Bispecific EGFR/C-Met Antibodies
Chiu; Mark ; et al.
U.S. patent application number 16/697249 was filed with the patent office on 2020-03-19 for bispecific egfr/c-met antibodies. The applicant listed for this patent is Janssen Biotech, Inc.. Invention is credited to Mark Chiu, Sheri Moores, Joost Neijssen, Paul Parren, Janine Schuurman.
| Application Number | 20200087407 16/697249 |
| Document ID | / |
| Family ID | 59896349 |
| Filed Date | 2020-03-19 |





View All Diagrams
| United States Patent Application | 20200087407 |
| Kind Code | A1 |
| Chiu; Mark ; et al. | March 19, 2020 |
Bispecific EGFR/C-Met Antibodies
Abstract
Bispecific EGFR/c-Met antibodies and methods of making and using the molecules.
| Inventors: | Chiu; Mark; (Paoli, PA) ; Moores; Sheri; (Phoenixville, PA) ; Neijssen; Joost; (Ultrecht, NL) ; Parren; Paul; (Ultrecht, NL) ; Schuurman; Janine; (Ultrecht, NL) | ||||||||||
| Applicant: |
|
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Family ID: | 59896349 | ||||||||||
| Appl. No.: | 16/697249 | ||||||||||
| Filed: | November 27, 2019 |
Related U.S. Patent Documents
| Application Number | Filing Date | Patent Number | ||
|---|---|---|---|---|
| 15989532 | May 25, 2018 | |||
| 16697249 | ||||
| 15616016 | Jun 7, 2017 | |||
| 15989532 | ||||
| 15386195 | Dec 21, 2016 | 9695242 | ||
| 15616016 | ||||
| 14283257 | May 21, 2014 | 9580508 | ||
| 15386195 | ||||
| 14086588 | Nov 21, 2013 | 9593164 | ||
| 14283257 | ||||
| 61728912 | Nov 21, 2012 | |||
| 61782550 | Mar 14, 2013 | |||
| 61809541 | Apr 8, 2013 | |||
| 61864717 | Aug 12, 2013 | |||
| 61892797 | Oct 18, 2013 | |||
| Current U.S. Class: | 1/1 |
| Current CPC Class: | A61K 31/517 20130101; A61K 39/395 20130101; A61K 2300/00 20130101; A61K 31/437 20130101; A61K 33/24 20130101; A61K 31/5377 20130101; C07K 2318/20 20130101; A61K 33/24 20130101; A61K 45/06 20130101; C07K 2317/76 20130101; A61K 2039/505 20130101; A61K 39/395 20130101; A61K 31/4545 20130101; A61K 2300/00 20130101; A61K 2300/00 20130101; A61K 2300/00 20130101; A61K 31/4545 20130101; C07K 2317/31 20130101; A61K 2300/00 20130101; C07K 16/2863 20130101; A61K 31/517 20130101; C07K 2317/73 20130101; A61K 39/3955 20130101 |
| International Class: | C07K 16/28 20060101 C07K016/28; A61K 39/395 20060101 A61K039/395; A61K 33/24 20060101 A61K033/24; A61K 31/5377 20060101 A61K031/5377; A61K 31/437 20060101 A61K031/437; A61K 45/06 20060101 A61K045/06; A61K 31/517 20060101 A61K031/517; A61K 31/4545 20060101 A61K031/4545 |
Claims
1. An isolated bispecific epidermal growth factor receptor (EGFR)/hepatocyte growth factor receptor (c-Met) antibody, comprising: a) a first heavy chain (HC1) comprising a HC1 constant domain 3 (HC1 CH3) and a HC1 variable region 1 (VH1); b) a first light chain (LC1) comprising a light chain variable region 1 (VL1); c) a second heavy chain (HC2) comprising a HC2 constant domain 3 (HC2 CH3) and a HC2 variable region 2 (VH2); and d) a second light chain (LC2) comprising a light chain variable region 2 (VL2), wherein the VH1 comprises a heavy chain complementarity determining region 1 (HCDR1), a HCDR2 and a HCDR3 amino acid sequences of SEQ ID NOs: 210, 211 and 212, respectively; the VL1 comprises a light chain complementarity determining region 1 (LCDR1), a LCDR2 and a LCDR3 amino acid sequences of SEQ ID NOs: 213, 214 and 215, respectively, the VH2 comprises the HCDR1, the HCDR2 and the HCDR3 amino acid sequences of SEQ ID NOs: 216, 217 and 218, respectively; and the VL2 comprises the LCDR1, the LCDR2 and the LCDR3 amino acid sequences of SEQ ID NOs: 219, 220 and 221, respectively.
2. The bispecific antibody of claim 1, wherein the antibody inhibits growth of NCI-H292 or NCI-H1975 cells with an IC.sub.50 value that is at least about 300-fold less, at least about 400-fold less, at least about 500-fold less, at least about 600-fold less, at least about 700-fold less or at least about 800-fold less when compared to the IC.sub.50 value of inhibition of growth of NCI-H292 or NCI-H1975 cells with cetuximab when NCI-H292 or NCI-H1975 cells are grown in low attachment conditions.
3. The bispecific antibody of claim 1, wherein the antibody inhibits growth of HGF-expressing SKMES-1 cell tumor in SCID Beige mice with a percentage (%) T/C value of at least 500-fold less on day 36 when compared to cetuximab, when the bispecific antibody and cetuximab are administered at 20 mg/kg dose.
4. The bispecific antibody of claim 1, wherein the VH1, the VL1, the VH2 and the VL2 comprise the amino acid sequences of SEQ ID NOs: 189, 190, 193 and 194, respectively.
5. The bispecific antibody of claim 4, wherein the HC1 and the HC2 are an IgG1 isotype.
6. The bispecific antibody of claim 5, wherein the HC1 CH3 comprises at least one substitution and the HC2 CH3 comprises at least one substitution at residue positions 405 or 409.
7. The bispecific antibody of claim 6, wherein the HC1 CH3 comprises a K409R or a F405L substitution and the HC2 CH3 comprises a K409R or a F405L substitution.
8. The bispecific antibody of claim 7, wherein a) the HC1 CH3 comprises the F405L substitution and the HC2 CH3 comprises the K409R substitution; or b) the HC1 CH3 comprises the K409R substitution and the HC2 CH3 comprises the F405L substitution.
9. The bispecific antibody of claim 1, wherein the HC1 comprises the amino acid sequence of SEQ ID NO: 204 and the HC2 comprises the amino acid sequence of SEQ ID NO: 203.
10. The bispecific antibody of claim 9, wherein the antibody has a biantennary glycan structure with a fucose content of about between 1% to about 15%.
11. A method of treating a subject having cancer, comprising administering a therapeutically effective amount of the bispecific EGFR/c-Met antibody of claim 1 to a patient in need thereof to treat the cancer.
12. The method of claim 11, wherein the cancer is associated with an EGFR activating mutation, an EGFR gene amplification, increased levels of circulating HGF, a c-Met activating mutation, a c-Met gene amplification or a mutant KRAS.
13. The method of claim 12, wherein the EGFR activating mutation is G719A, G719X (X being any amino acid), L861X (X being any amino acid), L858R, E746K, L747S, E749Q, A750P, A755V, V765M, L858P or T790M substitution, deletion of E746-A750, deletion of R748-P753, insertion of Ala (A) between M766 and A767, insertion of Ser, Val and Ala (SVA) between 5768 and V769, and insertion of Asn and Ser (NS) between P772 and H773.
14. The method of claim 12, wherein the mutant KRAS has a G12V or G12C substitution.
15. The method of claim 11, wherein the subject is resistant or has acquired resistance to treatment with erlotinib, gefitinib, afatinib, CO-1686, AZD9192 or cetuximab.
16. The method of claim 11, wherein the cancer is an epithelial cell cancer, breast cancer, ovarian cancer, lung cancer, non-small cell lung cancer (NSCLC), lung adenocarcinoma, small cell lung cancer, colorectal cancer, anal cancer, prostate cancer, kidney cancer, bladder cancer, head and neck cancer, pharynx cancer, cancer of the nose, pancreatic cancer, skin cancer, oral cancer, cancer of the tongue, esophageal cancer, vaginal cancer, cervical cancer, cancer of the spleen, testicular cancer, gastric cancer, cancer of the thymus, colon cancer, thyroid cancer, liver cancer, hepatocellular carcinoma (HCC) or sporadic or hereditary papillary renal cell carcinoma (PRCC).
17. The method of claim 11, wherein the subject is homozygous for phenylalanine at position 158 of CD16 or heterozygous for valine and phenylalanine at position 158 of CD16.
18. The method of claim 11, comprising administering a second therapeutic agent.
19. The method of claim 18, wherein the second therapeutic agent is a chemotherapeutic agent or a targeted anti-cancer therapy.
20. The method of claim 11, wherein the second therapeutic agent is cisplatin, vinblastine or a tyrosine kinase inhibitor of EGFR, c-Met, HER2, HER3, HER4 or VEGFR.
21. A method of inhibiting growth or proliferation of cells that express EGFR and/or c-Met, comprising contacting the cells with the bispecific antibody of claim 1.
22. A method of inhibiting growth or metastasis of EGFR and/or c-Met expressing tumor or cancer cells in a subject comprising administering to the subject an effective amount of the bispecific antibody of claim 1 to inhibit the growth or metastasis of EGFR and/or c-Met expressing tumor or cancer cells.
23. The method of claim 22, wherein the EGFR and/or c-Met expressing tumor is an epithelial cell cancer, breast cancer, ovarian cancer, lung cancer, non-small cell lung cancer (NSCLC), lung adenocarcinoma, small cell lung cancer, colorectal cancer, anal cancer, prostate cancer, kidney cancer, bladder cancer, head and neck cancer, pharynx cancer, cancer of the nose, pancreatic cancer, skin cancer, oral cancer, cancer of the tongue, esophageal cancer, vaginal cancer, cervical cancer, cancer of the spleen, testicular cancer, gastric cancer, cancer of the thymus, colon cancer, thyroid cancer, liver cancer, hepatocellular carcinoma (HCC) or sporadic or hereditary papillary renal cell carcinoma (PRCC).
24. The method of claim 23, wherein the EGFR and/or c-Met expressing tumor is associated with an EGFR activating mutation, an EGFR gene amplification, increased levels of circulating HGF, a c-Met activating mutation, a c-Met gene amplification or a mutant KRAS.
25. The method of claim 24, wherein the EGFR activating mutation is G719A, G719X (X being any amino acid), L861X (X being any amino acid), L858R, E746K, L747S, E749Q, A750P, A755V, V765M, L858P or T790M substitution, deletion of E746-A750, deletion of R748-P753, insertion of Ala (A) between M766 and A767, insertion of Ser, Val and Ala (SVA) between 5768 and V769, and insertion of Asn and Ser (NS) between P772 and H773.
26. The method of claim 24, wherein the mutant KRAS has a G12V or G12C substitution.
Description
CROSS REFERENCE TO RELATED APPLICATIONS
[0001] This application is a continuation of U.S. application Ser. No. 15/989,532, filed 25 May 2018, currently pending, which is a continuation of U.S. application Ser. No. 15/616,016, filed 20 Apr. 2018, now abandoned, which is a continuation-in-part of U.S. application Ser. No. 15/386,195, filed 21 Dec. 2016, now U.S. Pat. No. 9,695,242, which is a divisional application of U.S. application Ser. No. 14/283,257, filed 21 May 2014, now U.S. Pat. No. 9,580,508, which is a continuation-in-part of U.S. application Ser. No. 14/086,588, filed 21 Nov. 2013, now U.S. Pat. No. 9,593,164, which claims the benefit of U.S. Provisional Application No. 61/728,912, filed 21 Nov. 2012, U.S. Provisional Application No. 61/782,550, filed 14 Mar. 2013, U.S. Provisional Application No. 61/809,541, filed 8 Apr. 2013, U.S. Provisional Application No. 61/864,717 filed 12 Aug. 2013, and U.S. Provisional Application No. 61/892,797, filed 18 Oct. 2013, the entire contents of which are incorporated herein by reference.
FIELD OF THE INVENTION
[0002] The present invention relates to bispecific EGFR/c-Met antibodies and methods of making and using the molecules.
BACKGROUND OF THE INVENTION
[0003] Epidermal growth factor receptor (EGFR, ErbB1 or HER1) is a Type I transmembrane glycoprotein of 170 kDa that is encoded by the c-erbB1 proto-oncogene. EGFR is a member of the human epidermal growth factor receptor (HER) family of receptor tyrosine kinases (RTK) which includes HER2 (ErbB2), HER3 (ErbB3) and HER4 (ErbB4). EGFR signaling is initiated by ligand binding followed by induction of conformational change, homodimerization or heterodimerization of the receptor with other ErbB family members, and trans-autophosphorylation of the receptor (Ferguson et al., Annu Rev Biophys, 37: 353-73, 2008), which initiates signal transduction cascades that ultimately affect a wide variety of cellular functions, including cell proliferation and survival. Increases in expression or kinase activity of EGFR have been linked with a range of human cancers, making EGFR an attractive target for therapeutic intervention (Mendelsohn et al., Oncogene 19: 6550-6565, 2000; Grunwald et al., J Natl Cancer Inst 95: 851-67, 2003; Mendelsohn et al., Semin Oncol 33: 369-85, 2006). Increases in both the EGFR gene copy number and protein expression have been associated with favorable responses to the EGFR tyrosine kinase inhibitor, IRESSA.TM. (gefitinib), in non-small cell lung cancer (Hirsch et al., Ann Oncol 18:752-60, 2007).
[0004] EGFR therapies include both small molecules and anti-EGFR antibodies, approved for treatment of colorectal cancer, pancreatic cancer, head and neck cancer, and non-small cell lung cancer (NSCLC) (Baselga and Arteaga, J Clin Oncol 23:2445-2459 (20005; Gill et al., J Biol Chem, 259:7755-7760, 1984; Goldstein et al., Clin Cancer Res, 1:131 1-1318; 1995; Prewett et al., Clin Cancer Res, 4:2957-2966, 1998).
[0005] Efficacy of anti-EGFR therapies may depend on tumor type and EGFR mutation/amplification status in the tumor. Side effects of current therapeutics may include skin toxicity (De Roock et al., Lancet Oncol 11:753-762, 2010; Linardou et al., Nat Rev Clin Oncol, 6: 352-366, 2009; Li and Perez-Soler, Targ Oncol 4: 107-119, 2009). EGFR tyrosine kinase inhibitors (TKI) are commonly used as 2.sup.nd line therapies for non-small cell lung cancer (NSCLC), but often stop working within twelve months due to resistance pathways (Riely et al., Clin Cancer Res 12: 839-44, 2006).
[0006] c-Met encodes a transmembrane tyrosine kinase receptor. It was first identified as a proto-oncogene in 1984 after it was found that treatment with a carcinogen resulted in a constitutively active fusion protein TPR-MET (Cooper et al., Nature 311:29-33, 1984). Activation of c-Met by its ligand hepatocyte growth factor (HGF) stimulates a plethora of cell processes including growth, motility, invasion, metastasis, epithelial-mesenchymal transition, angiogenesis/wound healing, and tissue regeneration (Christensen et al., Cancer Lett 225:1-26, 2005; Peters and Adjei, Nat Rev Clin Oncol 9:314-26, 2012). c-Met is synthesized as a single chain protein that is proteolytically cleaved into a 50 kDa alpha- and 140 kDa beta-subunits that are linked by a disulphide bond (Ma et al., Cancer and Metastasis Reviews, 22: 309-325, 2003). c-Met is structurally similar to other membrane receptors such as RON and Sea. The exact stoichiometry of HGF:c-Met binding is unclear, but it is generally believed that two HGF molecules bind to two c-Met molecules leading to receptor dimerization and autophosphorylation at tyrosines 1230, 1234, and 1235 (Stamos et al., The EMBO Journal 23: 2325-2335, 2004). Ligand-independent c-Met autophosphorylation can also occur due to gene amplification, mutation or receptor over-expression.
[0007] c-Met is frequently amplified, mutated or over-expressed in many types of cancer including gastric, lung, colon, breast, bladder, head and neck, ovarian, prostate, thyroid, pancreatic, and CNS cancers. Missense mutations typically localized to the kinase domain are commonly found in hereditary papillary renal cell carcinomas (PRCC) and in 13% of sporadic PRCCs (Schmidt et al., Oncogene 18: 2343-2350, 1999). c-Met mutations localized to the semaphorin or juxtamembrane domains of c-Met are frequently found in gastric, head and neck, liver, ovarian, NSCLC and thyroid cancers (Ma et al., Cancer and Metastasis Reviews, 22: 309-325, 2003; Sakakura et al., Chromosomes and Cancer, 1999. 24:299-305). c-Met amplification has been detected in brain, colorectal, gastric, and lung cancers, often correlating with disease progression (Ma et al., Cancer and Metastasis Reviews, 22: 309-325, 2003). Up to 4% and 20% of non-small cell lung cancer (NSCLC) and gastric cancers, respectively, exhibit c-Met amplification (Sakakura et al., Chromosomes and Cancer, 1999. 24:299-305: Sierra and Tsao, Therapeutic Advances in Medical Oncology, 3:S21-35, 2011). Even in the absence of gene amplification, c-Met overexpression is frequently observed in lung cancer (Ichimura et al., Jpn J Cancer Res, 87:1063-9, 1996). Moreover, in clinical samples, nearly half of lung adenocarcinomas exhibited high levels of c-Met and HGF, both of which correlated with enhanced tumor growth rate, metastasis and poor prognosis (Sierra and Tsao, Therapeutic Advances in Medical Oncology, 3:S21-35, 2011; Siegfried et al., Ann Thorac Surg 66: 1915-8, 1998).
[0008] Nearly 60% of all tumors that become resistant to EGFR tyrosine kinase inhibitors increase c-Met expression, amplify c-Met, or increase c-Met only known ligand, HGF (Turke et al., Cancer Cell, 17:77-88, 2010), suggesting the existence of a compensatory pathway for EGFR through c-Met. c-Met amplification was first identified in cultured cells that became resistant to gefitinib, an EGFR kinase inhibitor, and exhibited enhanced survival through the Her3 pathway (Engelman et al., Science, 316:1039-43, 2007). This was further validated in clinical samples where nine of 43 patients with acquired resistance to either erlotinib or gefitinib exhibited c-Met amplification, compared to only two of 62 untreated patients. Four of the nine treated patients also acquired the EGFR activating mutation, T790M, demonstrating simultaneous resistance pathways (Beat et al., Proc Natl Acad Sci USA, 104:20932-7, 2007).
[0009] The individual roles of both EGFR and c-Met in cancer is well established, making these targets attractive for combination therapy. Both receptors signal through the same survival and anti-apoptotic pathways (ERK and AKT); thus, inhibiting the pair in combination may limit the potential for compensatory pathway activation thereby improving overall efficacy. Combination therapies targeting EGFR and c-Met are tested in clinical trials with Tarceva.RTM. (erlotinib) in combination with anti-c-Met monovalent antibody for NSCLC (Spigel et al., 2011 ASCO Annual Meeting Proceedings 2011, Journal of Clinical Oncology: Chicago, Ill. p. 7505) and Tarceva (erlotinib) in combination with ARQ-197, a small molecule inhibitor of c-Met (Adjei et al., Oncologist, 16:788-99, 2011). Combination therapies or bispecific anti-EGFR/c-Met molecules have been disclosed for example in: Intl. Pat. Publ. Nos. WO2008/127710, WO2009/111691, WO2009/126834, WO2010/039248, WO2010/115551 and U.S. Pat. Publ. No. US2009/0042906.
[0010] Current small molecule and large molecule therapeutic approaches to antagonize EGFR and/or c-Met signaling pathways for therapy may be sub-optimal due to possible lack of specificity, potential off-target activity and dose-limiting toxicity that may be encountered with small molecule inhibitors. Typical monospecific bivalent antibodies may result in clustering of membrane bound receptors and unwanted activation of the downstream signaling pathways. Monovalent antibodies having full length heavy chains (half arms) pose significant complexity and cost to the manufacturing process.
[0011] Accordingly, the need exists for additional monospecific and bispecific EGFR and/or c-Met inhibitors for both therapeutic and diagnostic purpose.
SUMMARY OF THE INVENTION
[0012] One embodiment of the invention is an isolated bispecific epidermal growth factor receptor (EGFR)/hepatocyte growth factor receptor (c-Met) antibody, comprising: [0013] a first heavy chain (HC1) comprising a HC1 constant domain 3 (HC1 CH3) and a HC1 variable region 1 (VH1); [0014] a second heavy chain (HC2) comprising a HC2 constant domain 3 (HC2 CH3) and a HC2 variable region 2 (VH2); [0015] a first light chain (LC1) comprising a light chain variable region 1 (VL1); and [0016] a second light chain (LC2) comprising a light chain variable region 2 (VL2), wherein the VH1 and the VL1 pair to form a first antigen-binding site that specifically binds EGFR, the VH2 and the VL2 pair to form a second antigen-binding site that specifically binds c-Met, the HC1 comprises at least one substitution in the HC1 CH3 and the HC2 comprises at least one substitution in the HC2 CH3, and the substitution in the HC1 CH3 and the substitution in the HC2 CH3 occur at different amino acid residue positions, when residue numbering is according to the EU index.
[0017] In other embodiments, the invention provides for bispecific EGFR/c-Met antibodies, wherein the antibody inhibits phosphorylation of extracellular signal-related kinases 1 and 2 (ERK1/2) in NCI-H292, NCI-H1975 or SKMES-1 cell line with an IC.sub.50 value that is at least about 10-fold less, at least about 20-fold less, at least about 30-fold less, at least about 40-fold less, at least about 50-fold less or at least about 60-fold less when compared to the IC.sub.50 value of inhibition of phosphorylation of ERK1/2 in NCI-H292, NCI-H1975 or SKMES-1 cell lines with a mixture of a control monovalent EGFR antibody comprising a heavy chain 3 (HC3) and a light chain 3 (LC3) and a control monovalent c-Met antibody comprising a heavy chain 4 (HC4) and a light chain 4 (LC4), wherein the HC3 and the HC1, the LC3 and the LC1, the HC4 and the HC2, and the LC4 and the LC2 have identical amino acid sequences, respectively, wherein the phosphorylation of ERK1/2 is measured in whole cell lysates using a sandwich immunoassay using an anti-phosphoERK1/2 antibody as a capture antibody and an antibody binding to unphosphorylated and phosphorylated ERK1/2 conjugated with an electrochemiluminescent compound as a detection antibody.
[0018] In other embodiments, the invention provides for bispecific EGFR/c-Met antibodies, wherein the antibody inhibits phosphorylation of protein kinase B (AKT) at Ser473 in NCI-H1975 cell line with an IC.sub.50 value that is at least about 70-fold less when compared to the IC.sub.50 value of inhibition of phosphorylation of AKT at Ser473 in NCI-H1975 cell line with the mixture of the control monovalent EGFR antibody comprising the HC3 and the LC3 and the control monovalent c-Met antibody comprising the HC4 and the LC4, wherein the HC3 and the HC1, the LC3 and the LC1, the HC4 and the HC2, and the LC4 and the LC2 have identical amino acid sequences, respectively, wherein the phosphorylation of AKT at Ser473 is measured in whole cell lysates using a sandwich immunoassay using an antibody binding to unphosphorylated and phosphorylated AKT as a capture antibody and an anti-phosphoAKT Ser473 antibody conjugated to an electrochemiluminescent compound as a detection antibody.
[0019] In other embodiments, the invention provides for bispecific EGFR/c-Met antibodies that bind EGFR of SEQ ID NO: 73 at EGFR residues K489, I491, K467 and S492 and c-Met at residues PEFRDSYPIKYVHAF (SEQ ID NO: 238) and FAQSKPDSAEPMDRSA (SEQ ID NO: 239).
[0020] In other embodiments, the invention provides for bispecific EGFR/c-Met antibodies that inhibit growth of NCI-H292 or NCI-H1975 cells with an IC.sub.50 value that is at least about 300-fold less, at least about 400-fold less, at least about 500-fold less, at least about 600-fold less, at least about 700-fold less or at least about 800-fold less when compared to the IC.sub.50 value of inhibition of growth of NCI-H292 or NCI-H1975 cells with cetuximab, when NCI-H292 or NCI-H1975 cells are grown in low attachment conditions.
[0021] In other embodiments, the invention provides for bispecific EGFR/c-Met antibodies that inhibit growth of HGF-expressing SKMES-1 cell tumor in SCID Beige mice with percentage (%) T/C value of at least 500-fold less on day 36 when compared to cetuximab, when the bispecific antibody and cetuximab are administered at 20 mg/kg dose.
[0022] In other embodiments, the invention provides for bispecific EGFR/c-Met antibodies wherein the HC1 CH3 comprises a K409R or a F405L substitution and the HC2 CH3 comprises a K409R or F405L substitution, wherein residue numbering is according to the EU index.
[0023] In other embodiments, the invention provides for bispecific EGFR/c-Met antibodies comprising certain heavy and light chain CDR, VH1, VL1, VH2, VL2, HC1, LC1, HC2 and LC2 sequences.
[0024] Another embodiment of the invention is an isolated synthetic polynucleotide encoding the HC1, the HC2, the LC1 or the LC2 of the invention.
[0025] Another embodiment of the invention is a vector comprising the polynucleotide of the invention.
[0026] Another embodiment of the invention is a host cell comprising the vector of the invention.
[0027] Another embodiment of the invention is a method of producing the isolated bispecific EGFR/c-Met antibody, comprising: [0028] combining an isolated monospecific bivalent anti-EGFR antibody comprising two heavy chains of SEQ ID NO: 199 and two light chains of SEQ ID NO: 200 and an isolated monospecific bivalent anti-c-Met antibody comprising two heavy chains of SEQ ID NO: 201 and two light chains of SEQ ID NO: 202 in a mixture of about 1:1 molar ratio; [0029] introducing a reducing agent into the mixture; [0030] incubating the mixture about ninety minutes to about six hours; [0031] removing the reducing agent; and [0032] purifying the bispecific EGFR/c-Met antibody that comprises a first heavy chain of SEQ ID NO: 199 and a second heavy chain of SEQ ID NO: 201, a first light chain of SEQ ID NO: 200 and a second light chain of SEQ ID NO: 202, wherein the first heavy chain of SEQ ID NO: 199 pairs with the first light chain of SEQ ID NO: 200 to form the first binding domain that specifically binds EGFR, and the second heavy chain of SEQ ID NO: 201 pairs with the second light chain of SEQ ID NO: 202 to form the second binding domain that specifically binds c-Met.
[0033] Another embodiment of the invention is a pharmaceutical composition comprising the bispecific antibody of the invention and a pharmaceutically acceptable carrier.
[0034] Another embodiment of the invention is method of treating a subject having cancer, comprising administering a therapeutically effective amount of the bispecific EGFR/c-Met antibody of the invention to a patient in need thereof for a time sufficient to treat the cancer.
[0035] Another embodiment of the invention is method of inhibiting growth or proliferation of cells that express EGFR and/or c-Met, comprising contacting the cells with the bispecific antibody of the invention.
[0036] Another embodiment of the invention is method of inhibiting growth or metastasis of EGFR and/or c-Met expressing tumor or cancer cells in a subject comprising administering to the subject an effective amount of the bispecific antibody of the invention to inhibit the growth or metastasis of EGFR and/or c-Met expressing tumor or cancer cells.
BRIEF DESCRIPTION OF THE DRAWINGS
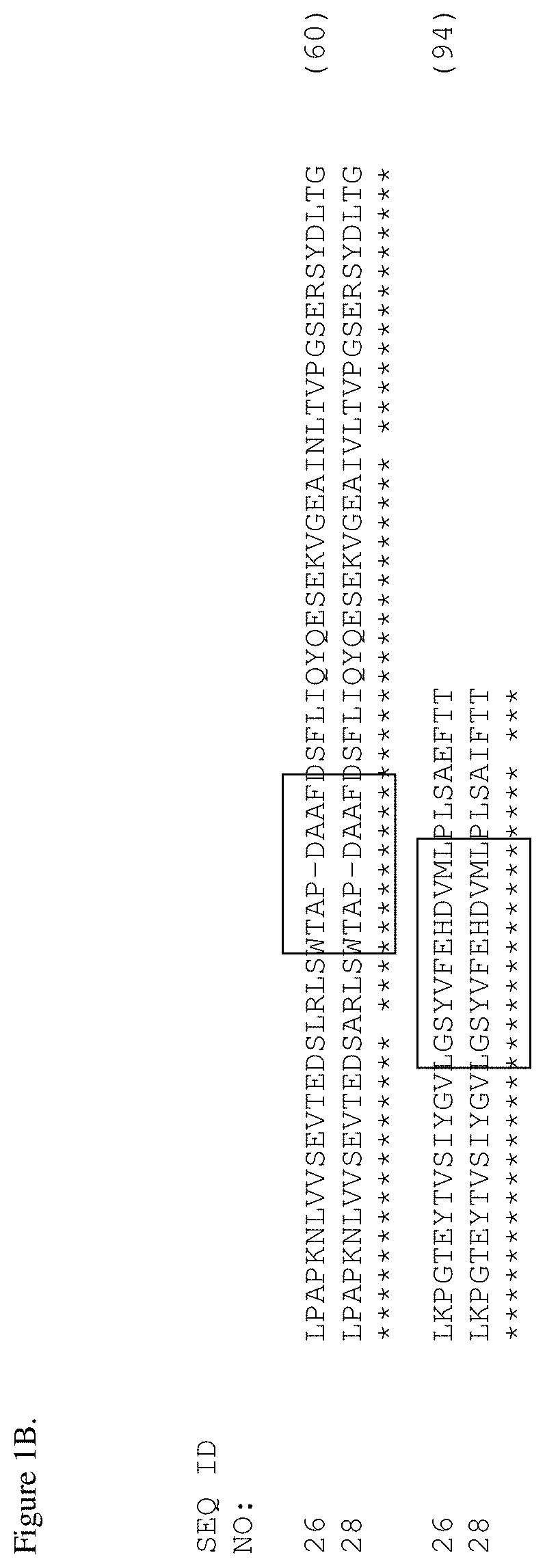
[0037] FIGS. 1A and 1B. Amino acid alignment of the EGFR-binding FN3 domains. The BC and FG loops are boxed at residues 22-28 and 75-86 of SEQ ID NO: 18. Some variants include thermal stability improving L17A, N46K and E86I substitutions (residue numbering according to Tencon SEQ ID NO: 1).
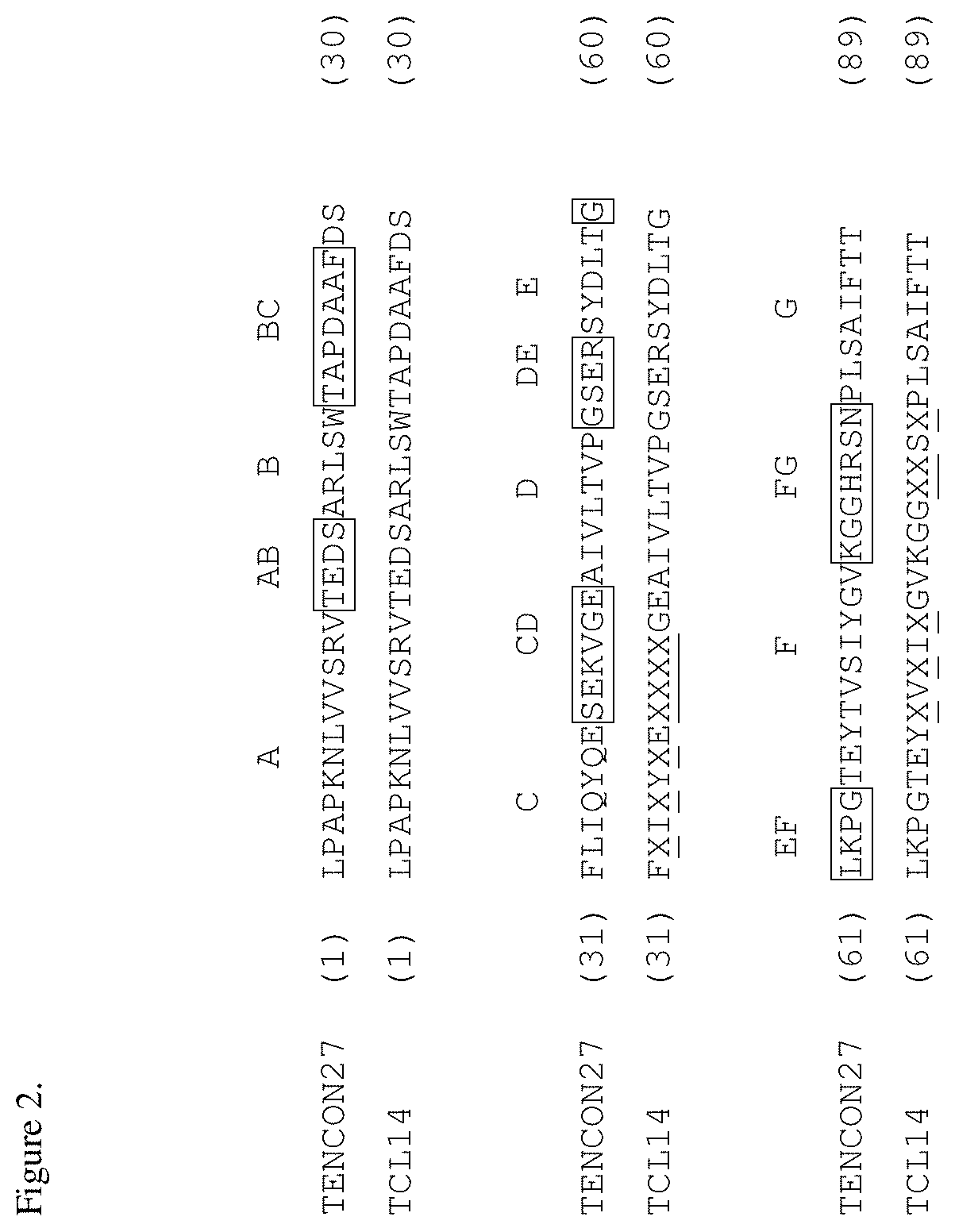
[0038] FIG. 2. Sequence alignment of the Tencon27 scaffold (SEQ ID NO: 99) and a TCL14 library (SEQ ID NO: 100) having randomized C-CD-F-FG alternative surface. The loop residues are boxed. Loops and strands are indicated above the sequences.
[0039] FIG. 3. Sequence alignment of the c-Met-binding FN3 domains. The C loop and the CD strand and the F loop and the FG strand are boxed and span residues 29-43 and 65-81.
