subjected to mammalian display and the scFv gene population was recovered and screened, 27/90 clones (30%) were positive in ELISA. This illustrates that it is possible to carry out mammalian display on a library and recover an enriched population of binders.
[0402] Example 15 describes the introduction of the population selected by 1 round of phage display on .beta.-galactosidase into HEK cells using flow electroporation. The mammalian display cell population was selected in blasticidin as before and was subjected to flow sorting using 10 nM of labelled .beta.-galactosidase. After 9 days growth 75% of cells were found by flow cytometry to be positive for .beta.-galactosidase binding using 10 nM .beta.-galactosidase). These were sorted and expanded further. To illustrate the potential to drive stringency, labelling was carried out using either 1 nM or 10 nM antigen concentrations. 20.3% and 55.9% of cells respectively were sorted from each population. After sorting, mRNA was prepared immediately from the sorted population without additional cell culture. Following cloning, expression in bacteria and ELISA screening (as before) it was found that the success rate in ELISA increased with increasing stringency during flow sorting. The clones displaying highest signal level came from this group and the number of positives clones was also improved (FIG. 24). This illustrates the ability to drive stringency of selection within display populations, reflected in the better performance of the resulting antibodies.
Example 17. Incorporation of T Cell Receptors (TCRs) Genes for Mammalian Library Production by Nuclease-Directed Integration
[0403] The methods described herein have application beyond display of antibodies. To demonstrate the potential for screening libraries of T-cell receptors using nuclease-directed integration, a vector construct (pINT20) allowing expression of T-cell receptors was constructed.
[0404] pINT20 (FIG. 25a) is a dual promoter vector for targeting the human AAVS locus. It has left and right homology arms as presented in FIG. 3. The left AAVS homology arm is flanked by unique AsiSI and Nsi1 sites and is followed by a splice acceptor and a puromycin gene. The sequence between the end of the left homology arm and the splice acceptor is the same as previously described and the puromycin gene commences with an ATG in frame with the upstream exon ((FIG. 25B and as also shown for the blasticidin gene in FIG. 3). Correct nuclease-directed integration will lead to in-frame splicing of a puromycin gene which is spliced to an endogenous upstream exon which itself driven by the endogenous AAVS promoter allowing selection of clones with correct integration. The puromycin resistance gene is followed by an SV40 polyadenylation site. The right AAVS homology arm is flanked by unique BstZ171 and Sbf1 sites.
[0405] pINT20 is configured with a pEF promoter (from pSF-pEF, Oxford Genetics Cat. No. OG43), which allows genes of interest to be cloned into Nhe1/Kpn1 sites. The Nhe1 site is preceded by a secretion leader sequence and the Kpn1 site is followed by the polyadenylation signal of bovine growth hormone (bGH poly A) as shown previously in FIG. 2. Downstream is a CMV promoter (from pSF-CMV-f1-Pac1, Oxford Genetics Cat No OG111) allowing cloning via Nco and Not (as shown in FIG. 2) or Hind3. The Nco1 site is preceded by a secretion leader sequence and the cassette is followed by a bGH polyA site.
[0406] To exemplify display and enrichment of T cell receptors (TCRs) a TCR recognising a cancer marker described by Li et al. (2005) and later by Zhao et al. (2007)[137, 138] was used. This TCR called c12c2 recognises the peptide SLLMWITQV (SE ID NO: 102) presented on HLA-A2 with an affinity of 450 nM. This peptide represents residues 157-165 from NY-ESO-1 (NY-ESO-1 157-165). This is an affinity-matured variant of a parental antibody called 1G4 with affinity of 32 .mu.M.
[0407] A second TCR was used which recognises another cancer marker. The parental MEL5 TCR recognises the peptide MART-1 26-35 ("Melanoma antigen recognised by T cells-1") presented on HLA-A2 with peptide sequence ELAGIGILTV (SEQ ID NO: 103). This TCR was affinity matured by phage display to give clone a24/017 with 0.5 nM affinity described in Madura et al. (2013) [139]. The structure of the complex between TCR and MHC:peptide complex has been solved (pdb code 4JFH) and the clone is hereafter named as "4JFH". The same parental TCR has also been engineered based on design by Pierce et al. (2014) [140] and the structure of the complex solved.
[0408] According to Debets and colleagues the attachment of the CD3 domain as shown tends to cause association of the heterologous gene even in the presence of native human TCRs [141, 142]. The CD3 .xi. element used is composed of an extracellular domain, a transmembrane domain and a complete cytoplasmic domain. In addition substitution of human constant domains by mouse constant domains in the heterologous genes also tends to drive their association over association with endogenous human constant domains [143]. Finally the use of mouse constant domains offers the option of detecting the heterologous TCR chains against a background of human TCRs. These elements were incorporated into the design of the TCR expression cassette.
[0409] Two synthetic genes were designed and synthesised giving rise to gene constructs with the following structure:
Human TCR V.alpha.-mouse .alpha. constant-human CD3 .xi. Human TCR V.beta.-mouse .beta. constant-human CD3 .xi.
[0410] The sequence of the synthetic gene incorporating the .alpha. chain and the .beta. chain constructs incorporating the variable domains of c12/c2 is shown in FIGS. 25c and d. These are cloned into the Nhe1/Kpn1 sites and the Nco1/Hind3 sites of pINT20 respectively. The construct encoding this TCR is called pINT20-c12/c2. In the first instance the synthetic gene was designed to incorporate a V.alpha. C.alpha. domain (flanked by Nhe1 and Not 1 sites) and V.beta. domain (flanked by Nco1/Xho1 sites encoding the TCR c12/c2). These elements can then be replaced by alternative TCRs using these restriction sites.
[0411] Two additional synthetic genes were made encoding the V.alpha. and V .beta. domains of 4JFH [139] (FIGS. 25e and f). The construct encoding this TCR is called pINT20-4JFH.
[0412] 10.sup.7 HEK293 cells were transfected using 3 .mu.g of pINT20-c12/c2 and pINT20-4JHF as donor DNA (300 ng donor DNA per 10.sup.6 cells) in the ratios shown in Table 7. pINT20-c12/c2 is referred to as TCR1 and pINT20-4JHF is referred to as TCR2. 5 g each of pZT-AAVS1 L1 and pZT-AAVS R1 TALENs were added to 10.sup.7 cells (500 ng each per 10.sup.6 cells) with the exception of sample 6 where this was replaced by 10 .mu.g of control DNA (pcDNA3.0). DNA was introduced by polyethyleneimine transfection, as described above.
TABLE-US-00020 TABLE 7 Sample TCR1/TCR2 ratio 1 1:100 2 100:1 3 50:50 4 100% TCR1 5 100% TCR2 6 50:50 (no TALE nuclease)
[0413] Following 12 days in selection (0.25 .mu.g/ml puromycin) cells were labelled with target antigen. The peptide:MHC complexes recognised by the TCRs described above are presented as a phycoerythrin labelled pentamer (ProImmune). c12/c2 recognises peptide SLLMWITQV presented on HLA-A2 representing NY-ESO-1 157-165 (Proimmune product code 390). 4JHF (also known as .alpha.24/.beta.17) recognises peptide ELAGIGILTV presented on HLA-A2 representing MelanA/MART 26-35 (Proimmune product code 082). In each case the MHC:peptide complex was labelled with phycoerythrin and used according to manufacturer's instructions. FIG. 26 (a-d) shows that each TCR is specific for the expected MHC: peptide complex (a, d) and fails to bind to the non-cognate peptide in complex (FIG. 26b, c). DNA encoding each TCR was mixed with a 100-fold excess of DNA encoding the other (samples 1-2, Table 7). HEK293 cells were transfected and selected in puromycin. Sorting of antigen positive cells was performed after 14 days of puromycin selection (FIG. 26g, h).
[0414] In order to quantitate the level of enrichment within the flow-sorted output populations, the TCR genes were recovered by PCR amplification and the relative amounts of each TCR species determined following cloning. Total RNA was isolated from the sorted population. cDNA synthesis was performed as described in example 16. Primers to amplify the TCR alpha and beta chain were 1999/2782 and 41679/2789 respectively (Table 8). PCR amplification employed KOD hot start polymerase using the manufacturer's recommended protocol (EMD Millipore, 71086, EMD Millipore). PCR reaction conditions were 95.degree. C. (2 mins) and 25 cycles of 95.degree. C. (20 s), 60.degree. C. (10 s), 70.degree. C. (15 s) followed by 70.degree. C. (5 mins). The amplified TCR alpha and beta chains were digested with NheI/NotI or NcoI/XhoI and sub-cloned into vectors with compatible sites (in this case pBIOCAM1-Tr-N NheI/NotI or pBIOCAM2-Tr-N(NcoI/XhoI) cut vectors respectively). PCR of individual clones were amplified with a c2c12 (TCR1) specific TCR alpha primer (2781) or a 4JFH (TCR2) specific TCR alpha primers (4JFH-V.alpha.-F) and vector specific primers to assay for TCR clone identity. After sorting of the sample 1 (Table 7) where a ratio of 1:100 TCR1/TCR2 donor plasmid was employed (Table 7) with enrichment for TCR1 specific clones using peptide SLLMWITQV presented on HLA-A2 (Proimmune, product code 390), the proportion of TCR1 clones, as determined by colony PCR, increased to 11/15 (73%). Enrichment by sorting the sample 2 where a ratio of 100:1 TCR1/TCR2 donor plasmid was employed (Table 7), with peptide ELAGIGILTV presented on HLA-A2 (Proimmune product code 082) resulted in an increase of the proportion of TCR2 clones to 4/15 (27%), determined by colony PCR.
[0415] To demonstrate library selection using nuclease-directed integration, a mutant library based on c12/c2 was created by cloning a repertoire of genes encoding mutant TCR alpha chains along with a repertoire of genes encoding mutant TCR beta chains. Such a library could be created using oligonucleotide-directed mutagenesis approaches. e.g., methods based on Kunkel mutagenesis [144]. As an alternative and by way of example, a PCR assembly approach was used to create a mutant TCR alpha chain (as a Nhe1/Kpn1 fragment) and a mutant TCR beta chain (as a Kpn/Hind 3 fragment, including the CMV promoter) which is cloned into the Nhe1/Hind 3 site of pINT20. This was done using primers shown in Table 8.
TABLE-US-00021 TABLE 8 Primers used in library construction and clone recovery 4JFH-V.alpha.-F ACACACGCTAGCCAGAAAGAGGTGGAACAG (SEQ ID NO: 104) 1999 AAAAAGCAGGCTACCATGAGGGCCTGGATC (SEQ ID NO: 87) TTCTTTCTCC 2770 CAAAGAACAGCTCGCCGGTSNNCCCGASSN (SEQ ID NO: 105) NGGAGCTGGCGCAAAAGTAC 2771 CTCGCCCGAAGGTGGGAATGTANGWTCCSN (SEQ ID NO: 106) NSNNAAGTGGGCGCACGGCGCAC 2780 CTGGCAGCTAGCAAGCAGGAAG (SEQ ID NO: 107) 2781 TACATTCCCACCTTCGGGCGAG (SEQ ID NO: 108) 2782 TTTTTTGCGGCCGCGGACAGGTTCTG (SEQ ID NO: 109) 2783 CGTAAGCTGGTACCTTATTATCTAGGG (SEQ ID NO: 110) 2785 CCCTAGATAATAAGGTACCAGCTTACG (SEQ ID NO: 111) 2787 ACCGGCGAGCTGTTCTTTG (SEQ ID NO: 112) 2788 AGTGACAAGCTTTTATTATCTGGGTG (SEQ ID NO: 113) 2789 CAGGTCCTCGAGCACTGTC (SEQ ID NO: 114) 41679 ATGAGTTGGAGCTGTATCATCC (SEQ ID NO: 100) (N = A, C, G, T. S = C OR G, W = A OR T)
[0416] A mutant oligonucleotide was designed (primer 2771) which randomised 2 amino acid positions within CDR3 of the c12/c2 alpha chain and provide the option of either serine or threonine at another position (primer 2771 is also represented by the lower strand of FIG. 25g). Primer 2771 was used in conjunction with primer 2780 to create mutant TCR alpha repertoire going from the Nhe1 cloning site incorporating the region of CDR3 mutagenesis with an invariant sequence at the end. Primer 2781 is complementary to the invariant 5' end of primer 2771. PCR with primers 2781 and 2783 provided the remainder of the TCR alpha-CD3 zeta cassette up to the Kpn1 site. PCR assembly of the 2 PCR fragments is used to create the TCR alpha-CD3 zeta fragment which can be cloned into pINT20 following digestion with Nhe1 and Kpn1.
[0417] A second mutant oligonucleotide (primer 2770) was designed which randomised 2 amino acid positions within CDR3 of the c12/c2 beta chain and provide the option of either valine or leucine at another position (primer 2770 represented by the lower strand of FIG. 25h). Primer 2770 was used in conjunction with primer 2785 to create a mutant TCR beta repertoire from the Kpnl cloning site incorporating the region of CDR3 mutagenesis with an invariant sequence at the end. Primer 2787 is complementary to the invariant 5' end of primer 2700. PCR with primers 2787 and 2788 provided the remainder of the TCR beta-CD3 zeta cassette up to the Hind 3 site. PCR assembly of these 2 PCR fragments is used to create a TCR beta-CD3 zeta fragment which can be cloned into pINT20. A complete repertoire incorporating mutations at both CDR 3 of alpha and beta chains was created by cloning of the Nhe1/Kpn fragment, the Kpn1/Hind3 fragment into Nhe1/Hind3 digested pINT20. Following ligation the library was cloned into electrocompetent DH10B cells. Plasmid DNA was prepared and the DNA transfected into HEK293 cells along with vectors encoding TALE nuclease as described earlier (an equimolar mix of pZT-AAVS1 L1 and pZT-AAVS R1 Systems Bioscience Cat. No. GE601A-1).
[0418] Following ligation of the mutant alpha and beta chains of the c12/c2 mutant library into pINT20, DNA was electroporated into DH10B cell, plasmid DNA was prepared and the library was co-transfected with TALE nuclease targeting the AAVS locus. Transfection was performed using Maxcyte electroporation. Growth and selection were as described above. Quantitation of the library size, by titration of transfected cells and plate-based selection in puromycin, indicated that a library size 5.times.10.sup.5 was created. After puromycin selection for 11 days cells were labelled with an APC-labelled antibody specific to the .beta. chain of the mouse TCR (Life Technologies Cat H57-957). FIG. 26i-j shows 38% of clones in the population express a T cell receptor. Of this TCR positive population, 13% also bound peptidel (5% of the total population). By this approach clones with improved expression or peptide:MHC binding activity can be isolated.
[0419] From FIG. 26 it can be seen that each T cell receptor recognised only its cognate antigen. Furthermore when a mixture of the two different specificities are used it is possible to distinguish each of them through the labelled antigen. This approach also allows TCR clones with improved affinity (or expression) to be distinguished within the mutant TCR library by identifying a subset of the library that was labelled to a greater extent than the parental clone.
[0420] T cell receptor genes were also introduced into Jurkat cells by electroporation. Jurkat cells were centrifuged and re-suspended in a final volume of 10.sup.8 cells/ml in the manufacturer's electroporation buffer (Maxcyte Electroporation buffer, Thermo Fisher Scientific Cat. no NC0856428)). An aliquot of 4.times.10.sup.7 cells (0.4 ml) was added to the OC400 electroporation cuvette with 40 .mu.g DNA (i.e., 1 .mu.g/10.sup.6 cells). DNA consisted of a mixture of donor plasmid DNA (pINT20-c12/c2 or pINT20-4JHF or pINT20-c12/c2 TCR library, 9.2 .mu.g) and an equimolar mix of DNA (30.8 .mu.g total) of DNA encoding the AAVS-SBI TALENs (pZT-AAVS1 L1 and pZT-AAVS R1 Systems Bioscience Cat No GE601A-1). In samples without added TALENs the input DNA was brought to 1 jg/10.sup.6 cells using control plasmid pcDNA3.0. An alternative method of introducing T cell receptor genes into Jurkat cells used the 4D-Nucleofector (Lonza). Here, the transfection protocol followed the manufacturer's instructions according to the SE cell-line kit (Lonza, Cat. PBC1-02250). Briefly, 2 .mu.g of DNA, consisting of a mixture of donor plasmid DNA (pINT20-c12/c2 or pINT20-4JHF or pINT20-c12/c2 TCR library, 0.46 .mu.g) and an equimolar mix of DNA (1.54 .mu.g total) of DNA encoding the AAVS-SBI TALENs (pZT-AAVS1 L1 and pZT-AAVS R1 Systems Bioscience Cat No GE601A-1) was transfected per 10.sup.6 Jurkat cells. The pulse code setting was C.sub.L120 and cell type programme was specific for Jurkat E6.1 (ATCC) cells.
[0421] FIG. 26 demonstrates that TCR expression was achieved and recognition of the appropriate peptide:MHC molecule was achieved. This was dependent on the use of TALE nuclease (compare FIGS. 26m and n). Signalling through the introduced TCR was also achieved using the relevant peptide:MHC molecule.
[0422] pINT20-c12/c2 transfected Jurkat cells or untransfected Jurkat cells were plated in a 96-well plate at a density of 1.times.10.sup.6/ml, 200 .mu.l per well. Cells were stimulated with either 2 .mu.l or 6 .mu.l per well of PE labelled peptide 1-MHC pentamer (ProImmune) or 2 .mu.g/ml anti-human CD3 (BD Pharmingen, Cat 555329) in the presence and absence of anti-human CD28 (BD Pharmingen Cat 555725) at 2 .mu.g/ml. After a 24 hour incubation at 37.degree. C. and 5% CO.sub.2, the activation of Jurkat cells was detected by investigating CD69 expression. Cells were stained with 50l PBS+1% BSA+0.5 .mu.l of anti-human CD69-APC (Invitrogen, Cat. MHCD6905) per well for 45 minutes at 4.degree. C.
[0423] FIG. 26 (sample o and p) demonstrates up-regulation of CD69 upon stimulation with CD3. The figure also shows the effect of adding 2 .mu.l (q) or 6 .mu.l (r) of peptidel:MHC. A population of double-positive cells which bind peptide:MHC and express CD69 is obvious. This example shows cells incubated in the presence of CD28 but the same effect was observed in the absence of CD28 (not shown).
[0424] This example demonstrates the potential of nuclease-directed integration of libraries of an alternative type of binder, i.e., T cell receptors. We demonstrate that it is possible to express and detect TCR expression on the cell surface using specific antibodies. We also demonstrate that these T cell receptors specifically recognise their respective targets. We have also constructed a mutant library allowing selection of improved binders. Finally we have demonstrated that library screening based on activation of TCR signalling in T cells is possible. Here we have used a cultured human T cell line. It is also possible to introduce DNA into primary T cells by Maxcyte electroporation. Methods for the isolation and preparation of primary T lymphocytes are known to those skilled in the art (e.g., Cribbs et al., 2013, Oelke et al. 2003 [145, 146]. Exposure of TCR transfected lymphocytes to multimeric peptide:MHC can then be used to achieve activation either through exposure to peptide:MHC multimers [146] or to antigen presenting cells loaded with the appropriate peptide [146, 147]. Activation can be detected either through expression of reporter genes or through up-regulation of endogenous genes such as CD69 [104, 148].
Example 18. Display of Libraries of Chimeric Antigen Receptors on Mammalian Cells
[0425] Activation of T cells normally occurs through interaction of the T Cell Receptor (TCR) with specific peptide:MHC complexes. This in turn leads to signalling directed via CD3 and other T cell signalling molecules. As an alternative to target recognition directed by the TCR it has been shown that alternative binding molecules such as single chain Fvs can be presented on T cells as fusions to downstream signalling molecules in a way that re-directs T cell activation to the molecule recognised by the scFv (or alternative binder). In this way T cell activation is no longer limited to the molecular recognition directed towards peptide:MHC complexes by TCRs but can be directed to other cell surface molecules. This alternative format wherein a non-TCR binding entity is fused to a signalling component is referred to as a "chimeric antigen receptor" (CAR). In the case of T cells this has been shown to be an important and valuable means of re-directing T cell activation.
[0426] For any given target it is still not clear what the optimal epitope or affinity features should be for incorporation into a CAR [103]. Features of the CAR design such as linker length, or choice of transmembrane domain may in turn affect what constitutes an optimal epitope. The combination of antigen density on target and non-target cells together with the choice of signalling domain could affect the optimal affinity requirements. The ability to present libraries of chimeric antigen receptors on T cells affords an opportunity to identify optimal binding specificity, binder format, linker length/sequence, variants of fused signalling module, etc., either alone or in combination. Here we demonstrate the utility of nuclease-directed integration for construction of libraries of chimeric antigen receptors in mammalian cells. The vector pINT21 (FIG. 27a) is a single CMV promoter vector for convenient expression and secretion of binders such as scFvs flanked by Nco1/Not1 restriction sites to allow in frame expression with an upstream leader sequence and a downstream fusion partner (as shown earlier in FIG. 8). The CAR expression cassette in pINT21 is flanked by AAVS homology arms as described earlier in FIG. 3.
[0427] The vector pINT21-CAR1 (FIG. 27a, c) fuses binders such as single chain Fvs to the transmembrane domain and intracellular domain of CD3.xi. (FIG. 27c and as described for TCR expression in FIG. 25). This format is often referred to as a "first generation" chimeric antigen receptor. Signalling domains from other co-stimulatory molecules have also been used to provide additional signals and these have been shown to give improved signalling. These have been referred to as second and third generation chimeric antigen receptors. For example pINT21-CAR2 (FIG. 27b, d)fuses the binder (conveniently cloned in this case as an Nco1/Not 1 fragment to a previously described second generation domain (WO 2012/079000 A1) consisting of:
The hinge and transmembrane domain from CD8 4-1BB signalling domain CD3.xi. signalling domain
[0428] By way of example a number of different binder groups were cloned into the Nco/Not1 sites of pINT20-CAR1 and pINT20-CAR2. CD19 has previously been used in a number of different studies to target B-cell malignancies references in Sadelain et al. (2013) [103]. A previously described anti-CD19 antibody (WO 2012/079000 A1) (called FMC63) was prepared as a synthetic scFv gene in either a VH-linker-VL configuration or a VL-linker-VH configuration (FMC63 H-L or FMC L-H respectively, FIG. 27E shows the sequence of FMC63 H-L. FMC63 L-H was configured with the variable domains in VL-linker-VH configuration flanked by Nco1 and Not1 at 5' and 3' ends respectively.
[0429] As controls, scFvs with alternative binding specificities were also cloned into pINT20. These include anti-FGFR1_A [105] and an anti-desmin control antibody [7]. In addition, Adhirons [152] recognising lox1 (FIG. 29a, b) were introduced as an example of an alternative format of binder configured as a CAR fusion (see example 19 for description).
[0430] To demonstrate the creation of libraries of binders presented in a CAR format, populations of scFv-formatted antibodies selected on mesothelin and CD229 were cloned. Mesothelin is a cell surface glycoprotein which is highly expressed in a number of cancers including mesothelioma. A number of antibody-based formats are under development and in clinical trial including CARs directed to mesothelin [149]. CD229 represents another potential tumour-associated antigen which could be targeted by immune therapy in cancers such as chronic lymphocytic leukaemia and multiple myeloma [150, 151].
[0431] A population of antibodies recognising either mesothelin or CD229 was created by selection using the "McCafferty" phage display library as described in example 6 and ref 7). Two rounds of selection were carried out and the scFv-encoding genes recovered using primers M13leadseq and Notmycseq (example 6). Products were cut with Nco1/Not 1, gel purified and cloned into pINT21-CAR2. These were directed into the AAVS locus of HEK293 cells by TALE nuclease cleavage to generate a library of 4.8.times.10.sup.5 for CD229 and 6.4.times.10.sup.5 for mesothelin (representing a 30.times. and 53.times. increase in library compared to samples transfected in the absence of TALE nuclease).
[0432] pINT20-CAR1 and pINT20-CAR2 were introduced into HEK293 cells by PEI transfection. Here donor plasmid DNA (pINT20-CAR1 or pINT20-CAR2, 6 .mu.g was mixed with an equimolar mix of DNA (20 .mu.g total) of DNA encoding the AAVS-SBI TALENs (pZT-AAVS1 L1 and pZT-AAVS R1 Systems Bioscience Cat No GE601A-1) in Freestyle 293 media (Lifetech, Cat. 12338-026), linear PEI (52 al, 1 mg/ml, Polysciences Inc.) added and incubated at room temperature for 10 mins. The mixture is then added to 20 ml HEK293 suspension cells (1.times.10.sup.6 cell/ml) in a 125 ml vented Erlenmeyer flask. pINT20-CAR1 and pINT20-CAR2 were also introduced into Jurkat cells by electroporation. Jurkat cells were centrifuged and re-suspended in a final volume of 10.sup.8 cells/ml in the manufacturer's electroporation buffer (Maxcyte Electroporation buffer, Thermo Fisher Scientific Cat. no NC0856428)). An aliquot of 4.times.10.sup.7 cells (0.4 ml) was added to the OC400 electroporation cuvette with 40 .mu.g DNA (i.e., 1 .mu.g/10.sup.6 cells). DNA consisted of a mixture of donor plasmid DNA (pINT20-CAR1 and pINT20-CAR2, 9.2 .mu.g) and an equimolar mix of DNA (30.8 jg total) of DNA encoding the AAVS-SBI TALENs (pZT-AAVS1 L1 and pZT-AAVS R1 Systems Bioscience Cat No GE601A-1). In samples without added TALENs, the input DNA was brought to 1 .mu.g/10.sup.6 cells using control plasmid pcDNA3.0.
[0433] Fluorescent labelling of the various antigens was performed using Lightning-Link Rapid Dye-Light 633 conjugation kit (Innova Biosciences, cat. 325-0010). Preparation of FGFR1 and FGFR2 is described in example 15. Lox1 and CD229 were from R and D Systems (Cat. Nos. 1798-LX-050, and 898-CD050 respectively), CD19-Fc and mesothelin were from (AcroBiosystems Cat. No. CD9-H5259 and. MSN-H526.times. respectively).
[0434] FIG. 28b illustrates display of nuclease-directed anti-FGFR1 antibody [105] within a second generation CAR construct (pINT21-CAR2-FGFR1_A). FIG. 28d also illustrates display of an alternative scaffold molecule (an Adhiron ref [152]) as a fusion with a second generation chimeric antigen receptor (pINT21-CAR2-lox1). FIGS. 28f and g also illustrate positive clones within a library of scFvs selected on mesothelin or CD229.
[0435] In this example CARs were introduced into HEK cells and Jurkat cells but this could equally be done by introducing the constructs into primary cells such as human T lymphocytes (e.g., as described by Sadelain et al. (2013) [103], for example using electroporation [135]. Expression constructs for CAR expression in lymphocytes may be further optimised, e.g. by optimising mRNA stability and translation through variation in 5' and 3'untranslated regions, poly A length etc., as has previously been described [135]. Signalling of CAR constructs introduced into primary T lymphocytes or T lymphocyte cell lines can be induced by exposure to cells expressing target antigen or using multimeric antigen, e.g. antigen immobilised on a surface or presented on beads [104, 148].
Example 19. Display of Libraries of Alternative Scaffolds Constructed in Mammalian Cells Via Nuclease-Directed Integration
[0436] The method described for constructing libraries of binders can be employed beyond display of antibodies and T cell receptors. A number of alternative scaffolds have been described allowing construction of libraries of variants from which novel binding specificities have been isolated, e.g., Tiede et al. (2014) [152] and references therein. In the example described by Tiede et al. (2014) a stable, versatile scaffold based on a consensus sequence from plant-derived phytocystatins was used. This scaffold is referred to as an Adhiron and FIG. 29a shows a synthetic gene encoding an Adhiron which was selected to bind to lox1 (WO 2014125290 A1). FIG. 29B shows an alternative lox1 binder (lox1B). Both were synthesised and cloned into the Nco1/Not1 site of pINT20_CAR2 to create a fusion with the downstream partner.
[0437] A library can be constructed by randomising loop residues (e.g., by Kunkel mutagenesis or PCR assembly as described above in example 17). By way of example, FIG. 29c shows the design of a mutant oligonucleotides useful to create a library following the same approach as described in example 17. In this case, randomisation is achieved by introducing a variable number of NNS residues, although alternative strategies known to those skilled in the art could be used.
[0438] As another example FIGS. 29d and e demonstrates the means to create a library of binders by nuclease-directed integration based on a knottin scaffold [156]. Knottins are peptides of approximately 30 amino acids which are stabilised by three disulphide bonds, with one threaded through the other two to create a "knotted" structure. FIG. 29d shows the trypsin binding knottin MCoTI-II with an Nco1 site at the 5' end and a Not site at the 3' end allowing in-frame expression with the vectors described herein. As an example for library construction the 6 amino acids of the first loop (underlined in FIG. 29d) can be mutated with variable number of amino acids. FIG. 29e illustrates a mutagenic strategy replacing the 6 amino acids of loop 1 with 10 randomised amino acids using the codons VNS (where V=A, C or G and S.dbd.C or G). The VNS codon encompasses 24 codons encoding 17 amino acids which exclude cysteines. This strategy is for illustrative purposes and alternative mutagenic strategies will be known to those skilled in the art.
Example 20. Nuclease-Directed Introduction of Antibody Libraries Using CRISPR/Cas9
[0439] Nuclease-directed integration via CRISPR/Cas9 was demonstrated using the "Geneart CRISPR nuclease vector kit" (Lifetech A21175). In this system, a U6 RNA polymerase III promoter drives expression of a target complementary CRIPSR RNA (crRNA) which is linked to a trans-activating crRNA (tracrRNA). The crRNA and tracrRNA together make up a guide RNA which directs the cleavage specificity of a Cas9 protein encoded on the same "GeneArt CRISPR nuclease vector" (see manufacturer's instructions). The vector is provided as a linearised plasmid into which a short double-stranded oligonucleotide with appropriate 3' overhangs is cloned. Cleavage specificity is then determined by the sequence of the cloned segment. Two different targeting sequences were designed to direct cleavage to the human AAVS locus describe above.
[0440] The sequences were:
TABLE-US-00022 CRISPR1 double-stranded DNA insert: (SEQ ID NO: 115) 5' GGGGCCACTAGGGACAGGATGTTTT (SEQ ID NO: 116) 3' GTGGCCCCCGGTGATCCCTGTCCTAC CRISPR2 double-stranded DNA insert: (SEQ ID NO: 117) 5' GTCACCAATCCTGTCCCTAGGTTTT (SEQ ID NO: 118) 3' GTGGCCAGTGGTTAGGACAGGGATC
[0441] The resulting guide RNAs target cleavage within the same region of the AAVS locus as the TALE nucleases described above (with CRISPR2 being in the reverse orientation from CRISPR1). Thus the AAVS homology arms used previously to direct integration of the expression cassette can be used for integration directed by these CRISPR guide RNAs. Linearised vector and double stranded oligonucleotides were ligated and transformed into electrocompetent DH10B cells. Cloning of the correct insert was confirmed by sequencing and plasmid DNA was prepared. The Cas9/CRISPR2 construct (encompassing the CRISPR2 oligonucleotide) was transfected together with donor DNA encoding a .beta.-galactosidase library selected by 1 round of phage display selection (example 15). These were transfected into HEK293 cells using Maxcyte electroporation system with the OC-400 assembly. 4.times.10.sup.7 cells were transfected with 23.2 .mu.g of donor DNA representing a population of scFv genes selected by one round of phage display on .beta.-galactosidase cloned into pD6. Cells were co-transfected with either 77 .mu.g of Cas9-CRISPR2 plasmid, or 77 .mu.g TALEN plasmid (38.5 .mu.g each of pZT-AAVS1 L1 and pZT-AAVS) or 77 .mu.g control plasmid
TABLE-US-00023 TABLE 9 Transfection Number of clones Sample no Nuclease used efficiency % per 10.sup.10 cells 1 Cas9/CRISPR 2 10.53 (29x) 10 .times. 10.sup.8 2 AAVS TALEN 5.1 (14x) 5.1 .times. 10.sup.8 3 None (pCDNA3.0) 0.36 0.36 .times. 10.sup.8
[0442] Titration of the number of transformants formed by Cas9/CRISPR2 transfection (by measuring blasticidin resistance colonies) revealed that 1053 blasticidin resistant colonies were generated from plating 10,000 cells, equating to a transfection efficiency of 10.5% (Table 9). In the case of TALE nuclease-directed a transfection efficiency of 5.1% was achieved. In contrast, in the absence of the Cas9/CRISPR2 construct only 0.36% transfection efficiency was achieved.
[0443] As an alternative to transfection using plasmid DNA to introduce the Cas9 protein and guide RNA into cells it is also possible to directly introduce a nucleoprotein complex consisting of Cas9 protein (Toolgen, Inc.) and a guide RNA. Guide RNA was prepared by Toolgen, Inc. using in vitro transcription from a T7 promoter as a single transcript which included the TRACR sequence (italicized) preceded by sequence complimentary to the target DNA (in bold) as shown below.
TABLE-US-00024 CRISPR 1 RNA (SEQ ID NO: 119): 5' GGGGGGCCACUAGGGACAGGAUGUUUUAGAGCUAGAAAUAGCAAGUU AAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUG CUUUU CRISPR 2 RNA (SEQ ID NO: 120): 5' GGGUCACCAAUCCUGUCCCUAGGUUUUAGAGCUAGAAAUAGCAAGUU AAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUG CUUUU
[0444] 6.6 .mu.g of Cs9 protein, 4.6 g of RNA and 10 g of donor DNA (encoding the anti-FGFR1_A antibody in pD6) were introduced into 10.sup.7 HEK293 cells by Maxcyte electroporation as described above. Transfection efficiencies of 2.2% and 2.9% were achieved for CRISPR 1 and CRISPR2 RNA respectively with 0.7% and 0.8% in the absence of added Cas9:RNA protein complex.
[0445] These guide RNAs target the same sequences encoded by CRISPR1 and CRISPR2. As an alternative, crRNA and tracrRNA can be made by chemical synthesis (e.g., GE Dharmacon).
Example 21: Nuclease Mediated Antibody Gene Insertion by Ligation or Microhomology-Mediated End-Joining (MMEJ)
[0446] Although homologous recombination (HR) is useful for the precise insertion of large DNA fragments, this requires the construction of large targeting vectors incorporating long homology arms. This can make the construction of large libraries more difficult due to the reduced transformation efficiency of larger DNA constructs. Alternatively, simple ligation reactions can occur between the chromosomal DNA and targeting vector, if a nuclease recognition sequence is incorporated into the targeting vector. The ligation reactions can either be "sticky-end" employing, for example, zinc finger nucleases (Orlando et al., 2010) [45] or TALENs (Cristea et al., 2013) [22] which can make double-strand breaks (DSBs) that leave 5' overhangs or "blunt-end" employing CRISP/Cas9 ribonucleoprotein. An example of nuclease gene integration by ligation using I-SceI meganuclease was shown by the construction of vector pD7-Sce1. pD7 is derived from pD6 (FIG. 8) but the left and right AAVS homology arms were replaced with short double stranded oligonucleotides. The left AAVS homology arm of the pD vector series is flanked by EcoR1 and Nsi1 restriction enzymes (see FIG. 3). To convert pD6 to pD7-Sce1, this was replaced by a double-stranded oligonucleotide insert formed by primers 2778 and 2779 encoding an I-SceI meganuclease recognition sequence with "sticky ends" compatible with the "sticky ends" formed by EcoRI/NsiI. The right hand AAVS homology arm is flanked by Asc1 and Mlu1 restriction sites (FIG. 3). The right homology arm was replaced by a double-stranded oligonucleotide insert with "sticky ends" compatible with the "sticky ends" formed by AscI/MluI digestion and is formed by primers 2723 and 2724.
TABLE-US-00025 TABLE 10 Primers for pD7 and pINT19 construction 2723 CGCGCCAGAAGTCTCACCAAGCCCA (SEQ ID NO: 121) 2724 CGCGTGGGCTTGGTGAGACTTCTGG (SEQ ID NO: 122) 2768 AATTCTCCCCTCCACCCCACAGTAGGGA (SEQ ID NO: 123) CAGTGGGGCCAGGATTGGTGACAGAAAA TGCA 2769 TTTTCTGTCACCAATCCTGGCCCCACTG (SEQ ID NO: 124) TCCCTACTGTGGGGTGGAGGGGAG 2778 AATTCTAGGGATAACAGGGTAATATGCA (SEQ ID NO: 125) 2779 TATTACCCTGTTATCCCTAG (SEQ ID NO: 126) 2808 AATTCTTTTCTGTCACCAATCCTGGGGC (SEQ ID NO: 127) CACTAGGGACACTGTGGGGTGGAGGGGA TGCA 2809 TCCCCTCCACCCCACAGTGTCCCTAGTG (SEQ ID NO: 128) GCCCCAGGATTGGTGACAGAAAAGAATT G
[0447] Antibodies recognising Fgfr1 and Fgfr2 (example 15) were cloned into pD7 to create pD7-SceI anti-Fgfr1 and pD7-SceI anti-Fgfr2 respectively. These were co-transfected with the I-SceI expression plasmid (example 11, FIG. 16) into the HEK293 clone 6F cell line (see example 11) which contains an integrated I-SceI recognition site.
[0448] Ligation of DSBs in the chromosome and targeting vector generated by zinc finger nuclease or TALE nucleases can also be achieved. By inverting the zinc finger nuclease or TALEN recognition sites on the targeting vector this can ensure that the product of the insertion is no longer a target for cleavage in a method termed "Obligate ligation-gated recombination" or ObLiGaRe (Maresca et al., 2013) [153]. pD7-ObLiGaRe vectors can be generated in the same way as described above for the creation of pD7-Scel. In this case, the left hand homology arm is replaced by an oligonucleotide consisting of primers 2808 and 2809 encoding an inverted TALEN recognition site (shown in bold) and spacer region. The right hand homology arm is replaced with primers 2723 and 2809 as described above.
[0449] An alternative to simple ligation reactions between DSBs in the chromosome and targeting vector, mediated by non-homologous end joining (NHEJ) is microhomology-mediated end-joining (MMEJ). MMEJ is a DSB repair mechanism that uses microhomologous sequences between 5 to 25 bp for error-prone end joining (McVey and Lee, 2008) [154]. A strategy for precise gene integration has been devised where the genomic sequence and the targeting vector contain the same TALE nuclease pair recognition sequence, but a different vector spacer sequence in which the anterior and posterior half are switched. The genomic sequence and vector can be cut by the same TALEN pair and MMEJ takes place via the microhomologous DNA ends. The resultant integrated targeting vector is no longer a target for TALE nuclease because of the shortened spacer region which are not optimal for TALE nuclease cleavage (Nakade et al., 2014) [155].
[0450] The MMEJ AAVS targeting vector pD7-MMEJ can be generated in the same way as described above for the creation of pD7-Scel. In this case the left hand homology arm is replaced by an oligonucleotide consisting of primers 2768 and 2769 encoding the TALEN recognition site (shown in bold) and switched spacer region (underlined). The right hand homology arm is replaced with primers 2723 and 2809 as described above.
Example 22: Design of Primers for Creation of Single (CMV) Promoter Cassette Flanked by ROSA26 Arms (pINT19-ROSA)
[0451] This example is intended to demonstrate that antibody or alternative binding molecule genes can be integrated into the genome of mammalian cells by nuclease directed methods and the resultant clones screened for a desired function by either reporter or phenotypic screening. An example of this was previously demonstrated where antibody genes were integrated into the chromosome of mouse embryonic stem (ES) cells and individual ES colonies screened for their ability to maintain pluripotency when subjected to differentiation conditions [105]. Antibody genes recovered from ES colonies which maintained a pluripotent phenotype were shown to block the FGFR1/FGF4 signaling pathway. A problem with this previously reported method is that homologous recombination can result in small library sizes, thus limiting its ability to directly screen for rare clones present in large binding molecule libraries. Nuclease-mediated gene integration methods for antibody and binding molecule gene integration are more efficient resulting in larger library size generation and thus more likely to generate mammalian cell libraries capable of identifying functional antibodies by phenotypic or reporter cell screening.
[0452] The donor targeting vector pINT 19 is designed to integrate antibody genes into the mouse ROSA26 locus by nuclease directed methods for direct functional screening. pINT19 is a single CMV promoter vector for scFv-Fc fusion expression. The expression cassette is flanked by mouse ROSA26 arms. Since the upstream exon is untranslated, the puromycin gene is preceded by a splice acceptor and further down has a KOZAK sequence leading into the puromycin gene.
[0453] The AAVS left homology arm and puromycin resistance gene of pINT 18 was replaced by a cassette encoding the ROSA26 left homology, splice acceptor, optimized kozak consensus sequence and puromycin resistance gene. The ROSA26 left homology arm was initially amplified from pGATOR (Melidoni et al., 2013 (105)) as two fragments which knocked out an internal NotI site. The two fragments, generated by primers J60/2716 and 2715/2706 were combined in an assembly PCR with primers J60 and 2706 and digested with AsiSI and NsiI. The splice acceptor was amplified from pGATOR using primers 2709 and 2710 and the puromycin resistance cassette amplified with primers 2745 (which included a region homologous to the splice acceptor and optimized Kozak consensus) and J59. The splice acceptor region and puromycin resistance cassette were combined in assembly PCR using primers 2709 and J59 and digested with Nsi1 and Bgl2. The ROSA26 left arm homology and splice acceptor-puromycin cassettes were ligated with pINT 18 (AsiS 1/Bgl2) vector.
[0454] To complete the ROSA targeting vector the right hand ROSA26 homology arm, downstream of CMV-scFv-Fc cassette, was introduced to replace the pINT18 AAVS right homology arm. This was performed by PCR of the ROSA right homology arm, present in pGATOR (Melidoni et al., 2013) using primers J61 and J62 to amplify a fragment with BstZ171 at one end and Sbf1 at the other. Primer 61 was positioned to exclude an endogenous Sbf1 site 65 bp up from ROSA ZFN cleavage site. FIG. 31 shows the sequences of ROSA26 left and right homology arms.
TABLE-US-00026 TABLE 11 Primers for nuclease-directed targeting of the mouse ROSA 26 locus J60 ACACACGGTACCGCGATCGCGCTGATT AsiSI-rosa26-L-F (SEQ ID NO: 129) GGCTTCTTTTCCTC 2706 TTTTTTATGCATTCTAGAAAGACTGGA NsiI-rosa26-L-R (SEQ ID NO: 130) GTTGCAGA 2715 GAGCGTCCGCCCACCCTC ROSA-Left- (SEQ ID NO: 131) NotI_knockout_F 2716 GAGGGTGGGCGGACGCTC ROSA-Left- (SEQ ID NO: 132) NotI_knockout_R 2709 TTTTTTATGCATTAAGGGATCTGTAGG Splice_acceptor- (SEQ ID NO: 133) GCGCAG F-NsiI 2710 GTGAATTCCTAGAGCGGCCTC Splice_acceptor- (SEQ ID NO: 134) R 2745 GAGGCCGCTCTAGGAATTCACGCCGCC Overlap-Puro- (SEQ ID NO: 135) ACCATGACCGAGTACAAGCCCAC F+kozak J59 AAAAAAAGATCTGTGTGTTTCGAATCA Bgl2-Puro-R (SEQ ID NO: 136) GGCACCGGGCTTGCGGGTCAT J61 ttttttGTATACGGGAATTGAACAGGT ROSA-Right_F- (SEQ ID NO: 137) GTAAAATTG BstZ171 J62 TTTTTTCCTGCAGGAGGTTGGATTCTC ROSA-Right_R- (SEQ ID NO: 138) AATACATCTATTGTTG SbfI 2701 GCCGACGTCTCGTCGCTGATGTTTT (SEQ ID NO: 139) 2702 ATCAGCGACGAGACGTCGGCCGGTG (SEQ ID NO: 140) 2703 CGCCCATCTTCTAGAAAGACGTTTT (SEQ ID NO: 141) 2704 GTCTTTCTAGAAGATGGGCGCGGTG (SEQ ID NO: 142)
[0455] Integration of pINT19, encoding antibody or alternative binding molecules, into the mouse ROSA26 locus could be achieved by nuclease-directed introduction of antibody libraries using CRISPR/Cas9 as described in Example 20. Here, nuclease-directed integration via CRISPR/Cas9 could be demonstrated using the "Geneart CRISPR nuclease vector kit" (Lifetech A21175). In this system, a U6 RNA polymerase III promoter drives expression of a target complementary CRIPSR RNA (crRNA) which is linked to a trans-activating crRNA (tracrRNA). The crRNA and tracrRNA together make up a guide RNA which directs the cleavage specificity of a Cas9 protein encoded on the same "GeneArt CRISPR nuclease vector" (see manufacturer's instructions). The vector is provided as a linearised plasmid into which a short double-stranded oligonucleotide with appropriate 3' overhangs is cloned. Cleavage specificity is then determined by the sequence of the cloned segment. 2 different targeting sequences were designed to direct cleavage to the mouse ROSA26 encoded by primers 2701/2702 and 2703/2704 (see Table 10).
[0456] As an alternative, Zinc finger nucleases which cleave within the ROSA26 locus have been described [34]. These could be constructed in an appropriate expression vector as described for Sce-I meganuclease (FIG. 16, example 11).