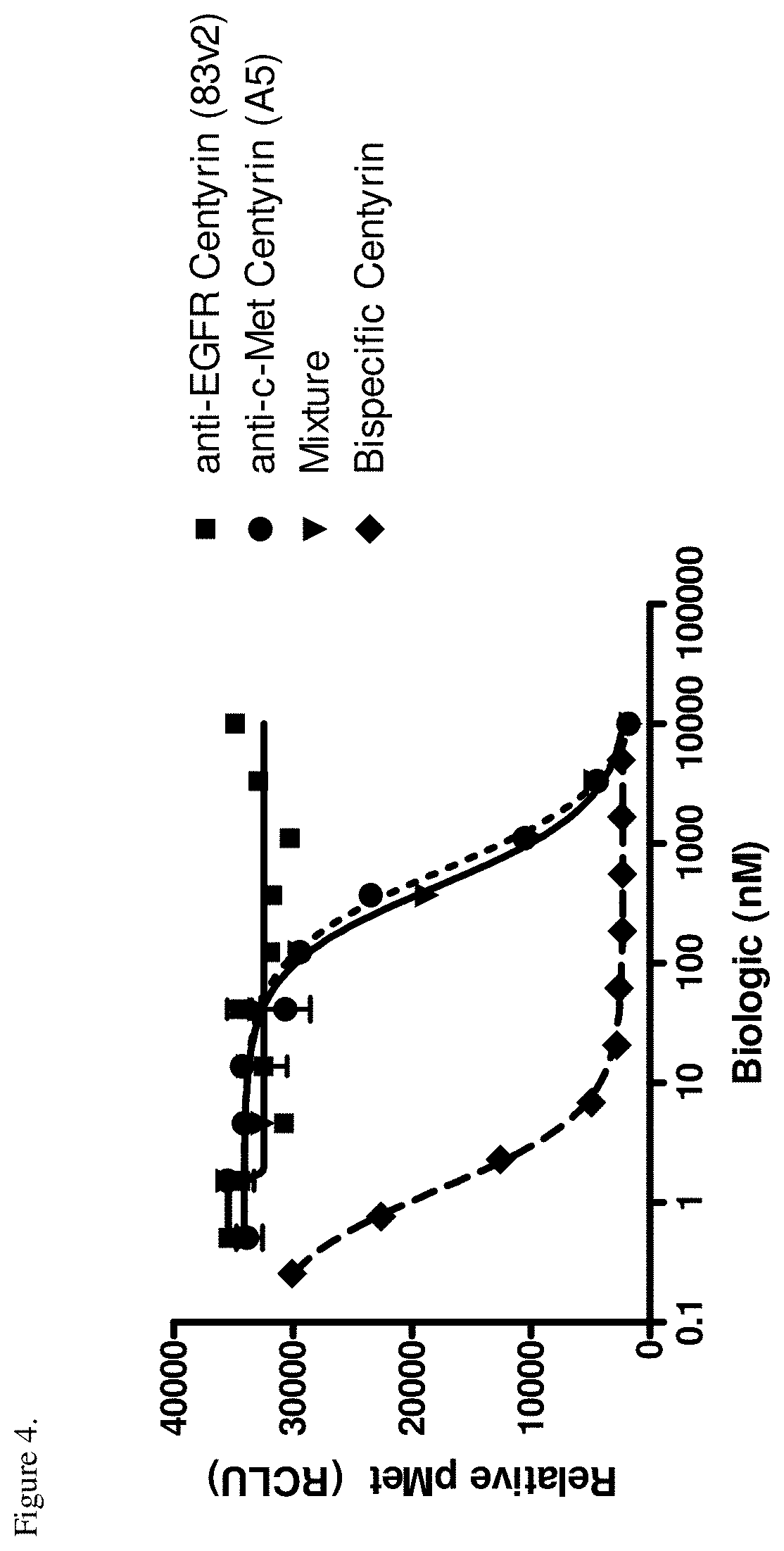
[0040] FIG. 4. Inhibition of c-Met phosphorylation in NCI-H292 cells pre-treated with monospecific or bispecific FN3 domain containing molecules and stimulated with HGF is shown. Substantial increase in the potency of the bispecific EGFR/c-Met molecule (ECB1) was observed when compared to a monospecific c-Met-binding FN3 domain (P114AR5P74-A5, shown as A5 in the Figure) on its own or in combination with an EGFR-binding FN3 domain (P54AR4-83v2, shown as 83v2 in the Figure).
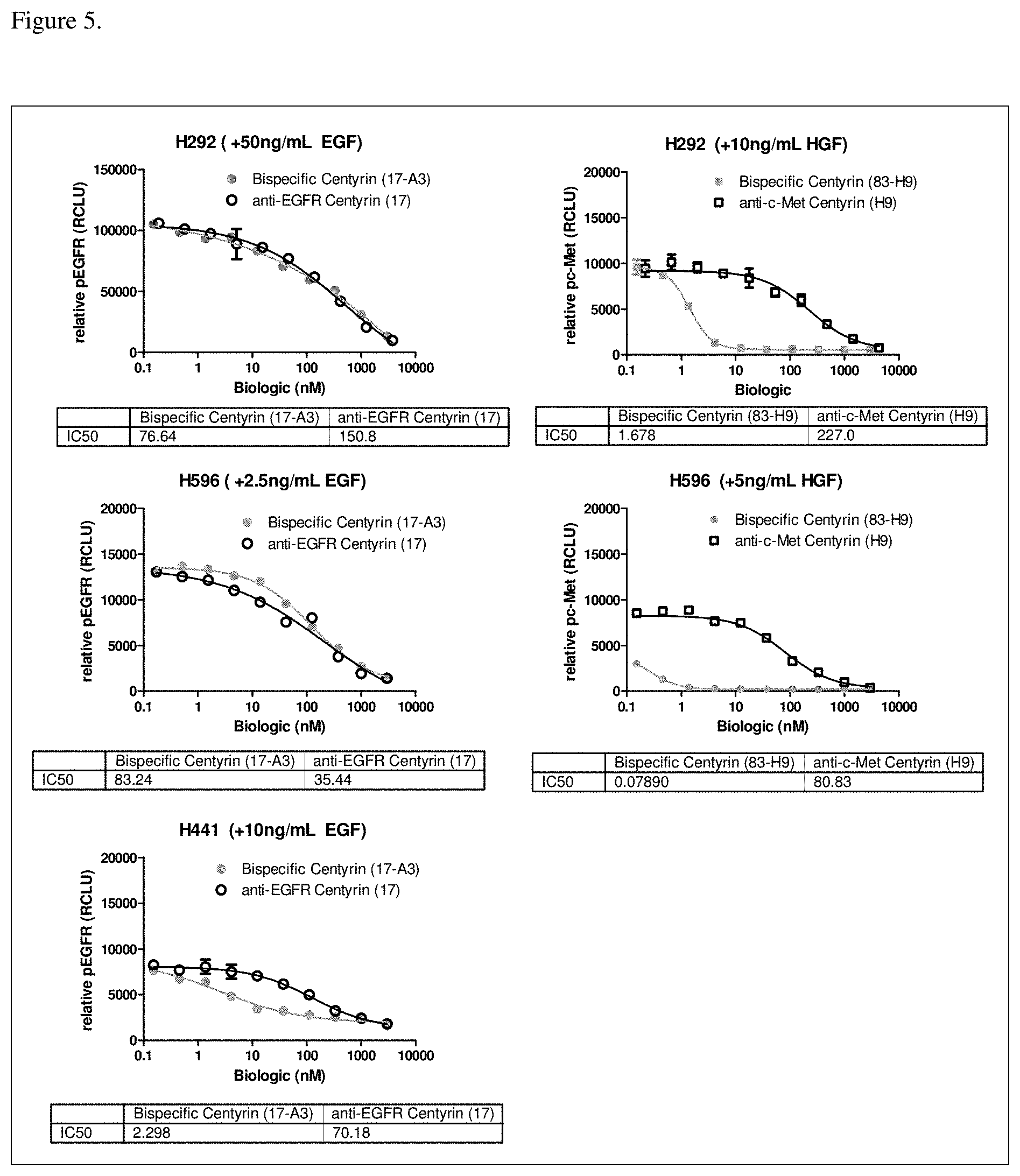
[0041] FIG. 5. Inhibition of EGFR and c-Met phosphorylation in cells pre-treated with monospecific or bispecific FN3 domain containing molecules. In cell lines expressing high levels of EGFR, NCI-H292 (FIG. 5A) and H596 (FIG. 5B), anti-EGFR monospecific and bispecific FN3 domain containing molecules are equally potent at decreasing EGFR phosphorylation. In cell lines expressing low levels of EGFR relative to c-Met, NCI-H441 (FIG. 5C), bispecific EGFR/c-Met molecules improve the potency for inhibition of EGFR phosphorylation compared to the monospecific EGFR-binding FN3 domain alone. In cell lines with low levels of c-Met, relative to EGFR, NCI-H292 (FIG. 5D) and H596 (FIG. 5E), inhibition of c-Met phosphorylation is significantly potentiated with bispecific EGFR/c-Met molecule, compared to monospecific c-Met-binding FN3 domain only. Molecules used in the study were: bispecific ECB5 (shown as 17-A3 in the Figure), monospecific EGFR-binding FN3 domain P53A1R5-17 (shown as "17" in the Figure), bispecific EGFR/c-Met molecule ECB3 (shown as 83-H9 in the Figure), and monospecific c-Met binding FN3 domain P114AR7P93-H9 (shown as H9 in the Figure).
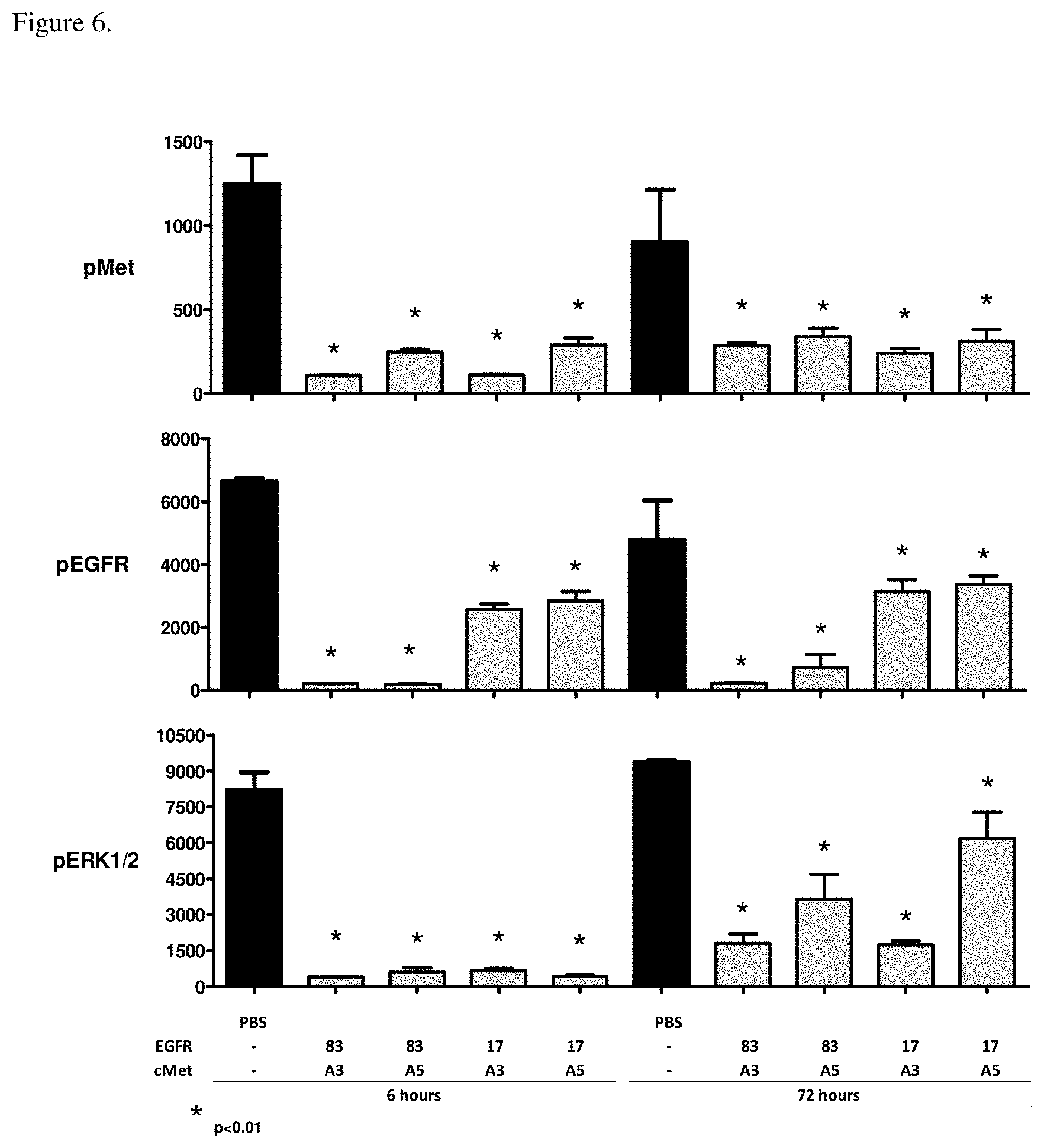
[0042] FIG. 6. Pharmacodynamic signaling in tumors isolated from mice dosed with bispecific EGFR/c-Met molecules for 6 h or 72 h. All molecules significantly reduced c-Met, EGFR and ERK phosphorylation at 6 h and 72 h, the degree if inhibition was dependent on the affinity of the FN3 domains to EGFR and/or c-Met. Bispecific molecules were generated by joining EGFR-binding FN3 domain with a high ("83" in the Figure is p54AR4-83v2) or medium ("17v2" in the Figure is P53A1R5-17v2) affinity to a c-Met-binding FN3 domain with high ("A3" in the Figure is P114AR7P94-A3) or medium ("A5" in the Figure is P114AR5P74-A5) affinity.
[0043] FIG. 7. Plasma (top) and tumor (bottom) accumulation of bispecific EGFR/cMet molecules of variable affinities linked to an albumin binding domain (ABD) are shown 6 h (left) and 72 h (right) after IP dosing. Six hours after dosing, tumor accumulation is maximal in mice dosed with a bispecific molecule harboring a medium affinity EGFR-binding FN3 domain (17v2) or high affinity EGFR binding domain (83v2). The bispecific molecules incorporated high or medium affinity EGFR or c-Met binding FN3 domains as follows: 83v2-A5-ABD (ECB18; high/medium for EGFR/cMet) 83v2-A3-ABD (ECB38; high/high) 17v2-A5 (ECB28; medium/medium) 17v2-A3-ABD (ECB39; medium/high). In the figure, 83v2 refers to p54AR4-83v2; 17v2 refers to p53A1R5-17v2; A3 refers to p114AR7P94-A3 and A5 refers to p114AR5P74-A5.
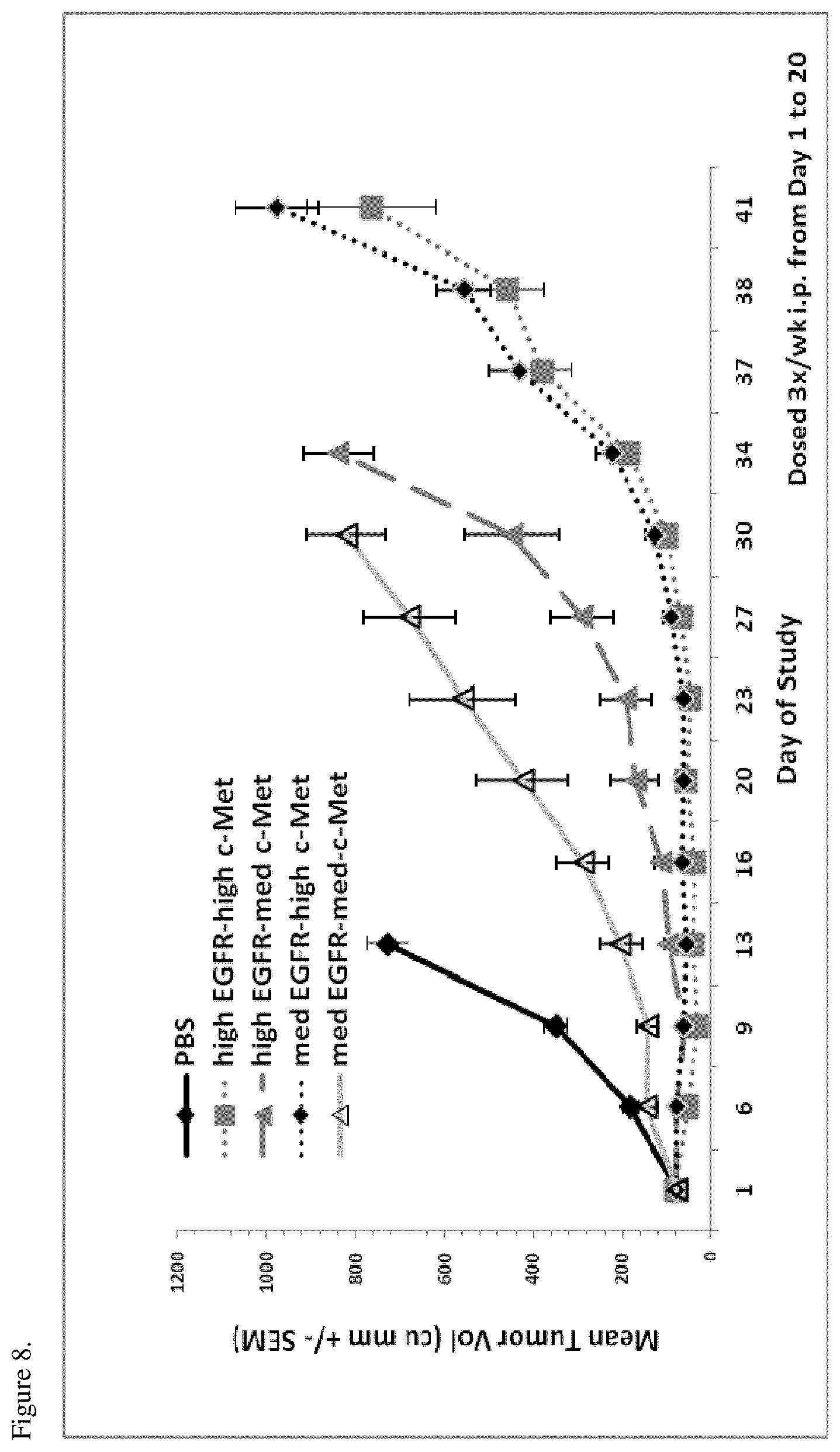
[0044] FIG. 8. H292-HGF tumor xenografts were implanted into SCID Beige mice. When tumors reached an average volume of approximately 80 mm.sup.3, mice were dosed three times per week with bispecific EGFR/c-Met molecules (25 mg/kg) or PBS vehicle. All bispecific molecules reduced tumor growth, the tumor growth inhibition (TGI) being dependent on the affinities of the molecules for c-Met and EGFR (high EGFR-high cMet refers to p54AR4-83v2-p114AR7P94-A3 (ECB38); high EGFR-med cMet refers to p54AR4-83v2-p114AR5P74-A5 (ECB18); med EGFR-high cMet refers to p53A1R5-17v2-p114AR7P94-A3 (ECB39); med EGFR-med-cMet refers to p53A1R5-17-p114AR5P74-A 5 (ECB28)).
[0045] FIG. 9. H292-HGF tumor xenografts were implanted into SCID Beige mice and they were treated with different therapies. The anti-tumor activity of the therapies is shown (bispecific EGFR/c-Met molecule refers to p54AR4-83v2-p114AR7P94-A3-ABD (ECB38); the other therapies are crizotinib, erlotinib, cetuximab, and the combination of crizotinib and erlotinib).
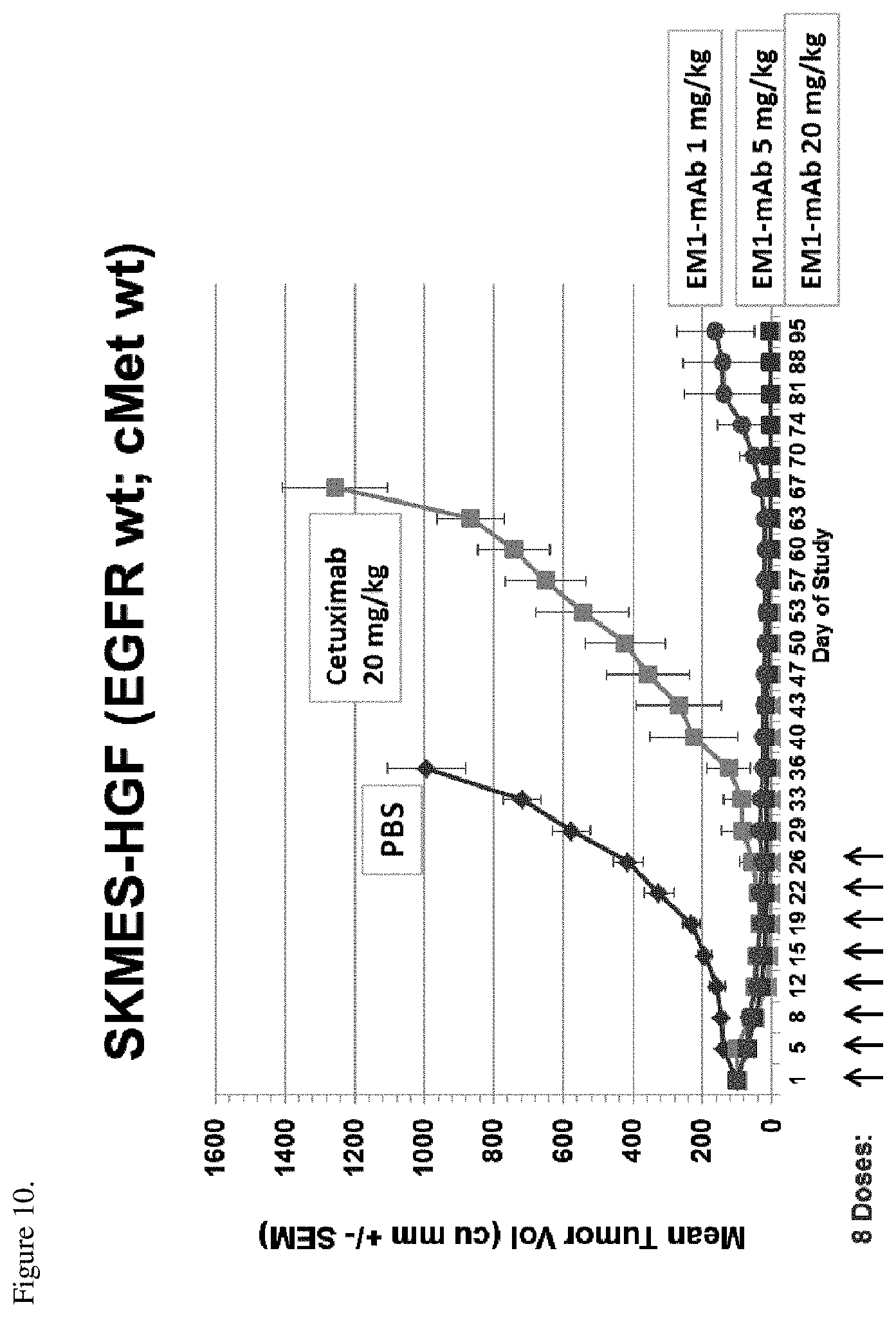
[0046] FIG. 10. SKMES-HGF tumor xenografts were implanted into SCID Beige mice and the mice were treated with different therapies. The anti-tumor activity of the therapies is shown as change in tumor size (mm.sup.3) over time. The bispecific EGFR/c-Met antibody EM1-mAb was dosed intraperitoneally (i.p.) twice a week at either 20 mg/kg, 5 mg/kg, or 1 mg/kg; cetuximab was dosed i.p. twice a week at 20 mg/kg. Arrows in the figure show the administration days. Numbers after the antibodies indicated the administered dose.
[0047] FIG. 11. HCC827 tumor xenografts were implanted into nude mice and the mice were treated with erlotinib or EM1-mAb at indicated doses. EM1-mAb was dosed biweekly and erlotinib once a day for four weeks. Arrows in the figure show the administration days. The anti-tumor activity of the therapies is shown as change in tumor size (mm.sup.3) over time.
[0048] FIG. 12. SNU-5 tumor xenografts were implanted into CB17/SCID mice and the mice were treated with 10 mg/kg cetuximab or 10 mg/kg or 1 mg/kg EM1-mAb. Antibodies were dosed biweekly for four weeks. Arrows in the figure show the administration days. The anti-tumor activity of the therapies is shown as change in tumor size (mm.sup.3) over time.
[0049] FIG. 13. H1975-HGF tumor xenografts were implanted into nude mice and the mice were treated with 10 mg/kg cetuximab, 10 mg/kg EM1-mAb, 50 mg/kg erlotinib, 15 mg/kg afatinib, or a combination of 10 mg/kg EM1-mAb and 15 mg/kg afatinib. Antibodies were dosed biweekly and the small molecules once a day for three weeks. Arrows in the figure show the administration days. The anti-tumor activity of the therapies is shown as change in tumor size (mm.sup.3) over time.
[0050] FIG. 14. HCC827-ER1 tumor xenografts were implanted into nude mice and the mice were treated with 10 mg/kg EM1-mAb, 25 mg/kg erlotinib, or a combination of the two. EM1-mAb was dosed biweekly and erlotinib once a day for 19 days. Arrows in the figure show the administration days. The anti-tumor activity of the therapies is shown as change in tumor size (mm.sup.3) over time.
[0051] FIG. 15. Average EGFR and c-Met levels in tumor lysates isolated from H1975 HGF tumor xenografts implanted into SCID Beige mice after administration of a single dose of 20 mg/kg EM1-mAb. Receptor levels are shown as % of PBS control at indicated times post-treatment.
[0052] FIG. 16. H1975-HGF tumor xenografts were implanted into nude mice and the mice were treated with 10 mg/kg EM1-mAb or 10 mg/kg EM1-mAb variant IgG2 V234A/G237A/P238S/H268AN309L/A330S/P331S having no Fc receptor binding and lacking effector functions. Antibodies were dosed biweekly at indicated days. The anti-tumor activity of the therapies is shown as change in tumor size (mm.sup.3) over time.
[0053] FIG. 17A. Correlation of EGFR surface density and inhibition of EGFR phosphorylation by the bispecific EM-1 mAb. IC.sub.50 values for inhibition of EGFR phosphorylation were plotted vs. EGFR molecules on the surface expressed as antibodies bound per cell (ABC). Pearson correlation p=0.00082, r.sup.2=0.7820.
[0054] FIG. 17B. Correlation of receptor density and inhibition of c-Met phosphorylation by the bispecific EM-1 mAb. IC.sub.50 values for inhibition of c-Met phosphorylation were plotted vs. c-Met molecules on the surface expressed as ABC. Pearson correlation p<0.0001, r.sup.2=0.9336.
[0055] FIG. 18. The bispecific EGFR/c-Met EM-1 mAb induced heterodimerization of EGFR and c-Met assessed in .beta.-galactosidase fragment mediated complementation assay in U-2 OS cells stably expressing EGFR and c-Met .beta.-gal fusion proteins. Control monovalent EGFR- or c-Met antibodies or parental bivalent anti-EGFR or c-Met antibodies 2F8 or 069 had no effect. E1-F405L-gp120-K409R (.circle-solid.), M1-K409R-gp120-F405L (.quadrature.), a combination of E1-F405L-gp120-K409R and M1-K409R-gp120-F405L (), EM-1 bispecific mAb (.diamond-solid.), anti-cMet parental 069 mAb (.diamond.), anti-EGFR parental 2F8mAb (.tangle-solidup.).
[0056] FIG. 19A. Inhibition of c-Met phosphorylation in H292 cells relative to a HGF-treated control. The ratio of EGFR to c-Met surface expression in H292 cells was 5.72. Negative control (.diamond-solid.), E1-F405L-gp120-K409R (.circle-solid.), M1-K409R-gp120-F405L (.box-solid.), EM-1 bispecific mAb (.tangle-solidup.). The dotted lines indicate the levels of the MSD signal from the EGF-treated control without antibody or unstimulated control as shown in the Figure. ug/mL indicates the antibody log concentration in .mu.g/ml.
[0057] FIG. 19B. Inhibition of EGFR phosphorylation in H292 cells relative to an EGFR-treated control. Negative control (.diamond-solid.), E1-F405L-gp120-K409R (.circle-solid.), M1-K409R-gp120-F405L (.box-solid.), EM-1 bispecific mAb (.tangle-solidup.). The dotted lines indicate the levels of the MSD signal from the EGF-treated control without antibody or unstimulated control as shown in the Figure. ug/mL indicates the antibody log concentration in .mu.g/ml.
[0058] FIG. 20. Inhibition of EGFR phosphorylation in SNU-5 cells relative to an EGF-treated control. Negative control (.diamond-solid.), E1-F405L-gp120-K409R (.circle-solid.), M1-K409R-gp120-F405L (.box-solid.), EM-1 bispecific mAb (.tangle-solidup.). The dotted lines indicaet the levels of the MSD signal from the negative controls or unstimulated control as shown in the Figure. ug/mL indicates the antibody log concentration in .mu.g/ml.
[0059] FIG. 21. Inhibition of ligand-induced receptor phosphorylation in H1993 cells relative to an EGF-treated control. E1-F405L-gp120-K409R (.circle-solid.), M1-K409R-gp120-F405L (.box-solid.), EM-1 bispecific mAb (.tangle-solidup.), negative control (.diamond-solid.), combination of E1-F405L-gp120-K409R and M1-K409R-gp120-F405L (). X-axis indicates the antibody log nM concentration. The dotted lines indicate the levels of the MSD signal from the unstimulated (vehicle) control or EGF-treated control without antibody.
DETAILED DESCRIPTION OF THE INVENTION
[0060] The term "fibronectin type III (FN3) domain" (FN3 domain) as used herein refers to a domain occurring frequently in proteins including fibronectins, tenascin, intracellular cytoskeletal proteins, cytokine receptors and prokaryotic enzymes (Bork and Doolittle, Proc Nat Acad Sci USA 89:8990-8994, 1992; Meinke et al., J Bacteriol 175:1910-1918, 1993; Watanabe et al., J Biol Chem 265:15659-15665, 1990). Exemplary FN3 domains are the 15 different FN3 domains present in human tenascin C, the 15 different FN3 domains present in human fibronectin (FN), and non-natural synthetic FN3 domains as described for example in U.S. Pat. Publ. No. 2010/0216708. Individual FN3 domains are referred to by domain number and protein name, e.g., the 3.sup.rd FN3 domain of tenascin (TN3), or the 10.sup.th FN3 domain of fibronectin (FN10).
[0061] The term "substituting" or "substituted" or `mutating" or "mutated" as used herein refers to altering, deleting or inserting one or more amino acids or nucleotides in a polypeptide or polynucleotide sequence to generate a variant of that sequence.
[0062] The term "randomizing" or "randomized" or "diversified" or "diversifying" as used herein refers to making at least one substitution, insertion or deletion in a polynucleotide or polypeptide sequence.
[0063] "Variant" as used herein refers to a polypeptide or a polynucleotide that differs from a reference polypeptide or a reference polynucleotide by one or more modifications for example, substitutions, insertions or deletions.
[0064] The term "specifically binds" or "specific binding" as used herein refers to the ability of an FN3 domain, a bispecific agent that specifically binds EGFR and c-Met, or a bispecific EGFR/c-Met antibody of the invention to bind to a predetermined antigen with a dissociation constant (K.sub.D) of about 1.times.10.sup.-6 M or less, for example about 1.times.10.sup.-7 M or less, about 1.times.10.sup.-8 M or less, about 1.times.10.sup.-9 M or less, about 1.times.10.sup.-10 M or less, about 1.times.10.sup.-11 M or less, about 1.times.10.sup.-12 M or less, or about 1.times.10.sup.-13 M or less. Typically the FN3 domain, the bispecific agent that specifically binds EGFR and c-Met or the bispecific EGFR/c-Met antibody of the invention binds to a predetermined antigen (i.e. EGFR or c-Met) with a K.sub.D that is at least ten fold less than its K.sub.D for a nonspecific antigen (for example BSA or casein) as measured by surface plasmon resonance using for example a Proteon Instrument (BioRad). Thus, the bispecific EGFR/c-Met FN3 domain containing molecule, the bispecific agent that specifically binds EGFR and c-Met or the bispecific EGFR/c-Met antibody of the invention specifically binds to each EGFR and c-Met with a binding affinity (K.sub.D) of at least about 1.times.10.sup.-6 M or less, for example about 1.times.10.sup.-7 M or less, about 1.times.10.sup.-8 M or less, about 1.times.10.sup.-9 M or less, about 1.times.10.sup.-10 M or less, about 1.times.10.sup.-11 M or less, about 1.times.10.sup.-12 M or less, or about 1.times.10.sup.-13 M or less. The bispecific EGFR/c-Met FN3 domain containing molecule, the bispecific agent that specifically binds EGFR and c-Met or the bispecific EGFR/c-Met antibody of the invention that specifically binds to a predetermined antigen may, however, have cross-reactivity to other related antigens, for example to the same predetermined antigen from other species (homologs).
[0065] The term "library" refers to a collection of variants. The library may be composed of polypeptide or polynucleotide variants.
[0066] The term "stability" as used herein refers to the ability of a molecule to maintain a folded state under physiological conditions such that it retains at least one of its normal functional activities, for example, binding to a predetermined antigen such as EGFR or c-Met.
[0067] "Epidermal growth factor receptor" or "EGFR" as used here refers to the human EGFR (also known as HER1 or ErbB1 (Ullrich et al., Nature 309:418-425, 1984) having the amino acid sequence shown in SEQ ID NO: 73 and in GenBank accession number NP_005219, as well as naturally-occurring variants thereof. Such variants include well-known EGFRvIII and other alternatively spliced variants (e.g., as identified by SwissProt Accession numbers P00533-1 (wild type; identical to SEQ ID NO: 73 and NP_005219), P00533-2 (F404L/L405S), P00533-3 (628-705: CTGPGLEGCP . . . GEAPNQALLR.fwdarw.PGNESLKAML . . . SVIITASSCH and 706-1210 deleted), P00533-4 (C628S and 629-1210 deleted), variants GlnQ98, R266, K521, 1674, G962, and P988 (Livingston et al., NIEHS-SNPs, environmental genome project, NIEHS ES15478), T790M, L858R/T790M and del(E746, A750).
[0068] "EGFR ligand" as used herein encompasses all (e.g., physiological) ligands for EGFR, including EGF, TGF.alpha., heparin binding EGF (HB-EGF), amphiregulin (AR), and epiregulin (EPI).
[0069] "Epidermal growth factor" (EGF) as used herein refers to the well-known 53 amino acid human EGF having the amino acid sequence shown in SEQ ID NO: 74.
[0070] "Hepatocyte growth factor receptor" or "c-Met" as used herein refers to the human c-Met having the amino acid sequence shown in SEQ ID NO: 101 or in GenBank Accession No: NP 001120972 and natural variants thereof.
[0071] "Hepatocyte growth factor" (HGF) as used herein refers to the well-known human HGF having the amino acid sequence shown in SEQ ID NO: 102 which is cleaved to form a dimer of an alpha and beta chain linked by a disulfide bond.