[0457] Nuclease-mediated integration of donor plasmid pINT 19 would give rise to clones expressing secreted antibody which could bind endogenous receptor or ligand resulting in either antagonism [105,107] or agonism [47,106,108] of receptor signalling pathways. To enable the linkage between cellular phenotype and the functional activity of the secreted antibody, cells can be plated at a low density in semi-solid media so that individual colonies propagate and antibody expression can be initiated via an inducible promoter [105]. Alternatively, a constitutive promoter could be employed for antibody gene expression. The semi-solid media would maintain an elevated local concentration of the endogenously-expressed antibody, so that any phenotypic change specific to a colony arising from an individual cell would be caused by the unique antibody expressed from that particular clone. If a rapid response reporter, such as Rex or Nanog promoter fused to a reporter gene, was employed it would be possible to plate cells at a low density in semi-solid media, harvest and then screen by flow cytometry. Alternatively, the stop codon downstream of the antibody gene in pINT 19 could be replaced by a transmembrane domain to enable tethering of the antibody to the cell surface. The stop codon downstream of the antibody gene in pINT19 could also be replaced by an endoplasmic reticulum (ER) retention signal sequence to enable retention of antibodies in the ER and potential down-regulation of an endogenously expressed target receptor or any secreted protein or peptide. pINT19 is designed specifically to target the mouse ROSA26 locus and can be employed for phenotypic screening of antibodies or alternative binding molecules in mouse ES cells. However, nuclease-directed antibody or binder molecule gene integration methods could also be applied to other functional screens such as those described using the lentiviral approach [47,106,107,108].
REFERENCES
[0458] All references listed below and others cited anywhere in this disclosure are incorporated herein by reference in their entirety. [0459] 1 Russell, S. J., Hawkins, R. E., & Winter, G. (1993). Retroviral vectors displaying functional antibody fragments. Nucleic Acids Res, 21(5), 1081-1085. [0460] 2 Boublik, Y., Di Bonito, P., & Jones, I. M. (1995). Eukaryotic virus display: Engineering the major surface glycoprotein of the Autographa californica nuclear polyhedrosis virus (AcNPV) for the presentation of foreign proteins on the virus surface. Nature Biotechnology, 13(10), 1079-1084. [0461] 3 Mottershead, D. G., Alfthan, K., Ojala, K., Takkinen, K., & Oker-Blom, C. (2000). Baculoviral display of functional scFv and synthetic IgG-binding domains. Biochemical and Biophysical Research Communications, 275(1), 84-90. [0462] 4 Oker-Blom, C., Airenne, K. J., & Grabherr, R. (2003). Baculovirus display strategies: Emerging tools for eukaryotic libraries and gene delivery. Briefings in Functional Genomics and Proteomics, 2(3), 244-253. [0463] 5 Edwards, B. M., Barash, S. C., Main, S. H., Choi, G. H., Minter, R., Ullrich, S., et al. (2003). The Remarkable Flexibility of the Human Antibody Repertoire; Isolation of Over One Thousand Different Antibodies to a Single Protein, BLyS. Journal of Molecular Biology, 334(1), 103-118. doi: 10.1016/j.jmb.2003.09.054 [0464] 6 Pershad, K., Pavlovic, J. D., Graslund, S., Nilsson, P., Colwill, K., Karatt-Vellatt, A., et al. (2010). Generating a panel of highly specific antibodies to 20 human SH2 domains by phage display. Protein Engineering, Design and Selection, 23(4), 279-288. doi:10.1093/protein/gzq003 [0465] 7 Schofield, D. J., Pope, A. R., Clementel, V., Buckell, J., Chapple, S., Clarke, K. F., et al. (2007). Application of phage display to high throughput antibody generation and characterization. Genome Biol, 8(11), R254. [0466] 8 Salema, V., Marin, E., Martinez-Arteaga, R., Ruano-Gallego, D., Fraile, S., Margolles, Y., et al. (2013). Selection of Single Domain Antibodies from Immune Libraries Displayed on the Surface of E. coli Cells with Two 13-Domains of Opposite Topologies. PLoS ONE, 8(9), e75126. doi: 10.1371/journal.pone.0075126.s007 [0467] 9 Chao, G., Lau, W. L., Hackel, B. J., Sazinsky, S. L., Lippow, S. M., & Wittrup, K. D. (2006). Isolating and engineering human antibodies using yeast surface display. Nat Protoc, 1(2), 755-768. [0468] 10 Higuchi, K., Araki, T., Matsuzaki, O., Sato, A., Kanno, K., Kitaguchi, N., & I to, H. (1997). Cell display library for gene cloning of variable regions of human antibodies to hepatitis B surface antigen. J Immunol Methods, 202(2), 193-204. [0469] 11 Ho, M., Nagata, S., & Pastan, I. (2006). Isolation of anti-CD22 Fv with high affinity by Fv display on human cells. Proc Natl Acad Sci USA, 103(25), 9637-9642. doi: 10.1073/pnas.0603653103 [0470] 12 Ho, M., & Pastan, I. (2009). Display and selection of scFv antibodies on HEK-293T cells. Methods Mol Biol, 562, 99-113. doi:10.1007/978-1-60327-302-2_8 [0471] 13 Akamatsu, Y., Pakabunto, K., Xu, Z., Zhang, Y., & Tsurushita, N. (2007). Whole IgG surface display on mammalian cells: Application to isolation of neutralizing chicken monoclonal anti-IL-12 antibodies. Journal of Immunological Methods, 327(1-2), 40-52. doi: 10.1016/j.jim.2007.07.007 [0472] 14 Beerli, R. R., Bauer, M., Buser, R. B., Gwerder, M., Muntwiler, S., Maurer, P., et al. Isolation of human monoclonal antibodies by mammalian cell display. Proc Natl Acad Sci USA. 2008 Sep. 23; 105(38):14336-41. doi: 10.1073/pnas.0805942105 [0473] 15 Breous-Nystrom, E., Schultze, K., Meier, M., Flueck, L., Holzer, C., Boll, M., et al. (2013). Retrocyte DisplayA.RTM. technology: Generation and screening of a high diversity cellular antibody library. Methods, 1-11. doi: 10.1016/j.ymeth.2013.09.003 [0474] 16 Grindley, Whiteson & Rice. Mechanisms of Site-Specific Recombination. Annu Rev Biochem 2006 75:567-605 [0475] 17 Zhou, C., Jacobsen, F. W., Cai, L., Chen, Q., & Shen, W. D. (2010). Development of a novel mammalian cell surface antibody display platform. mAbs, 2(5), 508-518 [0476] 18 Li, C. Z., Liang, Z. K., Chen, Z. R., Lou, H. B., Zhou, Y., Zhang, Z. H., et al. (2012). Identification of HBsAg-specific antibodies from a mammalian cell displayed full-length human antibody library of healthy immunized donor. Cellular and Molecular Immunology, 9(2), 184-190. [0477] 19 Buchholz, F., Ringrose, L., Angrand, P. O., Rossi, F., & Stewart, A. F. (1996). Different thermostabilities of FLP and Cre recombinases: Implications for applied site-specific recombination. Nucleic Acids Res, 24(21), 4256-4262. [0478] 20 Schaft, J., Ashery-Padan, R., Van Hoeven, F. D., Gruss, P., & Francis Stewart, A. (2001). Efficient FLP recombination in mouse ES cells and oocytes. Genesis, 31(1), 6-10. [0479] 21 Moehle, E. A., Moehle, E. A., Rock, J. M., Rock, J. M., Lee, Y.-L., Lee, Y. L., et al. (2007). Targeted gene addition into a specified location in the human genome using designed zinc finger nucleases. Proc Natl Acad Sci USA, 104(9), 3055-3060. doi:10.1073/pnas.0611478104 [0480] 22 Cristea, S., Freyvert, Y., Santiago, Y., Holmes, M. C., Urnov, F. D., Gregory, P. D., & Cost, G. J. (2013). In vivo cleavage of transgene donors promotes nuclease-mediated targeted integration. Biotechnology and Bioengineering, 110(3), 871-880. doi: 10.1002/bit.24733 [0481] 23 Letourneur, F., & Malissen, B. (1989). Derivation of a T cell hybridoma variant deprived of functional T cell receptor alpha and beta chain transcripts reveals a nonfunctional alpha-mRNA of BW5147 origin. European Journal of Immunology, 19(12), 2269-2274. doi:10.1002/eji. 1830191214 [0482] 24 Kanayama, N., Todo, K., Reth, M., & Ohmori, H. (2005). Reversible switching of immunoglobulin hypermutation machinery in a chicken B cell line. Biochem Biophys Res Commun, 327(1), 70-75. doi:10.1016/j.bbrc.2004.11.143 [0483] 25 Lin, W., Kurosawa, K., Murayama, A., Kagaya, E., & Ohta, K. (2011). B-cell display-based one-step method to generate chimeric human IgG monoclonal antibodies. Nucleic Acids Research, 39(3), e14-e14. doi:10.1093/nar/gkq1122 [0484] 26 Adachi, N., So, S., Iiizumi, S., Nomura, Y., Murai, K., Yamakawa, C., et al. (2006). The human pre-B cell line Nalm-6 is highly proficient in gene targeting by homologous recombination. DNA and Cell Biology, 25(1), 19-24. doi:10.1089/dna.2006.25.19 [0485] 27 Palacios, R., & Steinmetz, M. (1985). IL3-Dependent mouse clones that express B-220 surface antigen, contain Ig genes in germ-line configuration, and generate B lymphocytes in vivo. Cell, 41(3), 727-734 [0486] 28 Barretina, J., Caponigro, G., Stransky, N., Venkatesan, K., Margolin, A. A., Kim, S., et al. (2012). The Cancer Cell Line Encyclopedia enables predictive modelling of anticancer drug sensitivity. Nature, 483(7391), 603-607. doi:10.1038/nature 11003 [0487] 29 Forbes, S. A., Bindal, N., Bamford, S., Cole, C., Kok, C. Y., Beare, D., et al. (2011). COSMIC: mining complete cancer genomes in the Catalogue of Somatic Mutations in Cancer. Nucleic Acids Research, 39(Database issue), D945-50. doi:10.1093/nar/gkq929 [0488] 30 Silva, G., Poirot, L., Galetto, R., Smith, J., Montoya, G., Duchateau, P., & Paques, F. (2011). Meganucleases and other tools for targeted genome engineering: Perspectives and challenges for gene therapy. Current Gene Therapy, 11(1), 11-27 [0489] 31 Epinat, J. C., Silva, G. H., Paques, F., Smith, J., & Duchateau, P. (2013). Engineered meganucleases for genome engineering purposes. Topics in Current Genetics (Vol. 23, pp. 147-185). [0490] 32 Szczepek, M., Brondani, V., Bichel, J., Serrano, L., Segal, D. J., & Cathomen, T. (2007). Structure-based redesign of the dimerization interface reduces the toxicity of zinc-finger nucleases. Nature Biotechnology, 25(7), 786-793. doi: 10.1038/nbt1317 [0491] 33 Doyon, Y., Vo, T. D., Mendel, M. C., Greenberg, S. G., Wang, J., Xia, D. F., et al. (2011). Enhancing zinc-finger-nuclease activity with improved obligate heterodimeric architectures. Nat Methods, 8(1), 74-79. doi: 10.1038/nmeth. 1539 [0492] 34 Perez-Pinera, P., Ousterout, D. G., Brown, M. T., & Gersbach, C. A. (2012). Gene targeting to the ROSA26 locus directed by engineered zinc finger nucleases. Nucleic Acids Research, 40(8), 3741-3752. doi:10.1093/nar/gkr1214 [0493] 35 Urnov, F. D., Miller, J. C., Lee, Y.-L., Beausejour, C. M., Rock, J. M., Augustus, S., et al. (2005). Highly efficient endogenous human gene correction using designed zinc-finger nucleases. Nature, 435(7042), 646-651. doi:10.1038/nature03556 [0494] 36 Maresca, M., Lin, V. G., Guo, N., & Yang, Y. (2013). Obligate ligation-gated recombination (ObLiGaRe): custom-designed nuclease-mediated targeted integration through nonhomologous end joining. Genome Research, 23(3), 539-546. doi:10.1101/gr.145441.112 [0495] 37 Bogdanove, A. J., & Voytas, D. F. (2011). TAL effectors: customizable proteins for DNA targeting. Science, 333(6051), 1843-1846. doi:10.1126/science. 1204094 [0496] 38 Reyon, D., Tsai, S. Q., Khgayter, C., Foden, J. A., Sander, J. D., & Joung, J. K. (2012). FLASH assembly of TALENs for high-throughput genome editing. Nature Biotechnology, 30(5), 460-465. doi:10.1038/nbt.2170 [0497] 39 Boissel, S., Jarjour, J., Astrakhan, A., Adey, A., Gouble, A., Duchateau, P., et al. (2013). megaTALs: a rare-cleaving nuclease architecture for therapeutic genome engineering. Nucleic Acids Research. doi:10.1093/nar/gkt1224 [0498] 40 Beurdeley, M., Bietz, F., Li, J., Thomas, S., Stoddard, T., Juillerat, A., et al. (2013). Compact designer TALENs for efficient genome engineering. Nature Communications, 4, 1762. doi: 10.1038/ncomms2782 [0499] 41 Sampson, T. R., & Weiss, D. S. (2014). Exploiting CRISPR/Cas systems for biotechnology. BioEssays: News and Reviews in Molecular, Cellular and Developmental Biology, 36(1), 34-38. doi: 10.1002/bies.201300135 [0500] 42 Shalem, O., Sanjana, N. E., Hartenian, E., Shi, X., Scott, D. A., Mikkelsen, T. S., et al. (2014). Genome-scale CRISPR-Cas9 knockout screening in human cells. Science, 343(6166), 84-87. doi: 10.1126/science. 1247005 [0501] 43 Wang, T., Wei, J. J., Sabatini, D. M., & Lander, E. S. (2014). Genetic screens in human cells using the CRISPR-Cas9 system. Science, 343(6166), 80-84. doi:10.1126/science. 1246981 [0502] 44 Beard, C., Hochedlinger, K., Plath, K., Wutz, A., & Jaenisch, R. (2006). Efficient method to generate single-copy transgenic mice by site-specific integration in embryonic stem cells. Genesis, 44(1), 23-28. [0503] 45 Orlando, S. J., Santiago, Y., DeKelver, R. C., Freyvert, Y., Boydston, E. A., Moehle, E. A., et al. (2010). Zinc-finger nuclease-driven targeted integration into mammalian genomes using donors with limited chromosomal homology. Nucleic Acids Research, 38(15), e152. doi:10.1093/nar/gkq512 [0504] 46 Cadinanos, J; Bradley, A. (2007) Nucleic Acids Res. 35(12):e87 [0505] 47 Zhang, H., Yea, K., Xie, J., Ruiz, D., Wilson, I. A., & Lerner, R. A. (2013). Selecting agonists from single cells infected with combinatorial antibody libraries. Chemistry & Biology, 20(5), 734-741. doi:10.1016/j.chembiol.2013.04.012 [0506] 48 Porteus, M. H., & Baltimore, D. (2003). Chimeric nucleases stimulate gene targeting in human cells. Science, 300(5620), 763. doi:10.1126/science. 1078395 [0507] 49 Rouet, P., Smih, F., & Jasin, M. (1994). Introduction of double-strand breaks into the genome of mouse cells by expression of a rare-cutting endonuclease. Molecular and Cellular Biology, 14(12), 8096-8106. [0508] 50 Jasin, M. (1996). Genetic manipulation of genomes with rare-cutting endonucleases. Trends in Genetics, 12(6), 224-228 [0509] 51 Davis, L., & Maizels, N. (2011). DNA nicks promote efficient and safe targeted gene correction. PLoS ONE, 6(9), e23981. doi:10.1371/journal.pone.0023981 [0510] 52 Fujioka, K., Aratani, Y., Kusano, K., & Koyama, H. (1993). Targeted recombination with single-stranded DNA vectors in mammalian cells. Nucleic Acids Res, 21(3), 407-412 [0511] 53 Khan, I. F., Hirata, R. K., & Russell, D. W. (2011). AAV-mediated gene targeting methods for human cells. Nat Protoc, 6(4), 482-501. doi: 10.1038/nprot.2011.301 [0512] 54 Deyle, D. R., & Russell, D. W. (2009). Adeno-associated virus vector integration. Current Opinion in Molecular Therapeutics, 11(4), 442-447 [0513] 55 Benatuil, L., Perez, J. M., Belk, J., & Hsieh, C. M. (2010). An improved yeast transformation method for the generation of very large human antibody libraries. Protein Engineering, Design and Selection, 23(4), 155-159. [0514] 56 Feldhaus, M. J., Siegel, R. W., Opresko, L. K., Coleman, J. R., Feldhaus, J. M., Yeung, Y. A., et al. (2003). Flow-cytometric isolation of human antibodies from a nonimmune Saccharomyces cerevisiae surface display library. Nat Biotechnol. [0515] 57 Zhao, A., Nunez-Cruz, S., Li, C., Coukos, G., Siegel, D. L., & Scholler, N. (2011). Rapid isolation of high-affinity human antibodies against the tumor vascular marker Endosialin/TEM1, using a paired yeast-display/secretory scFv library platform. Journal of Immunological Methods, 363(2), 221-232. doi:10.1016/j.jim.2010.09.001 [0516] 58 Skerra, A. (2007). Alternative non-antibody scaffolds for molecular recognition. Curr Opin Biotechnol, 18(4), 295-304. doi:10.1016/j.copbio.2007.04.010 [0517] 59 Gebauer, M., & Skerra, A. (2009). Engineered protein scaffolds as next-generation antibody therapeutics. Current Opinion in Chemical Biology, 13(3), 245-255. doi: 10.1016/j.cbpa.2009.04.627 [0518] 60 Haan & Maggos (2004) BioCentury, 12(5): A1-A6 [0519] 61 Koide et al. (1998) Journal of Molecular Biology, 284: 1141-1151. [0520] 62 Nygren et al. (1997) Current Opinion in Structural Biology, 7: 463-469 [0521] 63 Wess, L. In: BioCentury, The Bernstein Report on BioBusiness, 12(42), A1-A7, 2004 [0522] 64 Chang, H.-J., Hsu, H.-J., Chang, C.-F., Peng, H.-P., Sun, Y.-K., Yu, H.-M., et al. (2009). Molecular Evolution of Cystine-Stabilized Miniproteins as Stable Proteinaceous Binders. Structure, 17(4), 620-631. doi: 10.1016/j.str.2009.01.011 [0523] 65 Ward, E. S. et al., Nature 341, 544-546 (1989) [0524] 66 McCafferty et al Nature, 348, 552-554 (1990) [0525] 67 Holt et al Trends in Biotechnology 21, 484-490 (2003) [0526] 68 Bird et al, Science, 242, 423-426, (1988) [0527] 69 Huston et al, PNAS USA, 85, 5879-5883, (1988) [0528] 70 Holliger, P. et al, PNAS USA 90 6444-6448, (1993) [0529] 71 Reiter, Y. et al, Nature Biotech, 14, 1239-1245, (1996) [0530] 72 Holliger & Hudson, Nature Biotechnology 23(9): 1126-1136 (2005) [0531] 73 Knappik et al. J. Mol. Biol. 296, 57-86 (2000) [0532] 74 Krebs et al. Journal of Immunological Methods 254, 67-84 (2001) [0533] 75 Holliger and Bohlen Cancer and metastasis rev. 18: 411-419 (1999) [0534] 76 Holliger, P. and Winter G. Current Opinion Biotechnol 4, 446-449 (1993) [0535] 77 Glennie M J et al., J. Immunol. 139, 2367-2375 (1987) [0536] 78 Repp R. et al., J. Hemat. 377-382 (1995) [0537] 79 Staerz U. D. and Bevan M. J. PNAS 83 (1986) [0538] 80 Suresh M. R. et al.,
Method Enzymol. 121: 210-228 (1986) [0539] 81 Merchand et al., Nature Biotech. 16:677-681 (1998) [0540] 82 Ridgeway, J. B. B. et al, Protein Eng., 9, 616-621, (1996) [0541] 83 Marks, J. D., Hoogenboom, H. R., Bonnert, T. P., McCafferty, J., Griffiths, A. D., & Winter, G. (1991). By-passing immunization. Human antibodies from V-gene libraries displayed on phage. J Mol Biol, 222(3), 581-597. [0542] 84 Gronwald, R. G. K., Grant, F. J., Haldeman, B. A., Hart, C. E., O'Hara, P. J., Hagen, F. S., et al. (1988). Cloning and expression of a cDNA coding for the human platelet-derived growth factor receptor: Evidence for more than one receptor class (Vol. 85, pp. 3435-3439). Presented at the Proceedings of the National Academy of Sciences of the United States of America. [0543] 85 Kumar, N., & Borth, N. (2012). Flow-cytometry and cell sorting: an efficient approach to investigate productivity and cell physiology in mammalian cell factories. Methods, 56(3), 366-374. doi:10.1016/j.ymeth.2012.03.004 [0544] 86 Brezinsky, S. C. G., Chiang, G. G., Szilvasi, A., Mohan, S., Shapiro, R. I., MacLean, A., et al. (2003). A simple method for enriching populations of transfected CHO cells for cells of higher specific productivity. J Immunol Methods, 277(1-2), 141-155. [0545] 87 Pichler, J., Hesse, F., Wieser, M., Kunert, R., Galosy, S. S., Mott, J. E., & Borth, N. (2009). A study on the temperature dependency and time course of the cold capture antibody secretion assay. Journal of Biotechnology, 141(1-2), 80-83. doi:10.1016/j.jbiotec.2009.03.001 [0546] 88 Anastassiadis, K., Fu, J., Patsch, C., Hu, S., Weidlich, S., Duerschke, K., et al. (2009). Dre recombinase, like Cre, is a highly efficient site-specific recombinase in E. coli, mammalian cells and mice. Disease Models & Mechanisms, 2(9-10), 508-515. doi: 10.1242/dmm.003087 [0547] 89 Horlick, R. A., Macomber, J. L., Bowers, P. M., Neben, T. Y., Tomlinson, G. L., Krapf, I. P., et al. (2013). Simultaneous surface display and secretion of proteins from mammalian cells facilitate efficient in vitro selection and maturation of antibodies. J Biol Chem, 288(27), 19861-19869. doi:10.1074/jbc.M113.452482 [0548] 90 Biffi, G., Tannahill, D., McCafferty, J., & Balasubramanian, S. (2013). Quantitative visualization of DNA G-quadruplex structures in human cells. Nature Chemistry, 5(3), 182-186. doi: 10.1038/nchem. 1548 [0549] 91 Gao, J., Sidhu, S. S., & Wells, J. A. (2009). Two-state selection of conformation-specific antibodies. Proc Natl Acad Sci US A, 106(9), 3071-3076. doi:10.1073/pnas.0812952106 [0550] 92 Gu, G. J., Friedman, M., Jost, C., Johnsson, K., Kamali-Moghaddam, M., Pluckthun, A., et al. (2013). Protein tag-mediated conjugation of oligonucleotides to recombinant affinity binders for proximity ligation. N Biotechnol, 30(2), 144-152. doi: 10.1016/j.nbt.2012.05.005 [0551] 93 Cho, Y. K., & Shusta, E. V. (2010). Antibody library screens using detergent-solubilized mammalian cell lysates as antigen sources. Protein Eng Des Sel, 23(7), 567-577. doi:10.1093/protein/gzq029 [0552] 94 Tillotson, B. J., Cho, Y. K., & Shusta, E. V. (2013). Cells and cell lysates: a direct approach for engineering antibodies against membrane proteins using yeast surface display. Methods, 60(1), 27-37. doi:10.1016/j.ymeth.2012.03.010 [0553] 95 Kunert, A., Straetemans, T., Govers, C., Lamers, C., Mathijssen, R., Sleijfer, S., & Debets, R. (2013). TCR-Engineered T Cells Meet New Challenges to Treat Solid Tumors: Choice of Antigen, T Cell Fitness, and Sensitization of Tumor Milieu. Frontiers in Immunology, 4, 363. doi: 10.3389/fimmu.2013.00363 [0554] 96 Liddy, N., Bossi, G., Adams, K. J., Lissina, A., Mahon, T. M., Hassan, N. J., et al. (2012). Monoclonal TCR-redirected tumor cell killing. Nature Medicine, 18(6), 980-987. doi: 10.1038/nm.2764 [0555] 97 Holler, P. D., Holman, P. O., Shusta, E. V., O'Herrin, S., Wittrup, K. D., & Kranz, D. M. (2000). In vitro evolution of a T cell receptor with high affinity for peptide/MHC. Proc Natl Acad Sci USA, 97(10), 5387-5392. doi:10.1073/pnas.080078297 [0556] 98 Weber, K. S., Donermeyer, D. L., Allen, P. M., & Kranz, D. M. (2005). Class II-restricted T cell receptor engineered in vitro for higher affinity retains peptide specificity and function. Proc Natl Acad Sci USA, 102(52), 19033-19038. doi:10.1073/pnas.0507554102 [0557] 99 Kessels, H. W., van Den Boom, M. D., Spits, H., Hooijberg, E., & Schumacher, T. N. (2000). Changing T cell specificity by retroviral T cell receptor display. Proc Natl Acad Sci USA, 97(26), 14578-14583. doi:10.1073/pnas.97.26.14578 [0558] 100 Chervin, A. S., Aggen, D. H., Raseman, J. M., & Kranz, D. M. (2008). Engineering higher affinity T cell receptors using a T cell display system. Journal of Immunological Methods, 339(2), 175-184. [0559] 101 Crawford, F., Jordan, K. R., Stadinski, B., Wang, Y., Huseby, E., Marrack, P., et al. (2006). Use of baculovirus MHC/peptide display libraries to characterize T-cell receptor ligands. Immunological Reviews, 210, 156-170. doi:10.1111/j.0105-2896.2006.00365.x [0560] 102 Hinrichs, C. S., & Restifo, N. P. (2013). Reassessing target antigens for adoptive T-cell therapy. Nat Biotechnol, 31(11), 999-1008. doi: 10.1038/nbt.2725 [0561] 103 Sadelain, M., Brentjens, R., & Riviere, I. (2013). The basic principles of chimeric antigen receptor design. Cancer Discovery, 3(4), 388-398. doi:10.1158/2159-8290.CD-12-0548 [0562] 104 Alonso-Camino, V., Sanchez-Martin, D., Compte, M., Sanz, L., & Alvarez-Vallina, L. (2009). Lymphocyte display: a novel antibody selection platform based on T cell activation. PLoS ONE, 4(9), e7174. doi:10.1371/journal.pone.0007174 [0563] 105 Melidoni, A. N., Dyson, M. R., Wormald, S., & McCafferty, J. (2013). Selecting antagonistic antibodies that control differentiation through inducible expression in embryonic stem cells. Proceedings of the National Academy of Sciences, 110(44), 17802-17807. doi: 10.1073/pnas.1312062110 [0564] 106 Zhang, H., Wilson, I. A., & Lerner, R. A. (2012). Selection of antibodies that regulate phenotype from intracellular combinatorial antibody libraries. Proceedings of the National Academy of Sciences, 109(39), 15728-15733. doi:10.1073/pnas.1214275109 [0565] 107 Xie, J., Yea, K., Zhang, H., Moldt, B., He, L., Zhu, J., & Lerner, R. A. (2014). Prevention of cell death by antibodies selected from intracellular combinatorial libraries. Chemistry & Biology, 21(2), 274-283. [0566] 108 Yea, K., Zhang, H., Xie, J., Jones, T. M., Yang, G., Song, B. D., & Lerner, R. A. (2013). Converting stem cells to dendritic cells by agonist antibodies from unbiased morphogenic selections (Vol. 110, pp. 14966-14971). Presented at the Proceedings of the National Academy of Sciences of the United States of America. doi: 10.1073/pnas.1313671110 [0567] 109 Kawahara, M., Kimura, H., Ueda, H., & Nagamune, T. (2004). Selection of genetically modified cell population using hapten-specific antibody/receptor chimera. Biochem