[0072] "Blocks binding" or "inhibits binding", as used herein interchangeably refers to the ability of the FN3 domains, the bispecific EGFR/c-Met FN3 domain containing molecule, the bispecific agent that specifically binds EGFR and c-Met or the bispecific EGFR/c-Met antibody of the invention to block or inhibit binding of the EGFR ligand such as EGF to EGFR and/or HGF to c-Met, and encompass both partial and complete blocking/inhibition. The blocking/inhibition of EGFR ligand such as EGF to EGFR and/or HGF to c-Met by the FN3 domains, the bispecific EGFR/c-Met FN3 domain containing molecule, the bispecific agent that specifically binds EGFR and c-Met or the bispecific EGFR/c-Met antibody of the invention reduces partially or completely the normal level of EGFR signaling and/or c-Met signaling when compared to the EGFR ligand binding to EGFR and/or HGF binding to c-Met without blocking or inhibition. The FN3 domains, the bispecific EGFR/c-Met FN3 domain containing molecule, the bispecific agent that specifically binds EGFR and c-Met or the bispecific EGFR/c-Met antibody of the invention "blocks binding" of the EGFR ligand such as EGF to EGFR and/or HGF to c-Met when the inhibition is at least 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% or 100%. Inhibition of binding can be measured using well known methods, for example by measuring inhibition of binding of biotinylated EGF on EGFR expressing A431 cells exposed to the FN3 domain, the bispecific EGFR/c-Met FN3 domain containing molecule, the bispecific agent that specifically binds EGFR and c-Met or the bispecific EGFR/c-Met antibody of the invention using FACS, and using methods described herein, or measuring inhibition of binding of biotinylated HGF on c-Met extracellular domain using well known methods and methods described herein.
[0073] The term "EGFR signaling" refers to signal transduction induced by EGFR ligand binding to EGFR resulting in autophosphorylation of at least one tyrosine residue in the EGFR. An exemplary EGFR ligand is EGF.
[0074] "Neutralizes EGFR signaling" as used herein refers to the ability of the FN3 domains, the bispecific EGFR/c-Met FN3 domain containing molecule, the bispecific agent that specifically binds EGFR and c-Met or the bispecific EGFR/c-Met antibody of the invention to inhibit EGFR signaling induced by EGFR ligand such as EGF by at least 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% or 100%.
[0075] The term "c-Met signaling" refers to signal transduction induced by HGF binding to c-Met resulting in autophosphorylation of at least one tyrosine residue in the c-Met. Typically at least one tyrosine residue at positions 1230, 1234, 1235 or 1349 is autophosphorylated upon HGF binding.
[0076] "Neutralizes c-Met signaling" as used herein refers to the ability of the FN3 domain, the bispecific EGFR/c-Met FN3 domain containing molecule, the bispecific agent that specifically binds EGFR and c-Met or the bispecific EGFR/c-Met antibody of the invention to inhibit c-Met signaling induced by HGF by at least 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% or 100%.
[0077] "Overexpress", "overexpressed" and "overexpressing" as used herein interchangeably refer to a cancer or malignant cell that has measurably higher levels of EGFR and/or c-Met on the surface compared to a normal cell of the same tissue type. Such overexpression may be caused by gene amplification or by increased transcription or translation. EGFR and/or c-Met expression and overexpression can be measured using well know assays using for example ELISA, immunofluorescence, flow cytometry or radioimmunoassay on live or lysed cells. Alternatively, or additionally, levels of EGFR and/or c-Met-encoding nucleic acid molecules may be measured in the cell for example using fluorescent in situ hybridization, Southern blotting, or PCR techniques. EGFR and/or c-Met is overexpressed when the level of EGFR and/or c-Met on the surface of the cell is at least 1.5-fold higher when compared to the normal cell.
[0078] "Tencon" as used herein refers to the synthetic fibronectin type III (FN3) domain having the sequence shown in SEQ ID NO: 1 and described in U.S. Pat. Publ. No. US2010/0216708.
[0079] A "cancer cell" or a "tumor cell" as used herein refers to a cancerous, pre-cancerous or transformed cell, either in vivo, ex vivo, and in tissue culture, that has spontaneous or induced phenotypic changes that do not necessarily involve the uptake of new genetic material. Although transformation can arise from infection with a transforming virus and incorporation of new genomic nucleic acid, or uptake of exogenous nucleic acid, it can also arise spontaneously or following exposure to a carcinogen, thereby mutating an endogenous gene. Transformation/cancer is exemplified by, e.g., morphological changes, immortalization of cells, aberrant growth control, foci formation, proliferation, malignancy, tumor specific marker levels, invasiveness, tumor growth or suppression in suitable animal hosts such as nude mice, and the like, in vitro, in vivo, and ex vivo (Freshney, Culture of Animal Cells: A Manual of Basic Technique (3rd ed. 1994)).
[0080] The term "vector" means a polynucleotide capable of being duplicated within a biological system or that can be moved between such systems. Vector polynucleotides typically contain elements, such as origins of replication, polyadenylation signal or selection markers that function to facilitate the duplication or maintenance of these polynucleotides in a biological system. Examples of such biological systems may include a cell, virus, animal, plant, and reconstituted biological systems utilizing biological components capable of duplicating a vector. The polynucleotide comprising a vector may be DNA or RNA molecules or a hybrid of these.
[0081] The term "expression vector" means a vector that can be utilized in a biological system or in a reconstituted biological system to direct the translation of a polypeptide encoded by a polynucleotide sequence present in the expression vector.
[0082] The term "polynucleotide" means a molecule comprising a chain of nucleotides covalently linked by a sugar-phosphate backbone or other equivalent covalent chemistry. Double and single-stranded DNAs and RNAs are typical examples of polynucleotides.
[0083] "Complementary DNA" or "cDNA" refers to the well-known synthetic polynucleotide that shares the arrangement of sequence elements found in native mature mRNA species with contiguous exons, with the intervening introns present in genomic DNA are removed. The codons encoding the initiator methionine may or may not be present in cDNA. cDNA may be synthesized for example by reverse transcription or synthetic gene assembly.
[0084] "Synthetic" or "non-natural" or "artificial" as used herein refers to a polynucleotide or a polypeptide molecule not present in nature.
[0085] The term "polypeptide" or "protein" means a molecule that comprises at least two amino acid residues linked by a peptide bond to form a polypeptide. Small polypeptides of less than about 50 amino acids may be referred to as "peptides".
[0086] The term "bispecific EGFR/c-Met molecule" or "bispecific EGFR/c-Met FN3 domain containing molecule" as used herein refers to a molecule comprising an EGFR binding FN3 domain and a distinct c-Met binding FN3 domain that are covalently linked together either directly or via a linker. An exemplary bispecific EGFR/c-Met binding molecule comprises a first FN3 domain specifically binding EGFR and a second FN3 domain specifically binding c-Met.
[0087] "Valent" as used herein refers to the presence of a specified number of binding sites specific for an antigen in a molecule. As such, the terms "monovalent", "bivalent", "tetravalent", and "hexavalent" refer to the presence of one, two, four and six binding sites, respectively, specific for an antigen in a molecule.
[0088] "Mixture" as used herein refers to a sample or preparation of two or more FN3 domains not covalently linked together. A mixture may consist of two or more identical FN3 domains or distinct FN3 domains. Mixture as used herein also refers to a sample or preparation of two or more monovalent antibodies that are monovalent towards EGFR and/or monovalent towards c-Met.
[0089] The term "bispecific agent that specifically binds EGFR and c-Met" as used herein refers to a molecule comprising a first domain that specifically binds EGFR and a second domain that specifically binds c-Met. An exemplary agent that specifically binds EGFR and c-Met is a bispecific antibody. Another exemplary bispecific agent that specifically binds EGFR and c-Met is a molecule comprising an EGFR binding FN3 domain and a distinct c-Met binding FN3 domain. The bispecific agent that specifically binds EGFR and c-Met may be composed of a single polypeptide or more than one polypeptide.
[0090] The term "bispecific anti-EGFR/c-Met antibody" or "bispecific EGFR/c-Met antibody" as used herein refers to a bispecific antibody having a first domain that specifically binds EGFR and a second domain that specifically binds c-Met. The domains specifically binding EGFR and c-Met are typically VH/VL pairs, and the bispecific anti-EGFR/c-Met antibody is monovalent in terms of binding to EGFR and c-Met.
[0091] The term "antibodies" as used herein is meant in a broad sense and includes immunoglobulin molecules including polyclonal antibodies, monoclonal antibodies including murine, human, human-adapted, humanized and chimeric monoclonal antibodies, antibody fragments, bispecific or multispecific antibodies, dimeric, tetrameric or multimeric antibodies, and single chain antibodies.
[0092] Immunoglobulins can be assigned to five major classes, namely IgA, IgD, IgE, IgG and IgM, depending on the heavy chain constant domain amino acid sequence. IgA and IgG are further sub-classified as the isotypes IgA.sub.1, IgA.sub.2, IgG.sub.1, IgG.sub.2, IgG.sub.3 and IgG.sub.4. Antibody light chains of any vertebrate species can be assigned to one of two clearly distinct types, namely kappa (.kappa.) and lambda (.lamda.), based on the amino acid sequences of their constant domains.
[0093] The term "antibody fragments" refers to a portion of an immunoglobulin molecule that retains the heavy chain and/or the light chain antigen binding site, such as heavy chain complementarity determining regions (HCDR) 1, 2 and 3, light chain complementarity determining regions (LCDR) 1, 2 and 3, a heavy chain variable region (VH), or a light chain variable region (VL). Antibody fragments include a Fab fragment, a monovalent fragment consisting of the VL, VH, CL and CHI domains; a F(ab).sub.2 fragment, a bivalent fragment comprising two Fab fragments linked by a disulfide bridge at the hinge region; a Fd fragment consisting of the VH and CHI domains; a Fv fragment consisting of the VL and VH domains of a single arm of an antibody; a domain antibody (dAb) fragment (Ward et al (1989) Nature 341:544-546), which consists of a VH domain. VH and VL domains can be engineered and linked together via a synthetic linker to form various types of single chain antibody designs where the VH/VL domains pair intramolecularly, or intermolecularly in those cases when the VH and VL domains are expressed by separate single chain antibody constructs, to form a monovalent antigen binding site, such as single chain Fv (scFv) or diabody; described for example in PCT Intl. Publ. Nos. WO1998/44001, WO1988/01649, WO1994/13804, and WO1992/01047. These antibody fragments are obtained using well known techniques known to those of skill in the art, and the fragments are screened for utility in the same manner as are full length antibodies.
[0094] The phrase "isolated antibody" refers to an antibody or antibody fragment that is substantially free of other antibodies having different antigenic specificities (e.g., an isolated bispecific antibody specifically binding EGFR and c-Met is substantially free of antibodies that specifically bind antigens other than human EGFR and c-Met). An isolated antibody that specifically binds EGFR and c-Met, however, can have cross-reactivity to other antigens, such as orthologs of human EGFR and/or c-Met, such as Macaca fascicularis (cynomolgus) EGFR and/or c-Met. Moreover, an isolated antibody may be substantially free of other cellular material and/or chemicals.
[0095] An antibody variable region consists of a "framework" region interrupted by three "antigen binding sites". The antigen binding sites are defined using various terms: (i) Complementarity Determining Regions (CDRs), three in the VH (HCDR1, HCDR2, HCDR3), and three in the VL (LCDR1, LCDR2, LCDR3), are based on sequence variability (Wu and Kabat J Exp Med 132:211-50, 1970; Kabat et al Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md., 1991). (ii) "Hypervariable regions", "HVR", or "HV", three in the VH (H1, H2, H3) and three in the VL (L1, L2, L3), refer to the regions of an antibody variable domains which are hypervariable in structure as defined by Chothia and Lesk (Chothia and Lesk Mol Biol 196:901-17, 1987). Other terms include "IMGT-CDRs" (Lefranc et al., Dev Comparat Immunol 27:55-77, 2003) and "Specificity Determining Residue Usage" (SDRU) (Almagro Mol Recognit 17:132-43, 2004). The International ImMunoGeneTics (IMGT) database (http://www_imgt_org) provides a standardized numbering and definition of antigen-binding sites. The correspondence between CDRs, HVs and IMGT delineations is described in Lefranc et al., Dev Comparat Immunol 27:55-77, 2003.
[0096] "Chothia residues" as used herein are the antibody VL and VH residues numbered according to Al-Lazikani (Al-Lazikani et al., J Mol Biol 273:927-48, 1997).
[0097] "Framework" or "framework sequences" are the remaining sequences of a variable region other than those defined to be antigen binding sites. Because the antigen binding sites can be defined by various terms as described above, the exact amino acid sequence of a framework depends on how the antigen-binding site was defined.
[0098] "Humanized antibody" refers to an antibody in which the antigen binding sites are derived from non-human species and the variable region frameworks are derived from human immunoglobulin sequences. Humanized antibodies may include substitutions in the framework regions so that the framework may not be an exact copy of expressed human immunoglobulin or germline gene sequences.
[0099] "Human-adapted" antibodies or "human framework adapted (HFA)" antibodies refers to humanized antibodies adapted according to methods described in U.S. Pat. Publ. No. US2009/0118127. Human-adapted antibodies are humanized by selecting the acceptor human frameworks based on the maximum CDR and FR similarities, length compatibilities and sequence similarities of CDR1 and CDR2 loops and a portion of light chain CDR3 loops.
[0100] "Human antibody" refers to an antibody having heavy and light chain variable regions in which both the framework and the antigen binding sites are derived from sequences of human origin. If the antibody contains a constant region, the constant region also is derived from sequences of human origin.
[0101] Human antibody comprises heavy or light chain variable regions that are "derived from" sequences of human origin if the variable regions of the antibody are obtained from a system that uses human germline immunoglobulin or rearranged immunoglobulin genes. Such systems include human immunoglobulin gene libraries displayed on phage, and transgenic non-human animals such as mice carrying human immunoglobulin loci as described herein. "Human antibody" may contain amino acid differences when compared to the human germline or rearranged immunoglobulin sequences due to for example naturally occurring somatic mutations or intentional introduction of substitutions in the framework or antigen binding sites. Typically, "human antibody" is at least about 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% or 100% identical in amino acid sequence to an amino acid sequence encoded by a human germline or rearranged immunoglobulin gene. In some cases, "human antibody" may contain consensus framework sequences derived from human framework sequence analyses, for example as described in Knappik et al., J Mol Biol 296:57-86, 2000), or synthetic HCDR3 incorporated into human immunoglobulin gene libraries displayed on phage, for example as described in Shi et al., J Mol Biol 397:385-96, 2010 and Intl. Pat. Publ. No. WO2009/085462). Antibodies in which antigen binding sites are derived from a non-human species are not included in the definition of "human antibody".
[0102] Isolated humanized antibodies may be synthetic. Human antibodies, while derived from human immunoglobulin sequences, may be generated using systems such as phage display incorporating synthetic CDRs and/or synthetic frameworks, or can be subjected to in vitro mutagenesis to improve antibody properties, resulting in antibodies that do not naturally exist within the human antibody germline repertoire in vivo.
[0103] The term "recombinant antibody" as used herein, includes all antibodies that are prepared, expressed, created or isolated by recombinant means, such as antibodies isolated from an animal (e.g., a mouse) that is transgenic or transchromosomal for human immunoglobulin genes or a hybridoma prepared therefrom (described further below), antibodies isolated from a host cell transformed to express the antibody, antibodies isolated from a recombinant, combinatorial antibody library, and antibodies prepared, expressed, created or isolated by any other means that involve splicing of human immunoglobulin gene sequences to other DNA sequences, or antibodies that are generated in vitro using Fab arm exchange.
[0104] The term "monoclonal antibody" as used herein refers to a preparation of antibody molecules of single molecular composition. A monoclonal antibody composition displays a single binding specificity and affinity for a particular epitope, or in a case of a bispecific monoclonal antibody, a dual binding specificity to two distinct epitopes.
[0105] The term "substantially identical" as used herein means that the two antibody variable region amino acid sequences being compared are identical or have "insubstantial differences". Insubstantial differences are substitutions of 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, or 15 amino acids in an antibody variable region sequence that do not adversely affect antibody properties Amino acid sequences substantially identical to the variable region sequences disclosed herein are within the scope of the invention. In some embodiments, the sequence identity can be about 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% or higher. Percent identity can be determined for example by pairwise alignment using the default settings of the AlignX module of Vector NTI v.9.0.0 (Invitrogen, Carlsbad, Calif.). The protein sequences of the present invention can be used as a query sequence to perform a search against public or patent databases to, for example, identify related sequences. Exemplary programs used to perform such searches are the XBLAST or BLASTP programs (http_//www_ncbi_nlm/nih_gov), or the GenomeQuest.TM. (GenomeQuest, Westborough, Mass.) suite using the default settings.
[0106] The term "epitope" as used herein means a portion of an antigen to which an antibody specifically binds. Epitopes usually consist of chemically active (such as polar, non-polar or hydrophobic) surface groupings of moieties such as amino acids or polysaccharide side chains and can have specific three-dimensional structural characteristics, as well as specific charge characteristics. An epitope can be composed of contiguous and/or discontiguous amino acids that form a conformational spatial unit. For a discontiguous epitope, amino acids from differing portions of the linear sequence of the antigen come in close proximity in 3-dimensional space through the folding of the protein molecule.
[0107] The term "in combination with" as used herein means that two or more therapeutics can be administered to a subject together in a mixture, concurrently as single agents or sequentially as single agents in any order.
[0108] The numbering of amino acid residues in the antibody constant region throughout the specification is performed according to the EU index as described in Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991), unless otherwise explicitly stated.
Compositions of Matter
[0109] The present invention provides bispecific agents that specifically bind EGFR and c-Met. The present invention provides polypeptides and polynucleotides encoding the bispecific agents of the invention or complementary nucleic acids thereof, vectors, host cells, and methods of making and using them.
Monospecific and Bispecific EGFR and/or c-Met FN3 Domain Containing Binding Molecules
Monospecific EGFR FN3 Domain Containing Binding Molecules
[0110] The present invention provides fibronectin type III (FN3) domains that bind specifically to epidermal growth factor receptor (EGFR) and block binding of epidermal growth factor (EGF) to EGFR, and thus can be widely used in therapeutic and diagnostic applications. The present invention provides polynucleotides encoding the FN3 domains of the invention or complementary nucleic acids thereof, vectors, host cells, and methods of making and using them.
[0111] The FN3 domains of the invention bind EGFR with high affinity and inhibit EGFR signaling, and may provide a benefit in terms of specificity and reduced off-target toxicity when compared to small molecule EGFR inhibitors, and improved tissue penetration when compared to conventional antibody therapeutics.
[0112] One embodiment of the invention is an isolated fibronectin type III (FN3) domain that specifically binds epidermal growth factor receptor (EGFR) and blocks binding of epidermal growth factor (EGF) to EGFR.
[0113] The FN3 domains of the invention may block EGF binding to the EGFR with an IC.sub.50 value of less than about 1.times.10.sup.-7 M, less than about 1.times.10.sup.-8 M, less than about 1.times.10.sup.-9 M, less than about 1.times.10.sup.-10 M, less than about 1.times.10.sup.-11 M, or less than about 1.times.10.sup.-12 M in a competition assay employing A431 cells and detecting amount of fluorescence from bound biotinylated EGF using streptavidin-phycoerythrin conjugate at 600 nM on A431 cells incubated with or without the FN3 domains of the invention. Exemplary FN3 domains may block EGF binding to the EGFR with an IC.sub.50 value between about 1.times.10.sup.-9 M to about 1.times.10.sup.-7 M, such as EGFR binding FN3 domains having the amino acid sequence of SEQ ID NOs: 18-29, 107-110, or 122-137. The FN3 domains of the invention may block EGF binding to the EGFR by at least 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% or 100% when compared to binding of EGF to the EGFR in the absence of the FN3 domains of the invention using the same assay conditions.
[0114] The FN3 domain of the invention may inhibit EGFR signaling by at least 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% or 100% when compared to the level of signaling in the absence of the FN3 domains of the invention using the same assay conditions.
[0115] Binding of a ligand such as EGF to EGFR stimulates receptor dimerization, autophosphorylation, activation of the receptor's internal, cytoplasmic tyrosine kinase domain, and initiation of multiple signal transduction and transactivation pathways involved in regulation of DNA synthesis (gene activation) and cell cycle progression or division. Inhibition of EGFR signaling may result in inhibition in one or more EGFR downstream signaling pathways and therefore neutralizing EGFR may have various effects, including inhibition of cell proliferation and differentiation, angiogenesis, cell motility and metastasis.
[0116] EGFR signaling may be measured using various well know methods, for example measuring the autophosphorylation of the receptor at any of the tyrosines Y1068, Y1148, and Y1173 (Downward et al., Nature 311:483-5, 1984) and/or phosphorylation of natural or synthetic substrates. Phosphorylation can be detected using well known methods such as an ELISA assay or a western plot using a phosphotyrosine specific antibody. Exemplary assays can be found in Panek et al., J Pharmacol Exp Thera 283:1433-44, 1997 and Batley et al., Life Sci 62:143-50, 1998, and assays described herein.
[0117] In one embodiment, the FN3 domain of the invention inhibits EGF-induced EGFR phosphorylation at EGFR residue position Tyrosine 1173 with an IC.sub.50 value of less than about 2.5.times.10.sup.-6 M, for example less than about 1.times.10.sup.-6 M, less than about 1.times.10.sup.-7 M, less than about 1.times.10.sup.-8 M, less than about 1.times.10.sup.-9 M, less than about 1.times.10.sup.-10 M, less than about 1.times.10.sup.-11 M, or less than about 1.times.10.sup.-12 M when measured in A431 cells using 50 ng/mL human EGF.
[0118] In one embodiment, the FN3 domain of the invention inhibits EGF-induced EGFR phosphorylation at EGFR residue position Tyrosine 1173 with an IC.sub.50 value between about 1.8.times.10.sup.-8 M to about 2.5.times.10.sup.-6 M when measured in A431 cells using 50 ng/mL human EGF. Such exemplary FN3 domains are those having the amino acid sequence of SEQ ID NOs: 18-29, 107-110, or 122-137.
[0119] In one embodiment, the FN3 domain of the invention binds human EGFR with a dissociation constant (K.sub.D) of less than about 1.times.10.sup.-8 M, for example less than about 1.times.10.sup.-9 M, less than about 1.times.10.sup.-10 M, less than about 1.times.10.sup.-11 M, less than about 1.times.10.sup.-12 M, or less than about 1.times.10.sup.-13 M as determined by surface plasmon resonance or the Kinexa method, as practiced by those of skill in the art. In some embodiments, the FN3 domain of the invention binds human EGFR with a K.sub.D of between about 2.times.10.sup.-1.degree. to about 1.times.10.sup.-8 M. The affinity of a FN3 domain for EGFR can be determined experimentally using any suitable method. (See, for example, Berzofsky, et al., "Antibody-Antigen Interactions," In Fundamental Immunology, Paul, W. E., Ed., Raven Press: New York, N.Y. (1984); Kuby, Janis Immunology, W. H. Freeman and Company: New York, N.Y. (1992); and methods described herein). The measured affinity of a particular FN3 domain-antigen interaction can vary if measured under different conditions (e.g., osmolarity, pH). Thus, measurements of affinity and other antigen-binding parameters (e.g., K.sub.D, K.sub.on, K.sub.off) are preferably made with standardized solutions of protein scaffold and antigen, and a standardized buffer, such as the buffer described herein.
[0120] Exemplary FN3 domains of the invention that bind EGFR include FN3 domains of SEQ ID NOs: 18-29, 107-110, or 122-137.
[0121] In one embodiment, the FN3 domain that specifically binds EGFR comprises an amino acid sequence at least 87% identical to the amino acid sequence of SEQ ID NO: 27.
[0122] In one embodiment, the FN3 domain that specifically binds EGFR comprises
[0123] an FG loop comprising the sequence HNVYKDTNX.sub.9RGL (SEQ ID NO: 179) or the sequence LGSYVFEHDVML (SEQ ID NO: 180), wherein X.sub.9 is M or I; and
[0124] a BC loop comprising the sequence X.sub.1X.sub.2X.sub.3X.sub.4X.sub.5X.sub.6X.sub.7X.sub.8 (SEQ ID NO: 181),
[0125] wherein [0126] X.sub.1 is A, T, G or D; [0127] X.sub.2 is A, D, Y or W; [0128] X.sub.3 is P, D or N; [0129] X.sub.4 is L or absent; [0130] X.sub.5 is D, H, R, G, Y or W; [0131] X.sub.6 is G, D or A; [0132] X.sub.7 is A, F, G, H or D; and [0133] X.sub.8 is Y, F or L.
[0134] The FN3 domains of the invention that specifically bind EGFR and inhibit autophosphorylation of EGFR may comprise as a structural feature an FG loop comprising the sequence HNVYKDTNX.sub.9RGL (SEQ ID NO: 179) or the sequence LGSYVFEHDVML (SEQ ID NO: 180), wherein X.sub.9 is M or I. Such FN3 domains may further comprise a BC loop of 8 or 9 amino acids in length and defined by the sequence X.sub.1X.sub.2X.sub.3X.sub.4X.sub.5X.sub.6X.sub.7X.sub.8 (SEQ ID NO: 181), and inhibit EGFR autophosphorylation with an IC.sub.50 value of less than about 2.5.times.10.sup.-6 M, or with an IC.sub.50 value of between about 1.8.times.10.sup.-8 M to about 2.5.times.10.sup.-6 M when measured in A431 cells using 50 ng/mL human EGF.
[0135] The FN3 domains of the invention that specifically bind EGFR and inhibit autophosphorylation of EGFR further comprise the sequence of
TABLE-US-00001 (SEQ ID NO: 182) LPAPKNLVVSEVTEDSLRLSWX.sub.1X.sub.2X.sub.3X.sub.4X.sub.5X.sub.6X.sub.7X.su- b.8DSFLIQYQESEKV GEAINLTVPGSERSYDLTGLKPGTEYTVSIYGVHNVYKDTNX.sub.9RGLPLSA EFTT, or the sequence (SEQ ID NO: 183) LPAPKNLVVSEVTEDSLRLSWX.sub.1X.sub.2X.sub.3X.sub.4X.sub.5X.sub.6X.sub.7X.su- b.8DSFLIQYQESEKV GEAINLTVPGSERSYDLTGLKPGTEYTVSIYGVLGSYVFEHDVMLPLSAE FTT,
wherein [0136] X.sub.1 is A, T, G or D; [0137] X.sub.2 is A, D, Y or W; [0138] X.sub.3 is P, D or N; [0139] X.sub.4 is L or absent; [0140] X.sub.5 is D, H, R, G, Y or W; [0141] X.sub.6 is G, D or A; [0142] X.sub.7 is A, F, G, H or D; [0143] X.sub.8 is Y, F or L; and [0144] X.sub.9 is M or I
[0145] The EGFR binding FN3 domains can be generated and tested for their ability to inhibit EGFR autophosphorylation using well known methods and methods described herein.
[0146] Another embodiment of the invention is an isolated FN3 domain that specifically binds EGFR, wherein the FN3 domain comprises the sequence shown in SEQ ID NOs: 18-29, 107-110, or 122-137.
[0147] In some embodiments, the EGFR binding FN3 domains comprise an initiator methionine (Met) linked to the N-terminus or a cysteine (Cys) linked to a C-terminus of a particular FN3 domain, for example to facilitate expression and/or conjugation of half-life extending molecules.
[0148] Another embodiment of the invention is an isolated fibronectin type III (FN3) domain that specifically binds EGFR and blocks binding of EGF to the EGFR, wherein the FN3 domain is isolated from a library designed based on Tencon sequence of SEQ ID NO: 1.
Monospecific c-Met FN3 Domain Containing Binding Molecules
[0149] The present invention provides fibronectin type III (FN3) domains that bind specifically to hepatocyte growth factor receptor (c-Met) and block binding of hepatocyte growth factor (HGF) to c-Met, and thus can be widely used in therapeutic and diagnostic applications. The present invention provides polynucleotides encoding the FN3 domains of the invention or complementary nucleic acids thereof, vectors, host cells, and methods of making and using them.
[0150] The FN3 domains of the invention bind c-Met with high affinity and inhibit c-Met signaling, and may provide a benefit in terms of specificity and reduced off-target toxicity when compared to small molecule c-Met inhibitors, and improved tissue penetration when compared to conventional antibody therapeutics. The FN3 domains of the invention are monovalent, therefore preventing unwanted receptor clustering and activation that may occur with other bivalent molecules.
[0151] One embodiment of the invention is an isolated fibronectin type III (FN3) domain that specifically binds hepatocyte growth factor receptor (c-Met) and blocks binding of hepatocyte growth factor (HGF) to c-Met.
[0152] The FN3 domains of the invention may block HGF binding to c-Met with an IC.sub.50 value of less than about 1.times.10.sup.-7 M, less than about 1.times.10.sup.-8 M, less than about 1.times.10.sup.-9 M, less than about 1.times.10.sup.-10 M, less than about 1.times.10.sup.-11 M, or less than about 1.times.10.sup.-12 M in an assay detecting inhibition of binding of biotinylated HGF to c-Met-Fc fusion protein in the presence of the FN3 domains of the invention. Exemplary FN3 domains may block HGF binding to the c-Met with an IC.sub.50 value between about 2.times.10.sup.-10 M to about 6.times.10.sup.-8 M. The FN3 domains of the invention may block HGF binding to c-Met by at least 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% or 100% when compared to binding of HGF to c-Met in the absence of the FN3 domains of the invention using the same assay conditions.
[0153] The FN3 domain of the invention may inhibit c-Met signaling by at least 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% or 100% when compared to the level of signaling in the absence of FN3 domains of the invention using the same assay conditions.
[0154] Binding of HGF to c-Met stimulates receptor dimerization, autophosphorylation, activation of the receptor's internal, cytoplasmic tyrosine kinase domain, and initiation of multiple signal transduction and transactivation pathways involved in regulation of DNA synthesis (gene activation) and cell cycle progression or division. Inhibition of c-Met signaling may result in inhibition of one or more c-Met downstream signaling pathways and therefore neutralizing c-Met may have various effects, including inhibition of cell proliferation and differentiation, angiogenesis, cell motility and metastasis.
[0155] c-Met signaling may be measured using various well know methods, for example measuring the autophosphorylation of the receptor on at least one tyrosine residues Y1230, Y1234, Y1235 or Y1349 and/or phosphorylation of natural or synthetic substrates. Phosphorylation may be detected, for example, using an antibody specific for phosphotyrosine in an ELISA assay or on a western blot. Assays for tyrosine kinase activity are described for example in: Panek et al., J Pharmacol Exp Thera 283:1433-44, 1997 and Batley et al., Life Sci 62:143-50, 1998, and assays described herein.
[0156] In one embodiment, the FN3 domain of the invention inhibits HGF-induced c-Met phosphorylation at c-Met residue position 1349 with an IC.sub.50 value of less than about 1.times.10.sup.-6 M, less than about 1.times.10.sup.-7 M, less than about 1.times.10.sup.-8 M, less than about 1.times.10.sup.-9 M, less than about 1.times.10.sup.-10 M, less than about 1.times.10.sup.-11 M, or less than about 1.times.10.sup.-12 M when measured in NCI-H441 cells using 100 ng/mL recombinant human HGF.
[0157] In one embodiment, the FN3 domain of the invention inhibits HGF-induced c-Met phosphorylation at c-Met tyrosine Y1349 with an IC.sub.50 value between about 4.times.10.sup.-9 M to about 1.times.10.sup.-6 M when measured in NCI-H441 cells using 100 ng/mL recombinant human HGF.
[0158] In one embodiment, the FN3 domain of the invention binds human c-Met with an dissociation constant (K.sub.D) of equal to or less than about 1.times.10.sup.-7 M, 1.times.10.sup.-8 M, 1.times.10.sup.-9 M, 1.times.10.sup.-10 M, 1.times.10.sup.-11M, 1.times.10.sup.-12 M, 1.times.10.sup.-13 M, 1.times.10.sup.-14 M, or 1.times.10.sup.-15 M as determined by surface plasmon resonance or the Kinexa method, as practiced by those of skill in the art. In some embodiments, the FN3 domain of the invention binds human c-Met with a K.sub.D of between about 3.times.10.sup.-10 M to about 5.times.10.sup.-8 M. The affinity of a FN3 domain for c-Met may be determined experimentally using any suitable method. (See, for example, Berzofsky, et al., "Antibody-Antigen Interactions," In Fundamental Immunology, Paul, W. E., Ed., Raven Press: New York, N.Y. (1984); Kuby, Janis Immunology, W. H. Freeman and Company: New York, N.Y. (1992); and methods described herein). The measured affinity of a particular FN3 domain-antigen interaction can vary if measured under different conditions (e.g., osmolarity, pH). Thus, measurements of affinity and other antigen-binding parameters (e.g., K.sub.D, K.sub.on, K.sub.off) are preferably made with standardized solutions of protein scaffold and antigen, and a standardized buffer, such as the buffer described herein.
[0159] Exemplary FN3 domains of the invention that bind c-Met include FN3 domains having the amino acid sequence of SEQ ID NOs: 32-49 or 111-114.
[0160] In one embodiment, the FN3 domain that specifically binds c-Met comprises an amino acid sequence at least 83% identical to the amino acid sequence of SEQ ID NO: 41.
[0161] In one embodiment, the FN3 domain that specifically binds c-Met comprises [0162] a C strand and a CD loop comprising the sequence DSFX.sub.10IRYX.sub.11E X.sub.12X.sub.13X.sub.14X.sub.15GX.sub.16 (SEQ ID NO: 184), wherein [0163] X.sub.10 is W, F or V; [0164] X.sub.11 is D, F or L; [0165] X.sub.12 is V, F or L; [0166] X.sub.13 is V, L or T; [0167] X.sub.14 is V, R, G, L, T or S; [0168] X.sub.15 is G, S, A, T or K; and [0169] X.sub.16 is E or D; and [0170] a F strand and a FG loop comprising the sequence TEYX.sub.17VX.sub.18IX.sub.19X.sub.20V KGGX.sub.21X.sub.22SX.sub.23 (SEQ ID NO: 185), wherein [0171] X.sub.17 is Y, W, I, V, G or A; [0172] X.sub.18 is N, T, Q or G; [0173] X.sub.19 is L, M, N or I; [0174] X.sub.20 is G or S; [0175] X.sub.21 is S, L, G, Y, T, R, H or K; [0176] X.sub.22 is I, V or L; and [0177] X.sub.23 is V, T, H, I, P, Y or L.
[0178] The FN3 domains of the invention that specifically bind c-Met and inhibit autophosphorylation of c-Met further comprises the sequence:
TABLE-US-00002 (SEQ ID NO: 186) LPAPKNLVVSRVTEDSARLSWTAPDAAFDSFX.sub.10IRYX.sub.11EX.sub.12X.sub.13X.sub.1- 4X.sub.15 GX.sub.16AIVLTVPGSERSYDLTGLKPGTEYX.sub.17VX.sub.18IX.sub.19X.sub.20VKGGX.s- ub.21X.sub.22S X.sub.23PLSAEFTT,
wherein [0179] X.sub.10 is W, F or V; and [0180] X.sub.11 is D, F or L; [0181] X.sub.12 is V, F or L; [0182] X.sub.13 is V, L or T; [0183] X.sub.14 is V, R, G, L, T or S; [0184] X.sub.15 is G, S, A, T or K; [0185] X.sub.16 is E or D; [0186] X.sub.17 is Y, W, I, V, G or A; [0187] X.sub.18 is N, T, Q or G; [0188] X.sub.19 is L, M, N or I; [0189] X.sub.20 is G or S; [0190] X.sub.21 is S, L, G, Y, T, R, H or K; [0191] X.sub.22 is I, V or L; and [0192] X.sub.23 is V, T, H, I, P, Y or L.
[0193] Another embodiment of the invention is an isolated FN3 domain that specifically binds c-Met, wherein the FN3 domain comprises the sequence shown in SEQ ID NOs: 32-49 or 111-114.
[0194] Another embodiment of the invention is an isolated fibronectin type III (FN3) domain that specifically binds c-Met and blocks binding of HGF to the c-Met, wherein the FN3 domain is isolated from a library designed based on Tencon sequence of SEQ ID NO: 1.
Isolation of EGFR or c-Met FN3 Domains from a Library Based on Tencon Sequence
[0195] Tencon (SEQ ID NO: 1) is a non-naturally occurring fibronectin type III (FN3) domain designed from a consensus sequence of fifteen FN3 domains from human tenascin-C (Jacobs et al., Protein Engineering, Design, and Selection, 25:107-117, 2012; U.S. Pat. Publ. No. 2010/0216708). The crystal structure of Tencon shows six surface-exposed loops that connect seven beta-strands as is characteristic to the FN3 domains, the beta-strands referred to as A, B, C, D, E, F and G, and the loops referred to as AB, BC, CD, DE, EF, and FG loops (Bork and Doolittle, Proc Natl Acad Sci USA 89:8990-8992, 1992; U.S. Pat. No. 6,673,901). These loops, or selected residues within each loop, can be randomized in order to construct libraries of fibronectin type III (FN3) domains that can be used to select novel molecules that bind EGFR or c-Met. Table 1 shows positions and sequences of each loop and beta-strand in Tencon (SEQ ID NO: 1).
[0196] Library designed based on Tencon sequence may thus have randomized FG loop, or randomized BC and FG loops, such as libraries TCL1 or TCL2 as described below. The Tencon BC loop is 7 amino acids long, thus 1, 2, 3, 4, 5, 6 or 7 amino acids may be randomized in the library diversified at the BC loop and designed based on Tencon sequence. The Tencon FG loop is 7 amino acids long, thus 1, 2, 3, 4, 5, 6 or 7 amino acids may be randomized in the library diversified at the FG loop and designed based on Tencon sequence. Further diversity at loops in the Tencon libraries may be achieved by insertion and/or deletions of residues at loops. For example, the FG and/or BC loops may be extended by 1-22 amino acids, or decreased by 1-3 amino acids. The FG loop in Tencon is 7 amino acids long, whereas the corresponding loop in antibody heavy chains ranges from 4-28 residues. To provide maximum diversity, the FG loop may be diversified in sequence as well as in length to correspond to the antibody CDR3 length range of 4-28 residues. For example, the FG loop can further be diversified in length by extending the loop by additional 1, 2, 3, 4 or 5 amino acids.
[0197] Library designed based on Tencon sequence may also have randomized alternative surfaces that form on a side of the FN3 domain and comprise two or more beta strands, and at least one loop. One such alternative surface is formed by amino acids in the C and the F beta-strands and the CD and the FG loops (a C-CD-F-FG surface). A library design based on Tencon alternative C-CD-F-FG surface and is shown in FIG. 1 and detailed generation of such libraries is described in U.S. Pat. Publ. No. US2013/0226834.
[0198] Library designed based on Tencon sequence also includes libraries designed based on Tencon variants, such as Tencon variants having substitutions at residues positions 11, 14, 17, 37, 46, 73, or 86 (residue numbering corresponding to SEQ ID NO: 1), and which variants display improved thermal stability. Exemplary Tencon variants are described in US Pat. Publ. No. 2011/0274623, and include Tencon27 (SEQ ID NO: 99) having substitutions E11R, L17A, N46V and E86I when compared to Tencon of SEQ ID NO: 1.
TABLE-US-00003 TABLE 1 FN3 Tencon domain (SEQ ID NO: 1) A strand 1-12 AB loop 13-16 B strand 17-21 BC loop 22-28 C strand 29-37 CD loop 38-43 D strand 44-50 DE loop 51-54 E strand 55-59 EF loop 60-64 F strand 65-74 FG loop 75-81 G strand 82-89
[0199] Tencon and other FN3 sequence based libraries can be randomized at chosen residue positions using a random or defined set of amino acids. For example, variants in the library having random substitutions can be generated using NNK codons, which encode all 20 naturally occurring amino acids. In other diversification schemes, DVK codons can be used to encode amino acids Ala, Trp, Tyr, Lys, Thr, Asn, Lys, Ser, Arg, Asp, Glu, Gly, and Cys. Alternatively, NNS codons can be used to give rise to all 20 amino acid residues and simultaneously reducing the frequency of stop codons. Libraries of FN3 domains with biased amino acid distribution at positions to be diversified can be synthesized for example using Slonomics.RTM. technology (http:_//www_sloning_com). This technology uses a library of pre-made double stranded triplets that act as universal building blocks sufficient for thousands of gene synthesis processes. The triplet library represents all possible sequence combinations necessary to build any desired DNA molecule. The codon designations are according to the well known IUB code.
[0200] The FN3 domains specifically binding EGFR or c-Met of the invention can be isolated by producing the FN3 library such as the Tencon library using cis display to ligate DNA fragments encoding the scaffold proteins to a DNA fragment encoding RepA to generate a pool of protein-DNA complexes formed after in vitro translation wherein each protein is stably associated with the DNA that encodes it (U.S. Pat. No. 7,842,476; Odegrip et al., Proc Natl Acad Sci USA 101, 2806-2810, 2004), and assaying the library for specific binding to EGFR and/or c-Met by any method known in the art and described in the Example. Exemplary well known methods which can be used are ELISA, sandwich immunoassays, and competitive and non-competitive assays (see, e.g., Ausubel et al., eds, 1994, Current Protocols in Molecular Biology, Vol. 1, John Wiley & Sons, Inc., New York). The identified FN3 domains specifically binding EGFR or c-Met are further characterized for their ability to block EGFR ligand such as EGF binding to EGFR, or HGF binding to c-Met, and for their ability to inhibit EGFR and/or c-Met signaling using methods described herein.
[0201] The FN3 domains specifically binding to EGFR or c-Met of the invention can be generated using any FN3 domain as a template to generate a library and screening the library for molecules specifically binding EGFR or c-Met using methods provided within. Exemplar FN3 domains that can be used are the 3rd FN3 domain of tenascin C (TN3) (SEQ ID NO: 75), Fibcon (SEQ ID NO: 76), and the 10.sup.th FN3 domain of fibronectin (FN10) (SEQ ID NO: 77). Standard cloning and expression techniques are used to clone the libraries into a vector or synthesize double stranded cDNA cassettes of the library, to express, or to translate the libraries in vitro. For example ribosome display (Hanes and Pluckthun, Proc Natl Acad Sci USA, 94, 4937-4942, 1997), mRNA display (Roberts and Szostak, Proc Natl Acad Sci USA, 94, 12297-12302, 1997), or other cell-free systems (U.S. Pat. No. 5,643,768) can be used. The libraries of the FN3 domain variants may be expressed as fusion proteins displayed on the surface for example of any suitable bacteriophage. Methods for displaying fusion polypeptides on the surface of a bacteriophage are well known (U.S. Pat. Publ. No. 2011/0118144; Int. Pat. Publ. No. WO2009/085462; U.S. Pat. Nos. 6,969,108; 6,172,197; 5,223,409; 6,582,915; 6,472,147).
[0202] The FN3 domains specifically binding EGFR or c-Met of the invention can be modified to improve their properties such as improve thermal stability and reversibility of thermal folding and unfolding. Several methods have been applied to increase the apparent thermal stability of proteins and enzymes, including rational design based on comparison to highly similar thermostable sequences, design of stabilizing disulfide bridges, mutations to increase alpha-helix propensity, engineering of salt bridges, alteration of the surface charge of the protein, directed evolution, and composition of consensus sequences (Lehmann and Wyss, Curr Opin Biotechnol, 12, 371-375, 2001). High thermal stability may increase the yield of the expressed protein, improve solubility or activity, decrease immunogenicity, and minimize the need of a cold chain in manufacturing. Residues that can be substituted to improve thermal stability of Tencon (SEQ ID NO: 1) are residue positions 11, 14, 17, 37, 46, 73, or 86, and are described in US Pat. Publ. No. 2011/0274623. Substitutions corresponding to these residues can be incorporated to the FN3 domains or the bispecific FN3 domain containing molecules of the invention.
[0203] Another embodiment of the invention is an isolated FN3 domain that specifically binds EGFR and blocks binding of EGF to EGFR, comprising the sequence shown in SEQ ID NOs: 18-29, 107-110, 122-137, further comprising substitutions at one or more residue positions corresponding to positions 11, 14, 17, 37, 46, 73, and 86 in Tencon (SEQ ID NO: 1).
[0204] Another embodiment of the invention is an isolated FN3 domain that specifically binds c-Met and blocks binding of HGF to c-Met, comprising the sequence shown in SEQ ID NOs: 32-49 or 111-114, further comprising substitutions at one or more residue positions corresponding to positions 11, 14, 17, 37, 46, 73, and 86 in Tencon (SEQ ID NO: 1).
[0205] Exemplary substitutions are substitutions E11N, E14P, L17A, E37P, N46V, G73Y and E86I (numbering according to SEQ ID NO: 1).
[0206] In some embodiments, the FN3 domains of the invention comprise substitutions corresponding to substitutions L17A, N46V, and E86I in Tencon (SEQ ID NO: 1).
[0207] The FN3 domains specifically binding EGFR (FIG. 1) have an extended FG loop when compared to Tencon (SEQ ID NO: 1). Therefore, the residues corresponding to residues 11, 14, 17, 37, 46, 73, and 86 in Tencon (SEQ ID NO: 1) are residues 11, 14, 17, 37, 46, 73 and 91 in EGFR FN3 domains shown in FIGS. 1A and 1B except for the FN3 domain of SEQ ID NO: 24, wherein the corresponding residues are residues 11, 14, 17, 38, 74, and 92 due to an insertion of one amino acid in the BC loop.
[0208] Another embodiment of the invention is an isolated FN3 domain that specifically binds EGFR and blocks binding of EGF to EGFR comprising the amino acid sequence shown in SEQ ID NOs: 18-29, 107-110, or 122-137, optionally having substitutions corresponding to substitutions L17A, N46V, and E86I in Tencon (SEQ ID NO: 1).
[0209] Another embodiment of the invention is an isolated FN3 domain that specifically binds c-Met and blocks binding of HGF to c-Met comprising the amino acid sequence shown in SEQ ID NOs: 32-49 or 111-114, optionally having substitutions corresponding to substitutions L17A, N46V, and E86I in Tencon (SEQ ID NO: 1).
[0210] Measurement of protein stability and protein lability can be viewed as the same or different aspects of protein integrity. Proteins are sensitive or "labile" to denaturation caused by heat, by ultraviolet or ionizing radiation, changes in the ambient osmolarity and pH if in liquid solution, mechanical shear force imposed by small pore-size filtration, ultraviolet radiation, ionizing radiation, such as by gamma irradiation, chemical or heat dehydration, or any other action or force that may cause protein structure disruption. The stability of the molecule can be determined using standard methods. For example, the stability of a molecule can be determined by measuring the thermal melting ("TM") temperature, the temperature in .degree. Celsius (.degree. C.) at which half of the molecules become unfolded, using standard methods. Typically, the higher the TM, the more stable the molecule. In addition to heat, the chemical environment also changes the ability of the protein to maintain a particular three dimensional structure.
[0211] In one embodiment, the FN3 domains binding EGFR or c-Met of the invention exhibit increased stability by at least 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, or 95% or more compared to the same domain prior to engineering measured by the increase in the TM.
[0212] Chemical denaturation can likewise be measured by a variety of methods. Chemical denaturants include guanidinium hydrochloride, guanidinium thiocyanate, urea, acetone, organic solvents (DMF, benzene, acetonitrile), salts (ammonium sulfate, lithium bromide, lithium chloride, sodium bromide, calcium chloride, sodium chloride); reducing agents (e.g. dithiothreitol, beta-mercaptoethanol, dinitrothiobenzene, and hydrides, such as sodium borohydride), non-ionic and ionic detergents, acids (e.g. hydrochloric acid (HC1), acetic acid (CH.sub.3COOH), halogenated acetic acids), hydrophobic molecules (e.g. phosopholipids), and targeted denaturants. Quantitation of the extent of denaturation can rely on loss of a functional property, such as ability to bind a target molecule, or by physiochemical properties, such as tendency to aggregation, exposure of formerly solvent inaccessible residues, or disruption or formation of disulfide bonds.
[0213] In one embodiment, the FN3 domain of the invention binding EGFR or c-Met exhibit increased stability by at least 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, or 95% or more compared to the same scaffold prior to engineering, measured by using guanidinium hydrochloride as a chemical denaturant. Increased stability can be measured as a function of decreased tryptophan fluorescence upon treatment with increasing concentrations of guanidine hydrochloride using well known methods.
[0214] The FN3 domains of the invention may be generated as monomers, dimers, or multimers, for example, as a means to increase the valency and thus the avidity of target molecule binding, or to generate bi- or multispecific scaffolds simultaneously binding two or more different target molecules. The dimers and multimers may be generated by linking monospecific, bi- or multispecific protein scaffolds, for example, by the inclusion of an amino acid linker, for example a linker containing poly-glycine, glycine and serine, or alanine and proline. Exemplary linker include (GS).sub.2, (SEQ ID NO: 78), (GGGGS).sub.5 (SEQ ID NO: 79), (AP).sub.2 (SEQ ID NO: 80), (AP).sub.5 (SEQ ID NO: 81), (AP).sub.10 (SEQ ID NO: 82), (AP).sub.20 (SEQ ID NO: 83) and A(EAAAK).sub.5AAA (SEQ ID NO: 84), linkers. The dimers and multimers may be linked to each other in an N to C-direction. The use of naturally occurring as well as artificial peptide linkers to connect polypeptides into novel linked fusion polypeptides is well known in the literature (Hallewell et al., J Biol Chem 264, 5260-5268, 1989; Alfthan et al., Protein Eng. 8, 725-731, 1995; Robinson & Sauer, Biochemistry 35, 109-116, 1996; U.S. Pat. No. 5,856,456).
Bispecific Agents Specifically Binding EGFR and c-Met
[0215] The bispecific agents that specifically bind EGFR and c-Met of the invention may provide a benefit in terms of specificity and reduced off-target toxicity when compared to small molecule EGFR and/or c-Met inhibitors. The present invention is based at least in part on the surprising finding that the bispecific agents specifically binding EGFR and c-Met provide a significantly improved synergistic inhibitory effect when compared to a mixture of EGFR-binding and c-Met-binding monospecific agents. The molecules may be tailored to specific affinity towards both EGFR and c-Met to maximize tumor penetration and retention. The bispecific agents that specifically bind EGFR and c-Met provide more efficient inhibition of EGFR and/or c-Met signaling pathways and inhibit tumor growth more efficiently than cetuximab (Erbitux.RTM.).
[0216] The bispecific agents specifically binding EGFR and c-Met may be formed by any polypeptide or a multimeric polypeptide that comprises an EGFR binding domain and a c-Met binding domain. The EGFR and the c-Met binding domains may be an antigen binding sites of an antibody, a VH/VL pair of an antibody, or another type of binding molecule such as a domain based on fibronectin type III (FN3) domain, a fibronectin type IX (FN9) domain, or any combination thereof.
[0217] The EGFR and c-Met binding polypeptides may be derived from existing monospecific EGFR and c-Met binding polypeptides or may be isolated de novo.
Bispecific EGFR/c/Met FN3 Domain Containing Molecules
[0218] One embodiment of the invention is an isolated bispecific FN3 domain containing molecule comprising a first fibronectin type III (FN3) domain and a second FN3 domain, wherein the first FN3 domain specifically binds epidermal growth factor receptor (EGFR) and blocks binding of epidermal growth factor (EGF) to EGFR, and the second FN3 domain specifically binds hepatocyte growth factor receptor (c-Met) and blocks binding of hepatocyte growth factor (HGF) to c-Met.
[0219] The bispecific EGFR/c-Met FN3 domain containing molecules of the invention may be generated by covalently linking any EGFR-binding FN3 domain and any c-Met-binding FN3 domain of the invention directly or via a linker. Therefore, the first FN3 domain of the bispecific molecule may have characteristics as described above for the EGFR-binding FN3 domains, and the second FN3 domain of the bispecific molecule may have characteristics as described above for the c-Met-binding FN3 domains.
[0220] In one embodiment, the first FN3 domain of the bispecific EGFR/c-Met FN3 domain containing molecule inhibits EGF-induced EGFR phosphorylation at EGFR residue Tyrosine 1173 with an IC.sub.50 value of less than about 2.5.times.10.sup.-6 M when measured in A431 cells using 50 ng/mL human EGF, and the second FN3 domain of the bispecific EGFR/c-Met FN3 domain containing molecule inhibits HGF-induced c-Met phosphorylation at c-Met residue Tyrosine 1349 with an IC.sub.50 value of less than about 1.5.times.10.sup.-6 M when measured in NCI-H441 cells using 100 ng/mL human HGF.
[0221] In another embodiment, the first FN3 domain of the bispecific EGFR/c-Met FN3 domain containing molecule inhibits EGF-induced EGFR phosphorylation at EGFR residue Tyrosine 1173 with an IC.sub.50 value of between about 1.8.times.10.sup.-8 M to about 2.5.times.10.sup.-6 M when measured in A431 cells using 50 ng/mL human EGF, and the second FN3 domain of the bispecific EGFR/c-Met FN3 domain containing molecule inhibits HGF-induced c-Met phosphorylation at c-Met residue Tyrosine 1349 with an IC.sub.50 value between about 4.times.10.sup.-9 M to about 1.5.times.10.sup.-6 M when measured in NCI-H441 cells using 100 ng/mL human HGF.
[0222] In another embodiment, the first FN3 domain of the bispecific EGFR/c-Met FN3 domain containing molecule binds human EGFR with a dissociation constant (K.sub.D) of less than about 1.times.10.sup.-8 M, and the second FN3 domain of the bispecific EGFR/c-Met FN3 domain containing molecule binds human c-Met with a K.sub.D of less than about 5.times.10.sup.-8 M.
[0223] In the bispecific molecule binding both EGFR and c-Met, the first FN3 domain binds human EGFR with a K.sub.D of between about 2.times.10.sup.-10 to about 1.times.10.sup.-8 M, and the second FN3 domain binds human c-Met with a K.sub.D of between about 3.times.10.sup.-10 to about 5.times.10.sup.-8 M.
[0224] The affinity of the bispecific EGFR/c-Met molecule for EGFR and c-Met can be determined as described above for the monospecific molecules.
[0225] The first FN3 domain in the bispecific EGFR/c-Met molecule of the invention may block EGF binding to EGFR with an IC.sub.50 value of between about 1.times.10.sup.-9 M to about 1.5.times.10.sup.-7 M in an assay employing A431 cells and detecting the amount of fluorescence from bound biotinylated EGF using streptavidin-phycoerythrin conjugate at 600 nM on A431 cells incubated with or without the first FN3 domain. The first FN3 domain in the bispecific EGFR/c-Met molecule of the invention may block EGF binding to the EGFR by at least 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% or 100% when compared to binding of EGF to EGFR in the absence of the first FN3 domains using the same assay conditions.
[0226] The second FN3 domain in the bispecific EGFR/c-Met molecule of the invention may block HGF binding to c-Met with an IC.sub.50 value of between about 2.times.10.sup.-10 M to about 6.times.10.sup.-8 M in an assay detecting inhibition of binding of biotinylated HGF to c-Met-Fc fusion protein in the presence of the second FN3 domain. The second FN3 domain in the bispecific EGFR/c-Met molecule may block HGF binding to c-Met by at least 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% or 100% when compared to binding of HGF to c-Met in the absence of the second FN3 domain using the same assay conditions.
[0227] The bispecific EGFR/c-Met molecule of the invention may inhibit EGFR and/or c-Met signaling by at least 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% or 100% when compared to the level of signaling in the absence of the bispecific EGFR/c-Met molecule of the invention using the same assay conditions.
[0228] EGFR and c-Met signaling may be measured using various well know methods as described above for the monospecific molecules.
[0229] The bispecific EGFR/c-Met molecules of the invention comprising the first FN3 domain specifically binding EGFR and the second FN3 domain specifically binding c-Met provide a significantly increased synergistic inhibition of EGFR and c-Met signaling and tumor cell proliferation when compared to the synergistic inhibition observed by a mixture of the first and the second FN3 domain. Synergistic inhibition can be assessed for example by measuring inhibition of ERK phosphorylation by the bispecific EGFR/c-Met FN3 domain containing molecules and by a mixture of two monospecific molecules, one binding EGFR and the other c-Met. The bispecific EGFR/c-Met molecules of the invention may inhibit ERK phosphorylation with an at least about 100 fold smaller, for example at least 500, 1000, 5000 or 10,000 fold smaller IC.sub.50 value when compared to the IC.sub.50 value for a mixture of two monospecific FN3 domains, indicating at least 100 fold increased potency for the bispecific EGFR/c-Met FN3 domain containing molecules when compared to the mixture of two monospecific FN3 domains. Exemplary bispecific EGFR-c-Met FN3 domain containing molecules may inhibit ERK phosphorylation with and IC.sub.50 value of about 5.times.10.sup.-9 M or less. ERK phosphorylation may be measured using standard methods and methods described herein.
[0230] The bispecific EGFR/c-Met FN3 domain containing molecule of the invention may inhibit NCI-H292 cell proliferation with an IC.sub.50 value that is at least 30-fold less when compared to the IC.sub.50 value of inhibition of NCI-H292 cell growth with a mixture of the first FN3 domain and the second FN3, wherein the cell proliferation is induced with medium containing 10% FBS supplemented with 7.5 ng/mL HGF. The bispecific molecule of the invention may inhibit tumor cell proliferation with an IC.sub.50 value that is about 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 300, 400, 500, 600, 700, 800, or about 1000 fold less when compared to the IC.sub.50 value of inhibition of tumor cell proliferation with a mixture of the first FN3 domain and the second FN3 domain. Inhibition of tumor cell proliferation may be measured using standard methods and methods described herein.
[0231] Another embodiment of the invention is a bispecific FN3 domain containing molecule comprising a first fibronectin type III (FN3) domain and a second FN3 domain, wherein the first FN3 domain specifically binds epidermal growth factor receptor (EGFR) and blocks binding of epidermal growth factor (EGF) to EGFR, and the second FN3 domain specifically binds hepatocyte growth factor receptor (c-Met), and blocks binding of hepatocyte growth factor (HGF) to c-Met, wherein
[0232] the first FN3 domain comprises [0233] an FG loop comprising the sequence HNVYKDTNX.sub.9RGL (SEQ ID NO: 179) or the sequence LGSYVFEHDVML (SEQ ID NO: 180), wherein X.sub.9 is M or I; and [0234] a BC loop comprising the sequence X.sub.1X.sub.2X.sub.3X.sub.4X.sub.5X.sub.6X.sub.7X.sub.8 (SEQ ID NO: 181), wherein [0235] X.sub.1 is A, T, G or D; [0236] X.sub.2 is A, D, Y or W; [0237] X.sub.3 is P, D or N; [0238] X.sub.4 is L or absent; [0239] X.sub.5 is D, H, R, G, Y or W; [0240] X.sub.6 is G, D or A; [0241] X.sub.7 is A, F, G, H or D; and [0242] X.sub.8 is Y, F or L; and
[0243] the second FN3 domain comprises [0244] a C strand and a CD loop comprising the sequence DSFX.sub.10IRYX.sub.11E X.sub.12X.sub.13X.sub.14X.sub.15GX.sub.16 (SEQ ID NO: 184), wherein [0245] X.sub.10 is W, F or V; [0246] X.sub.11 is D, F or L; [0247] X.sub.12 is V, F or L; [0248] X.sub.13 is V, L or T; [0249] X.sub.14 is V, R, G, L, T or S; [0250] X.sub.15 is G, S, A, T or K; and [0251] X.sub.16 is E or D; and [0252] a F strand and a FG loop comprising the sequence TEYX.sub.17VX.sub.18IX.sub.19X.sub.20V KGGX.sub.21X.sub.22SX.sub.23 (SEQ ID NO: 185), wherein [0253] X.sub.17 is Y, W, I, V, G or A; [0254] X.sub.18 is N, T, Q or G; [0255] X.sub.19 is L, M, N or I; [0256] X.sub.20 is G or S; [0257] X.sub.21 is S, L, G, Y, T, R, H or K; [0258] X.sub.22 is I, V or L; and [0259] X.sub.23 is V, T, H, I, P, Y or L.
[0260] In another embodiment, the bispecific molecule comprises the first FN3 domain that binds EGFR comprising the sequence:
TABLE-US-00004 (SEQ ID NO: 182) LPAPKNLVVSEVTEDSLRLSWX.sub.1X.sub.2X.sub.3X.sub.4X.sub.5X.sub.6X.sub.7X.su- b.8DSFLIQYQESEKV GEAINLTVPGSERSYDLTGLKPGTEYTVSIYGVHNVYKDTNX.sub.9RGLPLSA EFTT, or the sequence (SEQ ID NO: 183) LPAPKNLVVSEVTEDSLRLSWX.sub.1X.sub.2X.sub.3X.sub.4X.sub.5X.sub.6X.sub.7X.su- b.8DSFLIQYQESEKV GEAINLTVPGSERSYDLTGLKPGTEYTVSIYGVLGSYVFEHDVMLPLSAE FTT,
wherein in the SEQ ID NOs: 182 and 183; [0261] X.sub.1 is A, T, G or D; [0262] X.sub.2 is A, D, Y or W; [0263] X.sub.3 is P, D or N; [0264] X.sub.4 is L or absent; [0265] X.sub.5 is D, H, R, G, Y or W; [0266] X.sub.6 is G, D or A; [0267] X.sub.7 is A, F, G, H or D; [0268] X.sub.8 is Y, F or L; and [0269] X.sub.9 is M or I.
[0270] In another embodiment, the bispecific molecule comprises the second FN3 domain that binds c-Met comprising the sequence
TABLE-US-00005 (SEQ ID NO: 186) LPAPKNLVVSRVTEDSARLSWTAPDAAFDSFX.sub.10IRYX.sub.11EX.sub.12X.sub.13X.sub.1- 4X.sub.15 GX.sub.16AIVLTVPGSERSYDLTGLKPGTEYX.sub.17VX.sub.18IX.sub.19X.sub.20VKGGX.s- ub.21X.sub.22S X.sub.23PLSAEFTT,
wherein [0271] X.sub.10 is W, F or V; and [0272] X.sub.11 is D, F or L; [0273] X.sub.12 is V, F or L; [0274] X.sub.13 is V, L or T; [0275] X.sub.14 is V, R, G, L, T or S; [0276] X.sub.15 is G, S, A, T or K; [0277] X.sub.16 is E or D; [0278] X.sub.17 is Y, W, I, V, G or A; [0279] X.sub.18 is N, T, Q or G; [0280] X.sub.19 is L, M, N or I; [0281] X.sub.20 is G or S; [0282] X.sub.21 is S, L, G, Y, T, R, H or K; [0283] X.sub.22 is I, V or L; and [0284] X.sub.23 is V, T, H, I, P, Y or L.
[0285] Exemplary bispecific EGFR/c-Met FN3 domain containing molecules comprise the amino acid sequence shown in SEQ ID NOs: 50-72, 106, 118-121, or 138-167.
[0286] The bispecific EGFR/c-Met molecules of the invention comprise certain structural characteristics associated with their functional characteristics, such as inhibition of EGFR autophosphorylation, such as the FG loop of the first FN3 domain that binds EGFR comprising the sequence HNVYKDTNX.sub.9RGL (SEQ ID NO: 179) or the sequence LGSYVFEHDVML (SEQ ID NO: 180), wherein X.sub.9 is M or I.
[0287] In one embodiment, the bispecific EGFR/c-Met FN3 domain containing molecules of the invention
[0288] inhibit EGF-induced EGFR phosphorylation at EGFR residues Tyrosine 1173 with an IC.sub.50 value of less than about 8.times.10.sup.-7 M when measured in H292 cells using 50 ng/mL human EGF;
inhibit HGF-induced c-Met phosphorylation at c-Met residue Tyrosine 1349 with an IC.sub.50 value of less than about 8.4.times.10.sup.-7 M when measured in NCI-H441 cells using 100 ng/mL human HGF;
[0289] inhibit HGF-induced NCI-H292 cell proliferation with an IC.sub.50 value of less than about 9.5.times.10.sup.-6M wherein the cell proliferation is induced with 10% FBS containing 7.5 ng HGF;
[0290] bind EGFR with a K.sub.D of less than about 2.0.times.10.sup.-8 M; or
[0291] bind c-Met with a K.sub.D of less than about 2.0.times.10.sup.-8 M.
[0292] In another embodiment, the bispecific EGFR/c-Met FN3 domain containing molecules of the invention
inhibit EGF-induced EGFR phosphorylation at EGFR residues Tyrosine 1173 with and IC.sub.50 of between about 4.2.times.10.sup.-9 M and 8.times.10.sup.-7 M when measured in H292 cells using 50 ng/mL human EGF;
[0293] inhibit HGF-induced c-Met phosphorylation at c-Met residues Tyrosine 1349 with an IC.sub.50 value of between about 2.4.times.10.sup.-8 M to about 8.4.times.10.sup.-7 M when measured in NCI-H441 cells using 100 ng/mL human HGF;
[0294] inhibit HGF-induced NCI-H292 cell proliferation with an IC.sub.50 value between about 2.3.times.10.sup.-8 M to about 9.5.times.10.sup.-6M wherein the cell proliferation is induced with 10% FBS containing 7.5 ng HGF;
[0295] bind EGFR with a K.sub.D of between about 2.times.10.sup.-10 M to about 2.0.times.10.sup.-8 M; or
[0296] bind c-Met with a K.sub.D of between about 1.times.10.sup.-9 M to about 2.0.times.10.sup.-8 M.
[0297] In one embodiment, the bispecific EGFR/c-Met molecules comprise the EGFR-binding FN3 domain comprising the sequence
TABLE-US-00006 (SEQ ID NO: 182) LPAPKNLVVSEVTEDSLRLSWX.sub.1X.sub.2X.sub.3X.sub.4X.sub.5X.sub.6X.sub.7X.su- b.8DSFLIQYQESEKVGE AINLTVPGSERSYDLTGLKPGTEYTVSIYGVHNVYKDTNX.sub.9RGLPLSAEF TT,
wherein [0298] X.sub.1 is D; [0299] X.sub.2 is D; [0300] X.sub.3 is P; [0301] X.sub.4 is absent; [0302] X.sub.5 is H or W; [0303] X.sub.6 is A; [0304] X.sub.7 is F [0305] X.sub.8 is Y; and [0306] X.sub.9 is M or I; and
[0307] the c-Met-binding FN3 domain comprising the sequence
TABLE-US-00007 (SEQ ID NO: 186) LPAPKNLVVSRVTEDSARLSWTAPDAAFDSFX.sub.10IRYX.sub.11EX.sub.12X.sub.13X.sub.1- 4X.sub.15 GX.sub.16AIVLTVPGSERSYDLTGLKPGTEYX.sub.17VX.sub.18IX.sub.19X.sub.20VKGGX.s- ub.21X.sub.22S X.sub.23PLSAEFTT,
wherein [0308] X.sub.10 is W; [0309] X.sub.11 is F; [0310] X.sub.12 is F; [0311] X.sub.13 is V or L; [0312] X.sub.14 is G or S; [0313] X.sub.15 is S or K; [0314] X.sub.16 is E or D; [0315] X.sub.17 is V; [0316] X.sub.18 is N; [0317] X.sub.19 is L or M; [0318] X.sub.20 is G or S; [0319] X.sub.21 is S or K; [0320] X.sub.22 is I; and [0321] X.sub.23 is P.
[0322] Exemplary bispecific EGFR/c-Met molecules are those having the sequence shown in SEQ ID NOs: 57, 61, 62, 63, 64, 65, 66, 67 and 68.
[0323] The bispecific molecules of the invention may further comprise substitutions at one or more residue positions in the first FN3 domain and/or the second FN3 domain corresponding to positions 11, 14, 17, 37, 46, 73 and 86 in Tencon (SEQ ID NO: 1) as described above, and a substitution at position 29. Exemplary substitutions are substitutions E11N, E14P, L17A, E37P, N46V, G73Y, E86I and D29E (numbering according to SEQ ID NO: 1). Skilled in the art will appreciate that other amino acids can be used for substitutions, such as amino acids within a family of amino acids that are related in their side chains as described infra. The generated variants can be tested for their stability and binding to EGFR and/or c-Met using methods herein.
[0324] In one embodiment, the bispecific EGFR/c-Met FN3 domain containing molecule comprises the first FN3 domain that binds specifically EGFR and the second FN3 domain that binds specifically c-Met, wherein the first FN3 domain comprises the sequence:
TABLE-US-00008 (SEQ ID NO: 187) LPAPKNLVVSX.sub.24VTX.sub.25DSX.sub.26RLSWDDPX.sub.27AFYX.sub.28SFLIQYQX.s- ub.29SE KVGEAIX.sub.30LTVPGSERSYDLTGLKPGTEYTVSIYX.sub.31VHNVYKDTNX.sub.32R GLPLSAX.sub.33FTT,
wherein
X.sub.24 is E, N or R;
X.sub.25 is E or P;
X.sub.26 is L or A;
X.sub.27 is H or W;
X.sub.28 is E or D;
X.sub.29 is E or P;
X.sub.30 is N or V;
X.sub.31 is G or Y;
X.sub.32 is M or I; and
X.sub.33 is E or I;
[0325] and the second FN3 domain comprises the sequence:
TABLE-US-00009 (SEQ ID NO: 188) LPAPKNLVVSX.sub.34VTX.sub.35DSX.sub.36RLSWTAPDAAFDSFWIRYFX.sub.37FX.sub.38- X.sub.39 X.sub.40GX.sub.41AIX.sub.42LTVPGSERSYDLTGLKPGTEYVVNIX.sub.43X.sub.44VKGGX.- sub.45ISP PLSAX.sub.46FTT;
wherein
X.sub.34 is E, N or R;
X.sub.35 is E or P;
X.sub.36 is L or A;
X.sub.37 is E or P;
X.sub.38 is V or L;
X.sub.39 is G or S;
X.sub.40 is S or K;
X.sub.41 is E or D;
X.sub.42 is N or V;
X.sub.43 is L or M;
X.sub.44 is G or S;
X.sub.45 is S or K; and
X.sub.46 is E or I.
[0326] In other embodiments, the bispecific EGFR/c-Met FN3 domain containing molecule comprises the first FN3 domain comprising an amino acid sequence at least 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% or 99% identical to the amino acid sequence of SEQ ID NO: 27, and the second FN3 domain comprising an amino acid sequence at least 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% or 99% identical to the amino acid sequence of SEQ ID NO: 41.
[0327] The bispecific EGFR/c-Met FN3 domain containing molecules of the invention may be tailored to a specific affinity towards EGFR and c-Met to maximize tumor accumulation.
[0328] Another embodiment of the invention is an isolated bispecific FN3 domain containing molecule comprising a first fibronectin type III (FN3) domain and a second FN3 domain, wherein the first FN3 domain specifically binds epidermal growth factor receptor (EGFR) and blocks binding of epidermal growth factor (EGF) to EGFR, and the second FN3 domain specifically binds hepatocyte growth factor receptor (c-Met), and blocks binding of hepatocyte growth factor (HGF) to c-Met, wherein the first FN3 domain and the second FN3 domain is isolated from a library designed based on Tencon sequence of SEQ ID NO: 1.