Biophys Res Commun, 315(1), 132-138. doi:10.1016/j.bbrc.2004.01.030 [0568] 110 Kawahara, M., Shimo, Y., Sogo, T., Hitomi, A., Ueda, H., & Nagamune, T. (2008). Antigen-mediated migration of murine pro-B Ba/F3 cells via an antibody/receptor chimera. Journal of Biotechnology, 133(1), 154-161. doi:10.1016/j.jbiotec.2007.09.009 [0569] 111 Sogo, T., Kawahara, M., Ueda, H., Otsu, M., Onodera, M., Nakauchi, H., & Nagamune, T. (2009). T cell growth control using hapten-specific antibody/interleukin-2 receptor chimera. Cytokine, 46(1), 127-136. doi:10.1016/j.cyto.2008.12.020 [0570] 112 Kawahara, M., Chen, J., Sogo, T., Teng, J., Otsu, M., Onodera, M., et al. (2011). Growth promotion of genetically modified hematopoietic progenitors using an antibody/c-Mpl chimera. Cytokine, 55(3), 402-408. doi:10.1016/j.cyto.2011.05.024 [0571] 113 Ueda, H., Kawahara, M., Aburatani, T., Tsumoto, K., Todokoro, K., Suzuki, E., et al. (2000). Cell-growth control by monomeric antigen: the cell surface expression of lysozyme-specific Ig V-domains fused to truncated Epo receptor. J Immunol Methods, 241(1-2), 159-170. [0572] 114 Kerppola, T. K. (2009). Visualization of molecular interactions using bimolecular fluorescence complementation analysis: Characteristics of protein fragment complementation. Chemical Society Reviews, 38(10), 2876-2886. [0573] 115 Michnick, S. W., Ear, P. H., Manderson, E. N., Remy, I., & Stefan, E. (2007). Universal strategies in research and drug discovery based on protein-fragment complementation assays. Nature Reviews Drug Discovery, 6(7), 569-582. doi: 10.1038/nrd231 [0574] 116 Petschnigg, J., Groisman, B., Kotlyar, M., Taipale, M., Zheng, Y., Kurat, C. F., et al. (2014). The mammalian-membrane two-hybrid assay (MaMTH) for probing membrane-protein interactions in human cells. Nat Methods. doi:10.1038/nmeth.2895 [0575] 117 Renaut, L., Monnet, C., Dubreuil, O., Zaki, O., Crozet, F., Bouayadi, K., et al. (2012). Affinity maturation of antibodies. Optimized methods to generate high-quality scfv libraries and isolate igg candidates by high-throughput screening. Methods in Molecular Biology (Vol. 907, pp. 451-461) [0576] 118 Dyson, M. R., Zheng, Y., Zhang, C., Colwill, K., Pershad, K., Kay, B. K., et al. (2011). Mapping protein interactions by combining antibody affinity maturation and mass spectrometry. Anal Biochem, 417(1), 25-35. doi:10.1016/j.ab.2011.05.005 [0577] 119 de Felipe P (2002) Polycistronic viral vectors. Curr Gene Ther 2: 355-378. doi: 10.2174/1566523023347742. [0578] 120 Foote, J., & Winter, G. (1992). Antibody framework residues affecting the conformation of the hypervariable loops. J Mol Biol, 224(2), 487-499. [0579] 121 Massie, B., Dionne, J., Lamarche, N., Fleurent, J., & Langelier, Y. (1995). Improved adenovirus vector provides herpes simplex virus ribonucleotide reductase R1 and R2 subunits very efficiently. Nature Biotechnology, 13(6), 602-608. [0580] 122 Kim, D. W., Uetsuki, T., Kaziro, Y., Yamaguchi, N., & Sugano, S. (1990). Use of the human elongation factor 1 alpha promoter as a versatile and efficient expression system. Gene, 91(2), 217-223. [0581] 123 Holden, P., Keene, D. R., Lunstrum, G. P., Bachinger, H. P., & Horton, W. A. (2005). Secretion of cartilage oligomeric matrix protein is affected by the signal peptide. J Biol Chem, 280(17), 17172-17179. [0582] 124 Sadelain, M., Papapetrou, E. P., & Bushman, F. D. (2011). Safe harbours for the integration of new DNA in the human genome. Nature Reviews Cancer, 12(1), 51-58. [0583] 125 Sanjana, N. E., Cong, L., Zhou, Y., Cunniff, M. M., Feng, G., & Zhang, F. (2012). A transcription activator-like effector toolbox for genome engineering. Nat Protoc, 7(1), 171-192. doi: 10.1038/nprot.2011.431 [0584] 126 Falk, R., Falk, A., Dyson, M. R., Melidoni, A. N., Parthiban, K., Young, J. L., et al. (2012). Generation of anti-Notch antibodies and their application in blocking Notch signalling in neural stem cells. Methods, 58(1), 69-78. doi: 10.1016/j.ymeth.2012.07.008 [0585] 127 Martin, C. D., Rojas, G., Mitchell, J. N., Vincent, K. J., Wu, J., McCafferty, J., & Schofield, D. J. (2006). A simple vector system to improve performance and utilisation of recombinant antibodies. BMC Biotechnology, 6, 46. [0586] 128 Reyon, D., Tsai, S. Q., Khgayter, C., Foden, J. A., Sander, J. D., & Joung, J. K. (2012). FLASH assembly of TALENs for high-throughput genome editing. Nature Biotechnology, 30(5), 460-465. doi: 10.1038/nbt.2170 [0587] 129 Van Der Weyden, L., Adams, D. J., Harris, L. W., Tannahill, D., Arends, M. J., & Bradley, A. (2005). Null and conditional Semaphorin 3B alleles using a flexible puro.DELTA.tk LoxP/FRT vector. Genesis, 41(4), 171-178 [0588] 130 de Felipe, P., & Ryan, M. D. (2004). Targeting of proteins derived from self-processing polyproteins containing multiple signal sequences. Traffic, 5(8), 616-626. [0589] 131 Raymond, C. S., & Soriano, P. (2007). High-efficiency FLP and PhiC31 site-specific recombination in mammalian cells. PLoS ONE, 2(1), e162. doi: 10.1371/journal.pone.0000162 [0590] 132 Kranz, A., Fu, J., Duerschke, K., Weidlich, S., Naumann, R., Stewart, A. F., & Anastassiadis, K. (2010). An improved Flp deleter mouse in C57Bl/6 based on Flp.sub.o recombinase. Genesis, 48(8), 512-520. doi:10.1002/dvg.20641 [0591] 133 Szymczak A L, Vignali D A (2005) Development of 2 A peptide-based strategies in the design of multicistronic vectors. Expert Opin Biol Ther 5: 627-638. doi: 10.1517/14712598.5.5.627 [0592] 134 Chapple, S. D., Crofts, A. M., Shadbolt, S. P., McCafferty, J., and Dyson, M. R. (2006). Multiplexed expression and screening for recombinant protein production in mammalian cells. BMC biotechnology 6, 49. [0593] 135. Zhao, Y. et al. Multiple injections of electroporated autologous T cells expressing a chimeric antigen receptor mediate regression of human disseminated tumor. Cancer Research 70, 9053-9061 (2010). [0594] 136 Szymczak, A. L. et al. Correction of multi-gene deficiency in vivo using a single `self-cleaving` 2A peptide-based retroviral vector. Nature biotechnology 22, 589-594 (2004). [0595] 137. Li, Y. et al. Directed evolution of human T-cell receptors with picomolar affinities by phage display. Nat. Biotechnol. 23, 349-354 (2005). [0596] 138. Zhao, Y. et al. High-affinity TCRs generated by phage display provide CD4+ T cells with the ability to recognize and kill tumor cell lines. J Immunol. 179, 5845-5854 (2007). [0597] 139. Madura, F. et al. T-cell receptor specificity maintained by altered thermodynamics. The Journal of biological chemistry 288, 18766-18775 (2013). [0598] 140. Pierce, B. G. et al. Computational design of the affinity and specificity of a therapeutic T cell receptor. PLoS Comput Biol 10, e1003478 (2014). [0599] 141. Sebestyen, Z. et al. Human TCR that incorporate CD3zeta induce highly preferred pairing between TCRalpha and beta chains following gene transfer. J. Immunol. 180, 7736-7746 (2008). [0600] 142. Roszik, J. et al. T-cell synapse formation depends on antigen recognition but not CD3 interaction: studies with TCR:.xi., a candidate transgene for TCR gene therapy. Eur. J. Immunol. 41, 1288-1297 (2011). [0601] 143. Cohen, C. J., Zhao, Y., Zheng, Z., Rosenberg, S. A. & Morgan, R. A. Enhanced antitumor activity of murine-human hybrid T-cell receptor (TCR) in human lymphocytes is associated with improved pairing and TCR/CD3 stability. Cancer Research 66, 8878-8886 (2006). [0602] 144. Huovinen, T. et al. Primer extension mutagenesis powered by selective rolling circle amplification. PLoS ONE 7, e31817 (2012). [0603] 145. Cribbs, A. P., Kennedy, A., Gregory, B. & Brennan, F. M. Simplified production and concentration of lentiviral vectors to achieve high transduction in primary human T cells. BMC biotechnology 13, 98 (2013). [0604] 146. Oelke, M. et al. Ex vivo induction and expansion of antigen-specific cytotoxic T cells by HLA-Ig-coated artificial antigen-presenting cells. Nat Med 9, 619-624 (2003). [0605] 147. Wolff, M. & Greenberg, P. D. Antigen-specific activation and cytokine-facilitated expansion of na
ive, human CD8+ T cells. Nature Protocols 9, 950-966 (2014). [0606] 148. Lipowska-Bhalla, G., Gilham, D. E., Hawkins, R. E. & Rothwell, D. G. Isolation of tumor antigen-specific single-chain variable fragments using a chimeric antigen receptor bicistronic retroviral vector in a Mammalian screening protocol. Hum Gene Ther Methods 24, 381-391 (2013). [0607] 149. Kelly, R. J., Sharon, E., Pastan, I. & Hassan, R. Mesothelin-targeted agents in clinical trials and in preclinical development. Mol. Cancer Ther. 11, 517-525 (2012). [0608] 150. Atanackovic, D. et al. Surface molecule CD229 as a novel target for the diagnosis and treatment of multiple myeloma. Haematologica 96, 1512-1520 (2011). [0609] 151. Bund, D., Mayr, C., Kofler, D. M., Hallek, M. & Wendtner, C.-M. Human Ly9 (CD229) as novel tumor-associated antigen (TAA) in chronic lymphocytic leukemia (B-CLL) recognized by autologous CD8+ T cells. Exp. Hematol. 34, 860-869 (2006). [0610] 152. Tiede, C. et al. Adhiron: a stable and versatile peptide display scaffold for molecular recognition applications. Protein Eng. Des. Sel. 27, 145-155 (2014). [0611] 153. Maresca, M., Lin, V. G., Guo, N. & Yang, Y. Obligate ligation-gated recombination (ObLiGaRe): custom-designed nuclease-mediated targeted integration through nonhomologous end joining. Genome Res. 23, 539-546 (2013). [0612] 154. McVey, M. & Lee, S. E. MMEJ repair of double-strand breaks (director's cut): deleted sequences and alternative endings. Trends Genet. 24, 529-538 (2008). [0613] 155. Nakade, S. et al. Microhomology-mediated end-joining-dependent integration of donor DNA in cells and animals using TALENs and CRISPR/Cas9. Nat Commun 5, 5560 (2014). [0614] 156. Chiche, L. et al. Squash inhibitors: From structural motifs to macrocyclic knottins. Current Protein & Peptide Science 5, 341-349.
Sequence CWU 1
1
14215396DNAArtificial SequencepD1 dual promoter antibody expression
cassette for surface expression 1ggtaccgaat tccgtgaggc tccggtgccc
gtcagtgggc agagcgcaca tcgcccacag 60tccccgagaa gttgggggga ggggtcggca
attgaaccgg tgcctagaga aggtggcgcg 120gggtaaactg ggaaagtgat
gtcgtgtact ggctccgcct ttttcccgag ggtgggggag 180aaccgtatat
aagtgcagta gtcgccgtga acgttctttt tcgcaacggg tttgccgcca
240gaacacaggt aagtgccgtg tgtggttccc gcgggcctgg cctctttacg
ggttatggcc 300cttgcgtgcc ttgaattact tccacctggc tccagtacgt
gattcttgat cccgagctgg 360agccaggggc gggccttgcg ctttaggagc
cccttcgcct cgtgcttgag ttgaggcctg 420gcctgggcgc tggggccgcc
gcgtgcgaat ctggtggcac cttcgcgcct gtctcgctgc 480tttcgataag
tctctagcca tttaaaattt ttgatgacct gctgcgacgc tttttttctg
540gcaagatagt cttgtaaatg cgggccagga tctgcacact ggtatttcgg
tttttgggcc 600cgcggccggc gacggggccc gtgcgtccca gcgcacatgt
tcggcgaggc ggggcctgcg 660agcgcggcca ccgagaatcg gacgggggta
gtctcaagct ggccggcctg ctctggtgcc 720tggcctcgcg ccgccgtgta
tcgccccgcc ctgggcggca aggctggccc ggtcggcacc 780agttgcgtga
gcggaaagat ggccgcttcc cggccctgct ccagggggct caaaatggag
840gacgcggcgc tcgggagagc gggcgggtga gtcacccaca caaaggaaaa
gggcctttcc 900gtcctcagcc gtcgcttcat gtgactccac ggagtaccgg
gcgccgtcca ggcacctcga 960ttagttctgg agcttttgga gtacgtcgtc
tttaggttgg ggggaggggt tttatgcgat 1020ggagtttccc cacactgagt
gggtggagac tgaagttagg ccagcttggc acttgatgta 1080attctcgttg
gaatttgccc tttttgagtt tggatcttgg ttcattctca agcctcagac
1140agtggttcaa agtttttttc ttccatttca ggtgtcgtga gacgtggcca
ccatgagggc 1200ctggatcttc tttctccttt gcctggccgg gagggctctg
gcagctagcg acatccagat 1260gacccagagc ccaagcagcc tgagcgccag
cgtgggtgac agagtgacca tcacctgtag 1320agccagcggt aacatccaca
actacctggc ttggtaccag cagaagccag gtaaggctcc 1380aaagctgctg
atctactaca ccaccaccct ggctgacggt gtgccaagca gattcagcgg
1440tagcggtagc ggtaccgact acaccttcac catcagcagc ctccagccag
aggacatcgc 1500cacctactac tgccagcact tctggagcac cccaaggacg
ttcggccaag ggaccaaggt 1560ggaaatcaaa cgtaccgcgg ccgccccttc
cgtgttcatc ttccctccct ccgacgagca 1620gctgaagtcc ggcaccgcct
ctgtggtgtg cctgctgaac aacttctacc ctcgggaggc 1680caaggtgcag
tggaaggtgg acaacgccct gcagtccggc aactcccagg aatccgtcac
1740cgagcaggac tccaaggact ctacctactc cctgtcctcc accctgaccc
tgtccaaggc 1800cgactacgag aagcacaagc tgtacgcctg cgaagtgacc
caccagggcc tgtcctctcc 1860cgtgaccaag tccttcaacc ggggcgagtg
ctaataaaag cttacgacgt gatcagcctc 1920gactgtgcct tctagttgcc
agccatctgt tgtttgcccc tcccccgtgc cttccttgac 1980cctggaaggt
gccactccca ctgtcctttc ctaataaaat gaggaaattg catcgcattg
2040tctgagtagg tgtcattcta ttctgggggg tggggtgggg caggacagca
agggggagga 2100ttgggaagac aatagcaggc atgctgggga acattgatta
ttgactagtt attaatagta 2160atcaattacg gggtcattag ttcatagccc
atatatggag ttccgcgtta cataacttac 2220ggtaaatggc ccgcctggct
gaccgcccaa cgacccccgc ccattgacgt caataatgac 2280gtatgttccc
atagtaacgc caatagggac tttccattga cgtcaatggg tggagtattt
2340acggtaaact gcccacttgg cagtacatca agtgtatcat atgccaagtc
cgccccctat 2400tgacgtcaat gacggtaaat ggcccgcctg gcattatgcc
cagtacatga ccttacggga 2460ctttcctact tggcagtaca tctacgtatt
agtcatcgct attaccatag tgatgcggtt 2520ttggcagtac accaatgggc
gtggatagcg gtttgactca cggggatttc caagtctcca 2580ccccattgac
gtcaatggga gtttgttttg gcaccaaaat caacgggact ttccaaaatg
2640tcgtaataac cccgccccgt tgacgcaaat gggcggtagg cgtgtacggt
gggaggtcta 2700tataagcaga gctcgtttag tgaaccgtca gatcctcact
ctcttccgca tcgctgtctg 2760cgagggccag ctgttgggct cgcggttgag
gacaaactct tcgcggtctt tccagtactc 2820ttggatcgga aacccgtcgg
cctccgaacg gtactccgcc accgagggac ctgagcgagt 2880ccgcatcgac
cggatcggaa aacctctcgt gaaaggcgtc taaccagtca cagtcgcaag
2940gtaggctgag caccgtggcg ggcggcagcg ggtggcggtc ggggttgttt
ctggcggagg 3000tgctgctgat gatgtaatta aagtaggcgg tcttgagacg
gcggatggtc gaggtgaggt 3060gtggcaggct tgagatccag ctgttggggt
gagtactccc tctcaaaagc gggcattact 3120tctgcgctaa gattgtcagt
ttccaaaaac gaggaggatt tgatattcac ctggcccgat 3180ctggccatac
acttgagtga caatgacatc cactttgcct ttctctccac aggtgtccac
3240tcccaggtcc aagtttgtgg aaattaatac gacgtggcca ccatgagttg
gagctgtatc 3300atcctcttct tggtagcaac agctacaggt aaggggttaa
cagtagcagg cttgaggtct 3360ggacatatat atgggtgaca atgacatcca
ctttgccttt ctctccacag gcgccatggc 3420ccaggtccaa ctgcaggaga
gcggtccagg tcttgtgaga cctagccaga ccctgagcct 3480gacctgcacc
gtgtctggca gcaccttcag cggctatggt gtaaactggg tgagacagcc
3540acctggacga ggtcttgagt ggattggaat gatttggggt gatggaaaca
cagactataa 3600ttcagctctc aaatccagag tgacaatgct ggtagacacc
agcaagaacc agttcagcct 3660gagactcagc agcgtgacag ccgccgacac
cgcggtctat tattgtgcaa gagagagaga 3720ttataggctt gactactggg
gtcaaggcag cctcgtcaca gtctcgagtg ccagcaccaa 3780gggccccagc
gtgttccctc tggccccctg tagcagaagc accagcgaga gcacagccgc
3840cctgggctgc ctggtcaagg actacttccc cgagcccgtg accgtgtcct
ggaactctgg 3900cgctctgacc agcggcgtgc acacctttcc agccgtgctg
cagagcagcg gcctgtacag 3960cctgagcagc gtggtcaccg tgcccagcag
caacttcggc acccagacct acacctgtaa 4020cgtggaccac aagcccagca
acaccaaggt ggacaagacc gtggaacgga agtgctgcgt 4080ggaatgcccc
ccctgtcccg ctcctccagt ggctggacct tccgtgttcc tgttcccccc
4140aaagcccaag gacaccctga tgatcagccg gacccccgaa gtgacctgcg
tggtggtgga 4200cgtgtcccac gaggaccccg aggtgcagtt caattggtac
gtggacggcg tggaagtgca 4260caacgccaag accaagccca gagaggaaca
gttcaacagc accttccggg tggtgtccgt 4320gctgaccgtg gtgcaccagg
actggctgaa cggcaaagag tacaagtgcg ccgtctccaa 4380caagggcctg
cctgccccca tcgagaaaac catcagcaag accaagggcc agcctcgcga
4440gcctcaggtg tacacactgc cccccagccg ggaagagatg accaagaacc
aggtgtccct 4500gacctgcctc gtgaagggct tctaccccag cgatatcgcc
gtggaatggg agagcaacgg 4560ccagcccgag aacaactaca agaccacccc
ccccatgctg gacagcgacg gctcattctt 4620cctgtacagc aagctgacag
tggacaagag ccggtggcag cagggcaacg tgttcagctg 4680cagcgtgatg
cacgaggccc tgcacaacca ctacacccag aagtccctga gcctgagccc
4740cggcaaggga tccaaggtaa gtttaaacat atatataact ttaaataatt
ggcattattt 4800aaagttacta ctaactaacc ctgattattt aaattttcag
gaacaaaaac tcatctcaga 4860agaggatctg aatgctgtgg gccaggacac
gcaggaggtc atcgtggtgc cacactcctt 4920gccctttaag gtggtggtga
tctcagccat cctggccctg gtggtgctca ccatcatctc 4980ccttatcatc
ctcatcatgc tttggcagaa gaagccacgt tagtaacagg taagagtgta
5040actttaaata atgccaatta tttaaagtta ctgactctct ctgcttacga
cgcttcttct 5100tttttttttc ctgcaggggt agtaatcagc ctcgactgtg
ccttctagtt gccagccatc 5160tgttgtttgc ccctcccccg tgccttcctt
gaccctggaa ggtgccactc ccactgtcct 5220ttcctaataa aatgaggaaa
ttgcatcgca ttgtctgagt aggtgtcatt ctattctggg 5280gggtggggtg
gggcaggaca gcaaggggga ggattgggaa gacaatagca ggcatgctgg
5340ggatggcccg ggcatgataa cttcgtataa tgtatgctat acgaagttat gtatac
53962233PRTArtificial sequenceBM40 Leader, Humanised D1.3 VL and
Human C Kappa sequence 2Met Arg Ala Trp Ile Phe Phe Leu Leu Cys Leu
Ala Gly Arg Ala Leu1 5 10 15Ala Ala Ser Asp Ile Gln Met Thr Gln Ser
Pro Ser Ser Leu Ser Ala 20 25 30Ser Val Gly Asp Arg Val Thr Ile Thr
Cys Arg Ala Ser Gly Asn Ile 35 40 45His Asn Tyr Leu Ala Trp Tyr Gln
Gln Lys Pro Gly Lys Ala Pro Lys 50 55 60Leu Leu Ile Tyr Tyr Thr Thr
Thr Leu Ala Asp Gly Val Pro Ser Arg65 70 75 80Phe Ser Gly Ser Gly
Ser Gly Thr Asp Tyr Thr Phe Thr Ile Ser Ser 85 90 95Leu Gln Pro Glu
Asp Ile Ala Thr Tyr Tyr Cys Gln His Phe Trp Ser 100 105 110Thr Pro
Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr 115 120
125Ala Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
130 135 140Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
Tyr Pro145 150 155 160Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
Ala Leu Gln Ser Gly 165 170 175Asn Ser Gln Glu Ser Val Thr Glu Gln
Asp Ser Lys Asp Ser Thr Tyr 180 185 190Ser Leu Ser Ser Thr Leu Thr
Leu Ser Lys Ala Asp Tyr Glu Lys His 195 200 205Lys Leu Tyr Ala Cys
Glu Val Thr His Gln Gly Leu Ser Ser Pro Val 210 215 220Thr Lys Ser
Phe Asn Arg Gly Glu Cys225 230315PRTArtificial sequenceLeader 1
sequence 3Met Ser Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala
Thr1 5 10 154449PRTArtificial sequenceLeader 2 and D1.3 VH
sequence, CH1 domain, Hinge, CH2 and CH3 domains 4Gly Ala Met Ala
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val1 5 10 15Arg Pro Ser
Gln Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Ser Thr 20 25 30Phe Ser
Gly Tyr Gly Val Asn Trp Val Arg Gln Pro Pro Gly Arg Gly 35 40 45Leu
Glu Trp Ile Gly Met Ile Trp Gly Asp Gly Asn Thr Asp Tyr Asn 50 55
60Ser Ala Leu Lys Ser Arg Val Thr Met Leu Val Asp Thr Ser Lys Asn65
70 75 80Gln Phe Ser Leu Arg Leu Ser Ser Val Thr Ala Ala Asp Thr Ala
Val 85 90 95Tyr Tyr Cys Ala Arg Glu Arg Asp Tyr Arg Leu Asp Tyr Trp
Gly Gln 100 105 110Gly Ser Leu Val Thr Val Ser Ser Ala Ser Thr Lys
Gly Pro Ser Val 115 120 125Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr
Ser Glu Ser Thr Ala Ala 130 135 140Leu Gly Cys Leu Val Lys Asp Tyr
Phe Pro Glu Pro Val Thr Val Ser145 150 155 160Trp Asn Ser Gly Ala
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val 165 170 175Leu Gln Ser
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro 180 185 190Ser
Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys 195 200
205Pro Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val
210 215 220Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser
Val Phe225 230 235 240Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
Ile Ser Arg Thr Pro 245 250 255Glu Val Thr Cys Val Val Val Asp Val
Ser His Glu Asp Pro Glu Val 260 265 270Gln Phe Asn Trp Tyr Val Asp
Gly Val Glu Val His Asn Ala Lys Thr 275 280 285Lys Pro Arg Glu Glu
Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val 290 295 300Leu Thr Val
Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys305 310 315
320Ala Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
Pro Pro 340 345 350Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
Thr Cys Leu Val 355 360 365Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
Glu Trp Glu Ser Asn Gly 370 375 380Gln Pro Glu Asn Asn Tyr Lys Thr
Thr Pro Pro Met Leu Asp Ser Asp385 390 395 400Gly Ser Phe Phe Leu
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp 405 410 415Gln Gln Gly
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His 420 425 430Asn
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Ser 435 440
445Lys560PRTArtificialMyc epitope, PDGFR spacer and PDGFR
Transmembrane region (TM) 5Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu
Asn Ala Val Gly Gln Asp1 5 10 15Thr Gln Glu Val Ile Val Val Pro His
Ser Leu Pro Phe Lys Val Val 20 25 30Val Ile Ser Ala Ile Leu Ala Leu
Val Val Leu Thr Ile Ile Ser Leu 35 40 45Ile Ile Leu Ile Met Leu Trp
Gln Lys Lys Pro Arg 50 55 6061542DNAArtificial SequencepD1-huD1.3
donor construct fragment containing AAVS1 left homology arm, splice
acceptor site, Blasticidin gene, SV40 polyA 6ggtaccgaat tcgccctttg
ctttctctga cctgcattct ctcccctggg cctgtgccgc 60tttctgtctg cagcttgtgg
cctgggtcac ctctacggct ggcccagatc cttccctgcc 120gcctccttca
ggttccgtct tcctccactc cctcttcccc ttgctctctg ctgtgttgct
180gcccaaggat gctctttccg gagcacttcc ttctcggcgc tgcaccacgt
gatgtcctct 240gagcggatcc tccccgtgtc tgggtcctct ccgggcatct
ctcctccctc acccaacccc 300atgccgtctt cactcgctgg gttccctttt
ccttctcctt ctggggcctg tgccatctct 360cgtttcttag gatggccttc
tccgacggat gtctcccttg cgtcccgcct ccccttcttg 420taggcctgca
tcatcaccgt ttttctggac aaccccaaag taccccgtct ccctggcttt
480agccacctct ccatcctctt gctttctttg cctggacacc ccgttctcct
gtggattcgg 540gtcacctctc actcctttca tttgggcagc tcccctaccc
cccttacctc tctagtctgt 600gcaagctctt ccagccccct gtcatggcat
cttccagggg tccgagagct cagctagtct 660tcttcctcca acccgggccc
ctatgtccac ttcaggacag catgtttgct gcctccaggg 720atcctgtgtc
cccgagctgg gaccacctta tattcccagg gccggttaat gtggctctgg
780ttctgggtac ttttatctgt cccctccacc ccacagtggg gcaagatgca
tcttctgacc 840tcttctcttc ctcccacagg gcatggcaaa acctctgagc
caggaagaaa gcacactgat 900tgaaagagca accgctacta tcaacagcat
ccccatctcc gaagactatt ctgtggctag 960tgccgctctg tccagcgacg
ggagaatctt caccggtgtg aacgtctacc actttacagg 1020cggaccatgc
gcagagctgg tggtcctggg gactgcagcc gctgcagccg ctggtaatct
1080gacctgtatc gtggccattg gcaacgaaaa taggggcatc ctgtccccat
gcggcaggtg 1140tcggcaggtg ctgctggatc tgcatcctgg catcaaggca
attgtcaaag actctgatgg 1200acagcctacc gccgtcggta tccgtgaact
gctgcctagc ggctatgtct gggagggata 1260atgagcttgg cttcgaaatg
accgaccaag cgacgcccaa cctgccatca cgagatttcg 1320attccaccgc
cgccttctat gaaaggttgg gcttcggaat cgttttccgg gacgccggct
1380ggatgatcct ccagcgcggg gatctcatgc tggagttctt cgcccacccc
aacttgttta 1440ttgcagctta taatggttac aaataaagca atagcatcac
aaatttcaca aataaagcat 1500ttttttcact gcattctagt tgtggtttaa
ttaagtcaat tc 15427924DNAArtificial SequencepD1-huD1.3 fragment
containing AAVS1 right homology arm 7tggcccgggc atgataactt
cgtataatgt atgctatacg aagttatgta tacggcgcgc 60ccactaggga caggattggt
gacagaaaag ccccatcctt aggcctcctc cttcctagtc 120tcctgatatt
gggtctaacc cccacctcct gttaggcaga ttccttatct ggtgacacac
180ccccatttcc tggagccatc tctctccttg ccagaacctc taaggtttgc
ttacgatgga 240gccagagagg atcctgggag ggagagcttg gcagggggtg
ggagggaagg gggggatgcg 300tgacctgccc ggttctcagt ggccaccctg
cgctaccctc tcccagaacc tgagctgctc 360tgacgcggct gtctggtgcg
tttcactgat cctggtgctg cagcttcctt acacttccca 420agaggagaag
cagtttggaa aaacaaaatc agaataagtt ggtcctgagt tctaactttg
480gctcttcacc tttctagtcc ccaatttata ttgttcctcc gtgcgtcagt
tttacctgtg 540agataaggcc agtagccagc cccgtcctgg cagggctgtg
gtgaggaggg gggtgtccgt 600gtggaaaact ccctttgtga gaatggtgcg
tcctaggtgt tcaccaggtc gtggccgcct 660ctactccctt tctctttctc