[0329] The bispecific EGFR/c-Met FN3 domain containing molecule of the invention can be generated by covalently coupling the EGFR-binding FN3 domain and the c-Met binding FN3 domain of the invention using well known methods. The FN3 domains may be linked via a linker, for example a linker containing poly-glycine, glycine and serine, or alanine and proline. Exemplary linker include (GS).sub.2, (SEQ ID NO: 78), (GGGGS).sub.5 (SEQ ID NO: 79), (AP).sub.2 (SEQ ID NO: 80), (AP).sub.5 (SEQ ID NO: 81), (AP).sub.10 (SEQ ID NO: 82), (AP).sub.20 (SEQ ID NO: 83), A(EAAAK).sub.5AAA (SEQ ID NO: 84), linkers. The use of naturally occurring as well as artificial peptide linkers to connect polypeptides into novel linked fusion polypeptides is well known in the literature (Hallewell et al., J Biol Chem 264, 5260-5268, 1989; Alfthan et al., Protein Eng. 8, 725-731, 1995; Robinson & Sauer, Biochemistry 35, 109-116, 1996; U.S. Pat. No. 5,856,456). The bispecific EGFR/c-Met molecules of the invention may be linked together from a C-terminus of the first FN3 domain to the N-terminus of the second FN3 domain, or from the C-terminus of the second FN3 domain to the N-terminus of the first FN3 domain. Any EGFR-binding FN3 domain may be covalently linked to a c-Met-binding FN3 domain. Exemplary EGFR-binding FN3 domains are domains having the amino acid sequence shown in SEQ ID NOs: 18-29, 107-110, and 122-137, and exemplary c-Met binding FN3 domains are domains having the amino acid sequence shown in SEQ ID NOs: 32-49 and 111-114. The EGFR-binding FN3 domains to be coupled to a bispecific molecule may additionally comprise an initiator methionine (Met) at their N-terminus.
[0330] Variants of the bispecific EGFR/c-Met FN3 domain containing molecules are within the scope of the invention. For example, substitutions can be made in the bispecific EGFR/c-Met FN3 domain containing molecule as long as the resulting variant retains similar selectivity and potency towards EGFR and c-Met when compared to the parent molecule. Exemplary modifications are for example conservative substitutions that will result in variants with similar characteristics to those of the parent molecules. Conservative substitutions are those that take place within a family of amino acids that are related in their side chains. Genetically encoded amino acids can be divided into four families: (1) acidic (aspartate, glutamate); (2) basic (lysine, arginine, histidine); (3) nonpolar (alanine, valine, leucine, isoleucine, proline, phenylalanine, methionine, tryptophan); and (4) uncharged polar (glycine, asparagine, glutamine, cysteine, serine, threonine, tyrosine). Phenylalanine, tryptophan, and tyrosine are sometimes classified jointly as aromatic amino acids. Alternatively, the amino acid repertoire can be grouped as (1) acidic (aspartate, glutamate); (2) basic (lysine, arginine histidine), (3) aliphatic (glycine, alanine, valine, leucine, isoleucine, serine, threonine), with serine and threonine optionally be grouped separately as aliphatic-hydroxyl; (4) aromatic (phenylalanine, tyrosine, tryptophan); (5) amide (asparagine, glutamine); and (6) sulfur-containing (cysteine and methionine) (Stryer (ed.), Biochemistry, 2nd ed, WH Freeman and Co., 1981). Non-conservative substitutions can be made to the bispecific EGFR/c-Met FN3 domain containing molecule that involves substitutions of amino acid residues between different classes of amino acids to improve properties of the bispecific molecules. Whether a change in the amino acid sequence of a polypeptide or fragment thereof results in a functional homolog can be readily determined by assessing the ability of the modified polypeptide or fragment to produce a response in a fashion similar to the unmodified polypeptide or fragment using the assays described herein. Peptides, polypeptides or proteins in which more than one replacement has taken place can readily be tested in the same manner.
[0331] The bispecific EGFR/c-Met FN3 domain containing molecules of the invention may be generated as dimers or multimers, for example, as a means to increase the valency and thus the avidity of target molecule binding. The multimers may be generated by linking one or more EGFR-binding FN3 domain and one or more c-Met-binding FN3 domain to form molecules comprising at least three individual FN3 domains that are at least bispecific for either EGFR or c-Met, for example by the inclusion of an amino acid linker using well known methods.
[0332] Another embodiment of the invention is a bispecific FN3 domain containing molecule comprising a first fibronectin type III (FN3) domain and a second FN3 domain, wherein the first FN3 domain specifically binds epidermal growth factor receptor (EGFR) and blocks binding of epidermal growth factor (EGF) to EGFR, and the second FN3 domain specifically binds hepatocyte growth factor receptor (c-Met), and blocks binding of hepatocyte growth factor (HGF) to c-Met comprising the amino acid sequence shown in SEQ ID NOs: 50-72, 106 or 138-165.
Half-Life Extending Moieties
[0333] The bispecific EGFR/c-Met FN3 domain containing molecules or the monospecific EGFR or c-Met binding FN3 domains of the invention may incorporate other subunits for example via covalent interaction. In one aspect of the invention, the bispecific EGFR/c-Met FN3 domain containing molecules of the invention further comprise a half-life extending moiety. Exemplary half-life extending moieties are albumin, albumin variants, albumin-binding proteins and/or domains, transferrin and fragments and analogues thereof, and Fc regions. An exemplary albumin-binding domain is shown in SEQ ID NO: 117.
[0334] All or a portion of an antibody constant region may be attached to the molecules of the invention to impart antibody-like properties, especially those properties associated with the Fc region, such as Fc effector functions such as C1q binding, complement dependent cytotoxicity (CDC), Fc receptor binding, antibody-dependent cell-mediated cytotoxicity (ADCC), phagocytosis, down regulation of cell surface receptors (e.g., B cell receptor; BCR), and can be further modified by modifying residues in the Fc responsible for these activities (for review; see Strohl, Curr Opin Biotechnol. 20, 685-691, 2009).
[0335] Additional moieties may be incorporated into the bispecific molecules of the invention such as polyethylene glycol (PEG) molecules, such as PEG5000 or PEG20,000, fatty acids and fatty acid esters of different chain lengths, for example laurate, myristate, stearate, arachidate, behenate, oleate, arachidonate, octanedioic acid, tetradecanedioic acid, octadecanedioic acid, docosanedioic acid, and the like, polylysine, octane, carbohydrates (dextran, cellulose, oligo- or polysaccharides) for desired properties. These moieties may be direct fusions with the protein scaffold coding sequences and may be generated by standard cloning and expression techniques. Alternatively, well known chemical coupling methods may be used to attach the moieties to recombinantly produced molecules of the invention.
[0336] A pegyl moiety may for example be added to the bispecific or monospecific molecules of the invention by incorporating a cysteine residue to the C-terminus of the molecule and attaching a pegyl group to the cysteine using well known methods. Exemplary bispecific molecules with the C-terminal cysteine are those having the amino acid sequence shown in SEQ IN NO: 170-178.
[0337] Monospecific and bispecific molecules of the invention incorporating additional moieties may be compared for functionality by several well known assays. For example, altered properties of monospecific and/or bispecific molecules due to incorporation of Fc domains and/or Fc domain variants may be assayed in Fc receptor binding assays using soluble forms of the receptors, such as the Fc.gamma.RI, Fc.gamma.RII, Fc.gamma.RIII or FcRn receptors, or using well known cell-based assays measuring for example ADCC or CDC, or evaluating pharmacokinetic properties of the molecules of the invention in in vivo models.
Polynucleotides, Vectors, Host Cells
[0338] The invention provides for nucleic acids encoding the EGFR-binding or c-Met binding FN3 domains or the bispecific EGFR/c-Met FN3 domain containing molecules of the invention as isolated polynucleotides or as portions of expression vectors or as portions of linear DNA sequences, including linear DNA sequences used for in vitro transcription/translation, vectors compatible with prokaryotic, eukaryotic or filamentous phage expression, secretion and/or display of the compositions or directed mutagens thereof. Certain exemplary polynucleotides are disclosed herein, however, other polynucleotides which, given the degeneracy of the genetic code or codon preferences in a given expression system, encode the EGFR-binding or c-Met binding FN3 domains or the bispecific EGFR/c-Met FN3 domain containing molecules of the invention are also within the scope of the invention.
[0339] One embodiment of the invention is an isolated polynucleotide encoding the FN3 domain specifically binding EGFR having the amino acid sequence of SEQ ID NOs: 18-29, 107-110, or 122-137.
[0340] One embodiment of the invention is an isolated polynucleotide comprising the polynucleotide sequence of SEQ ID NOs: 97-98 or 168-169.
[0341] One embodiment of the invention is an isolated polynucleotide encoding the FN3 domain specifically binding c-Met having the amino acid sequence of the sequence shown in SEQ ID NOs: 32-49 or 111-114.
[0342] One embodiment of the invention is an isolated polynucleotide encoding the bispecific EGFR/-c-Met FN3 domain containing molecule having the amino acid sequence of SEQ ID NOs: 50-72, 106, 118-121 or 138-165.
[0343] One embodiment of the invention is an isolated polynucleotide comprising the polynucleotide sequence of SEQ ID NOs: 115-116 or 166-167.
[0344] The polynucleotides of the invention may be produced by chemical synthesis such as solid phase polynucleotide synthesis on an automated polynucleotide synthesizer and assembled into complete single or double stranded molecules. Alternatively, the polynucleotides of the invention may be produced by other techniques such as PCR followed by routine cloning. Techniques for producing or obtaining polynucleotides of a given known sequence are well known in the art.
[0345] The polynucleotides of the invention may comprise at least one non-coding sequence, such as a promoter or enhancer sequence, intron, polyadenylation signal, a cis sequence facilitating RepA binding, and the like. The polynucleotide sequences may also comprise additional sequences encoding additional amino acids that encode for example a marker or a tag sequence such as a histidine tag or an HA tag to facilitate purification or detection of the protein, a signal sequence, a fusion protein partner such as RepA, Fc or bacteriophage coat protein such as pIX or pIII.
[0346] Another embodiment of the invention is a vector comprising at least one polynucleotide of the invention. Such vectors may be plasmid vectors, viral vectors, vectors for baculovirus expression, transposon based vectors or any other vector suitable for introduction of the polynucleotides of the invention into a given organism or genetic background by any means. Such vectors may be expression vectors comprising nucleic acid sequence elements that can control, regulate, cause or permit expression of a polypeptide encoded by such a vector. Such elements may comprise transcriptional enhancer binding sites, RNA polymerase initiation sites, ribosome binding sites, and other sites that facilitate the expression of encoded polypeptides in a given expression system. Such expression systems may be cell-based, or cell-free systems well known in the art.
[0347] Another embodiment of the invention is a host cell comprising the vector of the invention. A monospecific EGFR-binding or c-Met binding FN3 domain or the bispecific EGFR/c-Met FN3 domain containing molecule of the invention can be optionally produced by a cell line, a mixed cell line, an immortalized cell or clonal population of immortalized cells, as well known in the art. See, e.g., Ausubel, et al., ed., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., NY, NY (1987-2001); Sambrook, et al., Molecular Cloning: A Laboratory Manual, 2.sup.nd Edition, Cold Spring Harbor, N.Y. (1989); Harlow and Lane, Antibodies, a Laboratory Manual, Cold Spring Harbor, N.Y. (1989); Colligan, et al., eds., Current Protocols in Immunology, John Wiley & Sons, Inc., NY (1994-2001); Colligan et al., Current Protocols in Protein Science, John Wiley & Sons, NY, NY, (1997-2001).
[0348] The host cell chosen for expression may be of mammalian origin or may be selected from COS-1, COS-7, HEK293, BHK21, CHO, BSC-1, He G2, SP2/0, HeLa, myeloma, lymphoma, yeast, insect or plant cells, or any derivative, immortalized or transformed cell thereof. Alternatively, the host cell may be selected from a species or organism incapable of glycosylating polypeptides, e.g. a prokaryotic cell or organism, such as BL21, BL21(DE3), BL21-GOLD(DE3), XL1-Blue, JM109, HMS174, HMS174(DE3), and any of the natural or engineered E. coli spp, Klebsiella spp., or Pseudomonas spp strains.
[0349] Another embodiment of the invention is a method of producing the isolated FN3 domain specifically binding EGFR or c-Met of the invention or the isolated bispecific EGFR/c-Met FN3 domain containing molecule of the invention, comprising culturing the isolated host cell of the invention under conditions such that the isolated FN3 domain specifically binding EGFR or c-Met or the isolated bispecific EGFR/c-Met FN3 domain containing molecule is expressed, and purifying the domain or molecule.
[0350] The FN3 domain specifically binding EGFR or c-Met or the isolated bispecific EGFR/c-Met FN3 domain containing molecule of the invention can be purified from recombinant cell cultures by well-known methods, for example by protein A purification, ammonium sulfate or ethanol precipitation, acid extraction, anion or cation exchange chromatography, phosphocellulose chromatography, hydrophobic interaction chromatography, affinity chromatography, hydroxylapatite chromatography and lectin chromatography, or high performance liquid chromatography (HPLC).
Bispecific EGFR/c-Met Antibodies
[0351] The bispecific EGFR/c-Met antibodies may be generated de novo or may be engineered from existing monospecific anti-EGFR and anti-c-Met antibodies.
[0352] Exemplary anti-EGFR antibodies that may be used to engineer bispecific molecules are for example panitumumab (ABX-EGF), nimotuzumab, necitumumab, matuzumab, and those described for example in: U.S. Pat. Nos. 7,595,378, 7,247,301, U.S. Pat. Publ. No. US2011/0256142, U.S. Pat. Nos. 5,891,996, 5,212,290, 5,558,864, or U.S. Pat. No. 7,589,180. For example, antibody VH domain having the amino acid sequence shown in SEQ ID NO: 189 or 191 and antibody VL domain having the amino acid sequences shown in SEQ ID NO: 190 or 192 may be used.
[0353] Exemplary anti-c-Met antibodies that may be used to engineer bispecific molecules are for example Rilotumumab, Onartuzumab, Ficlatuzumab, and those described for example in PCT Intl. Publ. No. WO2011/110642, US Pat. Publ. No. US2004/0166544, PCT Intl. Publ. No. WO2005/016382, or PCT Intl. Publ. No. WO2006/015371. For example, antibody VH domain having the amino acid sequence shown in SEQ ID NO: 193 or 195 and antibody VL domain having the amino acid sequences shown in SEQ ID NO: 194 or 196 may be used. The heavy and light chain amino acid sequences of the antibodies identified by their United States Adopted Names (USAN) is available via the American Medical Association at http://_www_ama-assn_org or via the CAS registry.
[0354] Monospecific EGFR and c-Met biding variable domains may be selected de novo from for example a phage display library, where the phage is engineered to express human immunoglobulins or portions thereof such as Fabs, single chain antibodies (scFv), or unpaired or paired antibody variable regions (Knappik et al., J Mol Biol 296:57-86, 2000; Krebs et al., J Immunol Meth 254:67-84, 2001; Vaughan et al., Nature Biotechnology 14:309-314, 1996; Sheets et al., PITAS (USA) 95:6157-6162, 1998; Hoogenboom and Winter, J Mol Biol 227:381, 1991; Marks et al., J Mol Biol 222:581, 1991), and subsequently engineered into a bispecific format. The monospecific EGFR and c-Met binding variable domains may be isolated for example from phage display libraries expressing antibody heavy and light chain variable regions as fusion proteins with bacteriophage pIX coat protein as described in Shi et al (2010) J. Mol. Biol. 397:385-96 and PCT Intl. Publ. No. WO09/085462). The antibody libraries are screened for binding to human EGFR or c-Met extracellular domains and the obtained positive clones are further characterized and the Fabs isolated from the clone lysates. Such phage display methods for isolating human antibodies are established in the art. See for example: U.S. Pat. Nos. 5,223,409; 5,403,484; and 5,571,698, 5,427,908, 5,580,717, 5,969,108, 6,172,197, 5,885,793; 6,521,404; 6,544,731; 6,555,313; 6,582,915 and 6,593,081. The obtained de novo variable regions binding EGFR or c-Met are engineered to bispecific formats using the methods described herein.
Bispecific Antibody Formats
[0355] Antibodies of the present invention have two or more antigen binding sites and are bispecific. Bispecific antibodies of the invention include antibodies having a full length antibody structure.
[0356] "Full length antibody" as used herein refers to an antibody having two full length antibody heavy chains and two full length antibody light chains. A full length antibody heavy chain (HC) consists of well known heavy chain variable and constant domains VH, CH1, CH2, and CH3. A full length antibody light chain (LC) consists of well known light chain variable and constant domains VL and CL. The full length antibody may be lacking the C-terminal lysine (K) in either one or both heavy chains.
[0357] The term "Fab-arm" or "half molecule" refers to one heavy chain-light chain pair that specifically binds an antigen.
[0358] Full length bispecific antibodies of the invention may be generated for example using Fab arm exchange (or half molecule exchange) between two monospecific bivalent antibodies by introducing substitutions at the heavy chain CH3 interface in each half molecule to favor heterodimer formation of two antibody half molecules having distinct specificity either in vitro in cell-free environment or using co-expression. The Fab arm exchange reaction is the result of a disulfide-bond isomerization reaction and dissociation-association of CH3 domains. The heavy-chain disulfide bonds in the hinge regions of the parent monospecific antibodies are reduced. The resulting free cysteines of one of the parent monospecific antibodies form an inter heavy-chain disulfide bond with cysteine residues of a second parent monospecific antibody molecule and simultaneously CH3 domains of the parent antibodies release and reform by dissociation-association. The CH3 domains of the Fab arms may be engineered to favor heterodimerization over homodimerization. The resulting product is a bispecific antibody having two Fab arms or half molecules which each bind a distinct epitope, i.e. an epitope on EGFR and an epitope on c-Met.
[0359] "Homodimerization" as used herein refers to an interaction of two heavy chains having identical CH3 amino acid sequences. "Homodimer" as used herein refers to an antibody having two heavy chains with identical CH3 amino acid sequences.
[0360] "Heterodimerization" as used herein refers to an interaction of two heavy chains having non-identical CH3 amino acid sequences. "Heterodimer" as used herein refers to an antibody having two heavy chains with non-identical CH3 amino acid sequences.
[0361] The "knob-in-hole" strategy (see, e.g., PCT Intl. Publ. No. WO 2006/028936) may be used to generate full length bispecific antibodies. Briefly, selected amino acids forming the interface of the CH3 domains in human IgG can be mutated at positions affecting CH3 domain interactions to promote heterodimer formation. An amino acid with a small side chain (hole) is introduced into a heavy chain of an antibody specifically binding a first antigen and an amino acid with a large side chain (knob) is introduced into a heavy chain of an antibody specifically binding a second antigen. After co-expression of the two antibodies, a heterodimer is formed as a result of the preferential interaction of the heavy chain with a "hole" with the heavy chain with a "knob". Exemplary CH3 substitution pairs forming a knob and a hole are (expressed as modified position in the first CH3 domain of the first heavy chain/modified position in the second CH3 domain of the second heavy chain): T366Y/F405A, T366W/F405W, F405W/Y407A, T394W/Y407T, T394S/Y407A, T366W/T394S, F405W/T394S and T366W/T366S_L368A_Y407V.
[0362] Other strategies such as promoting heavy chain heterodimerization using electrostatic interactions by substituting positively charged residues at one CH3 surface and negatively charged residues at a second CH3 surface may be used, as described in US Pat. Publ. No. US2010/0015133; US Pat. Publ. No. US2009/0182127; US Pat. Publ. No. US2010/028637 or US Pat. Publ. No. US2011/0123532. In other strategies, heterodimerization may be promoted by following substitutions (expressed as modified position in the first CH3 domain of the first heavy chain/modified position in the second CH3 domain of the second heavy chain): L351Y_F405A_Y407V/T394W, T366I_K392M_T394W/F405A_Y407V, T366L_K392M_T394W/F405A_Y407V, L351Y_Y407A/T366A_K409F, L351Y_Y407A/T366V_K409F, Y407A/T366A_K409F, or T350V_L351Y_F405A_Y407V/T350V_T366L_K392L_T394W as described in U.S. Pat. Publ. No. US2012/0149876 or U.S. Pat. Publ. No. US2013/0195849
[0363] In addition to methods described above, bispecific antibodies of the invention may be generated in vitro in a cell-free environment by introducing asymmetrical mutations in the CH3 regions of two monospecific homodimeric antibodies and forming the bispecific heterodimeric antibody from two parent monospecific homodimeric antibodies in reducing conditions to allow disulfide bond isomerization according to methods described in Intl. Pat. Publ. No. WO2011/131746. In the methods, the first monospecific bivalent antibody (e.g., anti-c-Met antibody) and the second monospecific bivalent antibody (e.g., anti-EGFR antibody) are engineered to have certain substitutions at the CH3 domain that promoter heterodimer stability; the antibodies are incubated together under reducing conditions sufficient to allow the cysteines in the hinge region to undergo disulfide bond isomerization; thereby generating the bispecific antibody by Fab arm exchange. The incubation conditions may optimally be restored to non-reducing. Exemplary reducing agents that may be used are 2-mercaptoethylamine (2-MEA), dithiothreitol (DTT), dithioerythritol (DTE), glutathione, tris(2-carboxyethyl)phosphine (TCEP), L-cysteine and beta-mercaptoethanol, preferably a reducing agent selected from the group consisting of: 2-mercaptoethylamine, dithiothreitol and tris(2-carboxyethyl)phosphine. For example, incubation for at least 90 min at a temperature of at least 20.degree. C. in the presence of at least 25 mM 2-MEA or in the presence of at least 0.5 mM dithiothreitol at a pH of from 5-8, for example at pH of 7.0 or at pH of 7.4 may be used.
Bispecific EGFR/c-Met Antibodies
[0364] The bispecific EGFR/c-Met antibodies of the invention may provide a benefit in terms of specificity and reduced off-target toxicity when compared to small molecule EGFR and/or c-Met inhibitors. The present invention is based at least in part on the surprising finding that the bispecific EGFR/c-Met antibodies of the invention provide a significantly improved synergistic inhibitory effect when compared to a mixture of EGFR-binding and c-Met-binding monospecific antibodies or published bispecific EGFR/c-Met antibodies. Depending on the assay, the synergistic effect observed varied between about 14- to over about 800-fold. The bispecific EGFR/c-Met antibodies of the invention provide more efficient inhibition of EGFR and c-Met signaling pathways and inhibit tumor growth more efficiently than cetuximab (Erbitux.RTM.). The bispecific EGFR/c-Met antibodies of the invention inhibit EGFR signaling in tumors and/or tumor cell lines having EGFR activating mutations and/or mutations in EGFR that are known to result in resistance to treatments with tyrosine kinase inhibitors such as gefitinib, and inhibit c-Met signaling pathway, a pathway identified to be upregulated and to provide a compensatory signaling upon treatment with EGFR tyrosine kinase inhibitors in cancers such as NSCLC. The bispecific EGFR/c-Met antibodies of the invention, in addition to directly inhibiting EGFR and c-Met signaling, display antitumor activity through enhanced antibody dependent cell cytotoxicity (ADCC) and degradation of the EGFR and c-Met receptors. Contrary to the current EGFR therapies (cetuximab and panitumumab), the bispecific EGFR/c-Met antibodies of the invention induce, via enhanced ADCC, killing of tumor cells having KRAS mutations.
[0365] Int. Pat. Publ. No. WO2010/115551 describes a bispecific EGFR/c-Met antibody (BSAB01) engineered in an IgG-scFv format using the EGFR binding VH/VL pair of cetuximab, and the c-Met binding VH/VL pair of an antibody 5D5 (MetMab, onartuzumab) currently in Phase III trials. BSAB01 demonstrates approximately two-fold (additive) increased inhibition of A431 cell proliferation when compared to the parental antibodies (Example 7, FIG. 8b in WO2010/115551), and a modest additive inhibition of Ovarc-8 cell proliferation (FIG. 10a, Example 16 in WO2010/115551) when compared to the combination of the two parental antibodies (15% vs. 10% inhibition). Therefore, surprisingly and unexpectedly, the present invention provides bispecific EGFR/c-Met antibodies that demonstrate a significant synergistic effect in inhibition of EGFR and c-Met signaling, cancer cell survival and tumor growth. By not wishing to be bound by any theory, it is believed that the significant synergistic effect of the bispecific antibodies of the invention at least partially results from the epitope specificity of both the EGFR and the c-Met binding arms, possibly resulting in the inhibition of signaling through not only the EGFR and c-Met homodimers but also the EGFR/HERx heterodimers.
[0366] One embodiment of the invention is an isolated bispecific epidermal growth factor receptor (EGFR)/hepatocyte growth factor receptor (c-Met) antibody, comprising: [0367] a) a first heavy chain (HC1) comprising a HC1 constant domain 3 (HC1 CH3) and a HC1 variable region 1 (VH1); [0368] b) a second heavy chain (HC2) comprising a HC2 constant domain 3 (HC2 CH3) and a HC2 variable region 2 (VH2); [0369] c) a first light chain (LC1) comprising a light chain variable region 1 (VL1); and
[0370] a second light chain (LC2) comprising a light chain variable region 2 (VL2), wherein the VH1 and the VL1 pair to form a first antigen-binding site that specifically binds EGFR and the VH2 and the VL2 pair to form a second antigen-binding site that specifically binds c-Met, wherein the HC1 comprises at least one substitution in the HC1 CH3 and the HC2 comprises at least one substitution in the HC2 CH3, wherein the substitution in the HC1 CH3 and the substitution in the HC2 CH3 occur at different amino acid residue positions, when residue numbering is according to the EU index.
[0371] In some embodiments described herein, the bispecific EGFR/c-Met antibody inhibits phosphorylation of extracellular signal-related kinases 1 and 2 (ERK1/2) in NCI-H292, NCI-H1975 or SKMES-1 cell line with an IC.sub.50 value that is at least about 10-fold less, at least about 20-fold less, at least about 30-fold less, at least about 40-fold less, at least about 50-fold less or at least about 60-fold less when compared to the IC.sub.50 value of inhibition of phosphorylation of ERK1/2 in NCI-H292, NCI-H1975 or SKMES-1 cell line with a mixture of a control monovalent EGFR antibody comprising a heavy chain 3 (HC3) and a light chain 3 (LC3) and a control monovalent c-Met antibody comprising a heavy chain 4 (HC4) and a light chain 4 (LC4), wherein the HC3 and the HC1, the LC3 and the LC1, the HC4 and the HC2, and the LC4 and the LC2 have identical amino acid sequences, respectively, and the phosphorylation of ERK1/2 is measured in whole cell lysates using a sandwich immunoassay using an anti-phosphoERK1/2 antibody as a capture antibody and an antibody binding to unphosphorylated and phosphorylated ERK1/2 conjugated with an electrochemiluminescent compound as a detection antibody. The bispecific EGFR/c-Met antibodies of the invention provide a synergistic more pronounced inhibition of EGFR and c-Met signaling when compared to the combination of monospecific EGFR antibodies and monospecific c-Met antibodies, when inhibition is assessed by inhibition of ERK1/2 phosphorylation. Such exemplary bispecific EGFR/c-Met antibody is the antibody EM1-mAb of the invention.
[0372] "Control monospecific EGFR antibody" as used herein refers to an antibody that has a first Fab arm that binds EGFR that is identical in amino acid sequence to the EGFR-binding Fab arm of the bispecific EGFR/c-Met antibody to be tested, and has a second Fab arm that is "inert" and binds an unrelated/irrelevant antigen, human immunodeficiency virus (HIV) gp120. The second Fab arm has a light chain having the sequence of SEQ ID NO: 209 and a heavy chain having the sequence of SEQ ID NO: 198 in instances when the EGFR binding Fab arm in the bispecific EGFR/c-Met antibody to be tested comprises the F405L substitution. The second Fab arm has a light chain having the sequence of SEQ ID NO: 209 and a heavy chain having the sequence of SEQ ID NO: 197 in instances when the EGFR binding Fab arm in the bispecific EGFR/c-Met antibody to be tested comprises the K409R substitution.
[0373] "Control monospecific c-Met antibody" as used herein refers to an antibody that has a first Fab arm that binds c-Met that is identical in amino acid sequence to the c-Met-binding Fab arm of the bispecific EGFR/c-Met antibody to be tested, and has a second Fab arm that is "inert" and binds the unrelated/irrelevant antigen HIV gp120. The second Fab Fab arm has a light chain having the sequence of SEQ ID NO: 209 and a heavy chain having the sequence of SEQ ID NO: 198 in instances when the c-Met binding Fab arm in the bispecific EGFR/c-Met antibody to be tested comprises the F405L substitution. The second inert Fab arm has a light chain having the sequence of SEQ ID NO: 209 and a heavy chain having the sequence of SEQ ID NO: 197 in instances when the c-Met binding Fab arm in the bispecific EGFR/c-Met antibody to be tested comprises the K409R substitution.
[0374] In some embodiments described herein, the bispecific EGFR/c-Met antibody inhibits phosphorylation of ERK1/2 with an IC.sub.50 value of about 2.times.10.sup.-9 M or less, about 1.times.10.sup.-9 M or less, or about 1.times.10.sup.-10 M or less.
[0375] In some embodiments described herein, ERK1 is phosphorylated at residues Thr202 and Tyr204, and ERK2 is phosphorylated at residues Thr185 and Tyr197.
[0376] In some embodiments described herein, the bispecific EGFR/c-Met antibody inhibits phosphorylation of protein kinase B (AKT) at Ser473 in NCI-H1975 cell line with an IC.sub.50 value that is at least about 70-fold less when compared to the IC.sub.50 value of inhibition of phosphorylation of AKT at Ser473 in NCI-H1975 cell line with the mixture of the control monovalent EGFR antibody comprising the HC3 and the LC3 and the control monovalent c-Met antibody comprising the HC4 and the LC4, wherein the HC3 and the HC1, the LC3 and the LC1, the HC4 and the HC2, and the LC4 and the LC2 have identical amino acid sequences, respectively, wherein the phosphorylation of AKT at Ser473 is measured in whole cell lysates using a sandwich immunoassay using an antibody binding to unphosphorylated and phosphorylated AKT as a capture antibody and an anti-phosphoAKT Ser473 antibody conjugated to an electrochemiluminescent compound as a detection antibody.
[0377] In some embodiments described herein, the bispecific EGFR/c-Met antibody inhibits phosphorylation of protein kinase B (AKT) at Thr308 in NCI-H1975 cell line with an IC.sub.50 value that is at least about 100-fold less when compared to the IC.sub.50 value of inhibition of phosphorylation of AKT at Thr308 in NCI-H1975 cell line with the mixture of the control monovalent EGFR antibody comprising the HC3 and the LC3 and the control monovalent c-Met antibody comprising the HC4 and the LC4, wherein the HC3 and the HC1, the LC3 and the LC1, the HC4 and the HC2, and the LC4 and the LC2 have identical amino acid sequences, respectively, wherein the phosphorylation of AKT at Thr308 is measured in whole cell lysates using a sandwich immunoassay using an antibody binding to unphosphorylated and phosphorylated AKT as a capture antibody and an anti-phosphoAKT Thr308 antibody conjugated to an electrochemiluminescent compound as a detection antibody.
[0378] The bispecific EGFR/c-Met antibodies of the invention provide a synergistic more pronounced inhibition of EGFR and c-Met signaling when compared to the combination of monospecific EGFR antibodies and monospecific c-Met antibodies, when inhibition is assessed by inhibition of AKT phosphorylation. Such exemplary bispecific EGFR/c-Met antibody is the antibody EM1-mAb of the invention.
[0379] In some embodiments described herein, the bispecific EGFR/c-Met antibody inhibits phosphorylation of AKT at Ser473 or at Thr308 with and IC.sub.50 value of about 1.times.10.sup.-9 M or less.
[0380] In some embodiments described herein, the bispecific EGFR/c-Met antibody binds EGFR of SEQ ID NO: 73 at EGFR residues K489, 1491, K467 and 5492 and c-Met at residues PEFRDSYPIKYVHAF (SEQ ID NO: 238) and FAQSKPDSAEPMDRSA (SEQ ID NO: 239). Such an exemplary bispecific antibody is the EM1-mAb. The bispecific EM-1 antibody binds EGFR and c-Met at distinct epitopes when compared to the antibody BSAB01 as described above and in Int. Pat. Publ. No. WO2010/115551. The parental EGFR binding arm (cetuximab) of BSAB01 binds EGFR amino acid residues R353, Q384, Q408, H409, F412, S418, S440, K443, K465, I467, S468, and N473 in mature EGFR, corresponding to residues R367, Q408, Q432, H433, F436, S442, S464, K467, K489, I491, S492 and N497 of full length EGFR of SEQ ID NO: 73 (Li et al., Cancer Cell 7:301-311, 2005). The parental c-Met binding arm of BSAB01 (mAb 5D5) binds c-Met residues 325-340 PGAQLARQIGASLNDD (SEQ ID NO: 240). Epitope mapping of the EGFR binding parental antibody (2F8) of the EM1-mAb is described in US. Pat. Publ. No. US2011/0256142A1. Cetuximab and the parental 2F8 antibody bind partially overlapping but distinct epitopes.
[0381] Epitope mapping can be done using standard methods. For example, when the structures of both individual components are known, in silica protein-protein docking can be carried out to identify compatible sites of interaction. Hydrogen-deuterium (H/D) exchange can be carried out with the antigen and antibody complex to map regions on the antigen that may be bound by the antibody. Segment and point mutagenesis of the antigen can be used to locate amino acids important for antibody binding.
[0382] In some embodiments described herein, the bispecific EGFR/c-Met antibody neutralizes EGFR and c-Met signaling.
[0383] The bispecific EGFR/c-Met antibody of the invention may neutralize EGFR and c-Met signaling by at least 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% or 100% when compared to the level of signaling in the absence of the bispecific EGFR/c-Met molecule of the invention using the same assay conditions.
[0384] Binding of a ligand such as EGF to EGFR stimulates receptor dimerization, autophosphorylation, activation of the receptor's internal, cytoplasmic tyrosine kinase domain, and initiation of multiple signal transduction and transactivation pathways involved in regulation of DNA synthesis (gene activation) and cell cycle progression or division. Neutralization of EGFR signaling may result in inhibition in one or more EGFR downstream signaling pathways and therefore neutralizing EGFR may have various effects, including inhibition of cell proliferation and differentiation, angiogenesis, cell motility and metastasis, and inhibition of downstream signaling pathways.
[0385] EGFR signaling and neutralization of EGFR signaling may be measured using various well know methods, for example measuring the autophosphorylation of the receptor at any of the tyrosines Y1068, Y1148, and Y1173 (Downward et al., Nature 311:483-5, 1984) and/or phosphorylation of natural or synthetic substrates, and inhibition of autophosphorylation and/or phosphorylation of natural or synthetic substrates by the bispecific antibodies of the invention. Phosphorylation can be detected using well known methods such as an ELISA assay or a western plot using a phosphotyrosine specific antibody. Exemplary assays can be found in Panek et al., J Pharmacol Exp Thera 283:1433-44, 1997 and Batley et al., Life Sci 62:143-50, 1998, and as described herein.
[0386] Binding of HGF to c-Met stimulates receptor dimerization, autophosphorylation, activation of the receptor's cytoplasmic tyrosine kinase domain, and initiation of multiple signal transduction and transactivation pathways involved in regulation of DNA synthesis (gene activation) and cell cycle progression or division. Inhibition of c-Met signaling may result in inhibition in one or more c-Met downstream signaling pathways and therefore neutralizing c-Met may have various effects, including inhibition of cell proliferation and differentiation, angiogenesis, cell motility and metastasis.