catccttctt tccttaaaga gtccccagtg ctatctggga 720catattcctc
cgcccagagc agggtcccgc ttccctaagg ccctgctctg ggcttctggg
780tttgagtcct tggcaagccc aggagaggcg ctcaggcttc cctgtccccc
ttcctcgtcc 840accatctcat gcccctggct ctcctgcccc ttccctacag
gggttcctgg ctctgctctg 900acgcgtgtat actcgatctt tccg
9248132PRTArtificial SequenceSynthetic Blasticidin resistance gene
8Met Ala Lys Pro Leu Ser Gln Glu Glu Ser Thr Leu Ile Glu Arg Ala1 5
10 15Thr Ala Thr Ile Asn Ser Ile Pro Ile Ser Glu Asp Tyr Ser Val
Ala 20 25 30Ser Ala Ala Leu Ser Ser Asp Gly Arg Ile Phe Thr Gly Val
Asn Val 35 40 45Tyr His Phe Thr Gly Gly Pro Cys Ala Glu Leu Val Val
Leu Gly Thr 50 55 60Ala Ala Ala Ala Ala Ala Gly Asn Leu Thr Cys Ile
Val Ala Ile Gly65 70 75 80Asn Glu Asn Arg Gly Ile Leu Ser Pro Cys
Gly Arg Cys Arg Gln Val 85 90 95Leu Leu Asp Leu His Pro Gly Ile Lys
Ala Ile Val Lys Asp Ser Asp 100 105 110Gly Gln Pro Thr Ala Val Gly
Ile Arg Glu Leu Leu Pro Ser Gly Tyr 115 120 125Val Trp Glu Gly
1309734DNAArtificial SequencepD6 vector for scFv-Fc expression,
fragment from Nco1 site to Pme1 site containing a fragment (last 4
amino acids) of the leader sequence, the complete CH2 domain and
CH3 domain 9ggcgccatgg cccaggtcgc ggccgccgtg gaatgccccc cctgtcccgc
tcctccagtg 60gctggacctt ccgtgttcct gttcccccca aagcccaagg acaccctgat
gatcagccgg 120acccccgaag tgacctgcgt ggtggtggac gtgtcccacg
aggaccccga ggtgcagttc 180aattggtacg tggacggcgt ggaagtgcac
aacgccaaga ccaagcccag agaggaacag 240ttcaacagca ccttccgggt
ggtgtccgtg ctgaccgtgg tgcaccagga ctggctgaac 300ggcaaagagt
acaagtgcgc cgtctccaac aagggcctgc ctgcccccat cgagaaaacc
360atcagcaaga ccaagggcca gcctcgcgag cctcaggtgt acacactgcc
ccccagccgg 420gaagagatga ccaagaacca ggtgtccctg acctgcctcg
tgaagggctt ctaccccagc 480gatatcgccg tggaatggga gagcaacggc
cagcccgaga acaactacaa gaccaccccc 540cccatgctgg acagcgacgg
ctcattcttc ctgtacagca agctgacagt ggacaagagc 600cggtggcagc
agggcaacgt gttcagctgc agcgtgatgc acgaggccct gcacaaccac
660tacacccaga agtccctgag cctgagcccc ggcaagggat ccaaggtaag
tttaaacata 720tatataactt taaa 734104PRTArtificial SequenceVector
pD6, fragment at the end of leader sequence 10Gly Ala Met
Ala111219PRTArtificial SequenceVector pD6 CH2 and CH3 domains 11Ala
Pro
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro1 5 10 15Lys
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val 20 25
30Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val
35 40 45Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
Gln 50 55 60Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val
His Gln65 70 75 80Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Ala Val
Ser Asn Lys Gly 85 90 95Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
Thr Lys Gly Gln Pro 100 105 110Arg Glu Pro Gln Val Tyr Thr Leu Pro
Pro Ser Arg Glu Glu Met Thr 115 120 125Lys Asn Gln Val Ser Leu Thr
Cys Leu Val Lys Gly Phe Tyr Pro Ser 130 135 140Asp Ile Ala Val Glu
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr145 150 155 160Lys Thr
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr 165 170
175Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
180 185 190Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
Gln Lys 195 200 205Ser Leu Ser Leu Ser Pro Gly Lys Gly Ser Lys 210
2151260DNAArtificial SequenceFragment of sequence of pD4 vector
preceding and including the start of the AAVS1 left homology arm
12ggtaccgaat tcgccctttg ctttctctga cctgcattct ctcccctggg cctgtgccgc
60133933DNAArtificial sequenceFragment of sequence of vector pD4
including the end of the AAVS1 LHA, FRT site, Lox2272 site, 1-Sce1
meganuclease site, GFP left TALEN binding site, GFP right TALEN
binding site, T2A, GFP, PGK promoter, puromycin delta TK, loxP site
and start of AAVS1 RHA 13ttctgggtac ttttatctgt cccctccacc
ccacagtggg gcaagatgca tgaagttcct 60attccgaagt tcctattctc tagaaagtat
aggaacttcg accataactt cgtataaagt 120atcctatacg aagttatgcg
atcgctcgcg cgtagggata acagggtaat aagtccaccg 180gtcgccacca
tggtgagcaa gggcgaggag ctgttcactt ctgacctctt ctcttcctcc
240cacagggcct agagagatct ggcagcggag agggcagagg aagtcttcta
acatgcggtg 300acgtggagga gaatcccgga ccgatggtga gcaagggaga
agaactcttc accggggtgg 360tgcccatcct ggtcgagctg gacggcgacg
tgaacggcca caagttcagc gtgtccggcg 420agggcgaggg cgatgccacc
tacggcaagc tgaccctgaa gttcatctgc accaccggca 480agctgcccgt
gccctggccc accctcgtga ccaccctgac ctacggcgtg cagtgcttca
540gccgctaccc cgaccacatg aagcagcacg acttcttcaa gtccgccatg
cccgaaggct 600acgtccagga gcgcaccatc ttcttcaagg acgacggcaa
ctacaagacc cgcgccgagg 660tgaagttcga gggcgacacc ctggtgaacc
gcatcgagct gaagggcatc gacttcaagg 720aggacggcaa catcctgggg
cacaagctgg agtacaacta caacagccac aacgtctata 780tcatggccga
caagcagaag aacggcatca aggtgaactt caagatccgc cacaacatcg
840aggacggcag cgtgcagctc gccgaccact accagcagaa cacccccatc
ggcgacggcc 900ccgtgctgct gcccgacaac cactacctga gcacccagtc
cgccctgagc aaagacccca 960acgagaagcg cgatcacatg gtcctgctgg
agttcgtgac cgccgccggg atcactcacg 1020gcatggacga gctgtacaag
aagctgagcc acggcttccc gccggcggtg gcggcgcagg 1080atgatggcac
gctgcccatg tcttgtgccc aggagagcgg gatggaccgt caccctgcag
1140cctgtgcttc tgctaggatc aatgtgtagg tgatcagcct cgactgtgcc
ttctagttgc 1200cagccatctg ttgtttgccc ctcccccgtg ccttccttga
ccctggaagg tgccactccc 1260actgtccttt cctaataaaa tgaggaaatt
gcatcgcatt gtctgagtag gtgtcattct 1320attctggggg gtggggtggg
gcaggacagc aagggggagg attgggaaga caatagcagg 1380catgctgggg
atggcccaat tctaccgggt aggggaggcg cttttcccaa ggcagtctgg
1440agcatgcgct ttagcagccc cgctgggcac ttggcgctac acaagtggcc
tctggcctcg 1500cacacattcc acatccaccg gtaggcgcca accggctccg
ttctttggtg gccccttcgc 1560gccaccttct actcctcccc tagtcaggaa
gttccccccc gccccgcagc tcgcgtcgtg 1620caggacgtga caaatggaag
tagcacgtct cactagtctc gtgcagatgg acagcaccgc 1680tgagcaatgg
aagcgggtag gcctttgggg cagcggccaa tagcagcttt gctccttcgc
1740tttctgggct cagaggctgg gaaggggtgg gtccgggggc gggctcaggg
gcgggctcag 1800gggcggggcg ggcgcccgaa ggtcctccgg aggcccggca
ttctgcacgc ttcaaaagcg 1860cacgtctgcc gcgctgttct cctcttcctc
atctccgggc ctttcgacct gcagccaacg 1920ccaccatggg gaccgagtac
aagcccacgg tgcgcctcgc cacccgcgac gacgtccccc 1980gggccgtacg
caccctcgcc gccgcgttcg ccgactaccc cgccacgcgc cacaccgtcg
2040acccggaccg ccacatcgag cgggtcaccg agctgcaaga actcttcctc
acgcgcgtcg 2100ggctcgacat cggcaaggtg tgggtcgcgg acgacggcgc
cgcggtggcg gtctggacca 2160cgccggagag cgtcgaagcg ggggcggtgt
tcgccgagat cggcccgcgc atggccgagt 2220tgagcggttc ccggctggcc
gcgcagcaac agatggaagg cctcctggcg ccgcaccggc 2280ccaaggagcc
cgcgtggttc ctggccaccg tcggcgtctc gcccgaccac cagggcaagg
2340gtctgggcag cgccgtcgtg ctccccggag tggaggcggc cgagcgcgcc
ggggtgcccg 2400ccttcctgga gacctccgcg ccccgcaacc tccccttcta
cgagcggctc ggcttcaccg 2460tcaccgccga cgtcgaggtg cccgaaggac
cgcgcacctg gtgcatgacc cgcaagcccg 2520gtgccggatc catgcccacg
ctactgcggg tttatataga cggtcctcac gggatgggga 2580aaaccaccac
cacgcaactg ctggtggccc tgggttcgcg cgacgatatc gtctacgtac
2640ccgagccgat gacttactgg caggtgctgg gggcttccga gacaatcgcg
aacatctaca 2700ccacacaaca ccgcctcgac cagggtgaga tatcggccgg
ggacgcggcg gtggtaatga 2760caagcgccca gataacaatg ggcatgcctt
atgccgtgac cgacgccgtt ctggctcctc 2820atatcggggg ggaggctggg
agctcacatg ccccgccccc ggccctcacc ctcatcttcg 2880accgccatcc
catcgccgcc ctcctgtgct acccggccgc gcgatacctt atgggcagca
2940tgacccccca ggccgtgctg gcgttcgtgg ccctcatccc gccgaccttg
cccggcacaa 3000acatcgtgtt gggggccctt ccggaggaca gacacatcga
ccgcctggcc aaacgccagc 3060gccccggcga gcggcttgac ctggctatgc
tggccgcgat tcgccgcgtt tacgggctgc 3120ttgccaatac ggtgcggtat
ctgcagggcg gcgggtcgtg gcgggaggat tggggacagc 3180tttcggggac
ggccgtgccg ccccagggtg ccgagcccca gagcaacgcg ggcccacgac
3240cccatatcgg ggacacgtta tttaccctgt ttcgggcccc cgagttgctg
gcccccaacg 3300gcgacctgta caacgtgttt gcctgggcct tggacgtctt
ggccaaacgc ctccgtccca 3360tgcacgtctt tatcctggat tacgaccaat
cgcccgccgg ctgccgggac gccctgctgc 3420aacttacctc cgggatggtc
cagacccacg tcaccacccc cggctccata ccgacgatct 3480gcgacctggc
gcgcacgttt gcccgggaga tgggggaggc taactgagct ctagagctcg
3540ctgatcagcc tcgactgtgc cttctagttg ccagccatct gttgtttgcc
cctcccccgt 3600gccttccttg accctggaag gtgccactcc cactgtcctt
tcctaataaa atgaggaaat 3660tgcatcgcat tgtctgagta ggtgtcattc
tattctgggg ggtggggtgg ggcaggacag 3720caagggggag gattgggaag
acaatagcag gcatgctggg gatgcggtgg gctctatggc 3780ttctgaggcg
gaaagaacca gctggggctc gagatccact agttctagcc tcgaggctag
3840agcggccggc cctataactt cgtataatgt atgctatacg aagttatcag
gtaagttaac 3900agggcgcgcc cactagggac aggattggtg aca
39331420DNAArtificial SequenceFragment of pD4 including the end of
the AAVS1 right homology arm (RHA) 14tctgctctga cgcgtgtata
201553PRTArtificial SequenceFragment of pD4 comprising T2A and
start of EGFP sequence 15Gly Ser Gly Glu Gly Arg Gly Ser Leu Leu
Thr Cys Gly Asp Val Glu1 5 10 15Glu Asn Pro Gly Pro Met Val Ser Lys
Gly Glu Glu Leu Phe Thr Gly 20 25 30Val Val Pro Ile Leu Val Glu Leu
Asp Gly Asp Val Asn Gly His Lys 35 40 45Phe Ser Val Ser Gly
501669PRTArtificial SequenceFragment of pD4 comprising terminal
part of EGFP sequence and Mu ornithine decarboxylase PEST sequence
16Asn Glu Lys Arg Asp His Met Val Leu Leu Glu Phe Val Thr Ala Ala1
5 10 15Gly Ile Thr His Gly Met Asp Glu Leu Tyr Lys Lys Leu Ser His
Gly 20 25 30Phe Pro Pro Ala Val Ala Ala Gln Asp Asp Gly Thr Leu Pro
Met Ser 35 40 45Cys Ala Gln Glu Ser Gly Met Asp Arg His Pro Ala Ala
Cys Ala Ser 50 55 60Ala Arg Ile Asn Val651718PRTArtificial
SequenceInitial fragment of of purodelta TK sequence encoded by pD4
17Met Gly Thr Glu Tyr Lys Pro Thr Val Arg Leu Ala Thr Arg Asp Asp1
5 10 15Val Pro1815PRTArtificial SequenceTerminal fragment of
puromycin delta TK sequence encoded by pD4 18Cys Asp Leu Ala Arg
Thr Phe Ala Arg Glu Met Gly Glu Ala Asn1 5 10 15191098DNAArtificial
SequenceSequence of donor plasmid for integration into Flp/GFP
TALEN sites (pD5), fragment preceding D1.3 antibody expression
cassette 19ggtaccgaat tcctagctct tccagccccc tgtcatggca tcttccaggg
gtccgagagc 60tcagctagtc ttcttcctcc aacccgggcc cctatgtcca cttcaggaca
gcatgtttgc 120tgcctccagg gatcctgtgt ccccgagctg ggaccacctt
atattcccag ggccggttaa 180tgtggctctg gttctgggta cttttatctg
tcccctccac cccacagtgg ggcaagatgc 240atgaagttcc tattccgaag
ttcctattct ctagaaagta taggaacttc gaccataact 300tcgtataaag
tatcctatac gaagttatgc gatcgctcgc gcgtagggat aacagggtaa
360taagtccacc ggtcgccacc atggtcttct gacctcttct cttcctccca
cagggcatgg 420caaaacctct gagccaggaa gaaagcacac tgattgaaag
agcaaccgct actatcaaca 480gcatccccat ctccgaagac tattctgtgg
ctagtgccgc tctgtccagc gacgggagaa 540tcttcaccgg tgtgaacgtc
taccacttta caggcggacc atgcgcagag ctggtggtcc 600tggggactgc
agccgctgca gccgctggta atctgacctg tatcgtggcc attggcaacg
660aaaatagggg catcctgtcc ccatgcggca ggtgtcggca ggtgctgctg
gatctgcatc 720ctggcatcaa ggcaattgtc aaagactctg atggacagcc
taccgccgtc ggtatccgtg 780aactgctgcc tagcggctat gtctgggagg
gataatgaat gagcttggct tcgaaatgac 840cgaccaagcg acgcccaacc
tgccatcacg agatttcgat tccaccgccg ccttctatga 900aaggttgggc
ttcggaatcg ttttccggga cgccggctgg atgatcctcc agcgcgggga
960tctcatgctg gagttcttcg cccaccccaa cttgtttatt gcagcttata
atggttacaa 1020ataaagcaat agcatcacaa atttcacaaa taaagcattt
ttttcactgc attctagttg 1080tggtttaatt aacaattc
109820931DNAArtificial SequenceSequence of donor plasmid for
integration into Flp/GFP TALEN sites (pD5), fragment following D1.3
antibody expression cassette 20caggcatgct ggggatggcc cgggcatgat
aacttcgtat aatgtatgct atacgaagtt 60atgtatacgg cgcgcccgag caagggcgag
gagctgttca cttctgacct cttctcttcc 120tcccacctga gcctagagag
atctggcagc ggagagggca gaggaagtct tctaacatgc 180ggtgacgtgg
aggagaatcc cggaccgtga gtgagcaagg gagaagaact cttcaccggg
240gtggtgccca tcctggtcga gctggacggc gacgtgaacg gccacaagtt
cagcgtgtcc 300ggcgagggcg agggcgatgc cacctacggc aagctgaccc
tgaagttcat ctgcaccacc 360ggcaagctgc ccgtgccctg gcccaccctc
gtgaccaccc tgacctacgg cgtgcagtgc 420ttcagccgct accccgacca
catgaagcag cacgacttct tcaagtccgc catgcccgaa 480ggctacgtcc
aggagcgcac catcttcttc aaggacgacg gcaactacaa gacccgcgcc
540gaggtgaagt tcgagggcga caccctggtg aaccgcatcg agctgaaggg
catcgacttc 600aaggaggacg gcaacatcct ggggcacaag ctggagtaca
actacaacag ccacaacgtc 660tatatcatgg ccgacaagca gaagaacggc
atcaaggtga acttcaagat ccgccacaac 720atcgaggacg gcagcgtgca
gctcgccgac cactaccagc agaacacccc catcggcgac 780ggccccgtgc
tgctgcccga caaccactac ctgagcaccc agtccgccct gagcaaagac
840cccaacgaga agcgcgatca catggtcctg ctggagttcg tgaccgccgc
cgggatcact 900cacggcatgg acgagcctga cgcgtgtata c
93121131PRTArtificial SequenceSequence of Blasticidin resistance
protein encoded by donor plasmid for integration into FLP/GFP TALEN
sites (pD5) 21Ala Lys Pro Leu Ser Gln Glu Glu Ser Thr Leu Ile Glu
Arg Ala Thr1 5 10 15Ala Thr Ile Asn Ser Ile Pro Ile Ser Glu Asp Tyr
Ser Val Ala Ser 20 25 30Ala Ala Leu Ser Ser Asp Gly Arg Ile Phe Thr
Gly Val Asn Val Tyr 35 40 45His Phe Thr Gly Gly Pro Cys Ala Glu Leu
Val Val Leu Gly Thr Ala 50 55 60Ala Ala Ala Ala Ala Gly Asn Leu Thr
Cys Ile Val Ala Ile Gly Asn65 70 75 80Glu Asn Arg Gly Ile Leu Ser
Pro Cys Gly Arg Cys Arg Gln Val Leu 85 90 95Leu Asp Leu His Pro Gly
Ile Lys Ala Ile Val Lys Asp Ser Asp Gly 100 105 110Gln Pro Thr Ala
Val Gly Ile Arg Glu Leu Leu Pro Ser Gly Tyr Val 115 120 125Trp Glu
Gly 13022813DNAArtificial SequenceSequence of I-Sce-1 meganuclease
construct 22ccatgggcta tccttacgat gtccctgatt acgccaacag tcctggtatc
cctggtatgg 60gtcctaaaaa gaagcgaaaa gtgggtagac tggaacccgg catgaagaac
attaagaaaa 120atcaggtgat gaacctggga cctaattcca agctgctgaa
agagtacaag tctcagctga 180tcgaactgaa cattgagcag tttgaagcag
ggatcggtct gattctgggg gacgcctaca 240tccggagcag ggatgagggc
aagacttatt gcatgcagtt cgaatggaag aataaggcct 300acatggacca
cgtgtgtctg ctgtatgatc agtgggtcct gtctccccct cacaagaaag
360agagagtgaa ccatctgggc aatctggtca ttacttgggg agcacagacc
ttcaagcatc 420aggcctttaa caaactggct aacctgttca tcgtgaacaa
caagaaaacc atccctaaca 480atctggtcga aaactacctg acaccaatga
gtctggccta ttggttcatg gacgatggcg 540gaaaatggga ctacaacaag
aacagcacaa acaaaagcat cgtgctgaat acccagtcct 600tcacatttga
ggaagtggag tatctggtca agggcctgcg gaacaaattc cagctgaact
660gctacgtgaa gatcaacaag aacaagccaa tcatctacat cgattctatg
agttacctga 720tcttttataa cctgattaag ccatacctga tcccccagat
gatgtataaa ctgcccaata 780caatcagctc cgagactttc ctgaaggtct aga
81323269PRTArtificial SequenceSequence encoded by I-Sce1
meganuclease construct HA tag, NLS, and I-Sce1 23Met Gly Tyr Pro
Tyr Asp Val Pro Asp Tyr Ala Asn Ser Pro Gly Ile1 5 10 15Pro Gly Met
Gly Pro Lys Lys Lys Arg Lys Val Gly Arg Leu Glu Pro 20 25 30Gly Met
Lys Asn Ile Lys Lys Asn Gln Val Met Asn Leu Gly Pro Asn 35 40 45Ser
Lys Leu Leu Lys Glu Tyr Lys Ser Gln Leu Ile Glu Leu Asn Ile 50 55
60Glu Gln Phe Glu Ala Gly Ile Gly Leu Ile Leu Gly Asp Ala Tyr Ile65
70 75 80Arg Ser Arg Asp Glu Gly Lys Thr Tyr Cys Met Gln Phe Glu Trp
Lys 85 90 95Asn Lys Ala Tyr Met Asp His Val Cys Leu Leu Tyr Asp Gln
Trp Val 100 105 110Leu Ser Pro Pro His Lys Lys Glu Arg Val Asn His
Leu Gly Asn Leu 115 120 125Val Ile Thr Trp Gly Ala Gln Thr Phe Lys
His Gln Ala Phe Asn Lys 130 135 140Leu Ala Asn Leu Phe Ile Val Asn
Asn Lys Lys Thr Ile Pro Asn Asn145 150 155 160Leu Val Glu Asn Tyr
Leu Thr Pro Met Ser Leu Ala Tyr Trp Phe Met 165 170 175Asp Asp Gly
Gly Lys Trp Asp Tyr Asn Lys Asn Ser Thr Asn Lys Ser 180 185 190Ile
Val Leu Asn Thr Gln Ser Phe Thr Phe Glu Glu Val Glu Tyr Leu 195 200
205Val Lys Gly Leu Arg Asn Lys Phe Gln Leu Asn Cys Tyr Val Lys Ile
210 215 220Asn Lys Asn Lys Pro Ile Ile Tyr Ile Asp Ser Met Ser Tyr
Leu Ile225 230 235 240Phe Tyr Asn Leu Ile Lys Pro Tyr Leu Ile Pro
Gln Met Met Tyr Lys 245 250 255Leu Pro Asn Thr Ile Ser Ser Glu Thr
Phe Leu Lys Val 260 2652467DNAArtificial SequencepINT20 Vector
fragment showing Nsi1 site, splice acceptor site and start of
Puromycin resistance gene sequence 24atgcatcttc tgacctcttc
tcttcctccc acagggcatg accgagtaca agcccacggt 60gcgcctc
672510PRTArtificial SequenceInitial fragment of puromycin
resistance protein encoded by pINT20 vector 25Met Thr Glu Tyr Lys
Pro Thr Val Arg Leu1 5 10261125DNAArtificial SequenceSequence of T
cell receptor clone c12/c2 alpha chain construct containing Nhe1,
Not1 and Acc651 restriction sites 26gctagcaagc aggaagtgac
tcagatccca gccgctctga gcgtgcctga gggagaaaac 60ctggtcctga attgcagttt
caccgactca gccatctata acctgcagtg gtttcgccag 120gatccaggca
agggactgac ctccctgctg ctgattcaga gctcccagag ggaacagaca
180tctggcagac tgaatgctag tctggacaaa tctagtggac ggtctaccct
gtacatcgca 240gccagccagc ctggagattc cgcaacatat ctgtgcgccg
tgcgcccact tacaggcgga 300agctacattc ccaccttcgg gcgaggtaca
agcctgatcg tgcacccaga catccagaat 360ccggagcccg ccgtatacca
gctgaaggac cccagaagcc aggacagcac cctgtgcctg 420ttcaccgact
tcgacagcca gatcaacgtg cccaagacaa tggaaagcgg caccttcatc
480accgacaaga ccgtgctgga catgaaggct atggacagca agagcaacgg
cgccattgcc 540tggtccaacc agaccagctt cacatgccag gacatcttca
aagagacaaa cgccacctac 600cccagcagcg acgtgccctg tgatgccacc
ctgaccgaga agtccttcga gacagacatg 660aacctgaact tccagaacct
gtccgcggcc gcaggcctgc tggatcccaa gctgtgctac 720ctgctggacg
ggatcctgtt catctacggt gtgatcctga ctgccctgtt cctgcgagtc
780aaattttctc ggagtgccga cgctcctgca taccagcagg ggcagaacca
gctgtataac 840gagctgaatc tgggtcggag ggaggaatat gacgtgctgg
ataagagacg cggcagggat 900ccagaaatgg ggggcaagcc ccagcgacgg
aaaaaccctc aggagggact gtataatgaa 960ctgcagaagg acaaaatggc
cgaggcttac tctgaaattg ggatgaaggg cgagaggaga 1020cgcggcaaag
gacacgatgg cctgtaccag ggactgagca ctgctaccaa ggacacatat
1080gatgctctgc atatgcaggc actgccccct agataataag gtacc
112527369PRTArtificial SequenceSequence of T cell receptor clone
c12/c2 alpha chain 27Lys Gln Glu Val Thr Gln Ile Pro Ala Ala Leu
Ser Val Pro Glu Gly1 5 10 15Glu Asn Leu Val Leu Asn Cys Ser Phe Thr
Asp Ser Ala Ile Tyr Asn 20 25 30Leu Gln Trp Phe Arg Gln Asp Pro
Gly Lys Gly Leu Thr Ser Leu Leu 35 40 45Leu Ile Gln Ser Ser Gln Arg
Glu Gln Thr Ser Gly Arg Leu Asn Ala 50 55 60Ser Leu Asp Lys Ser Ser
Gly Arg Ser Thr Leu Tyr Ile Ala Ala Ser65 70 75 80Gln Pro Gly Asp
Ser Ala Thr Tyr Leu Cys Ala Val Arg Pro Leu Thr 85 90 95Gly Gly Ser
Tyr Ile Pro Thr Phe Gly Arg Gly Thr Ser Leu Ile Val 100 105 110His
Pro Asp Ile Gln Asn Pro Glu Pro Ala Val Tyr Gln Leu Lys Asp 115 120
125Pro Arg Ser Gln Asp Ser Thr Leu Cys Leu Phe Thr Asp Phe Asp Ser
130 135 140Gln Ile Asn Val Pro Lys Thr Met Glu Ser Gly Thr Phe Ile
Thr Asp145 150 155 160Lys Thr Val Leu Asp Met Lys Ala Met Asp Ser
Lys Ser Asn Gly Ala 165 170 175Ile Ala Trp Ser Asn Gln Thr Ser Phe
Thr Cys Gln Asp Ile Phe Lys 180 185 190Glu Thr Asn Ala Thr Tyr Pro
Ser Ser Asp Val Pro Cys Asp Ala Thr 195 200 205Leu Thr Glu Lys Ser
Phe Glu Thr Asp Met Asn Leu Asn Phe Gln Asn 210 215 220Leu Ser Ala
Ala Ala Gly Leu Leu Asp Pro Lys Leu Cys Tyr Leu Leu225 230 235
240Asp Gly Ile Leu Phe Ile Tyr Gly Val Ile Leu Thr Ala Leu Phe Leu
245 250 255Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln
Gln Gly 260 265 270Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg
Arg Glu Glu Tyr 275 280 285Asp Val Leu Asp Lys Arg Arg Gly Arg Asp
Pro Glu Met Gly Gly Lys 290 295 300Pro Gln Arg Arg Lys Asn Pro Gln
Glu Gly Leu Tyr Asn Glu Leu Gln305 310 315 320Lys Asp Lys Met Ala
Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu 325 330 335Arg Arg Arg
Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr 340 345 350Ala
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro 355 360
365Arg281226DNAArtificial SequenceSequence of T cell receptor clone
c12/c2 beta chain construct containing Nco1, Xho1 and Hind3
restriction sites 28ccatggccaa cgctggagtg actcagaccc ctaagttcca
ggtcctgaaa actgggcaga 60gtatgaccct gcagtgcgca caggacatga atcacgagta
catgtcatgg tatcggcagg 120atccagggat gggtctgagg ctgatccatt
acagcgtggg cgctggaact accgaccagg 180gcgaggtgcc caacggatat
aatgtctcaa gaagcaccac agaagatttc ccactgcgac 240tgctgagcgc
cgctcctagc cagacatccg tgtacttttg cgccagctcc aatgtcggga
300acaccggcga gctgttcttt ggggaaggtt cccgcctgac agtgctcgag
gacctgagaa 360acgtgacccc ccccaaggtg tccctgttcg agcctagcaa
ggccgagatc gccaacaagc 420agaaagccac cctcgtgtgc ctggccagag
gcttcttccc cgaccacgtg gaactgtctt 480ggtgggtcaa cggcaaagag
gtgcacagcg gcgtgtccac cgatccccag gcctacaaag 540agagcaacta
cagctactgc ctgagcagca gactgcgggt gtccgccacc ttctggcaca
600acccccggaa ccacttcaga tgccaggtgc agtttcacgg cctgagcgaa
gaggacaagt 660ggcccgaggg cagccctaag cccgtgaccc agaatatctc
tgccgaagcc tggggcagag 720ccgactgtgg cattaccagc gccagctacc
agcagggcgt gctgtctgcc accatcctgt 780acgaggtcgc gagcggactg
ctggacccaa agctgtgcta cctgctggat gggatcctgt 840tcatctacgg
tgtgattctg acagccctgt tcctgcgagt caagttcagc cggagcgccg
900acgcaccagc ataccagcag gggcagaatc agctgtataa cgagctgaat
ctgggtcgga 960gggaggaata cgacgtgctg gataagagac gcggcaggga
tcccgaaatg ggcggaaagc 1020ctcagcgacg gaaaaaccca caggagggac
tgtacaatga actgcagaag gacaaaatgg 1080ctgaggcata ttctgaaatc
ggcatgaagg gagagaggag acgcggcaaa ggacacgatg 1140ggctgtacca
gggtctgagt acagccacta aggacaccta tgatgccctg catatgcagg
1200ctctgccacc cagataataa aagctt 122629404PRTArtificial
SequenceSequence of T cell receptor clone c12/c2 beta chain
construct 29Met Ala Asn Ala Gly Val Thr Gln Thr Pro Lys Phe Gln Val
Leu Lys1 5 10 15Thr Gly Gln Ser Met Thr Leu Gln Cys Ala Gln Asp Met
Asn His Glu 20 25 30Tyr Met Ser Trp Tyr Arg Gln Asp Pro Gly Met Gly
Leu Arg Leu Ile 35 40 45His Tyr Ser Val Gly Ala Gly Thr Thr Asp Gln
Gly Glu Val Pro Asn 50 55 60Gly Tyr Asn Val Ser Arg Ser Thr Thr Glu
Asp Phe Pro Leu Arg Leu65 70 75 80Leu Ser Ala Ala Pro Ser Gln Thr
Ser Val Tyr Phe Cys Ala Ser Ser 85 90 95Asn Val Gly Asn Thr Gly Glu
Leu Phe Phe Gly Glu Gly Ser Arg Leu 100 105 110Thr Val Leu Glu Asp
Leu Arg Asn Val Thr Pro Pro Lys Val Ser Leu 115 120 125Phe Glu Pro
Ser Lys Ala Glu Ile Ala Asn Lys Gln Lys Ala Thr Leu 130 135 140Val
Cys Leu Ala Arg Gly Phe Phe Pro Asp His Val Glu Leu Ser Trp145 150
155 160Trp Val Asn Gly Lys Glu Val His Ser Gly Val Ser Thr Asp Pro
Gln 165 170 175Ala Tyr Lys Glu Ser Asn Tyr Ser Tyr Cys Leu Ser Ser
Arg Leu Arg 180 185 190Val Ser Ala Thr Phe Trp His Asn Pro Arg Asn
His Phe Arg Cys Gln 195 200 205Val Gln Phe His Gly Leu Ser Glu Glu
Asp Lys Trp Pro Glu Gly Ser 210 215 220Pro Lys Pro Val Thr Gln Asn
Ile Ser Ala Glu Ala Trp Gly Arg Ala225 230 235 240Asp Cys Gly Ile
Thr Ser Ala Ser Tyr Gln Gln Gly Val Leu Ser Ala 245 250 255Thr Ile
Leu Tyr Glu Val Ala Ser Gly Leu Leu Asp Pro Lys Leu Cys 260 265
270Tyr Leu Leu Asp Gly Ile Leu Phe Ile Tyr Gly Val Ile Leu Thr Ala
275 280 285Leu Phe Leu Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro
Ala Tyr 290 295 300Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn
Leu Gly Arg Arg305 310 315 320Glu Glu Tyr Asp Val Leu Asp Lys Arg
Arg Gly Arg Asp Pro Glu Met 325 330 335Gly Gly Lys Pro Gln Arg Arg
Lys Asn Pro Gln Glu Gly Leu Tyr Asn 340 345 350Glu Leu Gln Lys Asp
Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met 355 360 365Lys Gly Glu
Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly 370 375 380Leu
Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala385 390
395 400Leu Pro Pro Arg30677DNAArtificial SequenceSequence of T cell
receptor clone 4JFH alpha chain construct with initial Nhe1 and
terminal Not1 restriction sites 30gctagccaga aagaggtgga acagaacagc
ggccctctga gcgtgccaga aggcgctatc 60gccagcctga actgcaccta cagctttctg
ggcagccaga gcttcttctg gtacagacag 120tacagcggca agagccccga
gctgatcatg ttcacctaca gagagggcga caaagaggac 180ggcagattca
ccgcccagct gaacaaggcc agccagcacg tgtccctgct gatcagagac
240agccagccta gcgacagcgc cacctacctg tgcgccgtga atgatggcgg
cagactgacc 300tttggcgacg gcaccaccct gaccgtgaag cctgacatcc
agaatccgga gcccgccgta 360taccagctga aggaccccag aagccaggac
agcaccctgt gcctgttcac cgacttcgac 420agccagatca acgtgcccaa
gacaatggaa agcggcacct tcatcaccga caagaccgtg 480ctggacatga
aggctatgga cagcaagagc aacggcgcca ttgcctggtc caaccagacc
540agcttcacat gccaggacat cttcaaagag acaaacgcca cctaccccag
cagcgacgtg 600ccctgtgatg ccaccctgac cgagaagtcc ttcgagacag
acatgaacct gaacttccag 660aacctgtccg cggccgc 67731221PRTArtificial
SequenceSequence of T cell receptor clone 4JFH alpha chain
construct 31Gln Lys Glu Val Glu Gln Asn Ser Gly Pro Leu Ser Val Pro
Glu Gly1 5 10 15Ala Ile Ala Ser Leu Asn Cys Thr Tyr Ser Phe Leu Gly
Ser Gln Ser 20 25 30Phe Phe Trp Tyr Arg Gln Tyr Ser Gly Lys Ser Pro
Glu Leu Ile Met 35 40 45Phe Thr Tyr Arg Glu Gly Asp Lys Glu Asp Gly
Arg Phe Thr Ala Gln 50 55 60Leu Asn Lys Ala Ser Gln His Val Ser Leu
Leu Ile Arg Asp Ser Gln65 70 75 80Pro Ser Asp Ser Ala Thr Tyr Leu
Cys Ala Val Asn Asp Gly Gly Arg 85 90 95Leu Thr Phe Gly Asp Gly Thr
Thr Leu Thr Val Lys Pro Asp Ile Gln 100 105 110Asn Pro Glu Pro Ala
Val Tyr Gln Leu Lys Asp Pro Arg Ser Gln Asp 115 120 125Ser Thr Leu
Cys Leu Phe Thr Asp Phe Asp Ser Gln Ile Asn Val Pro 130 135 140Lys
Thr Met Glu Ser Gly Thr Phe Ile Thr Asp Lys Thr Val Leu Asp145 150
155 160Met Lys Ala Met Asp Ser Lys Ser Asn Gly Ala Ile Ala Trp Ser
Asn 165 170 175Gln Thr Ser Phe Thr Cys Gln Asp Ile Phe Lys Glu Thr
Asn Ala Thr 180 185 190Tyr Pro Ser Ser Asp Val Pro Cys Asp Ala Thr
Leu Thr Glu Lys Ser 195 200 205Phe Glu Thr Asp Met Asn Leu Asn Phe
Gln Asn Leu Ser 210 215 22032353DNAArtificial SequenceSequence of T
cell receptor clone4JFH beta chain construct with initial Nco1
restriction site and terminal Xho1 restriction site 32ccatggccag
ccagaccatc catcagtggc ctgccaccct ggtgcagcct gtgggatctc 60ctctgagcct
ggaatgcacc gtggaaggca ccagcaaccc caacctgtac tggtacagac
120aggccgctgg cagaggcccc cagctgctgt tttactgggg cccctttggc
cagatcagca 180gcgaggtgcc ccagaacctg agcgccagca gaccccagga
ccggcagttt atcctgagca 240gcaagaagct gctgctgagc gacagcggct
tctacctgtg cgcttggagc gagacaggcc 300tgggcatggg cggatggcag
tttggcgagg gcagcagact gacagtgctc gag 35333117PRTArtificial
SequenceSequence of T cell receptor clone 4JFH beta chain construct
33Met Ala Ser Gln Thr Ile His Gln Trp Pro Ala Thr Leu Val Gln Pro1
5 10 15Val Gly Ser Pro Leu Ser Leu Glu Cys Thr Val Glu Gly Thr Ser
Asn 20 25 30Pro Asn Leu Tyr Trp Tyr Arg Gln Ala Ala Gly Arg Gly Pro
Gln Leu 35 40 45Leu Phe Tyr Trp Gly Pro Phe Gly Gln Ile Ser Ser Glu
Val Pro Gln 50 55 60Asn Leu Ser Ala Ser Arg Pro Gln Asp Arg Gln Phe
Ile Leu Ser Ser65 70 75 80Lys Lys Leu Leu Leu Ser Asp Ser Gly Phe
Tyr Leu Cys Ala Trp Ser 85 90 95Glu Thr Gly Leu Gly Met Gly Gly Trp
Gln Phe Gly Glu Gly Ser Arg 100 105 110Leu Thr Val Leu Glu
1153453DNAArtificial SequencePrimer for mutation of CDR3 of c12/c2
TCR alpha chainmisc_feature(20)..(21)n is a, c, g, or
tmisc_feature(23)..(24)n is a, c, g, or tmisc_feature(31)..(31)n is
a, c, g, or t 34cacgcggcac gcgggtgaan nsnnscctwc natgtaaggg
tggaagcccg ctc 533552DNAArtificial SequencePrimer used to mutate
CDR3 of c12/c2 TCR alpha chainmisc_feature(23)..(23)n is a, c, g,
or tmisc_feature(30)..(31)n is a, c, g, or tmisc_feature(33)..(34)n
is a, c, g, or t 35ctcgcccgaa ggtgggaatg tangwtccsn nsnnaagtgg
gcgcacggcg ca 523617PRTArtificial SequenceStrategy used to mutate
CDR3 of c12/c2 TCR alpha chainmisc_feature(7)..(8)Xaa can be any
naturally occurring amino acid 36Cys Ala Val Arg Pro Leu Xaa Xaa
Gly Ser Tyr Ile Pro Thr Phe Gly1 5 10 15Arg3717PRTArtificial
SequenceStrategy used to mutate CDR3 of c12/c2 TCR alpha
chainmisc_feature(7)..(8)Xaa can be any naturally occurring amino
acid 37Cys Ala Val Arg Pro Leu Xaa Xaa Gly Thr Tyr Ile Pro Thr Phe
Gly1 5 10 15Arg3850DNAArtificial SequencePrimer used to mutate CDR3
of c12/c2 TCR beta chainmisc_feature(20)..(21)n is a, c, g, or
tmisc_feature(29)..(30)n is a, c, g, or t 38gtacttttgc gccagctccn
nsstcgggnn saccggcgag ctgttctttg 503949DNAArtificial SequencePrimer
used to mutate CDR3 of c12/c2 TCR beta chainmisc_feature(21)..(22)n
is a, c, g, or tmisc_feature(30)..(31)n is a, c, g, or t
39caaagaacag ctcgccggts nncccgassn nggagctggc gcaaaagta
494016PRTArtificial SequenceStrategy used to mutate CDR3 of c12/c2
TCR beta chain 40Tyr Phe Cys Ala Ser Ser Asn Val Gly Asn Thr Gly
Glu Leu Phe Phe1 5 10 154116PRTArtificial SequenceStrategy used to
mutate CDR3 of c12/c2 TCR beta chain 41Tyr Phe Cys Ala Ser Ser Asn
Leu Gly Asn Thr Gly Glu Leu Phe Phe1 5 10 1542462DNAArtificial
SequenceSequence of CD3 zeta in pINT21 CAR1 42ggcgccatgg cccaggtcgc
ggccgcaagc ggactgctgg acccaaagct gtgctacctg 60ctggatggga tcctgttcat
ctacggtgtg attctgacag ccctgttcct gcgagtcaag 120ttcagccgga
gcgccgacgc accagcatac cagcaggggc agaatcagct gtataacgag
180ctgaatctgg gtcggaggga ggaatacgac gtgctggata agagacgcgg
cagggatccc 240gaaatgggcg gaaagcctca gcgacggaaa aacccacagg
agggactgta caatgaactg 300cagaaggaca aaatggctga ggcatattct
gaaatcggca tgaagggaga gaggagacgc 360ggcaaaggac acgatgggct
gtaccagggt ctgagtacag ccactaagga cacctatgat 420gccctgcata
tgcaggctct gccacccaga taataaaagc tt 462434PRTArtificial
SequenceFragment encoded by pINT21 CAR1 vector 43Gly Ala Met
Ala144141PRTArtificial SequenceCDR3 zeta sequence encoded by pINT21
CAR1 44Ser Gly Leu Leu Asp Pro Lys Leu Cys Tyr Leu Leu Asp Gly Ile
Leu1 5 10 15Phe Ile Tyr Gly Val Ile Leu Thr Ala Leu Phe Leu Arg Val
Lys Phe 20 25 30Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln
Asn Gln Leu 35 40 45Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
Asp Val Leu Asp 50 55 60Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly
Lys Pro Gln Arg Arg65 70 75 80Lys Asn Pro Gln Glu Gly Leu Tyr Asn
Glu Leu Gln Lys Asp Lys Met 85 90 95Ala Glu Ala Tyr Ser Glu Ile Gly
Met Lys Gly Glu Arg Arg Arg Gly 100 105 110Lys Gly His Asp Gly Leu
Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp 115 120 125Thr Tyr Asp Ala
Leu His Met Gln Ala Leu Pro Pro Arg 130 135 14045705DNAArtificial
SequencepINT21 CAR2 Sequence encoding CD3 zeta 45gccatggccc
aggtcgcggc cgcaacaaca accccagccc ccagacctcc tacccctgcc 60cctacaattg
ccagccagcc tctgagcctg aggcccgagg cttgtagacc agctgctggc
120ggagccgtgc acaccagagg actggatttc gcctgcgaca tctacatctg
ggcccctctg 180gccggcacat gtggcgtgct gctgctgagc ctcgtgatca
ccctgtactg caagcggggc 240agaaagaaac tgctgtacat ctttaagcag
cccttcatgc ggcccgtgca gaccacccag 300gaagaggacg gctgctcctg
cagattcccc gaggaagaag aaggcggctg cgagctgaga 360gtgaagttca
gcagatccgc cgacgcccct gcctacaagc agggccagaa ccagctgtac
420aacgagctga acctgggcag acgggaagag tacgacgtgc tggacaagcg
gagaggccgg 480gacccagaga tgggcggaaa gcccagaaga aagaaccccc
aggaaggcct gtataacgaa 540ctgcagaaag acaaaatggc cgaggcctac
agcgagatcg gaatgaaggg cgagcggaga 600agaggcaagg ggcacgatgg
cctgtaccag ggcctgagca ccgccaccaa ggacacctat 660gacgccctgc
acatgcaggc cctgccccct agataataaa agctt 70546223PRTArtificial
SequenceSequence encoded by pINT21 CAR2 CD8 hinge / transmembrane
region, 4-1BB and CD3 zeta 46Thr Thr Thr Pro Ala Pro Arg Pro Pro
Thr Pro Ala Pro Thr Ile Ala1 5 10 15Ser Gln Pro Leu Ser Leu Arg Pro
Glu Ala Cys Arg Pro Ala Ala Gly 20 25 30Gly Ala Val His Thr Arg Gly
Leu Asp Phe Ala Cys Asp Ile Tyr Ile 35 40 45Trp Ala Pro Leu Ala Gly
Thr Cys Gly Val Leu Leu Leu Ser Leu Val 50 55 60Ile Thr Leu Tyr Cys
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe65 70 75 80Lys Gln Pro
Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly 85 90 95Cys Ser
Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg 100 105
110Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln
115 120 125Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu
Tyr Asp 130 135 140Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met
Gly Gly Lys Pro145 150 155 160Arg Arg Lys Asn Pro Gln Glu Gly Leu
Tyr Asn Glu Leu Gln Lys Asp 165 170 175Lys Met Ala Glu Ala Tyr Ser
Glu Ile Gly Met Lys Gly Glu Arg Arg 180 185 190Arg Gly Lys Gly His
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr 195 200 205Lys Asp Thr
Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg 210 215
22047750DNAArtificial SequenceSequence of FMC63
H-L (anti-CD19 antibody) 47gccatggccg aagtgaaact gcaggagtct
ggacccggcc tggtggcccc atctcagtct 60ctgagcgtga cctgtaccgt gtccggcgtg
tccctgcctg actatggcgt gtcctggatc 120agacagcccc ccagaaaggg
cctggaatgg ctgggagtga tctggggcag cgaaaccacc 180tactacaaca
gcgccctgaa gtcccggctg accatcatca aggacaactc caagagccag
240gtgttcctga agatgaacag cctgcagacc gacgacaccg ccatctacta
ctgcgccaag 300cactactact acggcggcag ctacgctatg gactactggg
gccagggcac ctcggtcacc 360gtctcgagtg gtggaggcgg ttcaggcgga
ggtggctctg gcggtggcgc tagcgacatc 420cagatgaccc agaccaccag
cagcctgagc gccagcctgg gcgatagagt gaccatcagc 480tgcagagcca
gccaggacat cagcaagtac ctgaactggt atcagcagaa acccgacggc
540accgtgaagc tgctgatcta ccacaccagc agactgcaca gcggcgtgcc
cagcagattt 600tccggctctg gcagcggcac cgactacagc ctgaccatct
ccaacctgga acaggaagat 660atcgctacct acttctgtca gcaaggcaac
accctgccct acaccttcgg cggagggacc 720aagctggaga tcaaacgtac
cgcggccgca 75048250PRTArtificial SequenceSequence of FMC63 H-L
(anti-CD19 antibody) 48Ala Met Ala Glu Val Lys Leu Gln Glu Ser Gly
Pro Gly Leu Val Ala1 5 10 15Pro Ser Gln Ser Leu Ser Val Thr Cys Thr
Val Ser Gly Val Ser Leu 20 25 30Pro Asp Tyr Gly Val Ser Trp Ile Arg
Gln Pro Pro Arg Lys Gly Leu 35 40 45Glu Trp Leu Gly Val Ile Trp Gly
Ser Glu Thr Thr Tyr Tyr Asn Ser 50 55 60Ala Leu Lys Ser Arg Leu Thr
Ile Ile Lys Asp Asn Ser Lys Ser Gln65 70 75 80Val Phe Leu Lys Met
Asn Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr 85 90 95Tyr Cys Ala Lys
His Tyr Tyr Tyr Gly Gly Ser Tyr Ala Met Asp Tyr 100 105 110Trp Gly
Gln Gly Thr Ser Val Thr Val Ser Ser Gly Gly Gly Gly Ser 115 120
125Gly Gly Gly Gly Ser Gly Gly Gly Ala Ser Asp Ile Gln Met Thr Gln
130 135 140Thr Thr Ser Ser Leu Ser Ala Ser Leu Gly Asp Arg Val Thr
Ile Ser145 150 155 160Cys Arg Ala Ser Gln Asp Ile Ser Lys Tyr Leu
Asn Trp Tyr Gln Gln 165 170 175Lys Pro Asp Gly Thr Val Lys Leu Leu
Ile Tyr His Thr Ser Arg Leu 180 185 190His Ser Gly Val Pro Ser Arg
Phe Ser Gly Ser Gly Ser Gly Thr Asp 195 200 205Tyr Ser Leu Thr Ile
Ser Asn Leu Glu Gln Glu Asp Ile Ala Thr Tyr 210 215 220Phe Cys Gln
Gln Gly Asn Thr Leu Pro Tyr Thr Phe Gly Gly Gly Thr225 230 235
240Lys Leu Glu Ile Lys Arg Thr Ala Ala Ala 245
25049327DNAArtificial SequenceSequence of lox1 binding Adhiron
lox1A 49gccatggctg ctacaggcgt gcgggctgtg cccggcaatg agaacagcct
ggaaatcgag 60gaactggcca gattcgccgt ggacgagcac aacaagaaag agaacgccct
gctggaattc 120gtgcgggtcg tgaaggccaa agagcagtgg agcgaggccg
acaacgactg gcacaccatg 180tactacctga ccctggaagc caaggacggc
ggcaagaaga agctgtacga ggccaaagtg 240tgggtcaagc tggacctgga
aacctggcag cacttcaact tcaaagagct ccaggaattc 300aagcccgtgg
gcgacgctgc ggccgcg 32750109PRTArtificial SequenceSequence of Lox1
binding Adhiron lox1A 50Ala Met Ala Ala Thr Gly Val Arg Ala Val Pro
Gly Asn Glu Asn Ser1 5 10 15Leu Glu Ile Glu Glu Leu Ala Arg Phe Ala
Val Asp Glu His Asn Lys 20 25 30Lys Glu Asn Ala Leu Leu Glu Phe Val
Arg Val Val Lys Ala Lys Glu 35 40 45Gln Trp Ser Glu Ala Asp Asn Asp
Trp His Thr Met Tyr Tyr Leu Thr 50 55 60Leu Glu Ala Lys Asp Gly Gly
Lys Lys Lys Leu Tyr Glu Ala Lys Val65 70 75 80Trp Val Lys Leu Asp
Leu Glu Thr Trp Gln His Phe Asn Phe Lys Glu 85 90 95Leu Gln Glu Phe
Lys Pro Val Gly Asp Ala Ala Ala Ala 100 10551333DNAArtificial
SequenceSequence of lox1 binding Adhiron lox1B 51gccatggctg
ctacaggcgt gcgggctgtg cccggcaatg agaacagcct ggaaatcgag 60gaactggcca
gattcgccgt ggacgagcac aacaagaaag agaacgccct gctggaattc
120gtgcgggtcg tgaaggccaa agagcaggaa cagcccatcg gcgagcaccc
cgtgaacgac 180accatgtact acctgaccct ggaagccaag gacggcggca
agaagaagct gtacgaggcc 240aaagtgtggg tcaagcggtg gctgcggttc
accgagatct acaacttcaa agagctccag 300gaattcaagc ccgtgggcga
cgctgcggcc gcg 33352111PRTArtificial SequenceSequence of lox1
binding Adhiron lox1B 52Ala Met Ala Ala Thr Gly Val Arg Ala Val Pro
Gly Asn Glu Asn Ser1 5 10 15Leu Glu Ile Glu Glu Leu Ala Arg Phe Ala
Val Asp Glu His Asn Lys 20 25 30Lys Glu Asn Ala Leu Leu Glu Phe Val
Arg Val Val Lys Ala Lys Glu 35 40 45Gln Glu Gln Pro Ile Gly Glu His
Pro Val Asn Asp Thr Met Tyr Tyr 50 55 60Leu Thr Leu Glu Ala Lys Asp
Gly Gly Lys Lys Lys Leu Tyr Glu Ala65 70 75 80Lys Val Trp Val Lys
Arg Trp Leu Arg Phe Thr Glu Ile Tyr Asn Phe 85 90 95Lys Glu Leu Gln
Glu Phe Lys Pro Val Gly Asp Ala Ala Ala Ala 100 105
1105354DNAArtificial SequenceAdhiron_mut1
primermisc_feature(23)..(24)n is a, c, g, or
tmisc_feature(26)..(27)n is a, c, g, or tmisc_feature(29)..(30)n is
a, c, g, or tmisc_feature(32)..(33)n is a, c, g, or t 53cagggtcagg
tagtacatgg tsnnsnnsnn snnctgctct ttggccttca cgac
545448DNAArtificial SequenceAdhiron_mut2
primermisc_feature(20)..(21)n is a, c, g, or
tmisc_feature(23)..(24)n is a, c, g, or tmisc_feature(26)..(27)n is
a, c, g, or tmisc_feature(29)..(30)n is a, c, g, or t 54ctggagctct
ttgaagttsn nsnnsnnsnn cttgacccac actttggc 485554DNAArtificial
SequenceMutagenic primer for construction of library of binders
within loop 1 (adhiron mut 1)misc_feature(22)..(23)n is a, c, g, or
tmisc_feature(25)..(26)n is a, c, g, or tmisc_feature(28)..(29)n is
a, c, g, or tmisc_feature(31)..(32)n is a, c, g, or t 55gtcgtgaagg
ccaaagagca gnnsnnsnns nnsaccatgt actacctgac cctg
545618PRTArtificial SequenceAdhiron mut1 loop library variant
sequencemisc_feature(8)..(11)Xaa can be any naturally occurring
amino acid 56Val Val Lys Ala Lys Glu Gln Xaa Xaa Xaa Xaa Thr Met
Tyr Tyr Leu1 5 10 15Thr Leu5748DNAArtificial SequenceMutagenic
primer for construction of library of binders within Adhiron mut2
loopmisc_feature(19)..(20)n is a, c, g, or tmisc_feature(22)..(23)n
is a, c, g, or tmisc_feature(25)..(26)n is a, c, g, or
tmisc_feature(28)..(29)n is a, c, g, or t 57gccaaagtgt gggtcaagnn
snnsnnsnns aacttcaaag agctccag 485816PRTArtificial SequenceAdhiron
mut 2 loop library variant sequencemisc_feature(7)..(10)Xaa can be
any naturally occurring amino acid 58Ala Lys Val Trp Val Lys Xaa
Xaa Xaa Xaa Asn Phe Lys Glu Leu Gln1 5 10 1559105DNAArtificial
sequenceTrypsin binding Knottin MCoTI-IIsequence 59gccatggccg
gtgtgtgccc caagatcttg aaaaagtgcc gccgtgacag cgattgtccc 60ggcgcctgca
tctgccgcgg caatggctat tgcggagcgg ccgca 1056032PRTArtificial
SequenceTrypsin binding Knottin MCo-TI-II sequence 60Ala Met Ala
Gly Val Cys Pro Lys Ile Leu Lys Lys Cys Arg Arg Asp1 5 10 15Ser Asp
Cys Pro Gly Ala Cys Ile Cys Arg Gly Asn Gly Tyr Cys Gly 20 25
306199DNAArtificial SequencePrimer for creation of library of
Knottin mutantsmisc_feature(11)..(11)n is a, c, g, or
tmisc_feature(14)..(14)n is a, c, g, or tmisc_feature(17)..(17)n is
a, c, g, or tmisc_feature(20)..(20)n is a, c, g, or
tmisc_feature(23)..(23)n is a, c, g, or tmisc_feature(26)..(26)n is
a, c, g, or tmisc_feature(29)..(29)n is a, c, g, or
tmisc_feature(32)..(32)n is a, c, g, or tmisc_feature(35)..(35)n is
a, c, g, or tmisc_feature(38)..(38)n is a, c, g, or t 61ggtgtgtgcv
nsvnsvnsvn svnsvnsvns vnsvnsvnst gccgccgtga cagcgattgt 60cccggcgcct
gcatctgccg cggcaatggc tattgcgga 996299DNAArtificial
SequenceSequence varied in library of knottin
mutantsmisc_feature(62)..(62)n is a, c, g, or
tmisc_feature(65)..(65)n is a, c, g, or tmisc_feature(68)..(68)n is
a, c, g, or tmisc_feature(71)..(71)n is a, c, g, or
tmisc_feature(74)..(74)n is a, c, g, or tmisc_feature(77)..(77)n is
a, c, g, or tmisc_feature(80)..(80)n is a, c, g, or
tmisc_feature(83)..(83)n is a, c, g, or tmisc_feature(86)..(86)n is
a, c, g, or tmisc_feature(89)..(89)n is a, c, g, or t 62tccgcaatag
ccattgccgc ggcagatgca ggcgccggga caatcgctgt cacggcggca 60snbsnbsnbs
nbsnbsnbsn bsnbsnbsnb gcacacacc 996333PRTArtificial
SequenceSequence of Knottin varied in
librarymisc_feature(4)..(13)Xaa can be any naturally occurring
amino acid 63Gly Val Cys Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa
Cys Arg Arg1 5 10 15Asp Ser Asp Cys Pro Gly Ala Cys Ile Cys Arg Gly
Asn Gly Tyr Cys 20 25 30Gly64121DNAArtificial SequencepD7-Sce1
sequence nucleotides 1 - 120 64ggtaccgaat tctagggata acagggtaat
atgcatcttc tgacctcttc tcttcctccc 60acagggcatg gcaaaacctc tgagccagga
agaaagcaca ctgattgaaa gagcaaccgc 120t 1216518PRTArtificial
SequenceFragment of blasticidin resistance gene encoded by pD7-Sce1
65Met Ala Lys Pro Leu Ser Gln Glu Glu Ser Thr Leu Ile Glu Arg Ala1
5 10 15Thr Ala66120DNAArtificial SequencepD7-ObLiGaRe sequence
fragment 66ggtaccgaat tcttttctgt caccaatcct ggggccacta gggacactgt
ggggtggagg 60aaatgcatct tctgacctct tctcttcctc ccacagggca tggcaaaacc
tctgagccag 120677PRTArtificial SequenceFragment of Blasticidin
resistance gene encoded by pD7-ObLiGaRe 67Met Ala Lys Pro Leu Ser
Gln1 5681061DNAArtificial SequenceROSA 26 left homology arm
sequence 68gcgatcgcgc tgattggctt cttttcctcc cgccgtgtgt gaaaacacaa
atggcgtgtt 60ttggttggcg taaggcgcct gtcagttaac ggcagccgga gtgcgcagcc
gccggcagcc 120tcgctctgcc cactgggtgg ggcgggaggt aggtggggtg
aggcgagctg gacgtgcggg 180cgcggtcggc ctctggcggg gcgggggagg
ggagggaggg tcagcgaaag tagctcgcgc 240gcgagcggcc gcccaccctc
cccttcctct gggggagtcg ttttacccgc cgccggccgg 300gcctcgtcgt
ctgattggct ctcggggccc agaaaactgg cccttgccat tggctcgtgt
360tcgtgcaagt tgagtccatc cgccggccag cgggggcggc gaggaggcgc
tcccaggttc 420cggccctccc ctcggccccg cgccgcagag tctggccgcg
cgcccctgcg caacgtggca 480ggaagcgcgc gctgggggcg gggacgggca
gtagggctga gcggctgcgg ggcgggtgca 540agcacgtttc cgacttgagt
tgcctcaaga ggggcgtgct gagccagacc tccatcgcgc 600actccgggga
gtggagggaa ggagcgaggg ctcagttggg ctgttttgga ggcaggaagc
660acttgctctc ccaaagtcgc tctgagttgt tatcagtaag ggagctgcag
tggagtaggc 720ggggagaagg ccgcaccctt ctccggaggg gggaggggag
tgttgcaata cctttctggg 780agttctctgc tgcctcctgg cttctgagga
ccgccctggg cctgggagaa tcccttcccc 840ctcttccctc gtgatctgca
actccagtct ttctagaatg cattaaggga tctgtagggc 900gcagtagtcc
agggtttcct tgatgatgtc atacttatcc tgtccctttt ttttccacag
960ctcgcggttg aggacaaact cttcgcggtc tttccagtgg ggatcgacgg
tatcgtagag 1020tcgaggccgc tctaggaatt cacgccgcca ccatgaccga g
106169703DNAArtificial SequenceROSA 26 right homology arm sequence
69gtatacggga attgaacagg tgtaaaattg gagggacaag acttcccaca gattttcggt
60tttgtcggga agttttttaa taggggcaaa taaggaaaat gggaggatag gtagtcatct
120ggggttttat gcagcaaaac tacaggttat tattgcttgt gatccgcctc
ggagtatttt 180ccatcgaggt agattaaaga catgctcacc cgagttttat
actctcctgc ttgagatcct 240tactacagta tgaaattaca gtgtcgcgag
ttagactatg taagcagaat tttaatcatt 300tttaaagagc ccagtacttc
atatccattt ctcccgctcc ttctgcagcc ttatcaaaag 360gtattttaga
acactcattt tagccccatt ttcatttatt atactggctt atccaacccc
420tagacagagc attggcattt tccctttcct gatcttagaa gtctgatgac
tcatgaaacc 480agacagatta gttacataca ccacaaatcg aggctgtagc
tggggcctca acactgcagt 540tcttttataa ctccttagta cactttttgt
tgatcctttg ccttgatcct taattttcag 600tgtctatcac ctctcccgtc
aggtggtgtt ccacatttgg gcctattctc agtccaggga 660gttttacaac
aatagatgta ttgagaatcc aacctcctgc agg 7037018DNAArtificial
SequenceLeft TALEN recognition sequence 70tcccctccac cccacagt
187115DNAArtificial SequenceTALEN spacer 71ggggccacta gggac
157218DNAArtificial SequenceRight TALEN target sequence
72aggattggtg acagaaaa 187318DNAArtificial SequenceRight TALEN
sequence 73ttttctgtca ccaatcct 187420DNAArtificial
SequenceAAVS-Left-arm-junction-PCR-Forw (9625) primer 74ccggaactct
gccctctaac 207523DNAArtificial SequenceBSD_Junction PCR-rev (9626)
primer 75tagccacaga atagtcttcg gag 237621DNAArtificial
SequenceDonor_plasmid_seq_PDGFRTM-2 Forw primer 76acacgcagga
ggccatcgtg g 217720DNAArtificial SequenceAAVS1_right
arm_junction_PCR_rev primer 77tcctgggata ccccgaagag
207854DNAArtificial SequencePrimer 2598 78ttttttttaa ttaagattat
tgactagtta ttaatagtaa tcaattacgg ggtc 547952DNAArtificial
SequencePrimer 2619 79ttttttgttt aaacttacct tggatccctt gccggggctc
aggctcaggg ac 528033DNAArtificial SequenceM13Leadseq primer
80aaattattat tcgcaattcc tttggttgtt cct 338130DNAArtificial
SequenceNotmycseq primer 81ggccccattc agatcctctt ctgagatgag
308222DNAArtificial SequencePrimer 2623 82taaagtaggc ggtcttgaga cg
228322DNAArtificial SequencePrimer 2624 83gaaggtgctg ttgaactgtt cc
228419DNAArtificial SequencePrimer 2595 84gagggctctg gcagctagc
198520DNAArtificial SequencePrimer 2597 85tcgagactgt gacgaggctg
208619DNAArtificial SequencePrimer 2625 86ccttggtgct ggcactcga
198740DNAArtificial SequencePrimer 1999 87aaaaagcagg ctaccatgag
ggcctggatc ttctttctcc 408844DNAArtificial SequenceTALEN recognition
sequence 88tccaccggtc gccaccatgg tgagcaaggg cgaggagctg ttca
448933DNAArtificial SequenceAAVS1_HA-L_Nested_Forw1 primer
89gtgcccttgc tgtgccgccg gaactctgcc ctc 339023DNAArtificial
SequenceEGFP_Synthetic_gene_Rev_Assembly primer 90ttcacgtcgc
cgtccagctc gac 239121DNAArtificial SequencePurotk_seq_fow2 primer
91tccataccga cgatctgcga c 219227DNAArtificial SequenceJ44 primer
92aaaagcgcct cccctacccg gtagaat 279327DNAArtificial SequenceJ46
primer 93ggcgacacgg aaatgttgaa tactcat 279427DNAArtificial
SequenceJ48 primer 94cactacaccc agaagtccct gagcctg
279520DNAArtificial SequencepSANG10pelB primer 95cgctgcccag
ccggccatgg 209626DNAArtificial SequencePrimer 2097 96gatggtgatg
atgatgtgcg gatgcg 269738DNAArtificial SequencePrimer 2423
97ttttttccat gggccggccc tccttcagtt tagttgag 389839DNAArtificial
SequencePrimer 2437 98ttttttgcgg ccgcggaagc cgtgatctcc ttctctctc
399920DNAArtificial SequencePrimer 2622 99gaacaggaac acggaaggtc
2010022DNAArtificial SequencePrimer 41679 100atgagttgga gctgtatcat
cc 2210118DNAArtificial SequencePrimer 2621 101gcattccacg gcggccgc
181029PRTArtificial SequencePeptide SLLMWITQV (NY-ESO-1 157-165)
recognised by c12c2 102Ser Leu Leu Met Trp Ile Thr Gln Val1
510310PRTArtificial SequencePeptide MART-1 26-35 recognised by MEL5
TCR 103Glu Leu Ala Gly Ile Gly Ile Leu Thr Val1 5
1010430DNAArtificial SequencePrimer 4JFH-Valpha-F 104acacacgcta
gccagaaaga ggtggaacag 3010550DNAArtificial SequencePrimer
2770misc_feature(21)..(22)n is a, c, g, or tmisc_feature(30)..(31)n
is a, c, g, or t 105caaagaacag ctcgccggts nncccgassn nggagctggc
gcaaaagtac 5010653DNAArtificial SequencePrimer
2771misc_feature(23)..(23)n is a, c, g, or tmisc_feature(30)..(31)n
is a, c, g, or tmisc_feature(33)..(34)n is a, c, g, or t
106ctcgcccgaa ggtgggaatg tangwtccsn nsnnaagtgg gcgcacggcg cac
5310722DNAArtificial SequencePrimer 2780 107ctggcagcta gcaagcagga
ag 2210822DNAArtificial SequencePrimer 2781 108tacattccca
ccttcgggcg ag
2210926DNAArtificial SequencePrimer 2782 109ttttttgcgg ccgcggacag
gttctg 2611027DNAArtificial SequencePrimer 2783 110cgtaagctgg
taccttatta tctaggg 2711127DNAArtificial SequencePrimer 2785
111ccctagataa taaggtacca gcttacg 2711219DNAArtificial
SequencePrimer 2787 112accggcgagc tgttctttg 1911326DNAArtificial
SequencePrimer 2788 113agtgacaagc ttttattatc tgggtg
2611419DNAArtificial SequencePrimer 2789 114caggtcctcg agcactgtc
1911525DNAArtificial SequenceCRISPR1 double-stranded DNA insert
strand 115ggggccacta gggacaggat gtttt 2511626DNAArtificial
SequenceCRISPR1 double-stranded DNA insert strand 116catcctgtcc
ctagtggccc ccggtg 2611725DNAArtificial SequenceCRISPR2
double-stranded DNA insert strand 117gtcaccaatc ctgtccctag gtttt
2511825DNAArtificial SequenceCRISPR2 double-stranded DNA insert
strand 118ctagggacag gattggtgac cggtg 25119102RNAArtificial
SequenceCRISPR 1 RNA 119ggggggccac uagggacagg auguuuuaga gcuagaaaua
gcaaguuaaa auaaggcuag 60uccguuauca acuugaaaaa guggcaccga gucggugcuu
uu 102120102RNAArtificial SequenceCRISPR 2 RNA 120gggucaccaa
uccugucccu agguuu
References
D00001
D00002
D00003
D00004
D00005
D00006
D00007
D00008
D00009
D00010
D00011
D00012
D00013