[0387] c-Met signaling and neutralization of c-Met signaling may be measured using various well know methods, for example measuring the autophosphorylation of the receptor on at least one tyrosine residues Y1230, Y1234, Y1235 or Y1349, and/or phosphorylation of natural or synthetic substrates. Phosphorylation can be detected, for example, using an antibody specific for phosphotyrosine in an ELISA assay or on a western blot. Exemplary assays can be found in Panek et al., J Pharmacol Exp Thera 283:1433-44, 1997 and Batley et al., Life Sci 62:143-50, 1998, and as described herein.
[0388] EGFR and c-Met signaling may be measured using various well know methods as described herein, such as measuring inhibition of ERK1/2 and AKT phosphorylation. Inhibition of ERK1 phosphorylation at Thr202 and Tyr204 and ERK2 phosphorylation at Thr185 and Tyr187 and inhibition of AKT at Ser473 or Thr308 can be measured for example in NCI-H1975 cell lysates utilizing a sandwich assay with capture antibody coated on solid support, and the detection antibody conjugated with an electrochemiluminescent compound such as Meso Scale Discover (MSD) SULFO-TAG label, followed by detection of the signal with a plate reader.
[0389] In some embodiments described herein, the bispecific EGFR/c-Met antibody inhibits growth of NCI-H292 or NCI-H1975 cells with an IC.sub.50 value that is at least about 300-fold less, at least about 400-fold less, at least about 500-fold less, at least about 600-fold less, at least about 700-fold less or at least about 800-fold less when compared to the IC.sub.50 value of inhibition of growth of NCI-H292 or NCI-H1975 cells with cetuximab, when NCI-H292 or NCI-H1975 cells are grown in low attachment conditions.
[0390] Inhibition of cell growth may be assessed by known methods. For example, the cells may be plated in plates coated with hydrogels or biomimetic polymers (for example Ultra Low Attachment plates by Corning) to prevent or reduce cell attachment, and the effect of antibodies on 7.5 ng/mL HGF-induced cell growth can be assessed by measuring percent cell viability after incubation for 72 hours using standard methods.
[0391] The bispecific EGFR/c-Met antibodies of the invention provide a synergistic more pronounced inhibition of EGFR and/or c-Met expressing cancer cells when compared to the combination of monospecific EGFR antibodies and monospecific c-Met antibodies and to the standard of care cetuximab. Such an exemplary bispecific EGFR/c-Met antibody is the antibody EM1-mAb of the invention. The bispecific EGFR/c-Met antibodies of the invention inhibit cancer cells that express the wild type EGFR and the wild type c-Met, and also cancer cells that express the EGFR L858R/T790M mutant, which mutation is identified to contribute to resistance to treatments with small molecule tyrosine kinase inhibitors (TKIs) such as gefitinib. Therefore the bispecific EGFR/c-Met antibodies of the invention may provide a benefit in a broader patient population when compared to cetuximab and TKIs.
[0392] In some embodiments described herein, the bispecific EGFR/c-Met antibody inhibits growth of HGF-expressing SKMES-1 cell tumor in SCID Beige mice with a percentage (%) T/C value of at least 500-fold less on day 36 when compared to cetuximab, when the bispecific antibody and cetuximab are administered at 20 mg/kg dose.
[0393] Tumor xenograft models using SCID Beige mice are well known. SKMES-1 cells may be engineered to express human HGF using standard methods. Typically, SCID Beige mice may be subcutaneously inoculated with SKMES-1 cells expressing human HGF embedded in extracellular matrix such as Culturex in the dorsal flank of each animal. One week after implantation, mice may be stratified into groups with equivalent tumor volumes, and thereafter dosed for example three times per week with the bispecific EGFR/c-Met antibodies of the invention, control or benchmark antibodies or small molecules. Tumor volumes may be recorded twice weekly, and tumor growth inhibition (TGI) may be observed by calculating the percentage (%) T/C value. The % T/C value is indicative of anti-tumor efficacy. T and C are the mean volumes of the treated and control groups, respectively, on a given day.
[0394] The bispecific EGFR/c-Met antibodies of the invention provide a significantly improved efficacy in in vivo tumor killing when compared to the standard of care cetuximab, and therefore may provide a benefit in a patient population when compared to cetuximab.
[0395] In some embodiments described herein, the bispecific EGFR/c-Met antibody heterodimerizes EGFR and c-Met on the cell surface. While not wishing to be bound by any particular theory, it is believed that the binding of the antibody of the invention to EGFR homodimers, c-Met homodimers, and EGFR/c-Met heterodimers results in simultaneous inhibition of ligand mediated signaling downstream of EGFR homodimers, c-Met homodimers and EGFR/c-Met heterodimers, providing synergistic inhibition when compared to the combination of monospecific anti-EGFR and anti-c-Met antibodies.
[0396] In some embodiments described herein, the bispecific EGFR/c-Met antibody comprises the HC1 and the HC2 of IgG1, IgG2, IgG3 or IgG4 isotype.
[0397] In some embodiments described herein, the bispecific EGFR/c-Met antibody comprises the HC1 and the HC2 of IgG1 isotype.
[0398] In some embodiments described herein, the bispecific EGFR/c-Met antibody HC1 CH3 comprises at least one, two, three, four, five, six, seven or eight substitutions and the HC2 CH3 comprises at least one, two, three, four, five, six, seven or eight substitutions at residue positions 350, 366, 368, 370, 399, 405, 407 or 409, when residue numbering is according to the EU index.
[0399] In some embodiments described herein, the bispecific EGFR/c-Met antibody HC1 CH3 comprises at least one, two, three or four substitutions and the HC2 CH3 comprises at least one, two, three or four substitutions at residue positions 350, 370, 405 or 409, when residue numbering is according to the EU index.
[0400] Antibody domains and numbering are well known. Two CH3 domains (or CH3 regions) are non-identical when they differ with at least one amino acid substitution from each other. An IgG1 CH3 region typically consists of residues 341-446 on IgG1 (residue numbering according to the EU index). An exemplary IgG1 constant region is shown in SEQ ID NO: 203. The CH3 domain spans residues 224-329 of SEQ ID NO: 203, and correspond to residues 341-446 according to EU index.
[0401] In some embodiments described herein, the bispecific EGFR/c-Met antibody HC1 CH3 comprises at least one substitution and the HC2 CH3 comprises at least one substitution at residue positions 405 or 409, when residue numbering is according to the EU index.
[0402] In some embodiments described herein, the bispecific EGFR/c-Met antibody HC1 CH3 comprises a K409R or a F405L substitution and the HC2 CH3 comprises a K409R or a F405L substitution, wherein residue numbering is according to the EU index.
[0403] In some embodiments described herein, the bispecific EGFR/c-Met antibody HC1 CH3 comprises the F405L substitution and the HC2 CH3 comprises the K409R substitution.
[0404] In some embodiments described herein, the HC1 CH3 and the HC2 CH3 substitutions are substitutions at position 366, 368, 370, 399, 405, 407 or 409 (numbering according to the EU index). These positions correspond to linear residue positions 248, 250, 252, 281, 287, 289 and 291, respectively, in a heavy chain constant region of SEQ ID NO: 203 and 204.
[0405] In some embodiments described herein, the HC1 CH3 position 409 has an amino acid substitution other than Lys, Leu or Met and the HC2 CH3 position 405 has an amino acid substitution other than Phe.
[0406] In some embodiments described herein, the HC1 CH3 position 405 has an amino acid substitution other than Phe and the HC2 CH3 position 409 has an amino acid substitution other than Lys, Leu or Met.
[0407] In some embodiments described herein, the HC1 CH3 position 409 has an amino acid substitution other than Lys, Leu or Met and the HC2 CH3 position 405 has an amino acid substitution other than Phe, Arg or Gly.
[0408] In some embodiments described herein, the HC1 CH3 position 405 has an amino acid substitution other than Phe, Arg or Gly and the HC2 CH3 position 409 has an amino acid substitution other than Lys, Leu or Met
[0409] In some embodiments described herein, the HC1 CH3 has Phe at position 405 and an amino acid other than Lys, Leu or Met at position 409 and the HC2 CH3 has an amino acid other than Phe at position 405 and a Lys at position 409.
[0410] In some embodiments described herein, the HC1 CH3 has an amino acid other than Phe at position 405 and Lys at position 409 and the HC2 CH3 has Phe at position 405 and an amino acid other than Lys, Leu or Met at position 409.
[0411] In some embodiments described herein, the HC1 CH3 has Phe at position 405 and an amino acid other than Lys, Leu or Met at position 409 and the HC2 CH3 has a substitution other than Phe, Arg or Gly at position 405 and Lys at position 409.
[0412] In some embodiments described herein, the HC1 CH3 has a substitution other than Phe, Arg or Gly at position 405 and Lys at position 409 and the HC2 CH3 has Phe at position 405 and an amino acid other than Lys, Leu or Met at position 409.
[0413] In some embodiments described herein, the HC1 CH3 has Phe at position 405 and an amino acid other than Lys, Leu or Met at position 409 and the HC2 CH3 has Leu at position 405 and Lys at position 409.
[0414] In some embodiments described herein, the HC1 CH3 has Leu at position 405 and Lys at position 409 and the HC2 CH3 has Phe at position 405 and an amino acid other than Lys, Leu or Met at position 409.
[0415] In some embodiments described herein, the HC1 CH3 has Phe at position 405 and aArg at position 409 and the HC2 CH3 has an amino acid other than Phe, Arg or Gly at position 405 and Lys at position 409.
[0416] In some embodiments described herein, the HC1 CH3 has an amino acid other than Phe, Arg or Gly at position 405 and Lys at position 409 and the HC2 CH3 has Phe at position 405 and Arg at position 409.
[0417] In some embodiments described herein, the HC1 CH3 has Phe at position 405 and Arg at position 409 and the HC2 CH3 has Leu at position 405 and Lys at position 409.
[0418] In some embodiments described herein, the HC1 CH3 has Leu at position 405 and Lys at position 409 and the HC2 CH3 has Phe at position 405 and Arg at position 409.
[0419] In some embodiments described herein, the HC1 CH3 has Phe at position 405 and Lys at position 409 and the HC2 CH3 has Leu at position 405 and aArg at position 409.
[0420] In some embodiments described herein, the HC1 CH3 has Leu at position 405 and aArg at position 409 and the HC2 CH3 has Phe at position 405 and Lys at position 409.
[0421] In some embodiments described herein, the HC1 CH3 has an amino acid other than Lys, Leu or Met at position 409 and the HC2 CH3 has Lys at position 409, Thr at position 370 and Leu at position 405.
[0422] In some embodiments described herein, the HC1 CH3 has Lys at position 409, Thr at position 370 and Leu at position 405 and the HC2 CH3 has an amino acid other than Lys, Leu or Met at position 409.
[0423] In some embodiments described herein, the HC1 CH3 has Arg at position 409 and the HC2 CH3 has Lys at position 409, Thr at position 370 and Leu at position 405.
[0424] In some embodiments described herein, the HC1 CH3 has Lys at position 409, Thr at position 370 and Leu at position 405 and the HC2 CH3 has Arg at position 409.
[0425] In some embodiments described herein, the HC1 CH3 has Lys at position 370, Phe at position 405 and aArg at position 409 and the HC2 CH3 has Lys at position 409, Thr at position 370 and Leu at position 405.
[0426] In some embodiments described herein, the HC1 CH3 has Lys at position 409, Thr at position 370 and Leu at position 405 and the HC2 CH3 has Lys at position 370, Phe at position 405 and Arg at position 409.
[0427] In some embodiments described herein, the HC1 CH3 has an amino acid other than Lys, Leu or Met at position 409 and the HC2 CH3 has an amino acid other than Tyr, Asp, Glu, Phe, Lys, Gln, Arg, Ser or Thr at position 407.
[0428] In some embodiments described herein, the HC1 CH3 has an amino acid other than Tyr, Asp, Glu, Phe, Lys, Gln, Arg, Ser or Thr at position 407 and the HC2 CH3 has an amino acid other than Lys, Leu or Met at position 409.
[0429] In some embodiments described herein, the HC1 CH3 has an amino acid other than Lys, Leu or Met at position 409 and the HC2 CH3 has Ala, Gly, His, Ile, Leu, Met, Asn, Val or Trp at position 407.
[0430] In some embodiments described herein, the HC1 CH3 has Ala, Gly, His, Ile, Leu, Met, Asn, Val or Trp at position 407 and the HC2 CH3 has an amino acid other than Lys, Leu or Met at position 409.
[0431] In some embodiments described herein, the HC1 CH3 has an amino acid other than Lys, Leu or Met at position 409 and the HC2 CH3 has Gly, Leu, Met, Asn or Trp at position 407.
[0432] In some embodiments described herein, the HC1 CH3 has Gly, Leu, Met, Asn or Trp at position 407 and the HC2 CH3 has an amino acid other than Lys, Leu or Met at position 409.
[0433] In some embodiments described herein, the HC1 CH3 has Tyr at position 407 and an amino acid other than Lys, Leu or Met at position 409 and the HC2 CH3 has an amino acid other than Tyr, Asp, Glu, Phe, Lys, Gln, Arg, Ser or Thr at position 407 and Lys at position 409.
[0434] In some embodiments described herein, the HC1 CH3 has an amino acid other than Tyr, Asp, Glu, Phe, Lys, Gln, Arg, Ser or Thr at position 407 and Lys at position 409 and the HC2 CH3 has Tyr at position 407 and an amino acid other than Lys, Leu or Met at position 409.
[0435] In some embodiments described herein, the HC1 CH3 has Tyr at position 407 and an amino acid other than Lys, Leu or Met at position 409 and the HC2 CH3 has Ala, Gly, His, Ile, Leu, Met, Asn, Val or Trp at position 407 and Lys at position 409.
[0436] In some embodiments described herein, the HC1 CH3 has Ala, Gly, His, Ile, Leu, Met, Asn, Val or Trp at position 407 and Lys at position 409 and the HC2 CH3 has Tyr at position 407 and an amino acid other than Lys, Leu or Met at position 409.
[0437] In some embodiments described herein, the HC1 CH3 has Tyr at position 407 and an amino acid other than Lys, Leu or Met at position 409 and the HC2 CH3 has Gly, Leu, Met, Asn or Trp at position 407 and Lys at position 409.
[0438] In some embodiments described herein, the HC1 CH3 has Gly, Leu, Met, Asn or Trp at position 407 and Lys at position 409 and the HC2 CH3 has Tyr at position 407 and an amino acid other than Lys, Leu or Met at position 409.
[0439] In some embodiments described herein, the HC1 CH3 has Tyr at position 407 and Arg at position 409 and the HC2 CH3 has an amino acid other than Tyr, Asp, Glu, Phe, Lys, Gln, Arg, Ser or Thr at position 407 and Lys at position 409.
[0440] In some embodiments described herein, the HC1 CH3 has an amino acid other than Tyr, Asp, Glu, Phe, Lys, Gln, Arg, Ser or Thr at position 407 and Lys at position 409 and the HC2 CH3 has Tyr at position 407 and Arg at position 409.
[0441] In some embodiments described herein, the HC1 CH3 has Tyr at position 407 and Arg at position 409 and the HC2 CH3 has Ala, Gly, His, Ile, Leu, Met, Asn, Val or Trp at position 407 and Lys at position 409.
[0442] In some embodiments described herein, the HC1 CH3 has Ala, Gly, His, Ile, Leu, Met, Asn, Val or Trp at position 407 and Lys at position 409 and the HC2 CH3 has Tyr at position 407 and Arg at position 409.
[0443] In some embodiments described herein, the HC1 CH3 has Tyr at position 407 and Arg at position 409 and the HC2 CH3 has Gly, Leu, Met, Asn or Trp at position 407 and Lys at position 409.
[0444] In some embodiments described herein, the HC1 CH3 has Gly, Leu, Met, Asn or Trp at position 407 and Lys at position 409 and the HC2 CH3 has Tyr at position 407 and Arg at position 409.
[0445] In some embodiments described herein, the HC1 CH3 has an amino acid other than Lys, Leu or Met at position 409, and the HC2 CH3 has (i) an amino acid other than Phe, Leu and Met at position 368, or (ii) a Trp at position 370, or (iii) an amino acid other than Asp, Cys, Pro, Glu or Gln at position 399.
[0446] In some embodiments described herein, the HC1 CH3 has (i) an amino acid other than Phe, Leu and Met at position 368, or (ii) a Trp at position 370, or (iii) an amino acid other than Asp, Cys, Pro, Glu or Gln at position 399 and the HC2 CH3 has an amino acid other than Lys, Leu or Met at position 409.
[0447] In some embodiments described herein, the HC1 CH3 has Arg, Ala, His or Gly at position 409, and the HC2 CH3 has (i) Lys, Gln, Ala, Asp, Glu, Gly, His, Ile, Asn, Arg, Ser, Thr, Val, or Trp at position 368, or (ii) Trp at position 370, or (iii) Ala, Gly, Ile, Leu, Met, Asn, Ser, Thr, Trp, Phe, His, Lys, Arg or Tyr at position 399.
[0448] In some embodiments described herein, the HC1 CH3 has (i) Lys, Gln, Ala, Asp, Glu, Gly, His, Ile, Asn, Arg, Ser, Thr, Val, or Trp at position 368, or (ii) Trp at position 370, or (iii) Ala, Gly, Ile, Leu, Met, Asn, Ser, Thr, Trp, Phe, His, Lys, Arg or Tyr at position 399 and the HC2 CH3 has Arg, Ala, His or Gly at position 409.
[0449] In some embodiments described herein, the HC1 CH3 has Arg at position 409, and the HC2 CH3 has (i) Asp, Glu, Gly, Asn, Arg, Ser, Thr, Val, or Trp at position 368, or (ii) Trp at position 370, or (iii) Phe, His, Lys, Arg or Tyr at position 399.
[0450] In some embodiments described herein, the HC1 CH3 has (i) Asp, Glu, Gly, Asn, Arg, Ser, Thr, Val, or Trp at position 368, or (ii) Trp at position 370, or (iii) Phe, His, Lys, Arg or Tyr at position 399 and the HC2 CH3 has Arg at position 409.
[0451] In some embodiments described herein, the HC1 CH3 comprises a K409R substitution or a F405L substitution and the HC2 CH3 comprises a K409R substitution or a F405L substitution, wherein the residue numbering is according to the EU index.
[0452] In some embodiments described herein, the HC1 CH3 comprises the F405L substitution and the HC2 CH3 comprises the K409R substitution.
[0453] Substitutions are typically made at the DNA level to a molecule such as the constant domain of the antibody using standard methods.
[0454] In some embodiments described herein, the bispecific EGFR/c-Met antibody comprises the VH1 and the VL1, wherein
[0455] the VH1 comprises the heavy chain complementarity determining region (HCDR) 1 (HCDR1), HCDR 2 (HCDR2) and HCDR 3 (HCDR3) amino acid sequences of SEQ ID NOs: 210, 211 and 212, respectively; and
[0456] the VL1 comprises the light chain complementarity determining region (LCDR) 1 (LCDR1), LCDR 2 (LCDR2) and LCDR 3 (LCDR3) amino acid sequences of SEQ ID NOs: 213, 214 and 215, respectively.
[0457] In some embodiments described herein, the bispecific EGFR/c-Met antibody comprises the VH2 and the VL2, wherein
[0458] the VH2 comprises the HCDR1, the HCDR2, and the HCDR3 amino acid sequences of SEQ ID NOs: 216, 217 and 218, respectively; and
[0459] the VL2 comprises the LCDR1, the LCDR2 and the LCDR3 amino acid sequences of SEQ ID NOs: 219, 220 and 221, respectively.
[0460] In some embodiments described herein, the bispecific EGFR/c-Met antibody comprises the VH1, the VL1, the VH2 and the VL2 amino acid sequences of SEQ ID NOs: 189, 190, 193 and 194, respectively.
[0461] In some embodiments described herein, the bispecific EGFR/c-Met antibody comprises the HC1, the LC1, the HC2 and the LC2 amino acid sequences of SEQ ID NOs: 199, 200, 201 and 202, respectively, optionally having a C-terminal lysine removed from the HC1, the HC2, or both the HC1 and the HC2.
[0462] In some embodiments described herein, the bispecific EGFR/c-Met antibody comprises the VH1 and the VL1, wherein
[0463] the VH1 comprises the HCDR1, the HCDR2, and the HCDR3 amino acid sequences of SEQ ID NOs: 222, 223 and 224, respectively; and
[0464] the VL1 comprises the LCDR1, the LCDR2 and the LCDR3 amino acid sequences of SEQ ID NOs: 225, 226 and 227, respectively.
[0465] In some embodiments described herein, the bispecific EGFR/c-Met antibody comprises the VH2 and the VL2, wherein
[0466] the VH2 comprises the HCDR1, the HCDR2, and the HCDR3 amino acid sequences of SEQ ID NOs: 228, 229 and 230, respectively; and
[0467] the VL2 comprises the LCDR1, the LCDR2 and the LCDR3 amino acid sequences of SEQ ID NOs: 231, 232 and 233, respectively.
[0468] In some embodiments described herein, the bispecific EGFR/c-Met antibody comprises the VH1, the VL1, the VH2 and the VL2 amino acid sequences of SEQ ID NOs: 191, 192, 195 and 196, respectively.
[0469] In some embodiments described herein, the bispecific EGFR/c-Met antibody comprises the HC1, the LC1, the HC2 and the LC2 amino acid sequences of SEQ ID NOs: 234, 235, 236 and 237, respectively, optionally having the C-terminal lysine removed from the HC1, the HC2, or both the HC1 and the HC2.
[0470] In some embodiments described herein, the bispecific EGFR/c-Met antibodies may block EGF binding to the EGFR and HGF binding to c-Met with an IC.sub.50 value of less than about 1.times.10.sup.-8 M, less than about 1.times.10.sup.-9 M, less than about 1.times.10.sup.-10 M, less than about 1.times.10.sup.-11 M, or less than about 1.times.10.sup.-12 M in a competition assay employing recombinant human EGFR or recombinant human c-Met extracellular domains coated on plates and incubated with or without the bispecific EGFR/c-Met antibodies of the invention. The bispecific EGFR/c-Met antibodies described herein may block EGF binding to EGFR and HGF binding to c-Met by at least 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% or 100% when compared to binding of EGF to the EGFR and HGF binding to c-Met in the absence of the bispecific EGFR/c-Met antibodies of the invention described herein using the same assay conditions.
[0471] In some embodiments described herein, the bispecific EGFR/c-Met antibody comprises the HC1, LC1, HC2 and LC2, wherein the HC1, the LC1, the HC2 and the LC2 are encoded by synthetic polynucleotides comprising the sequence of SEQ ID NOs: 205, 206, 207 and 208, respectively.
[0472] The bispecific EGFR/c-Met antibodies of the invention may be generated using techniques described herein, such as utilizing CH3 engineering and generating the antibodies using in vitro Fab arm exchange. An exemplary bispecific antibody may be generated from two monospecific antibodies by combining about 1-20 mg/mL of each antibody at a 1:1 molar ratio in PBS at pH 7.0-7.4 in a buffer having a final concentration of 75 mM 2-mercaptoethanolamine (2-MEA), incubating for 2-6 hours at 25-37.degree. C., followed by removal of 2-MEA via dialysis, diafiltration, tangential flow filtration, and spinned cell filtration. The yield of the bispecific antibody may be more than about 80%, more than about 90%, more than about 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% or 99%.
[0473] Some embodiments described herein provide for methods of producing the isolated bispecific EGFR/c-Met antibody, comprising:
[0474] combining an isolated monospecific bivalent anti-EGFR antibody comprising two heavy chains of SEQ ID NO: 199 and two light chains of SEQ ID NO: 200 and an isolated monospecific bivalent anti-c-Met antibody comprising two heavy chains of SEQ ID NO: 201 and two light chains of SEQ ID NO: 202 in a mixture of about 1:1 molar ratio;
[0475] introducing a reducing agent into the mixture;
[0476] incubating the mixture about ninety minutes to about six hours;
[0477] removing the reducing agent; and
[0478] purifying the bispecific EGFR/c-Met antibody that comprises a first heavy chain of SEQ ID NO: 199 and a second heavy chain of SEQ ID NO: 201, a first light chain of SEQ ID NO: 200 and a second light chain of SEQ ID NO: 202, wherein the first heavy chain of SEQ ID NO: 199 pairs with the first light chain of SEQ ID NO: 200 to form the first binding domain that specifically binds EGFR, and the second heavy chain of SEQ ID NO: 201 pairs with the second light chain of SEQ ID NO: 202 to form the second binding domain that specifically binds c-Met.
[0479] In some embodiments described herein, the reducing agent is 2-mercaptoethanolamine (2-MEA).
[0480] In some embodiments described herein, 2-MEA is present at a concentration of about 25 mM to about 75 mM.
[0481] In some embodiments described herein, the incubating step is performed at a temperature of about 25.degree. C. to about 37.degree. C.
[0482] Some embodiments described herein provide for an isolated bispecific EGFR/-c-Met antibody comprising a HC1, a LC1, a HC2 and a LC2, wherein the HC1 comprises the sequence of SEQ ID NO: 199, the LC1 comprises the sequence of SEQ ID NO: 200, the HC2 comprises the sequence of SEQ ID NO: 201, and the LC2 comprises the sequence of SEQ ID NO: 202, wherein the HC1, the LC1, the HC2 and/or the LC2 further comprises 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 or 15 conservative amino acid substitutions.
[0483] Some embodiments described herein provide for an isolated bispecific EGFR/-c-Met antibody comprising the HC1, the LC1, the HC2 and the LC2, wherein the HC1 comprises the sequence of SEQ ID NO: 234, the LC1 comprises the sequence of SEQ ID NO: 235, the HC2 comprises the sequence of SEQ ID NO: 236, and the LC2 comprises the sequence of SEQ ID NO: 237, wherein the HC1, the LC1, the HC2 and/or the LC2 further comprises 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 or 15 conservative amino acid substitutions.
[0484] Bispecific EGFR/c-Met antibodies whose HC1, LC1, HC2 and LC2 amino acid sequences differ insubstantially from those antibodies disclosed herein are encompassed within the scope of the invention. Typically, this involves one or more conservative amino acid substitutions with an amino acid having similar charge, hydrophobic, or stereochemical characteristics in the antigen-binding sites or in the frameworks without adversely altering the properties of the antibody. Conservative substitutions may also be made to improve antibody properties, for example stability or affinity. 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, or 15 amino acid substitutions may be made for example to the VH1, the VL1, the VH2 and/or the VL2. For example, a "conservative amino acid substitution" may involve a substitution of a native amino acid residue with a nonnative residue such that there is little or no effect on the polarity or charge of the amino acid residue at that position. Furthermore, any native residue in the polypeptide may also be substituted with alanine, as has been previously described for alanine scanning mutagenesis (MacLennan et al., Acta Physiol Scand Suppl 643:55-67, 1998; Sasaki et al., Adv Biophys 35:1-24, 1998). Desired amino acid substitutions may be determined by those skilled in the art at the time such substitutions are desired. For example, amino acid substitutions can be used to identify important residues of the molecule sequence, or to increase or decrease the affinity of the molecules described herein. Exemplary conservative amino acid substitutions are described supra.
[0485] Amino acid substitutions may be done for example by PCR mutagenesis (U.S. Pat. No. 4,683,195). Libraries of variants may be generated using well known methods, for example using random (NNK) or non-random codons, for example DVK codons, which encode 11 amino acids (Ala, Cys, Asp, Glu, Gly, Lys, Asn, Arg, Ser, Tyr, Trp) and screening the libraries for variants with desired properties.
[0486] In some embodiments described herein, amino acid substitutions can be made to the constant region of the antibody. For example different IgG1 allotypes can be used in the bispecific EGFR/c-Met antibodies of the invention, such as well known G1m17 allotype, G1m3 allotype or G1m1 allotype, or a combination thereof.
[0487] In some embodiments described herein, pharmacokinetic properties of the bispecific EGFR/c-Met antibodies may be enhanced by substitutions in the Fc domain that modulate antibody half-life. In some embodiments described herein, the bispecific EGFR/c-Met antibody comprises a substitution M252Y/S254T/T256E in the HC1 and/or the HC2, wherein residue numbering is according to the EU index. M252Y/S254T/T256E substitutions have been show to increase antibody half-life (Dall'Acqua et al., J Biol Chem 281:23514-24, 2006).
[0488] The bispecific EGFR/c-Met antibodies having conservative substitutions and/or additional substitutions in their Fc region are tested for their characteristics using the methods described herein.
[0489] In some embodiment described herein, immune effector properties of the bispecific EGFR/c-Met antibodies may be enhanced or silenced through Fc modifications by techniques known to those skilled in the art. For example, Fc effector functions such as C1q binding, complement dependent cytotoxicity (CDC), antibody-dependent cell-mediated cytotoxicity (ADCC), antibody-dependent cell-mediated phagocytosis (ADCP), down regulation of cell surface receptors (e.g., B cell receptor; BCR), etc. may be provided and/or controlled by modifying residues in the Fc responsible for these activities.
[0490] "Antibody-dependent cell-mediated cytotoxicity" or "ADCC" refers to a cell-mediated reaction in which non-specific cytotoxic cells that express Fc receptors (FcRs) (e.g. Natural Killer (NK) cells, neutrophils, and macrophages) recognize bound antibody on a target cell and subsequently cause lysis of the target cell.
[0491] The ability of monoclonal antibodies to induce ADCC can be enhanced by engineering their oligosaccharide component. Human IgG1 or IgG3 are N-glycosylated at Asn297 with the majority of the glycans in the well known biantennary G0, G0F, G1, G1F, G2 or G2F forms. Antibodies produced by non-engineered CHO cells typically have a glycan fucose content of about at least 85%. The removal of the core fucose from the biantennary complex-type oligosaccharides attached to the Fc regions enhances the ADCC of antibodies via improved Fc.gamma.RIIIa binding without altering antigen binding or CDC activity. Such mAbs can be achieved using different methods reported to lead to the successful expression of relatively high defucosylated antibodies bearing the biantennary complex-type of Fc oligosaccharides such as control of culture osmolality (Konno et al., Cytotechnology 64(:249-65, 2012), application of a variant CHO line Lec13 as the host cell line (Shields et al., J Biol Chem 277:26733-26740, 2002), application of a variant CHO line EB66 as the host cell line (Olivier et al., MAbs; 2(4), 2010; Epub ahead of print; PMID:20562582), application of a rat hybridoma cell line YB2/0 as the host cell line (Shinkawa et al., J Biol Chem 278:3466-3473, 2003), introduction of small interfering RNA specifically against the .alpha. 1,6-fucosyltrasferase (FUT8) gene (Mori et al., Biotechnol Bioeng 88:901-908, 2004), or coexpression of .beta.-1,4-N-acetylglucosaminyltransferase III and Golgi a-mannosidase II or a potent alpha-mannosidase I inhibitor, kifunensine (Ferrara et al., J Biol Chem 281:5032-5036, 2006, Ferrara et al., Biotechnol Bioeng 93:851-861, 2006; Xhou et al., Biotechnol Bioeng 99:652-65, 2008).
[0492] In some embodiments described herein, ADCC elicited by the bispecific EGFR/c-Met antibodies may also be enhanced by certain substitutions in the antibody Fc. Exemplary substitutions are for example substitutions at amino acid positions 256, 290, 298, 312, 356, 330, 333, 334, 360, 378 or 430 (residue numbering according to the EU index) as described in U.S. Pat. No. 6,737,056.
[0493] In some embodiments described herein, the bispecific EGFR/c-Met antibody of the invention has a biantennary glycan structure with fucose content of about between 1% to about 15%, for example 15%, 14%, 13%, 12%, 11% 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% or 1% In some embodiments, the bispecific EGFR/c-Met antibody has a glycan structure with fucose content of about 50%, 40%, 45%, 40%, 35%, 30%, 25%, or 20%.
[0494] "Fucose content" means the amount of the fucose monosaccharide within the sugar chain at Asn297. The relative amount of fucose is the percentage of fucose-containing structures related to all glycostructures. These may be characterized and quantified by multiple methods, for example: 1) using MALDI-TOF of N-glycosidase F treated sample (e.g. complex, hybrid and oligo- and high-mannose structures) as described in Int Pat. Publ. No. WO2008/077546 2); 2) by enzymatic release of the Asn297 glycans with subsequent derivatization and detection/quantitation by HPLC (UPLC) with fluorescence detection and/or HPLC-MS (UPLC-MS); 3) intact protein analysis of the native or reduced mAb, with or without treatment of the Asn297 glycans with Endo S or other enzyme that cleaves between the first and the second GlcNAc monosaccharides, leaving the fucose attached to the first GlcNAc; 4) digestion of the mAb to constituent peptides by enzymatic digestion (e.g., trypsin or endopeptidase Lys-C), and subsequent separation, detection and quantitation by HPLC-MS (UPLC-MS); 5) Separation of the mAb oligosaccharides from the mAb protein by specific enzymatic deglycosylation with PNGase F at Asn 297. The oligosaccharides thus released can be labeled with a fluorophore, separated and identified by various complementary techniques which allow: fine characterization of the glycan structures by matrix-assisted laser desorption ionization (MALDI) mass spectrometry by comparison of the experimental masses with the theoretical masses, determination of the degree of sialylation by ion exchange HPLC (GlycoSep C), separation and quantification of the oligosaccharide forms according to hydrophilicity criteria by normal-phase HPLC (GlycoSep N), and separation and quantification of the oligosaccharides by high performance capillary electrophoresis-laser induced fluorescence (HPCE-LIF).
[0495] "Low fucose" or "low fucose content" as used in the application refers to antibodies with fucose content of about between 1%-15%.
[0496] "Normal fucose" or `normal fucose content" as used herein refers to antibodies with fucose content of about over 50%, typically about over 80% or over 85%.
[0497] Some embodiments of the invention provide a synthetic nucleic acid encoding the heavy chains and the light chains of the bispecific EGFR/c-Met binding antibodies of the invention as described herein as isolated polynucleotides or as portions of expression vectors or as portions of linear DNA sequences, including linear DNA sequences used for in vitro transcription/translation, vectors compatible with prokaryotic, eukaryotic or filamentous phage expression, secretion and/or display of the compositions or directed mutagens thereof.
[0498] Some embodiments of the invention provide an isolated polynucleotide comprising the polynucleotide sequence of SEQ ID NOs: 205, 206, 207 or 208.
[0499] The polynucleotides of the invention may be produced by chemical synthesis such as solid phase polynucleotide synthesis on an automated polynucleotide synthesizer and assembled into complete single or double stranded molecules. Alternatively, the polynucleotides of the invention may be produced by other techniques such as PCR followed by routine cloning. Techniques for producing or obtaining polynucleotides of a given known sequence are well known in the art.
[0500] The polynucleotides of the invention may comprise at least one non-coding sequence, such as a promoter or enhancer sequence, intron, polyadenylation signal, a cis sequence facilitating RepA binding, and the like. The polynucleotide sequences may also comprise additional sequences encoding additional amino acids that encode for example a marker or a tag sequence such as a histidine tag or an HA tag to facilitate purification or detection of the protein, a signal sequence, a fusion protein partner such as RepA, Fc or bacteriophage coat protein such as pIX or pIII.