D00014

D00015

D00016

D00017
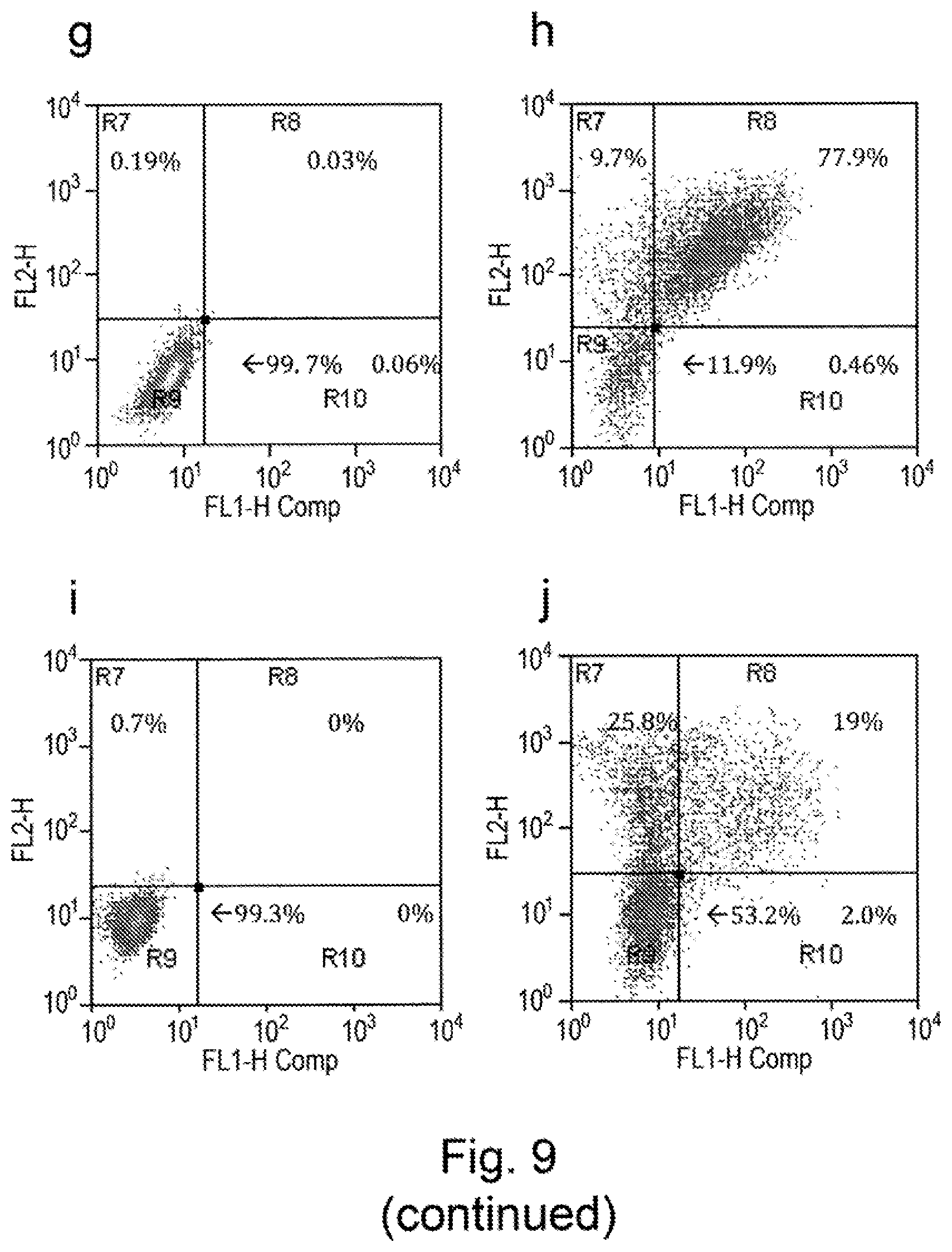
D00018
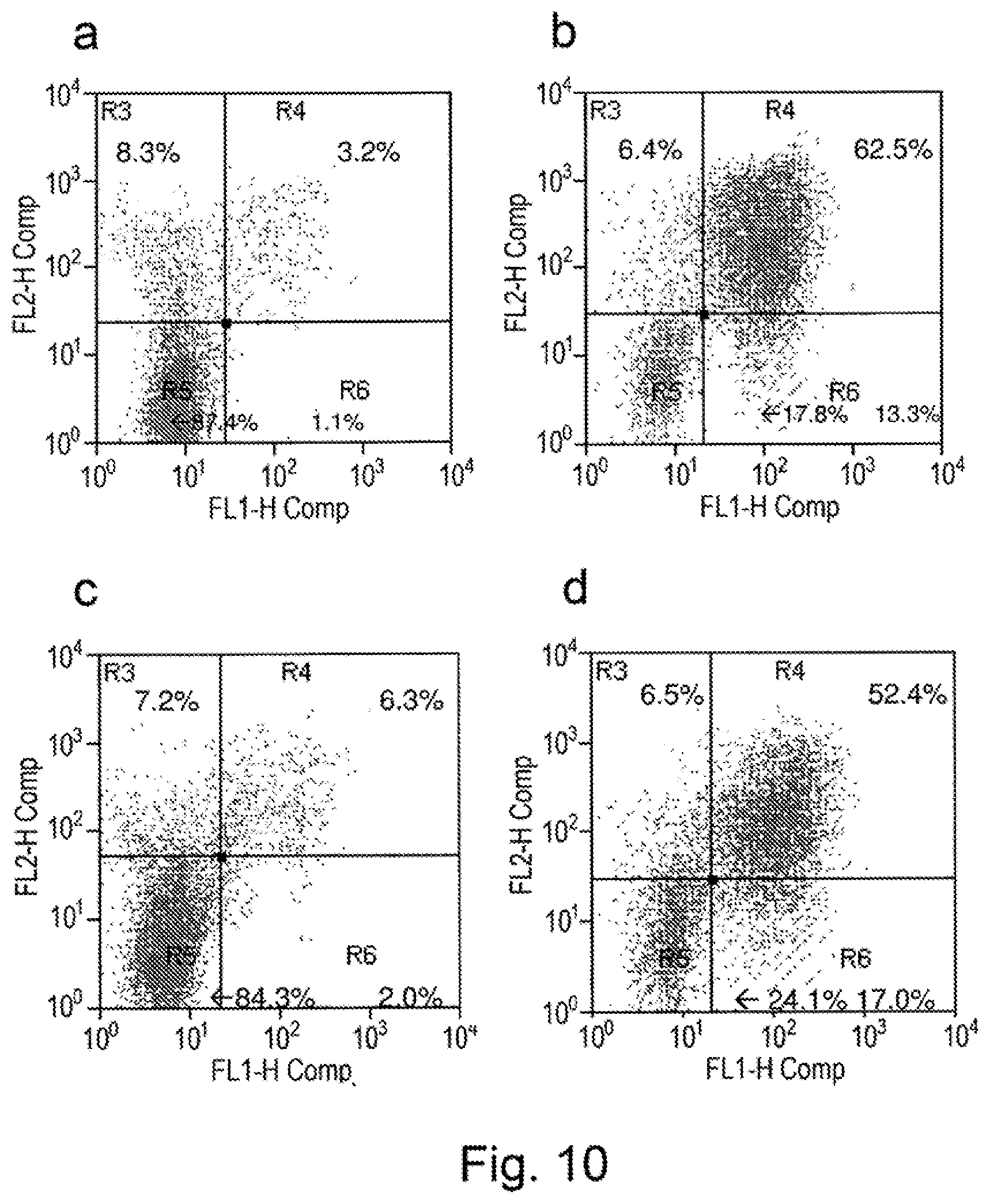
D00019