[0501] Some embodiments described herein provide for a vector comprising the polynucleotide of the invention. Such vectors may be plasmid vectors, viral vectors, vectors for baculovirus expression, transposon based vectors or any other vector suitable for introduction of the polynucleotide of the invention into a given organism or genetic background by any means. For example, polynucleotides encoding heavy and light chains of the bispecific antibodies of the invention may be inserted into expression vectors. The light and heavy chains may be cloned in the same or different expression vectors. The DNA segments encoding immunoglobulin chains may be operably linked to control sequences in the expression vector(s) that ensure the expression of immunoglobulin polypeptides. Such control sequences include signal sequences, promoters (e.g. naturally associated or heterologous promoters), enhancer elements, and transcription termination sequences, and may be chosen to be compatible with the host cell chosen to express the antibody. Once the vector has been incorporated into the appropriate host, the host may be maintained under conditions suitable for high level expression of the proteins encoded by the incorporated synthetic polynucleotides.
[0502] Suitable expression vectors are typically replicable in the host organisms either as episomes or as an integral part of the host chromosomal DNA. Commonly, expression vectors contain selection markers such as ampicillin-resistance, hygromycin-resistance, tetracycline resistance, kanamycin resistance or neomycin resistance to permit detection of those cells transformed with the desired DNA sequences.
[0503] Some embodiments described herein provide for a host cell comprising the vector of the invention. The term "host cell" refers to a cell into which a vector has been introduced. It is understood that the term host cell is intended to refer not only to the particular subject cell but to the progeny of such a cell. Because certain modifications may occur in succeeding generations due to either mutation or environmental influences, such progeny may not be identical to the parent cell, but are still included within the scope of the term "host cell" as used herein. Such host cells may be eukaryotic cells, prokaryotic cells, plant cells or archeal cells.
[0504] Exemplary eukaryotic cells may be of mammalian, insect, avian or other animal origins. Mammalian eukaryotic cells include immortalized cell lines such as hybridomas or myeloma cell lines such as SP2/0 (American Type Culture Collection (ATCC), Manassas, Va., CRL-1581), NSO (European Collection of Cell Cultures (ECACC), Salisbury, Wiltshire, UK, ECACC No. 85110503), FO (ATCC CRL-1646) and Ag653 (ATCC CRL-1580) murine cell lines. An exemplary human myeloma cell line is U266 (ATTC CRL-TIB-196). Other useful cell lines include those derived from Chinese Hamster Ovary (CHO) cells such as CHO-K1SV (Lonza Biologics, Walkersville, Md.), CHO-Kt (ATCC CRL-61) or DG44.
Uses of Bispecific EGFR/c-Met FN3 Domain Containing Molecules, Bispecific EGFR/-c-Met Antibodies and EGFR-Binding or c-Met Binding FN3 Domains of the Invention
[0505] The bispecific EGFR/c-Met FN3 domain containing molecules, the EGFR binding FN3 domains, the c-Met binding FN3 domains or the bispecific EGFR-c-Met antibodies of the invention may be used to diagnose, monitor, modulate, treat, alleviate, help prevent the incidence of, or reduce the symptoms of human disease or specific pathologies in cells, tissues, organs, fluid, or, generally, a host. The methods of the invention may be used to treat an animal patient belonging to any classification. Examples of such animals include mammals such as humans, rodents, dogs, cats and farm/domestic animals.
[0506] One aspect of the invention is a method for inhibiting growth or proliferation of cells that express EGFR and/or c-Met, comprising contacting the cells with the isolated bispecific EGFR/c-Met FN3 domain containing molecule, the EGFR binding FN3 domain, the c-Met binding FN3 domain or the bispecific EGFR/c-Met antibody of the invention.
[0507] Another aspect of the invention is a method for inhibiting growth or metastasis of EGFR and/or c-Met-expressing tumor or cancer cells in a subject comprising administering to the subject an effective amount of the isolated bispecific EGFR/c-Met FN3 domain containing molecule, the EGFR binding FN3 domain, the c-Met binding FN3 domain or the bispecific EGFR/c-Met antibody of the invention so that the growth or metastasis of EGFR- and/or c-Met-expressing tumor or cancer cell is inhibited.
[0508] Another aspect of the invention is a method of treating a subject having cancer, comprising administering a therapeutically effective amount of the isolated bispecific EGFR/c-Met FN3 domain containing molecule, the EGFR binding FN3 domain, the c-Met binding FN3 domain or the bispecific EGFR/c-Met antibody of the invention to a patient in need thereof for a time sufficient to treat the cancer.
[0509] The bispecific EGFR/c-Met FN3 domain containing molecule, the EGFR binding FN3 domain, the c-Met binding FN3 domain or the bispecific EGFR/c-Met antibodies of the invention may be used for treatment of any disease or disorder characterized by abnormal activation or production of EGFR, c-Met, EGF, soluble EGFR, soluble c-Met or other EGFR ligand or HGF, or disorder related to EGFR or c-Met expression, which may or may not involve malignancy or cancer, where abnormal activation and/or production of EGFR, c-Met, EGF or other EGFR ligand, or HGF is occurring in cells or tissues of a subject having, or predisposed to, the disease or disorder.
[0510] The FN3 domains that specifically bind c-Met and block binding of HGF to c-Met of the invention may be for treatment of tumors, including cancers and benign tumors. Cancers that are amenable to treatment by the c-Met binding FN3 domains of the invention include those that overexpress c-Met. Exemplary cancers that are amenable to treatment by the FN3 domains of the invention include epithelial cell cancers, breast cancer, ovarian cancer, lung cancer, colorectal cancer, anal cancer, prostate cancer, kidney cancer, bladder cancer, head and neck cancer, gastric cancer, ovarian cancer, pancreatic cancer, skin cancer, oral cancer, esophageal cancer, vaginal cancer, cervical cancer, cancer of the spleen, testicular cancer, and cancer of the thymus.
[0511] The FN3 domains that specifically bind EGFR and blocks binding of EGF to the EGFR of the invention may be used for treatment of tumors, including cancers and benign tumors. Cancers that are amenable to treatment by the FN3 domains of the invention include those that overexpress EGFR or variants. Exemplary cancers that are amenable to treatment by the FN3 domains of the invention include epithelial cell cancers, breast cancer, ovarian cancer, lung cancer, colorectal cancer, anal cancer, prostate cancer, kidney cancer, bladder cancer, head and neck cancer, ovarian cancer, pancreatic cancer, skin cancer, oral cancer, esophageal cancer, vaginal cancer, cervical cancer, cancer of the spleen, testicular cancer, and cancer of the thymus. The bispecific EGFR/c-Met FN3 domain containing molecules or the bispecific EGFR/c-Met antibodies of the invention may be used for treatment of tumors, including cancers and benign tumors. Exemplary cancers that are amenable to treatment by the bispecific EGFR/c-Met FN3 domain containing molecule or the bispecific EGFR/c-Met antibody of the invention include those that over-express EGFR and/or c-Met, cancers associated with elevated EGFR activity and/or expression levels (such as, for example, an EGFR activating mutation, an EGFR gene amplification, or ligand mediated EGFR activation) and elevated c-Met activity and/or expression levels (such as, for example, a c-Met activating mutation, a c-Met gene amplification, or HGF mediated c-Met activation).
[0512] Exemplary EGFR activating mutations that may be associated with cancer include point mutations, deletion mutations, insertion mutations, inversions or gene amplifications that lead to an increase in at least one biological activity of EGFR, such as elevated tyrosine kinase activity, formation of receptor homodimers and heterodimers, enhanced ligand binding etc. Mutations can be located in any portion of an EGFR gene or regulatory region associated with an EGFR gene and include mutations in exon 18, 19, 20 or 21 or mutations in the kinase domain. Exemplary activating EGFR mutations are G719A, L861X (X being any amino acid), L858R, E746K, L747S, E749Q, A750P, A755V, V765M, L858P or T790M substitutions, deletion of E746-A750, deletion of R748-P753, insertion of Ala between M766 and A767, insertion of SVA (Ser, Val, Ala) between S768 and V769, and insertion of NS (Asn, Ser) between P772 and H773. Other examples of EGFR activating mutations are known in the art (see e.g., U.S. Pat. Publ. No. US2005/0272083). Information about EGFR and other ErbB receptors including receptor homo- and hetero-dimers, receptor ligands, autophosphorylation sites, and signaling molecules involved in ErbB mediated signaling is known in the art (see e.g., Hynes and Lane, Nature Reviews Cancer 5: 341-354, 2005).
[0513] Exemplary c-Met activating mutations include point mutations, deletion mutations, insertion mutations, inversions or gene amplifications that lead to an increase in at least one biological activity of a c-Met protein, such as elevated tyrosine kinase activity, formation of receptor homodimers and heterodimers, enhanced ligand binding etc. Mutations can be located in any portion of the c-Met gene or regulatory regions associated with the gene, such as mutations in the kinase domain of c-Met. Exemplary c-Met activating mutations are mutations at residue positions N375, V13, V923, R175, V136, L229, S323, R988, S1058/T1010 and E168. Methods for detecting EGFR and c-Met mutations or gene amplifications are well known.
[0514] Exemplary cancers that are amenable to treatment by the bispecific molecules of the invention such as the bispecific EGFR/c-Met antibodies of the invention include epithelial cell cancers, breast cancer, ovarian cancer, lung cancer, non-small cell lung cancer (NSCLC), lung adenocarcinoma, small cell lung cancer, colorectal cancer, anal cancer, prostate cancer, kidney cancer, bladder cancer, head and neck cancer, pharynx cancer, cancer of the nose, pancreatic cancer, skin cancer, oral cancer, cancer of the tongue, esophageal cancer, vaginal cancer, cervical cancer, cancer of the spleen, testicular cancer, gastric cancer, cancer of the thymus, colon cancer, thyroid cancer, liver cancer (hepatocellular carcinoma (HCC)) or sporadic or hereditary papillary renal cell carcinoma (PRCC).
[0515] Another aspect of the invention is a method of treating a subject having cancer, comprising administering a therapeutically effective amount of the isolated bispecific EGFR/c-Met antibody of the invention to a patient in need thereof for a time sufficient to treat the cancer, wherein the subject is homozygous for phenylalanine at position 158 of CD16 (Fc.gamma.RIIIa-158F/F genotype) or heterozygous for valine and phenylalanine at position 158 of CD16 (Fc.gamma.RIIIa-158F/V genotype). CD16 is also known as the Fc gamma receptor Ma (Fc.gamma.RIIIa) or the low affinity immunoglobulin gamma Fc region receptor III-A isoform. Valine/phenylalanine (V/F) polymorphism at Fc.gamma.RIIIa protein residue position 158 has been shown to affect Fc.gamma.RIIIa affinity to human IgG. Receptor with Fc.gamma.RIIIa-158F/F or Fc.gamma.RIIIa-158F/V polymorphisms demonstrates reduced Fc engagement and therefore reduced ADCC when compared to the Fc.gamma.RIIIa-158V/V. The lack of or low amount of fucose on human N-linked oligosaccharides improves the ability of the antibodies to induce ADCC due to improved binding of the antibodies to human Fc.gamma.RIIIa (CD16) (Shields et al., J Biol Chem 277:26733-40, 2002). The antibodies of the invention have reduced fucose content of about between 1% to about 10%. In some embodiments, the bispecific EGFR/c-Met antibody has a glycan structure with fucose content of about 50%, 40%, 45%, 40%, 35%, 30%, 25%, 20%, 15%, 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% or 1%. Therefore, the antibodies of the invention may be more efficacious in the treatment of patients with Fc.gamma.RIIIa-158F/F or Fc.gamma.RIIIa-158FN genotypes. Patients can be analyzed for their Fc.gamma.RIIIa polymorphism using routine methods.
[0516] In some methods described herein, the antibodies of the invention may be used to treat a subject having cancer that is resistant or has acquired resistance to treatment with one or more EGFR inhibitors. Exemplary EGFR inhibitors for which cancer may acquire resistance are anti-EGFR antibodies cetuximab (Erbitux.RTM.), panitumumab (Vectibix.RTM.), matuzumab, nimotuzumab, small molecule EGFR inhibitors Tarceva.RTM. (erlotinib), IRESSA (gefitinib), EKB-569 (pelitinib, irreversible EGFR TKI), pan-ErbB and other receptor tyrosine kinase inhibitors, lapatinib (EGFR and HER2 inhibitor), pelitinib (EGFR and HER2 inhibitor), vandetanib (ZD6474, ZACTIMA.TM., EGFR, VEGFR2 and RET TKI), PF00299804 (dacomitinib, irreversible pan-ErbB TKI), CI-1033 (irreversible pan-erbB TKI), afatinib (BIBW2992, irreversible pan-ErbB TKI), AV-412 (dual EGFR and ErbB2 inhibitor), EXEL-7647 (EGFR, ErbB2, GEVGR and EphB4 inhibitor), CO-1686 (irreversible mutant-selective EGFR TKI), AZD9291 (irreversible mutant-selective EGFR TKI), and HKI-272 (neratinib, irreversible EGFR/ErbB2 inhibitor). The methods described herein may be used to treat cancer that is resistant to treatment with gefitinib, erlotinib, afatinib, CO-1686, AZD9291 and/or cetuximab. An exemplary antibody that can be used is EM1-mAb.
[0517] Another aspect of the invention is a method of treating a subject having cancer, comprising administering a therapeutically effective amount of the isolated bispecific EGFR/c-Met antibody of the invention to a patient in need thereof for a time sufficient to treat the cancer, wherein the subject is resistant or has acquired resistance to treatment with erlotinib, gefitinib, afatinib, CO-1686, AZD9291 or cetuximab.
[0518] Various qualitative and/or quantitative methods may be used to determine if a subject is resistant, has developed or is susceptible to developing a resistance to treatment with an EGFR inhibitor. Symptoms that may be associated with resistance to an EGFR inhibitor include, for example, a decline or plateau of the well-being of the patient, an increase in the size of a tumor, arrested or slowed decline in growth of a tumor, and/or the spread of cancerous cells in the body from one location to other organs, tissues or cells. Re-establishment or worsening of various symptoms associated with cancer may also be an indication that a subject has developed or is susceptible to developing resistance to EGFR inhibitors, such as anorexia, cognitive dysfunction, depression, dyspnea, fatigue, hormonal disturbances, neutropenia, pain, peripheral neuropathy, and sexual dysfunction. The symptoms associated with cancer may vary according to the type of cancer. For example, symptoms associated with cervical cancer may include abnormal bleeding, unusual heavy vaginal discharge, pelvic pain that is not related to the normal menstrual cycle, bladder pain or pain during urination, and bleeding between regular menstrual periods, after sexual intercourse, douching, or pelvic exam. Symptoms associated with lung cancer may include persistent cough, coughing up blood, shortness of breath, wheezing chest pain, loss of appetite, losing weight without trying and fatigue. Symptoms for liver cancer may include loss of appetite and weight, abdominal pain, especially in the upper right part of abdomen that may extend into the back and shoulder, nausea and vomiting, general weakness and fatigue, an enlarged liver, abdominal swelling (ascites), and a yellow discoloration of the skin and the whites of eyes (jaundice). One skilled in oncology may readily identify symptoms associated with a particular cancer type.
[0519] Others means to determine if a subject has developed a resistance to an EGFR inhibitor include examining EGFR phosphorylation, ERK1/2 phosphorylation and/or AKT phosphorylation in cancer cells, where increased phosphorylation may be indicative that the subject has developed or is susceptible to developing resistance to an EGFR inhibitor. Methods of determining EGFR, ERK1/2 and/or AKT phosphorylation are well known and described herein. Identification of a subject who has developed a resistance to an EGFR inhibitor may involve detection of elevated c-Met expression levels or elevated c-Met activity, for example, arising from increased levels of circulating HGF, an activating mutation of the c-Met gene or a c-Met gene amplification.
[0520] Another embodiment of the invention is a method of treating NSCLC in a patient having an NSCLC tumor or tumor metastasis having an activating EGFR mutation or EGFR gene amplification, comprising administering to the patient a therapeutically effective amount of the bispecific EGFR/c-Met antibody of the invention.
[0521] The bispecific EGFR/c-Met antibodies of the invention can be used to treat non-small cell lung cancer (NSCLC), which includes squamous cell carcinoma, adenocarcinoma, and large cell carcinoma. In some embodiments, cells of the NSCLC have an epithelial phenotype. In some embodiments, the NSCLC has acquired resistance to treatment with one or more EGFR inhibitors.
[0522] In NSCLC, specific mutations in the EGFR gene are associated with high response rates (70-80%) to EGFR tyrosine kinase inhibitors (EGFR-TKIs). A 5 amino acid deletion in exon 19 or the point mutation L858R in EGFR are associated with EGFR-TKI sensitivity (Nakata and Gotoh, Expert Opin Ther Targets 16:771-781, 2012). These mutations result in a ligand-independent activation of the EGFR kinase activity. Activating EGFR mutations occur in 10-30% of NSCLC patients and are significantly more common in East Asians, women, never smokers, and patients with adenocarcinoma histology (Janne and Johnson Clin Cancer Res 12(14 Suppl): 4416s-4420s, 2006). EGFR gene amplification is also strongly correlated with response after EGFR-TKI treatment (Cappuzzo et al., J Natl Cancer hist 97:643-55, 2005).
[0523] Although the majority of NSCLC patients with EGFR mutations initially respond to EGFR TKI therapy, virtually all acquire resistance that prevents a durable response. 50-60% of patients acquire resistance due to a second-site point mutation in the kinase domain of EGFR (T790M). Nearly 60% of all tumors that become resistant to EGFR tyrosine kinase inhibitors increase c-Met expression, amplify the c-Met gene, or increase its only known ligand, HGF (Turke et al., Cancer Cell, 17:77-88, 2010).
[0524] Another embodiments of the invention is a method of treating patient having cancer, comprising administering a therapeutically effective amount of the bispecific EGFR/c-Met antibody of the invention to a patient in need thereof for a time sufficient to treat the cancer, wherein the cancer is associated with an EGFR activating mutation, an EGFR gene amplification, increased levels of circulating HGF, a c-Met activating mutation, a c-Met gene amplification or a mutant KRAS.
[0525] In some embodiments the EGFR activating mutation is G719A, G719X (X being any amino acid), L861X (X being any amino acid), L858R, E746K, L747S, E749Q, A750P, A755V, V765M, L858P or T790M substitution, deletion of E746-A750, deletion of R748-P753, insertion of Ala (A) between M766 and A767, insertion of Ser, Val and Ala (SVA) between 5768 and V769, and insertion of Asn and Ser (NS) between P772 and H773.
[0526] Another embodiments of the invention is a method of treating patient having cancer, comprising administering a therapeutically effective amount of the bispecific EGFR/c-Met antibody of the invention to a patient in need thereof for a time sufficient to treat the cancer, wherein the cancer is associated with an EGFR mutation L858R, T790M or deletion of residues E746-A750 (del(E746, A750)), EGFR amplification or c-Met amplification.
[0527] In some embodiments, the cancer is associated with wild type EGFR and wild type c-Met.
[0528] In some embodiments, the cancer is associated with wild type EGFR and c-Met amplification.
[0529] In some embodiments, the cancer is associated with EGFR L858R and T790M mutations and wild type c-Met.
[0530] In some embodiments, the cancer is associated with EGFR deletion del (E764, A750) and wild type c-Met.
[0531] In some embodiments, the cancer is associated with EGFR deletion del(E764, A750) and c-Met amplification.
[0532] hi some embodiments, the cancer is associated with EGFR deletion del(E764, A750), EGFR amplification and c-Met amplification.
[0533] hi some embodiments, the patient has a NSCLC associated with EGFR L858R and T790M mutations and wild type c-Met.
[0534] In some embodiments, the patient has a NSCLC associated with EGFR amplification and wild type c-Met.
[0535] In some embodiments, the patient has a NSCLC associated with EGFR amplification and c-Met amplification.
[0536] In some embodiments, the patient has a NSCLC associated with EGFR deletion del(E764, A750) and wild type c-Met.
[0537] In some embodiments, the patient has a NSCLC associated with EGFR deletion del(E764, A750) and c-Met amplification.
[0538] In some embodiments, the patients are treated with the EM1-mAb of the invention. The EM1-mAb of the invention shows efficacy in in vivo tumor animal models, when the tumors are associated with L858R, T790M, del(E746, A750) EGFR, EGFR amplification, wild type c-Met and/or c-Met amplification. Amplification of EGFR or c-Met may be evaluated by standard methods, for example by determining the copy number of the EGFR or c-Met gene by southern blotting, FISH, or comparative genomic hybridization (CGH).
[0539] Another embodiments of the invention is a method of treating patient having cancer, comprising administering a therapeutically effective amount of the bispecific EGFR/c-Met antibody of the invention to a patient in need thereof for a time sufficient to treat the cancer, wherein the cancer is associated with EGFR mutations L858R, T790M or deletion of residues E746-A750 (del(E746, A750)), EGFR amplification or c-Met amplification, and mutant KRAS.
[0540] In some embodiments, the mutant KRAS has a G12V substitution. KRAS belongs to the family of RAS proto-oncogenes encoding guanosine triphosphatases (GTPases), and mediates EGFR signal transduction downstream of the receptor. Tumors with proto-oncogenic KRAS mutations such as the activating G12V or G12C mutation would therefore not be expected to be treatable by EGFR antibodies. Clinical studies with anti-EGFR antibodies cetuximab or panitumumab demonstrated that patients with KRAS-mutated colorectal tumors do not respond to these agents (Van Cutsem et al., N Eng J Med 360:1408-1417, 2009; Lievre et al., J Clin Oncol 26:374-379, 2008; Amado et al., J Clin Oncol 26:1626-1634m 2008). The bispecific EGFR/c-Met antibodies of the invention mediate KRAS mutant cell line killing via effective ADCC, and therefore, contrary to the current anti-EGFR therapies, may be efficacious in treatment of patients whose cancer is associated with KRAS activating mutations. Such exemplary antibody is the EM1-mAb.
[0541] The terms "treat" or "treatment" refers to both therapeutic treatment and prophylactic or preventative measures, wherein the object is to prevent or slow down (lessen) an undesired physiological change or disorder, such as the development or spread of cancer. For purposes of this invention, beneficial or desired clinical results include, but are not limited to, alleviation of symptoms, diminishment of extent of disease, stabilized (i.e., not worsening) state of disease, delay or slowing of disease progression, amelioration or palliation of the disease state, and remission (whether partial or total), whether detectable or undetectable. "Treatment" can also mean prolonging survival as compared to expected survival if not receiving treatment. Those in need of treatment include those already with the condition or disorder as well as those prone to have the condition or disorder or those in which the condition or disorder is to be prevented.
[0542] A "therapeutically effective amount" refers to an amount effective, at dosages and for periods of time necessary, to achieve a desired therapeutic result. A therapeutically effective amount of the bispecific EGFR/c-Met antibody of the invention may vary according to factors such as the disease state, age, sex, and weight of the individual, and the ability of the bispecific EGFR/c-Met antibody of the invention to elicit a desired response in the individual. Exemplary indicators of an effective EGFR/c-Met therapeutic that may decline or abate in association with resistance include, for example, improved well-being of the patient, decrease or shrinkage of the size of a tumor, arrested or slowed growth of a tumor, and/or absence of metastasis of cancer cells to other locations in the body.
Administration/Pharmaceutical Compositions
[0543] The invention provides for pharmaceutical compositions comprising the bispecific EGFR/c-Met antibody of the invention and a pharmaceutically acceptable carrier. For therapeutic use, the bispecific EGFR/c-Met FN3 domain containing molecules, the EGFR-binding FN3 domains, the c-Met-binding FN3 domains or the bispecific EGFR/c-Met antibodies of the invention may be prepared as pharmaceutical compositions containing an effective amount of the domain, molecule or antibody as an active ingredient in a pharmaceutically acceptable carrier. The term "carrier" refers to a diluent, adjuvant, excipient, or vehicle with which the active compound is administered. Such vehicles may be liquids, such as water and oils, including those of petroleum, animal, vegetable or synthetic origin, such as peanut oil, soybean oil, mineral oil, sesame oil and the like. For example, 0.4% saline and 0.3% glycine can be used. These solutions are sterile and generally free of particulate matter. They may be sterilized by conventional, well-known sterilization techniques (e.g., filtration). The compositions may contain pharmaceutically acceptable auxiliary substances as required to approximate physiological conditions such as pH adjusting and buffering agents, stabilizing, thickening, lubricating and coloring agents, etc. The concentration of the molecules or antibodies of the invention in such pharmaceutical formulation may vary widely, i.e., from less than about 0.5%, usually to at least about 1% to as much as 15 or 20% by weight and will be selected primarily based on required dose, fluid volumes, viscosities, etc., according to the particular mode of administration selected. Suitable vehicles and formulations, inclusive of other human proteins, e.g., human serum albumin, are described, for example, in e.g. Remington: The Science and Practice of Pharmacy, 21' Edition, Troy, D. B. ed., Lipincott Williams and Wilkins, Philadelphia, Pa. 2006, Part 5, Pharmaceutical Manufacturing pp 691-1092, See especially pp. 958-989.
[0544] The mode of administration for therapeutic use of the bispecific EGFR/c-Met FN3 domain containing molecules, the EGFR-binding FN3 domains, the c-Met-binding FN3 domains or the bispecific EGFR/c-Met antibodies of the invention may be any suitable route that delivers the agent to the host, such as parenteral administration, e.g., intradermal, intramuscular, intraperitoneal, intravenous or subcutaneous, pulmonary, transmucosal (oral, intranasal, intravaginal, rectal), using a formulation in a tablet, capsule, solution, powder, gel, particle; and contained in a syringe, an implanted device, osmotic pump, cartridge, micropump; or other means appreciated by the skilled artisan, as well known in the art. Site specific administration may be achieved by for example intrarticular, intrabronchial, intraabdominal, intracapsular, intracartilaginous, intracavitary, intracelial, intracerebellar, intracerebroventricular, intracolic, intracervical, intragastric, intrahepatic, intracardial, intraosteal, intrapelvic, intrapericardiac, intraperitoneal, intrapleural, intraprostatic, intrapulmonary, intrarectal, intrarenal, intraretinal, intraspinal, intrasynovial, intrathoracic, intrauterine, intravascular, intravesical, intralesional, vaginal, rectal, buccal, sublingual, intranasal, or transdermal delivery.
[0545] Thus, a pharmaceutical composition of the invention for intramuscular injection may be prepared to contain 1 ml sterile buffered water, and between about 1 ng to about 100 mg/kg, e.g. about 50 ng to about 30 mg/kg or more preferably, about 5 mg to about 25 mg/kg, of the bispecific EGFR/c-Met FN3 domain containing molecules, the EGFR-binding FN3 domains or the c-Met-binding FN3 domains of the invention.
[0546] The bispecific EGFR/c-Met antibodies of the invention may be administered to a patient by any suitable route, for example parentally by intravenous (IV) infusion or bolus injection, intramuscularly or subcutaneously or intraperitoneally. IV infusion can be given over as little as 15 minutes, but more often for 30 minutes, 60 minutes, 90 minutes or even 2 or 3 hours. The bispecific EGFR/c-Met antibodies of the invention may also be injected directly into the site of disease (e.g., the tumor itself). The dose given to a patient having a cancer is sufficient to alleviate or at least partially arrest the disease being treated ("therapeutically effective amount") and may be sometimes 0.1 to 10 mg/kg body weight, for example 1, 2, 3, 4, 5, 6, 7, 8, 9 or 10 mg/kg, but may even higher, for example 15, 20, 30, 40, 50, 60, 70, 80, 90 or 100 mg/kg. A fixed unit dose may also be given, for example, 50, 100, 200, 500 or 1000 mg, or the dose may be based on the patient's surface area, e.g., 400, 300, 250, 200, or 100 mg/m.sup.2. Usually between 1 and 8 doses, (e.g., 1, 2, 3, 4, 5, 6, 7 or 8) may be administered to treat cancer, but 10, 12, 20 or more doses may be given. Administration of the bispecific EGFR/c-Met antibody of the invention may be repeated after one day, two days, three days, four days, five days, six days, one week, two weeks, three weeks, one month, five weeks, six weeks, seven weeks, two months, three months, four months, five months, six months or longer. Repeated courses of treatment are also possible, as is chronic administration. The repeated administration may be at the same dose or at a different dose.
[0547] For example, a pharmaceutical composition comprising the bispecific EGFR/c-Met antibody of the invention for intravenous infusion may be made up to contain about 200 ml of sterile Ringer's solution, and about 8 mg to about 2400 mg, about 400 mg to about 1600 mg, or about 400 mg to about 800 mg of the bispecific EGFR/c-Met antibody for administration to a 80 kg patient. Methods for preparing parenterally administrable compositions are well known and are described in more detail in, for example, "Remington's Pharmaceutical Science", 15th ed., Mack Publishing Company, Easton, Pa.
[0548] The bispecific EGFR/c-Met FN3 domain containing molecules, the EGFR-binding FN3 domains, the c-Met-binding FN3 domains or the bispecific EGFR/c-Met antibodies of the invention may be lyophilized for storage and reconstituted in a suitable carrier prior to use. This technique has been shown to be effective with conventional protein preparations and well known lyophilization and reconstitution techniques can be employed.
[0549] The bispecific EGFR/c-Met FN3 domain containing molecules, the EGFR-binding FN3 domains, the c-Met-binding FN3 domains or the bispecific EGFR/c-Met antibodies of the invention may be administered in combination with a second therapeutic agent simultaneously, sequentially or separately. The second therapeutic agent may be a chemotherapeutic agent or a targeted anti-cancer therapy.
[0550] The bispecific EGFR/c-Met antibody may be administered together with any one or more of the chemotherapeutic drugs or other anti-cancer therapeutics known to those of skill in the art. Chemotherapeutic agents are chemical compounds useful in the treatment of cancer and include growth inhibitory agents or other cytotoxic agents and include alkylating agents, anti-metabolites, anti-microtubule inhibitors, topoisomerase inhibitors, receptor tyrosine kinase inhibitors, angiogenesis inhibitors and the like. Examples of chemotherapeutic agents include alkylating agents such as thiotepa and cyclosphosphamide (CYTOXAN.RTM.); alkyl sulfonates such as busulfan, improsulfan and piposulfan; aziridines such as benzodopa, carboquone, meturedopa, and uredopa; ethylenimines and methylamelamines including altretamine, triethylenemelamine, trietylenephosphoramide, triethylenethiophosphaoramide and trimethylolomelamine; nitrogen mustards such as chlorambucil, chlornaphazine, cholophosphamide, estramustine, ifosfamide, mechlorethamine, mechlorethamine oxide hydrochloride, melphalan, novembichin, phenesterine, prednimustine, trofosfamide, uracil mustard; nitrosureas such as carmustine, chlorozotocin, fotemustine, lomustine, nimustine, ranimustine; antibiotics such as aclacinomysins, actinomycin, authramycin, azaserine, bleomycins, cactinomycin, calicheamicin, carabicin, carminomycin, carzinophilin, chromomycins, dactinomycin, daunorubicin, detorubicin, 6-diazo-5-oxo-L-norleucine, doxorubicin, epirubicin, esorubicin, idarubicin, marcellomycin, mitomycins, mycophenolic acid, nogalamycin, olivomycins, peplomycin, potfiromycin, puromycin, quelamycin, rodorubicin, streptonigrin, streptozocin, tubercidin, ubenimex, zinostatin, zorubicin; anti-metabolites such as methotrexate and 5-FU; folic acid analogues such as denopterin, methotrexate, pteropterin, trimetrexate; purine analogues such as fludarabine, 6-mercaptopurine, thiamiprine, thioguanine; pyrimidine analogues such as ancitabine, azacitidine, 6-azauridine, carmofur, cytarabine, dideoxyuridine, doxifluridine, enocitabine, floxuridine; androgens such as calusterone, dromostanolone propionate, epitiostanol, mepitiostane, testolactone; anti-adrenals such as aminoglutethimide, mitotane, trilostane; folic acid replenisher such as frolinic acid; aceglatone; aldophosphamide glycoside; aminolevulinic acid; amsacrine; bestrabucil; bisantrene; edatraxate; defofamine; demecolcine; diaziquone; elfornithine; elliptinium acetate; etoglucid; gallium nitrate; hydroxyurea; lentinan; lonidamine; mitoguazone; mitoxantrone; mopidamol; nitracrine; pentostatin; phenamet; pirarubicin; podophyllinic acid; 2-ethylhydrazide; procarbazine; PSK.RTM.; razoxane; sizofiran; spirogermanium; tenuazonic acid; triaziquone; 2,2',2''-trichlorotriethylamine; urethan; vindesine; dacarbazine; mannomustine; mitobronitol; mitolactol; pipobroman; gacytosine; arabinoside ("Ara-C"); cyclophosphamide; thiotepa; members of taxoid or taxane family, such as paclitaxel (TAXOL.RTM. docetaxel (TAXOTERE.RTM.) and analogues thereof; chlorambucil; gemcitabine; 6-thioguanine; mercaptopurine; methotrexate; platinum analogues such as cisplatin and carboplatin; vinblastine; platinum; etoposide (VP-16); ifosfamide; mitomycin C; mitoxantrone; vincristine; vinorelbine; navelbine; novantrone; teniposide; daunomycin; aminopterin; xeloda; ibandronate; CPT-11; topoisomerase inhibitor RFS 2000; difluoromethylornithine (DMFO); retinoic acid; esperamicins; capecitabine; inhibitors of receptor tyrosine kinases and/or angiogenesis, including sorafenib (NEXAVAR.RTM.), sunitinib (SUTENT.RTM.), pazopanib (VOTRIENT.TM.), toceranib (PALLADIA.TM.), vandetanib (ZACTIMA.TM.), cediranib (RECENTIN.RTM.), regorafenib (BAY 73-4506), axitinib (AG013736), lestaurtinib (CEP-701), erlotinib (TARCEVA.RTM.), gefitinib (IRESSA.TM.), BIBW 2992 (TOVOK.TM.), lapatinib (TYKERB.RTM.), neratinib (HKI-272), and the like, and pharmaceutically acceptable salts, acids or derivatives of any of the above. Also included in this definition are anti-hormonal agents that act to regulate or inhibit hormone action on tumors such as anti-estrogens including for example tamoxifen, raloxifene, aromatase inhibiting 4(5)-imidazoles, 4-hydroxytamoxifen, trioxifene, keoxifene, LY 117018, onapristone, and toremifene (FARESTON.RTM.); and anti-androgens such as flutamide, nilutamide, bicalutamide, leuprolide, and goserelin; and pharmaceutically acceptable salts, acids or derivatives of any of the above. Other conventional cytotoxic chemical compounds as those disclosed in Wiemann et al., 1985, in Medical Oncology (Calabresi et aL, eds.), Chapter 10, McMillan Publishing, are also applicable to the methods of the present invention.
[0551] Exemplary agents that may be used in combination with the bispecific EGFR/c-Met FN3 domain containing molecules, the EGFR-binding FN3 domains, the c-Met-binding FN3 domains or the bispecific EGFR/c-Met antibodies of the invention include tyrosine kinase inhibitors and targeted anti-cancer therapies such as Iressa.RTM. (gefitinib) and Tarceva (erlotinib) and other antagonists of HER2, HER3, HER4 or VEGF. Exemplary HER2 antagonists include CP-724-714, HERCEPTIN.TM. (trastuzumab), OMNITARG.TM. (pertuzumab), TAK-165, lapatinib (EGFR and HER2 inhibitor), and GW-282974. Exemplary HER3 antagonists include anti-Her3 antibodies (see e.g., U.S. Pat. Publ. No. US2004/0197332). Exemplary HER4 antagonists include anti-HER4 siRNAs (see e.g., Maatta et al., Mol Biol Cell 17: 67-79, 2006. An exemplary VEGF antagonist is Bevacizumab (Avastin.TM.)