D00020
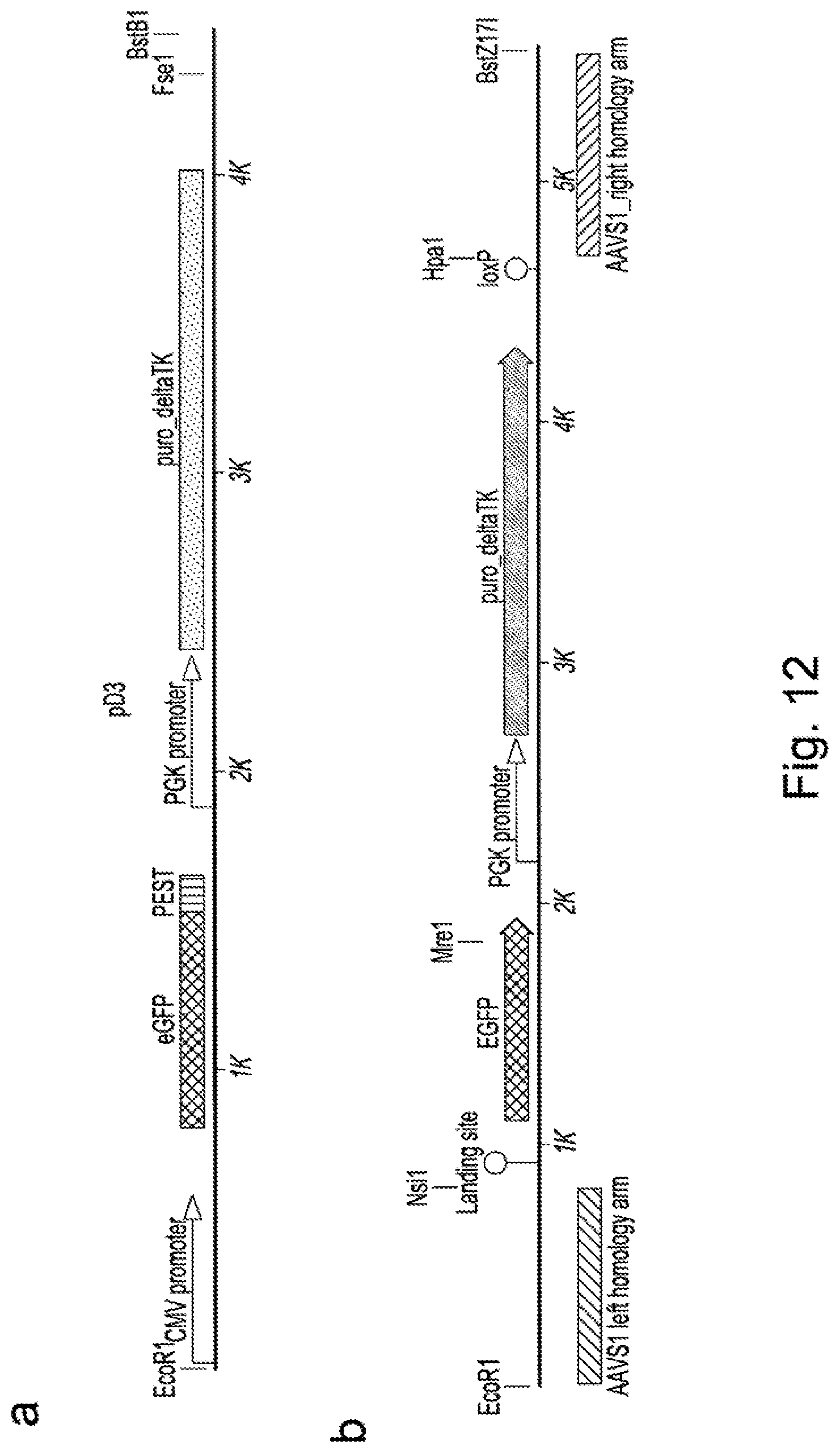
D00021

D00022

D00023
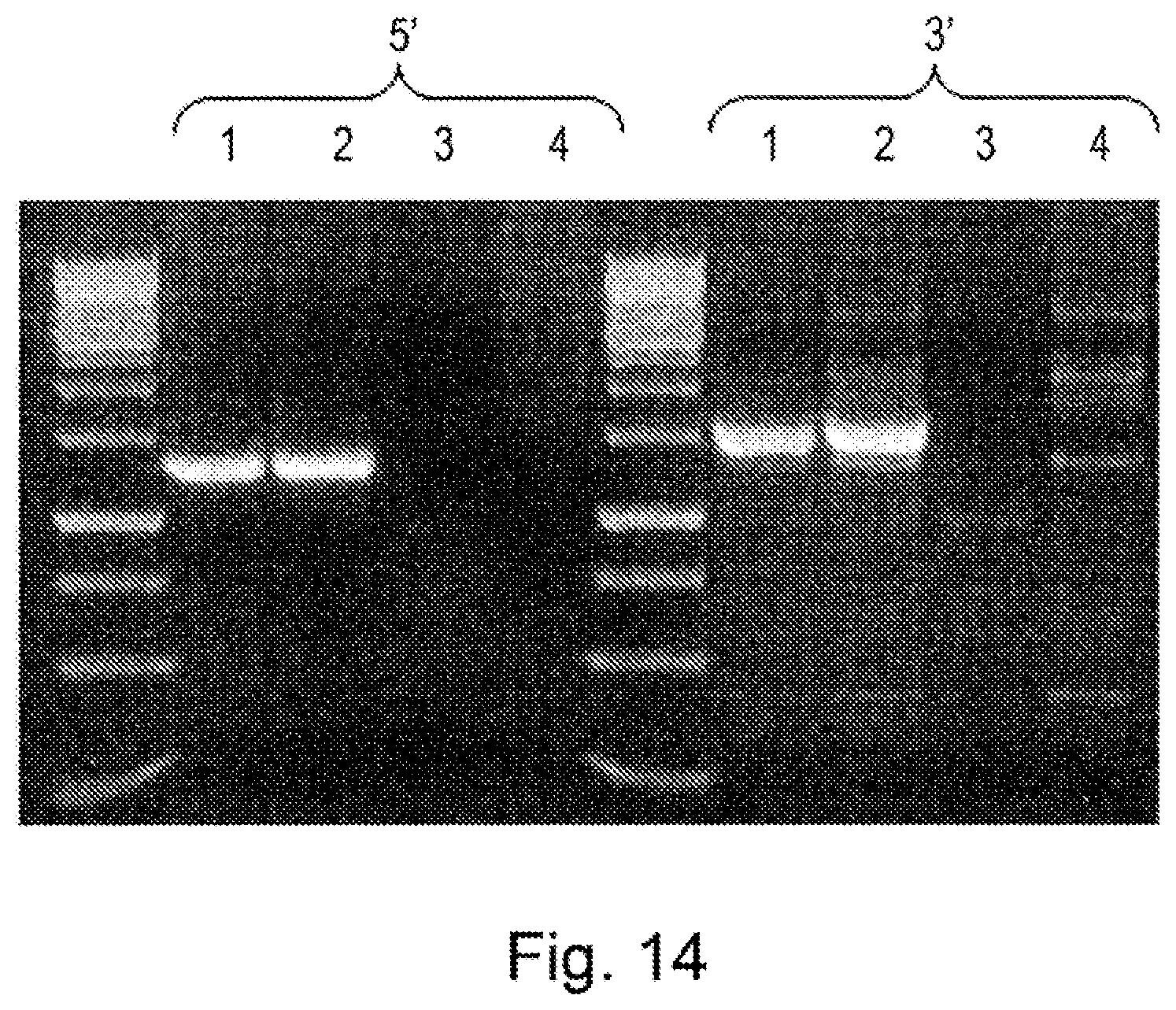
D00024

D00025

D00026

D00027

D00028

D00029
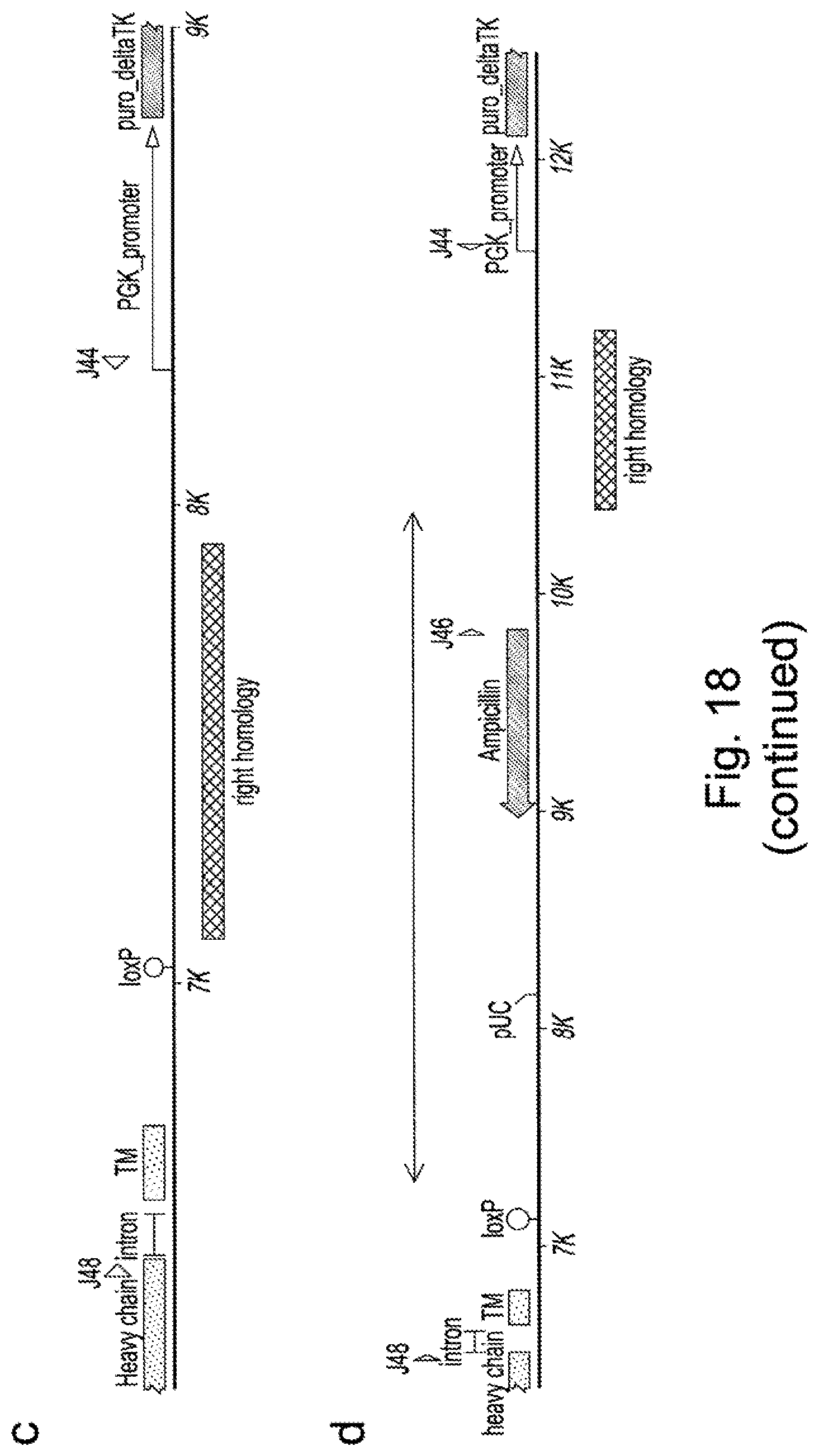
D00030
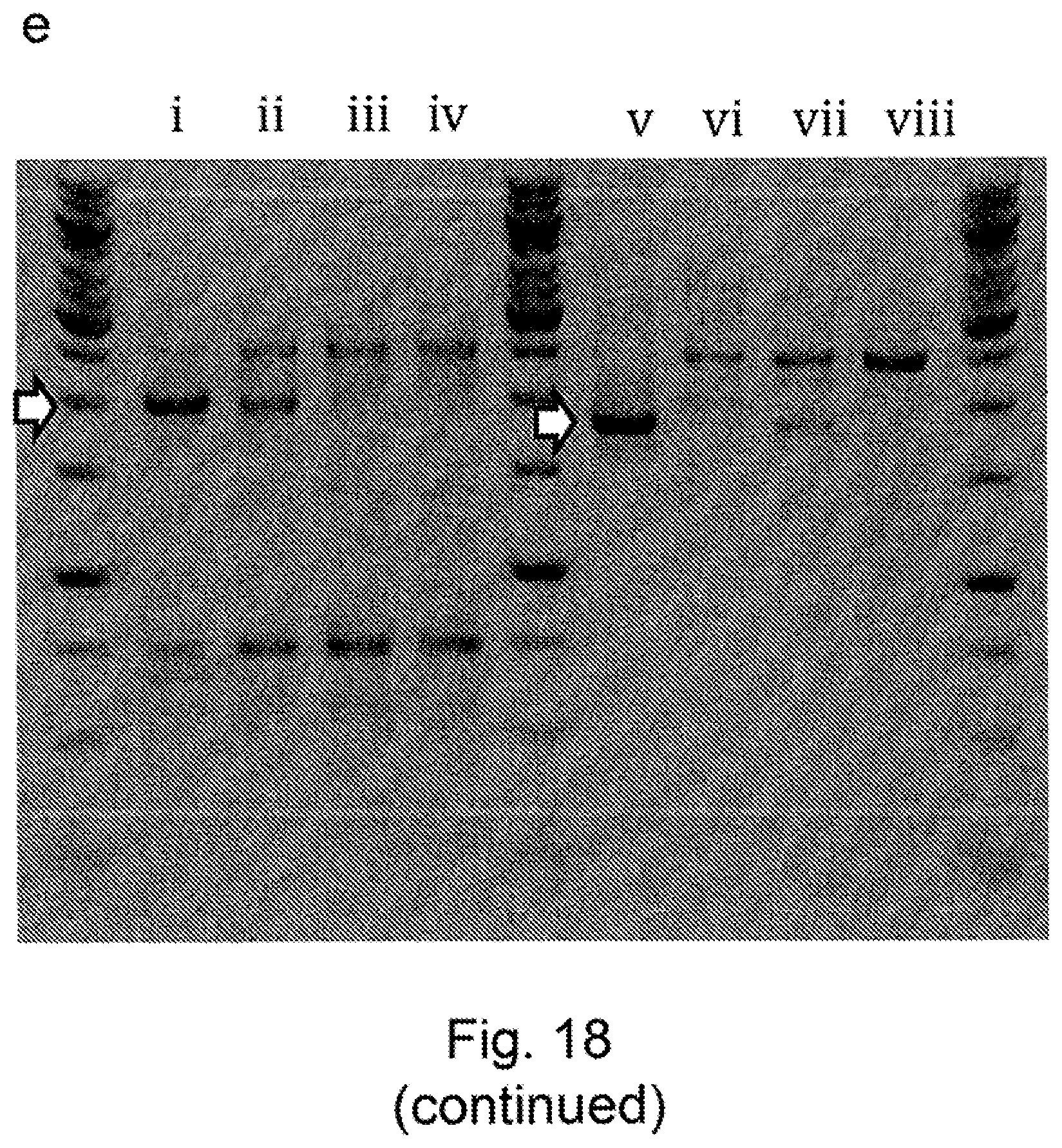
D00031
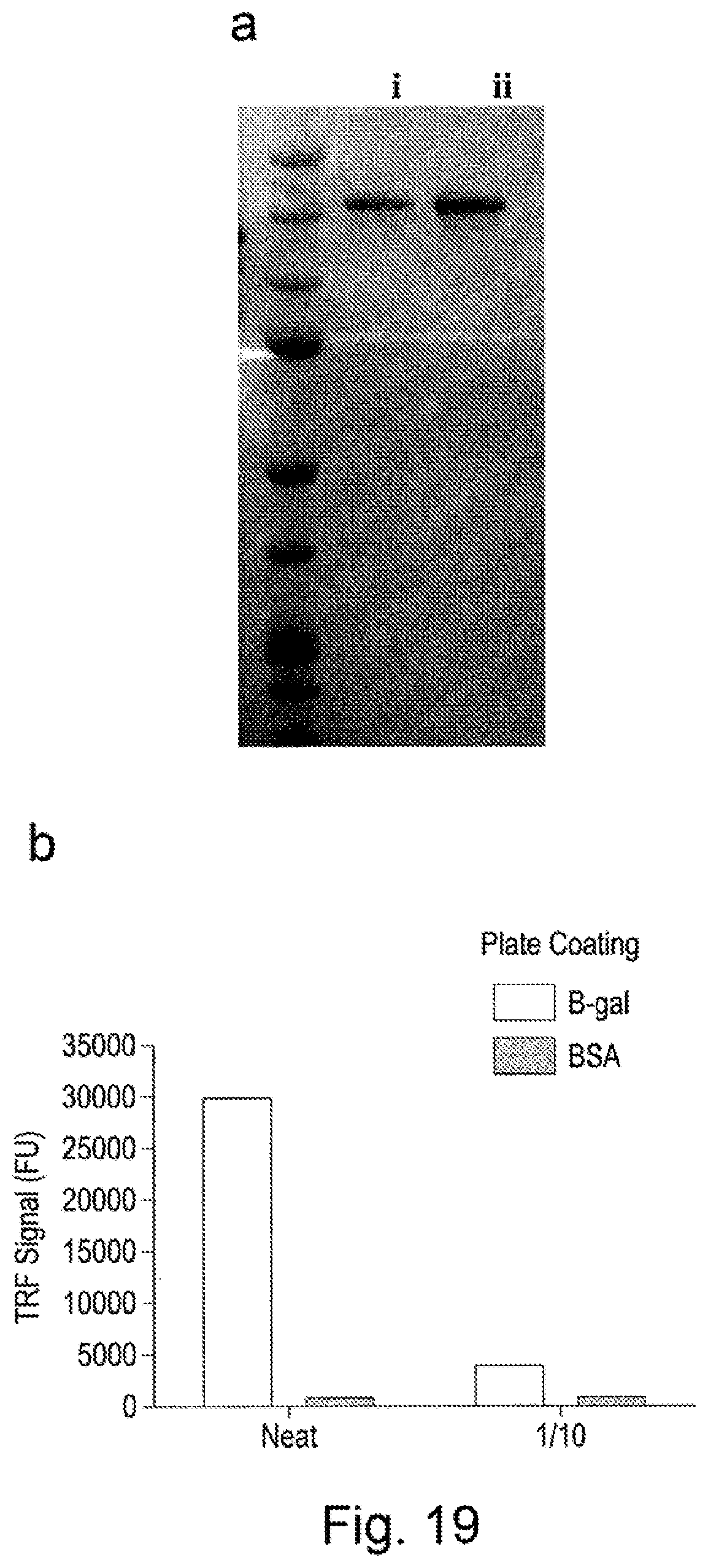
D00032

D00033

D00034

D00035

D00036
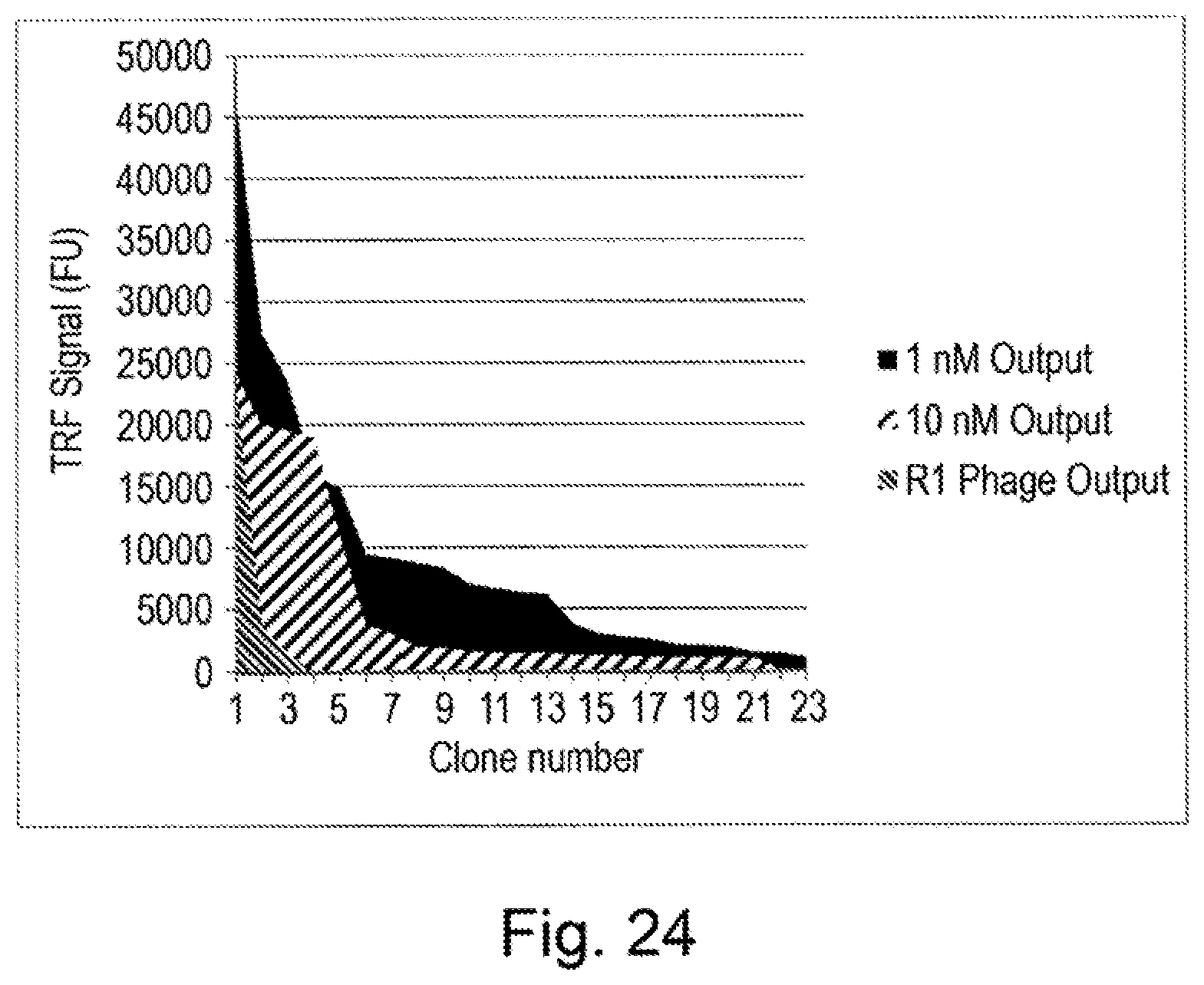
D00037
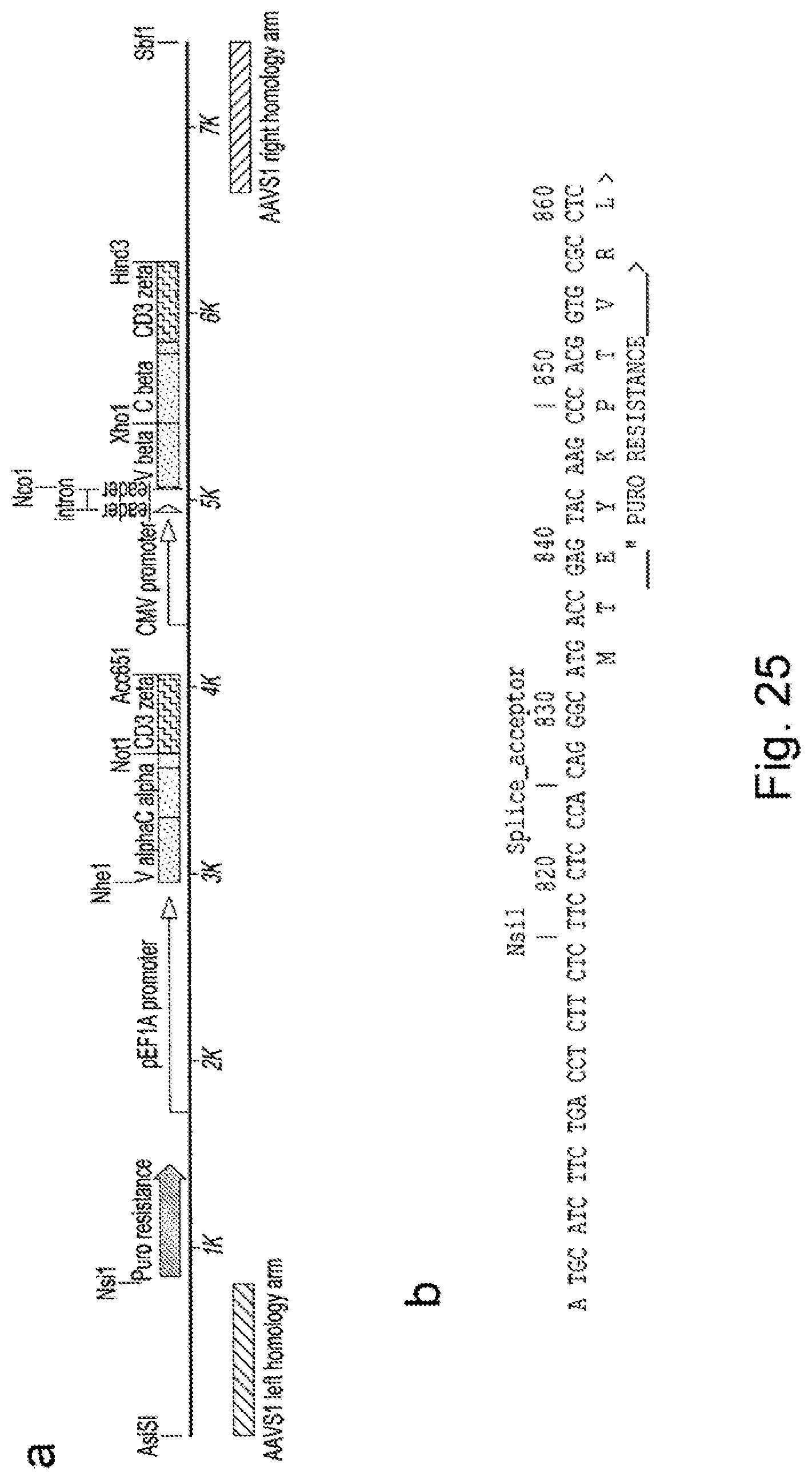
D00038

D00039

D00040

D00041

D00042
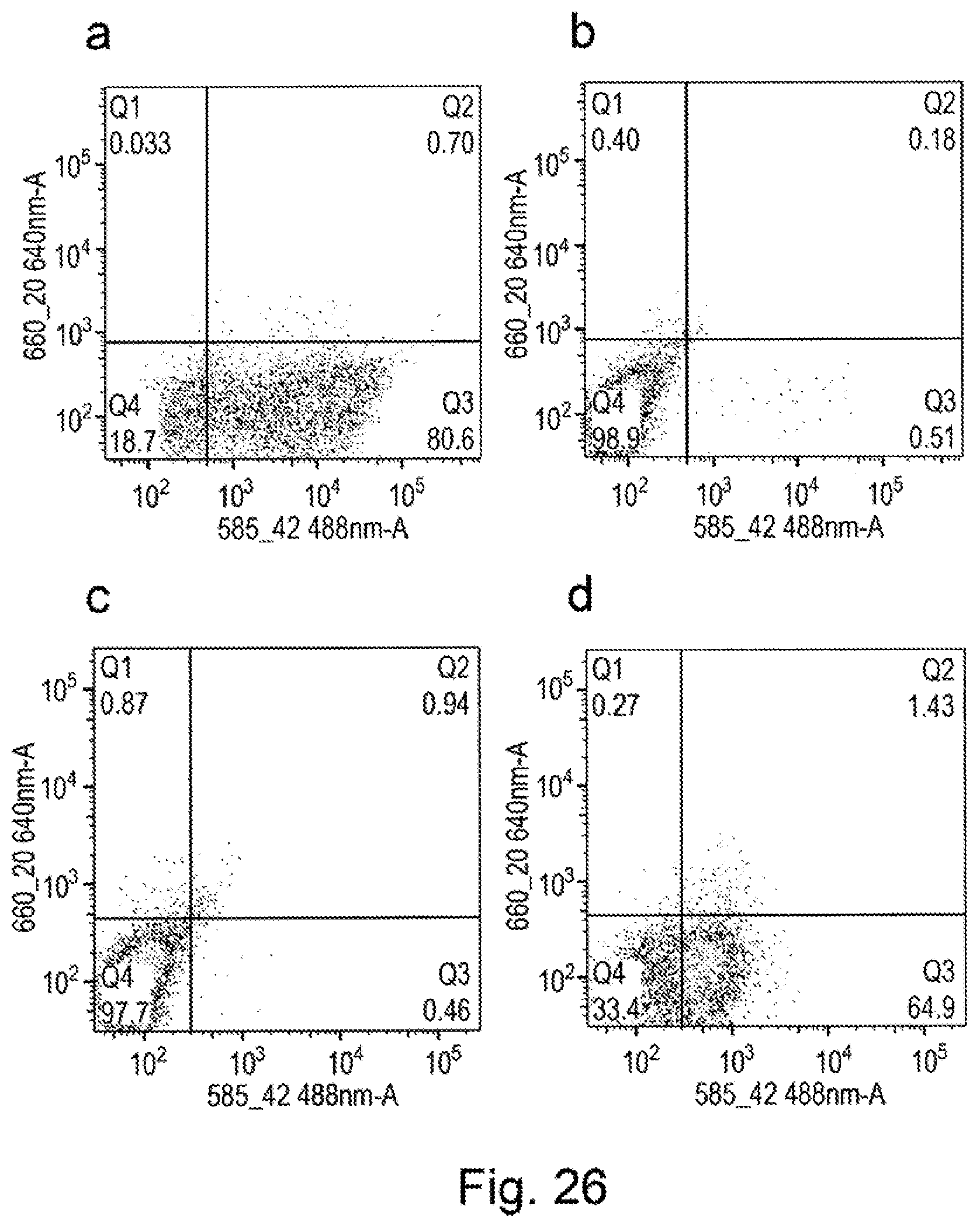
D00043

D00044

D00045

D00046

D00047

D00048

D00049

D00050

D00051

D00052

D00053

D00054

D00055

D00056

S00001
XML
uspto.report is an independent third-party trademark research tool that is not affiliated, endorsed, or sponsored by the United States Patent and Trademark Office (USPTO) or any other governmental organization. The information provided by uspto.report is based on publicly available data at the time of writing and is intended for informational purposes only.
While we strive to provide accurate and up-to-date information, we do not guarantee the accuracy, completeness, reliability, or suitability of the information displayed on this site. The use of this site is at your own risk. Any reliance you place on such information is therefore strictly at your own risk.
All official trademark data, including owner information, should be verified by visiting the official USPTO website at www.uspto.gov. This site is not intended to replace professional legal advice and should not be used as a substitute for consulting with a legal professional who is knowledgeable about trademark law.