[0552] When a small molecule is used in combination with the bispecific EGFR/c-Met antibody of the invention, it is typically administered more often, preferably once a day, but 2, 3, 4 or more times per day is also possible, as is every two days, weekly or at some other interval. Small molecule drugs are often taken orally but parenteral administration is also possible, e.g., by IV infusion or bolus injection or subcutaneously or intramuscularly. Doses of small molecule drugs may typically be from 10 to 1000 mg, or about 100, 150, 200 or 250 mg.
[0553] When the bispecific EGFR/c-Met antibody of the invention is administered in combination with a second therapeutic agent, the combination may take place over any convenient timeframe. For example, the bispecific EGFR/c-Met antibody and the second therapeutic agent may be administered to a patient on the same day, and even in the same intravenous infusion. However, the bispecific EGFR/c-Met antibody and the second therapeutic agent may also be administered on alternating days or alternating weeks, fortnights or months, and so on. In some methods, the bispecific EGFR/c-Met antibody and the second therapeutic agent are administered with sufficient proximity in time that they are simultaneously present (e.g., in the serum) at detectable levels in the patient being treated. In some methods, an entire course of treatment of the bispecific EGFR/c-Met antibody consisting of a number of doses over a time period is followed or preceded by a course of treatment of the second therapeutic agent also consisting of a number of doses. In some methods, treatment with the bispecific EGFR/c-Met antibody administered second is begun if the patient has resistance or develops resistance to the second therapeutic agent administered initially. The patient may receive only a single course or multiple courses of treatment with one or both the bispecific EGFR/c-Met antibody and the second therapeutic agent. A recovery period of 1, 2 or several days or weeks may be used between administration of the bispecific EGFR/c-Met antibody and the second therapeutic agent. When a suitable treatment regimen has already been established for the second therapeutic agent, that regimen may be used in combination with the bispecific EGFR/c-Met antibody of the invention. For example, Tarceva.RTM. (erlotinib) is taken as a 100 mg or 150 mg pill once a day, and Iressa.RTM. (gefitinib) is taken as 250 mg tablet daily.
[0554] The bispecific EGFR/c-Met antibody, optionally in combination with the second therapeutic agent may be administered together with any form of radiation therapy including external beam radiation, intensity modulated radiation therapy (IMRT) and any form of radiosurgery including Gamma Knife, Cyberknife, Linac, and interstitial radiation (e.g. implanted radioactive seeds, GliaSite balloon), and/or with surgery. Combination with radiation therapy can be especially appropriate for head and neck cancer and brain tumors.
[0555] While having described the invention in general terms, the embodiments of the invention will be further disclosed in the following examples that should not be construed as limiting the scope of the claims.
Example 1. Construction of Tencon Libraries
[0556] Tencon (SEQ ID NO: 1) is an immunoglobulin-like scaffold, fibronectin type III (FN3) domain, designed from a consensus sequence of fifteen FN3 domains from human tenascin-C (Jacobs et al., Protein Engineering, Design, and Selection, 25:107-117, 2012; U.S. Pat. Publ. No. 2010/0216708). The crystal structure of Tencon shows six surface-exposed loops that connect seven beta-strands. These loops, or selected residues within each loop, can be randomized in order to construct libraries of fibronectin type III (FN3) domains that can be used to select novel molecules that bind to specific targets.
Tencon:
TABLE-US-00010 [0557] (SEQ ID NO 1) LPAPKNLVVSEVTEDSLRLSWTAPDAAFDSFLIQYQESEKVGEAINLTVP GSERSYDLTGLKPGTEYTVSIYGVKGGHRSNPLSAEFTT:
Construction of TCL1 Library
[0558] A library designed to randomize only the FG loop of Tencon (SEQ ID NO: 1), TCL1, was constructed for use with the cis-display system (Jacobs et al., Protein Engineering, Design, and Selection, 25:107-117, 2012). In this system, a single-strand DNA incorporating sequences for a Tac promoter, Tencon library coding sequence, RepA coding sequence, cis-element, and ori element is produced. Upon expression in an in vitro transcription/translation system, a complex is produced of the Tencon-RepA fusion protein bound in cis to the DNA from which it is encoded. Complexes that bind to a target molecule are then isolated and amplified by polymerase chain reaction (PCR), as described below.
[0559] Construction of the TCL1 library for use with cis-display was achieved by successive rounds of PCR to produce the final linear, double-stranded DNA molecules in two halves; the 5' fragment contains the promoter and Tencon sequences, while the 3' fragment contains the repA gene and the cis- and ori elements. These two halves are combined by restriction digest in order to produce the entire construct. The TCL1 library was designed to incorporate random amino acids only in the FG loop of Tencon, KGGHRSN (SEQ ID NO: 86). NNS codons were used in the construction of this library, resulting in the possible incorporation of all 20 amino acids and one stop codon into the FG loop. The TCL1 library contains six separate sub-libraries, each having a different randomized FG loop length, from 7 to 12 residues, in order to further increase diversity. Design of Tencon-based libraries are shown in Table 2.
TABLE-US-00011 TABLE 2 Library BC Loop Design FG Loop Design WT Tencon TAPDAAFD* KGGHRSN** TCL1 TAPDAAFD* XXXXXXX XXXXXXXX XXXXXXXXX XXXXXXXXXX XXXXXXXXXXX XXXXXXXXXXXX TCL2 ######## #####S## *TAPDAAFD: residues 22-28 of SEQ ID NO: 1; **KGGHRSN: SEQ ID NO: 86 X refers to degenerate amino acids encoded by NNS codons. #refers to the ''designed distribution of amino acids'' described in the text.
[0560] To construct the TCL1 library, successive rounds of PCR were performed to append the Tac promoter, build degeneracy into the FG loop, and add necessary restriction sites for final assembly. First, a DNA sequence containing the promoter sequence and Tencon sequence 5' of the FG loop was generated by PCR in two steps. DNA corresponding to the full Tencon gene sequence was used as a PCR template with primers POP2220 (SEQ ID NO: 2) and TC5' to FG (SEQ ID NO: 3). The resulting PCR product from this reaction was used as a template for the next round of PCR amplification with primers 130mer (SEQ ID NO: 4) and Tc5' to FG to complete the appending of the 5' and promoter sequences to Tencon. Next, diversity was introduced into the FG loop by amplifying the DNA product produced in the first step with forward primer POP2222 (SEQ ID NO: 5), and reverse primers TCF7 (SEQ ID NO: 6), TCF8 (SEQ ID NO: 7), TCF9 (SEQ ID NO: 8), TCF10 (SEQ ID NO: 9), TCF11 (SEQ ID N NO: 10), or TCF12 (SEQ ID NO: 11), which contain degenerate nucleotides. At least eight 100 .mu.L, PCR reactions were performed for each sub-library to minimize PCR cycles and maximize the diversity of the library. At least 5 .mu.g of this PCR product were gel-purified and used in a subsequent PCR step, with primers POP2222 (SEQ ID NO: 5) and POP2234 (SEQ ID NO: 12), resulting in the attachment of a 6.times.His tag and NotI restriction site to the 3' end of the Tencon sequence. This PCR reaction was carried out using only fifteen PCR cycles and at least 500 ng of template DNA. The resulting PCR product was gel-purified, digested with Nod restriction enzyme, and purified by Qiagen column.
[0561] The 3' fragment of the library is a constant DNA sequence containing elements for display, including a PspOMI restriction site, the coding region of the repA gene, and the cis- and ori elements. PCR reactions were performed using a plasmid (pCR4Blunt) (Invitrogen) containing this DNA fragment with M13 Forward and M13 Reverse primers. The resulting PCR products were digested by PspOMI overnight and gel-purified. To ligate the 5' portion of library DNA to the 3' DNA containing the repA gene, 2 pmol of 5' DNA were ligated to an equal molar amount of 3' repA DNA in the presence of NotI and PspOMI enzymes and T4 ligase. After overnight ligation at 37.degree. C., a small portion of the ligated DNA was run on a gel to check ligation efficiency. The ligated library product was split into twelve PCR amplifications and a 12-cycle PCR reaction was run with primer pair POP2250 (SEQ ID NO: 13) and DidLigRev (SEQ ID NO: 14). The DNA yield for each sub-library of TCL1 library ranged from 32-34 .mu.g.
[0562] To assess the quality of the library, a small portion of the working library was amplified with primers Tcon5new2 (SEQ ID NO: 15) and Tcon6 (SEQ ID NO: 16), and was cloned into a modified pET vector via ligase-independent cloning. The plasmid DNA was transformed into BL21-GOLD (DE3) competent cells (Stratagene) and 96 randomly picked colonies were sequenced using a T7 promoter primer. No duplicate sequences were found. Overall, approximately 70-85% of clones had a complete promoter and Tencon coding sequence without frame-shift mutation. The functional sequence rate, which excludes clones with STOP codons, was between 59% and 80%.
Construction of TCL2 Library
[0563] TCL2 library was constructed in which both the BC and the FG loops of Tencon were randomized and the distribution of amino acids at each position was strictly controlled. Table 3 shows the amino acid distribution at desired loop positions in the TCL2 library. The designed amino acid distribution had two aims. First, the library was biased toward residues that were predicted to be structurally important for Tencon folding and stability based on analysis of the Tencon crystal structure and/or from homology modeling. For example, position 29 was fixed to be only a subset of hydrophobic amino acids, as this residue was buried in the hydrophobic core of the Tencon fold. A second layer of design included biasing the amino acid distribution toward that of residues preferentially found in the heavy chain HCDR3 of antibodies, to efficiently produce high-affinity binders (Birtalan et al., J Mol Biol 377:1518-28, 2008; Olson et al., Protein Sci 16:476-84, 2007). Towards this goal, the "designed distribution" of Table 3 refers to the distribution as follows: 6% alanine, 6% arginine, 3.9% asparagine, 7.5% aspartic acid, 2.5% glutamic acid, 1.5% glutamine, 15% glycine, 2.3% histidine, 2.5% isoleucine, 5% leucine, 1.5% lysine, 2.5% phenylalanine, 4% proline, 10% serine, 4.5% threonine, 4% tryptophan, 17.3% tyrosine, and 4% valine. This distribution is devoid of methionine, cysteine, and STOP codons.
TABLE-US-00012 TABLE 3 Residue WT Position* residues Distribution in the TCL2 library 22 T designed distribution 23 A designed distribution 24 P 50% P + designed distribution 25 D designed distribution 26 A 20% A + 20% G + designed distribution 27 A designed distribution 28 F 20% F, 20% I, 20% L, 20% V, 20% Y 29 D 33% D, 33% E, 33% T 75 K designed distribution 76 G designed distribution 77 G designed distribution 78 H designed distribution 79 R designed distribution 80 S 100% S 81 N designed distribution 82 P 50% P + designed distribution *residue numbering is based on Tencon sequence of SEQ ID NO: 1
[0564] The 5' fragment of the TCL2 library contained the promoter and the coding region of Tencon (SEQ ID NO: 1), which was chemically synthesized as a library pool (Sloning Biotechnology). This pool of DNA contained at least 1.times.10.sup.11 different members. At the end of the fragment, a BsaI restriction site was included in the design for ligation to RepA.
[0565] The 3' fragment of the library was a constant DNA sequence containing elements for display including a 6.times.His tag, the coding region of the repA gene, and the cis-element. The DNA was prepared by PCR reaction using an existing DNA template (above), and primers LS1008 (SEQ ID NO: 17) and DidLigRev (SEQ ID NO: 14). To assemble the complete TCL2 library, a total of 1 .mu.g of BsaI-digested 5' Tencon library DNA was ligated to 3.5 .mu.g of the 3' fragment that was prepared by restriction digestion with the same enzyme. After overnight ligation, the DNA was purified by Qiagen column and the DNA was quantified by measuring absorbance at 260 nm. The ligated library product was amplified by a 12-cycle PCR reaction with primer pair POP2250 (SEQ ID NO: 13) and DidLigRev (SEQ ID NO: 14). A total of 72 reactions were performed, each containing 50 ng of ligated DNA products as a template. The total yield of TCL2 working library DNA was about 100 .mu.g. A small portion of the working library was sub-cloned and sequenced, as described above for library TCL1. No duplicate sequences were found. About 80% of the sequences contained complete promoter and Tencon coding sequences with no frame-shift mutations.
Construction of TCL14 Library
[0566] The top (BC, DE, and FG) and the bottom (AB, CD, and EF) loops, e.g., the reported binding surfaces in the FN3 domains are separated by the beta-strands that form the center of the FN3 structure. Alternative surfaces residing on the two "sides" of the FN3 domains having different shapes than the surfaces formed by loops only are formed at one side of the FN3 domain by two anti-parallel beta-strands, the C and the F beta-strands, and the CD and FG loops, and is herein called the C-CD-F-FG surface.
[0567] A library randomizing an alternative surface of Tencon was generated by randomizing select surface exposed residues of the C and F strands, as well as portions of the CD and FG loops as shown in FIG. 1. A Tencon variant, Tencon27 (SEQ ID NO: 99) having following substitutions when compared to Tencon (SEQ ID NO: 1) was used to generate the library; E11R L17A, N46V, E86I. A full description of the methods used to construct this library is described in US. Pat. Publ. No. US2013/0226834
Example 2: Selection of Fibronectin Type III (FN3) Domains that Bind EGFR and Inhibit EGF Binding
Library Screening
[0568] Cis-display was used to select EGFR binding domains from the TCL1 and TCL2 libraries. A recombinant human extracellular domain of EGFR fused to an IgG1 Fc (R&D Systems) was biotinylated using standard methods and used for panning (residues 25-645 of full length EGFR of SEQ ID NO: 73). For in vitro transcription and translation (ITT), 2-6 .mu.g of library DNA were incubated with 0.1 mM complete amino acids, 1X S30 premix components, and 30 .mu.L of S30 extract (Promega) in a total volume of 100 .mu.L and incubated at 30.degree. C. After 1 hour, 450 .mu.L of blocking solution (PBS pH 7.4, supplemented with 2% bovine serum albumin, 100 .mu.g/mL herring sperm DNA, and 1 mg/mL heparin) were added and the reaction was incubated on ice for 15 minutes. EGFR-Fc:EGF complexes were assembled at molar ratios of 1:1 and 10:1 EGFR to EGF by mixing recombinant human EGF (R&D Systems) with biotinylated recombinant EGFR-Fc in blocking solution for 1 hour at room temperature. For binding, 500 .mu.L of blocked ITT reactions were mixed with 100 .mu.L of EGFR-Fc:EGF complexes and incubated for 1 hour at room temperature, after which bound complexes were pulled down with magnetic neutravidin or streptavidin beads (Seradyne). Unbound library members were removed by successive washes with PBST and PBS. After washing, DNA was eluted from the bound complexes by heating to 65.degree. C. for 10 minutes, amplified by PCR, and attached to a DNA fragment encoding RepA by restriction digestion and ligation for further rounds of panning. High affinity binders were isolated by successively lowering the concentration of target EGFR-Fc during each round from 200 nM to 50 nM and increasing the washing stringency. In rounds 4 and 5, unbound and weakly bound FN3 domains were removed by washing in the presence of a 10-fold molar excess of non-biotinylated EGFR-Fc overnight in PBS.
[0569] Following panning, selected FN3 domains were amplified by PCR using oligonucleotides Tcon5new2 (SEQ ID NO: 15) and Tcon6 (SEQ ID NO: 16), subcloned into a pET vector modified to include a ligase independent cloning site, and transformed into BL21-GOLD (DE3) (Stratagene) cells for soluble expression in E. coli using standard molecular biology techniques. A gene sequence encoding a C-terminal poly-histidine tag was added to each FN3 domain to enable purification and detection. Cultures were grown to an optical density of 0.6-0.8 in 2YT medium supplemented with 100 .mu.g/mL carbenicillin in 1-mL 96-well blocks at 37.degree. C. before the addition of IPTG to 1 mM, at which point the temperature was reduced to 30.degree. C. Cells were harvested approximately 16 hours later by centrifugation and frozen at -20.degree. C. Cell lysis was achieved by incubating each pellet in 0.6 mL of BugBuster.RTM. HT lysis buffer (Novagen EMD Biosciences) with shaking at room temperature for 45 minutes.
Selection of FN3 Domains that Bind EGFR on Cells
[0570] To assess the ability of different FN3 domains to bind EGFR in a more physiological context, their ability to bind A431 cells was measured. A431 cells (American Type Culture Collection, cat. # CRL-1555) over-express EGFR with -2.times.10.sup.6 receptors per cell. Cells were plated at 5,000/well in opaque black 96-well plates and allowed to attach overnight at 37.degree. C., in a humidified 5% CO.sub.2 atmosphere. FN3 domain-expressing bacterial lysates were diluted 1,000-fold into FACS stain buffer (Becton Dickinson) and incubated for 1 hour at room temperature in triplicate plates. Lysates were removed and cells were washed 3 times with 150 .mu.L/well of FACS stain buffer. Cells were incubated with 50 .mu.L/well of anti-penta His-Alexa488 antibody conjugate (Qiagen) diluted 1:100 in FACS stain buffer for 20 minutes at room temperature. Cells were washed 3 times with 150 .mu.L/well of FACS stain buffer, after which wells were filled with 100 .mu.L of FACS stain buffer and read for fluorescence at 488 nm using an Acumen eX3 reader. Bacterial lysates containing FN3 domains were screened for their ability to bind A431 cells (1320 crude bacterial lysates for TCL1 and TCL2 libraries) and 516 positive clones were identified, where binding was .gtoreq.10-fold over the background signal. 300 lysates from the TCL14 library were screened for binding, resulting in 58 unique FN3 domain sequences that bind to EGFR.
Selection of FN3 Domains that Inhibit EGF Binding to EGFR on Cells
[0571] To better characterize the mechanism of EGFR binding, the ability of various identified FN3 domain clones to bind EGFR in an EGF-competitive manner was measured using A431 cells and run in parallel with the A431 binding assay screen. A431 cells were plated at 5,000/well in opaque black 96-well plates and allowed to attach overnight at 37.degree. C. in a humidified 5% CO.sub.2 atmosphere. Cells were incubated with 50 .mu.L/well of 1:1,000 diluted bacterial lysate for 1 hour at room temperature in triplicate plates. Biotinylated EGF (Invitrogen, cat. # E-3477) was added to each well for a final concentration of 30 ng/mL and incubated for 10 minutes at room temperature. Cells were washed 3 times with 150 .mu.L/well of FACS stain buffer. Cells were incubated with 50 .mu.L/well of streptavidin-phycoerythrin conjugate (Invitrogen) diluted 1:100 in FACS stain buffer for 20 minutes at room temperature. Cells were washed 3 times with 150 .mu.L/well of FACS stain buffer, after which wells were filled with 100 .mu.L of FACS stain buffer and read for fluorescence at 600 nm using an Acumen eX3 reader.
[0572] Bacterial lysates containing the FN3 domains were screened in the EGF competition assay described above. 1320 crude bacterial lysates from TCL1 and TCL2 libraries were screened resulting in 451 positive clones that inhibited EGF binding by >50%.
Expression and Purification of Identified FN3 Domains Binding EGFR
[0573] His-tagged FN3 domains were purified from clarified E. coli lysates with His MultiTrap.TM. HP plates (GE Healthcare) and eluted in buffer containing 20 mM sodium phosphate, 500 mM sodium chloride, and 250 mM imidazole at pH 7.4. Purified samples were exchanged into PBS pH 7.4 for analysis using PD MultiTrap.TM. G-25 plates (GE Healthcare).
Size Exclusion Chromatography Analysis
[0574] Size exclusion chromatography was used to determine the aggregation state of the FN3 domains binding EGFR. Aliquots (10 .mu.L) of each purified FN3 domain were injected onto a Superdex 75 5/150 column (GE Healthcare) at a flow rate of 0.3 mL/min in a mobile phase of PBS pH 7.4. Elution from the column was monitored by absorbance at 280 nm. FN3 domains that exhibited high levels of aggregation by SEC were excluded from further analysis.
Off-Rate of Selected EGFR-Binding FN3 Domains from EGFR-Fc
[0575] Select EGFR-binding FN3 domains were screened to identify those with slow off-rates (k.sub.off) in binding to EGFR-Fc on a ProteOn XPR-36 instrument (Bio-Rad) to facilitate selection of high affinity binders. Goat anti-human Fc IgG (R&D systems), at a concentration of 5 .mu.g/mL, was directly immobilized via amine coupling (at pH 5.0) on all 6 ligand channels in horizontal orientation on the chip with a flow rate of 30 .mu.L/min in PBS containing 0.005% Tween-20. The immobilization densities averaged about 1500 Response Units (RU) with less than 5% variation among different channels. EGFR-Fc was captured on the anti-human Fc IgG surface to a density around 600 RU in vertical ligand orientation. All tested FN3 domains were normalized to a concentration of 1 .mu.M and tested for their binding in horizontal orientation. All 6 analyte channels were used for the FN3 domains to maximize screening throughput. The dissociation phase was monitored for 10 minutes at a flow rate of 100 .mu.L/min. The inter-spot binding signals were used as references to monitor non-specific binding between analytes and the immobilized IgG surface, and were subtracted from all binding responses. The processed binding data were locally fit to a 1:1 simple Langmuir binding model to extract the k.sub.off for each FN3 domain binding to captured EGFR-Fc.
Inhibition of EGF-Stimulated EGFR Phosphorylation
[0576] Purified EGFR-binding FN3 domains were tested for their ability to inhibit EGF-stimulated phosphorylation of EGFR in A431 cells at a single concentration. EGFR phosphorylation was monitored using the EGFR phospho(Tyr1173) kit (Meso Scale Discovery). Cells were plated at 20,000/well in clear 96-well tissue culture-treated plates (Nunc) in 100 .mu.L/well of RPMI medium (Gibco) containing GlutaMAX.TM. with 10% fetal bovine serum (FBS) (Gibco) and allowed to attach overnight at 37.degree. C. in a humidified 5% CO.sub.2 atmosphere. Culture medium was removed completely and cells were starved overnight in 100 .mu.L/well of medium containing no FBS at 37.degree. C. in a humidified 5% CO.sub.2 atmosphere. Cells were then treated with 100 .mu.L/well of pre-warmed (37.degree. C.) starvation medium containing EGFR-binding FN3 domains at a concentration of 2 .mu.M for 1 hour at 37.degree. C. in a humidified 5% CO.sub.2 atmosphere. Controls were treated with starvation medium only. Cells were stimulated by the addition and gentle mixing of 100 .mu.L/well of pre-warmed (37.degree. C.) starvation medium containing 100 ng/mL recombinant human EGF (R&D Systems, cat. #236-EG), for final concentrations of 50 ng/mL EGF and 1 .mu.M EGFR-binding FN3 domain, and incubation at 37.degree. C., 5% CO.sub.2 for 15 minutes. One set of control wells was left un-stimulated as negative controls. Medium was completely removed and cells were lysed with 100 .mu.L/well of Complete Lysis Buffer (Meso Scale Discovery) for 10 minutes at room temperature with shaking, as per the manufacturer's instructions. Assay plates configured for measuring EGFR phosphorylated on tyrosine 1173 (Meso Scale Discovery) were blocked with the provided blocking solution as per the manufacturer's instructions at room temperature for 1.5-2 hours. Plates were then washed 4 times with 200 .mu.L/well of 1.times. Tris Wash Buffer (Meso Scale Discovery). Aliquots of cell lysate (30 .mu.L/well) were transferred to assay plates, which were covered with plate sealing film (VWR) and incubated at room temperature with shaking for 1 hour. Assay plates were washed 4 times with 200 .mu.L/well of Tris Wash Buffer, after which 25 .mu.L of ice-cold Detection Antibody Solution (Meso Scale Discovery) were added to each well, being careful not to introduce bubbles. Plates were incubated at room temperature with shaking for 1 hour, followed by 4 washes with 200 .mu.L/well of Tris Wash Buffer. Signals were detected by addition of 150 .mu.L/well of Read Buffer (Meso Scale Discovery) and reading on a SECTOR.RTM. Imager 6000 instrument (Meso Scale Discovery) using manufacturer-installed assay-specific default settings. Percent inhibition of the EGF-stimulated positive control signal was calculated for each EGFR-binding FN3 domain.
[0577] Inhibition of EGF-stimulated EGFR phosphorylation was measured for 232 identified clones from the TCL1 and TCL2 libraries. 22 of these clones inhibited EGFR phosphorylation by .gtoreq.50% at 1 .mu.M concentration. After removal of clones that either expressed poorly or were judged to be multimeric by size exclusion chromatography, nine clones were carried forward for further biological characterization. The BC and FG loop sequences of these clones are shown in Table 4. Eight of the nine selected clones had a common FG loop sequence (HNVYKDTNMRGL; SEQ ID NO: 95) and areas of significant similarity were seen between several clones in their BC loop sequences.
TABLE-US-00013 TABLE 4 FN3 Domain BC Loop FG Loop SEQ ID SEQ ID SEQ ID Clone ID NO: Sequence NO: Sequence NO: P53A1R5-17 18 ADPHGFYD 87 HNVYKDTNMRGL 95 P54AR4-17 19 TYDRDGYD 88 HNVYKDTNMRGL 95 P54AR4-47 20 WDPFSFYD 89 HNVYKDTNMRGL 95 P54AR4-48 21 DDPRGFYE 90 HNVYKDTNMRGL 95 P54AR4-73 22 TWPYADLD 91 HNVYKDTNMRGL 95 P54AR4-74 23 GYNGDHFD 92 HNVYKDTNMRGL 95 P54AR4-81 24 DYDLGVYD 93 HNVYKDTNMRGL 95 P54AR4-83 25 DDPWDFYE 94 HNVYKDTNMRGL 95 P54CR4-31 26 TAPDAAFD 85 LGSYVFEHDVM 96
Example 3: Characterization of EGFR-Binding FN3 Domains that Inhibit EGF Binding
Large-Scale Expression, Purification, and Endotoxin Removal
[0578] The FN3 domains shown in Table 4 were scaled up to provide more material for detailed characterization. An overnight culture containing each EGFR-binding FN3 domain variant was used to inoculate 0.8 L of Terrific broth medium supplemented with 100 .mu.g/mL ampicillin at a 1/80 dilution of overnight culture into fresh medium, and incubated with shaking at 37.degree. C. The culture was induced when the optical density at 600 nm reached .about.1.2-1.5 by addition of IPTG to a final concentration of 1 mM and the temperature was reduced to 30.degree. C. After 4 hours, cells were collected by centrifugation and the cell pellet was stored at -80.degree. C. until needed.
[0579] For cell lysis, the thawed pellet was resuspended in 1X BugBuster.RTM. supplemented with 25 U/mL Benzonase.RTM. (Sigma-Aldrich) and 1 kU/mL rLysozyme.TM. (Novagen EMD Biosciences) at a ratio of 5 mL of BugBuster.RTM. per gram of pellet. Lysis proceeded for 1 hour at room temperature with gentle agitation, followed by centrifugation at 56,000.times.g for 50 minutes at 4.degree. C. The supernatant was collected and filtered through a 0.2 .mu.m filter, then loaded on to a 5-mL HisTrap FF column pre-equilibrated with Buffer A (50 mM Tris-HC1 pH 7.5, 500 mM NaCl, 10 mM imidazole) using a GE Healthcare AKTAexplorer 100s chromatography system. The column was washed with 20 column volumes of Buffer A and further washed with 16% Buffer B (50 mM Tris-HC1 pH7.5, 500 mM NaCl, 250 mM imidazole) for 6 column volumes. The FN3 domains were eluted with 50% B for 10 column volumes, followed by a gradient from 50-100% B over 6 column volumes. Fractions containing the FN3 domain protein were pooled, concentrated using a Millipore 10K MWCO concentrator, and filtered before loading onto a HiLoad.TM. 16/60 Superdex.TM. 75 column (GE Healthcare) pre-equilibrated with PBS. The protein monomer peak eluting from the size exclusion column was retained.
[0580] Endotoxins were removed using a batch approach with ActiClean Etox resin (Sterogene Bioseparations). Prior to endotoxin removal, the resin was pre-treated with 1 N NaOH for 2 hours at 37.degree. C. (or overnight at 4.degree. C.) and washed extensively with PBS until the pH had stabilized to .about.7 as measured with pH indicator paper. The purified protein was filtered through a 0.2 .mu.m filter before adding to 1 mL of Etox resin at a ratio of 10 mL of protein to 1 mL of resin. The binding of endotoxin to resin was allowed to proceed at room temperature for at least 2 hours with gentle rotation. The resin was removed by centrifugation at 500.times.g for 2 minutes and the protein supernatant was retained. Endotoxin levels were measured using EndoSafe.RTM.-PTS.TM. cartridges and analyzed on an EndoSafe.RTM.-MCS reader (Charles River). If endotoxin levels were above 5 EU/mg after the first Etox treatment, the above procedure was repeated until endotoxin levels were decreased to .gtoreq.5 EU/mg. In cases where the endotoxin level was above 5 EU/mg and stabilized after two consecutive treatments with Etox, anion exchange or hydrophobic interaction chromatography conditions were established for the protein to remove the remaining endotoxins.
Affinity Determination of Selected EGFR-Binding FN3 Domains to EGFR-Fc (EGFR-Fc Affinity)
[0581] Binding affinity of selected EGFR-binding FN3 domains to recombinant EGFR extracellular domain was further characterized by surface Plasmon resonance methods using a Proteon Instrument (BioRad). The assay set-up (chip preparation, EGFR-Fc capture) was similar to that described above for off-rate analysis. Selected EGFR binding FN3 domains were tested at 1 .mu.M concentration in 3-fold dilution series in the horizontal orientation. A buffer sample was also injected to monitor the baseline stability. The dissociation phase for all concentrations of each EGFR-binding FN3 domain was monitored at a flow rate of 100 .mu.L/min for 30 minutes (for those with k.sub.off .about.10.sup.-2 s.sup.-1 from off-rate screening), or 1 hour (for those with k.sub.off .about.10.sup.-3 s.sup.-1 or slower). Two sets of reference data were subtracted from the response data: 1) the inter-spot signals to correct for the non-specific interactions between the EGFR-binding FN3 domain and the immobilized IgG surface; 2) the buffer channel signals to correct for baseline drifting due to the dissociation of captured EGFR-Fc surface over time. The processed binding data at all concentrations for each FN3 domain were globally fit to a 1:1 simple Langmuir binding model to extract estimates of the kinetic (k.sub.on, k.sub.off) and affinity (K.sub.D) constants. Table 5 shows the kinetic constants for each of the constructs, with the affinity varying from 200 pM to 9.6 nM.
Binding of Selected EGFR-Binding FN3 Domains to EGFR on Cells ("A431 Cell Binding Assay")
[0582] A431 cells were plated at 5,000/well in opaque black 96-well plates and allowed to attach overnight at 37.degree. C., in a humidified 5% CO.sub.2 atmosphere. Purified EGFR-binding FN3 domains (1.5 nM to 30 .mu.M) were added to the cells (in 50 uL) for 1 hour at room temperature in triplicate plates. Supernatant was removed and cells were washed 3 times with 150 .mu.L/well of FACS stain buffer. Cells were incubated with 50 .mu.L/well of anti-penta His-Alexa488 antibody conjugate (Qiagen) diluted 1:100 in FACS stain buffer for 20 minutes at room temperature. Cells were washed 3 times with 150 .mu.L/well of FACS stain buffer, after which wells were filled with 100 .mu.L of FACS stain buffer and read for fluorescence at 488 nm using an Acumen eX3 reader. Data were plotted as raw fluorescence signal against the logarithm of the FN3 domain molar concentration and fitted to a sigmoidal dose-response curve with variable slope using GraphPad Prism 4 (GraphPad Software) to calculate EC.sub.50 values. Table 5 reports the EC.sub.50 for each of the constructs ranging from 2.2 nM to >20 .mu.M.
Inhibition of EGF Binding to EGFR on Cells Using Selected EGFR-Binding FN3 Domains (A431 Cell EGF Competition Assay)
[0583] A431 cells were plated at 5,000/well in opaque black 96-well plates and allowed to attach overnight at 37.degree. C., in a humidified 5% CO.sub.2 atmosphere. Purified EGFR-binding FN3 domains (1.5 nM to 30 .mu.M) were added to the cells (50 .mu.L/well) for 1 hour at room temperature in triplicate plates. Biotinylated EGF (Invitrogen, Cat #: E-3477) was added to each well to give a final concentration of 30 ng/mL and incubated for 10 minutes at room temperature. Cells were washed 3 times with 150 .mu.L/well of FACS stain buffer. Cells were incubated with 50 .mu.L/well of streptavidin-phycoerythrin conjugate (Invitrogen) diluted 1:100 in FACS stain buffer for 20 minutes at room temperature. Cells were washed 3 times with 150 .mu.L/well of FACS stain buffer, after which wells were filled with 100 .mu.L, of FACS stain buffer and read for fluorescence at 600 nm using an Acumen eX3 reader. Data were plotted as the raw fluorescence signal against the logarithm of FN3 domain molar concentration and fitted to a sigmoidal dose-response curve with variable slope using GraphPad Prism 4 (GraphPad Software) to calculate IC.sub.50 values. Table 5 reports the IC.sub.50 values ranging from 1.8 nM to 121 nM.
Inhibition of EGF-Stimulated EGFR Phosphorylation (Phoshpo EGFR Assay)
[0584] Select FN3 domains that significantly inhibited EGF-stimulated EGFR phosphorylation were assessed more completely by measuring IC.sub.50 values for inhibition. Inhibition of EGF-stimulated EGFR phosphorylation was assessed at varying FN3 domain concentrations (0.5 nM to 10 .mu.M) as described above in "inhibition of EGF stimulated EGFR phosphorylation". Data were plotted as electrochemiluminescence signal against the logarithm of the FN3 domain molar concentration and IC.sub.50 values were determined by fitting data to a sigmoidal dose response with variable slope using GraphPad Prism 4 (GraphPad Software). Table 5 shows the IC.sub.50 values which ranged from 18 nM to >2.5 .mu.m.
Inhibition of Human Tumor Cell Growth (NCI-H292 Growth and NCI-H322 Growth Assay)
[0585] Inhibition of EGFR-dependent cell growth was assessed by measuring viability of the EGFR over-expressing human tumor cell lines, NCI-H292 and NCI-H322 (American Type Culture Collection, cat. # CRL-1848 & # CRL-5806, respectively), following exposure to EGFR-binding FN3 domains. Cells were plated at 500 cells/well (NCI-H292) or 1,000 cells/well (NCI-H322) in opaque white 96-well tissue culture-treated plates (Nunc) in 100 .mu.L/well of RPMI medium (Gibco) containing GlutaMAX.TM. and 10 mM HEPES, supplemented with 10% heat inactivated fetal bovine serum (Gibco) and 1% penicillin/streptomycin (Gibco), and allowed to attach overnight at 37.degree. C. in a humidified 5% CO.sub.2 atmosphere. Cells were treated by addition of 5 .mu.L/well of phosphate-buffered saline (PBS) containing a concentration range of EGFR-binding FN3 domains. Controls were treated with 54/well of PBS only or 25 mM ethylenediaminetetraacetic acid in PBS. Cells were incubated at 37.degree. C., 5% CO.sub.2 for 120 hours. Viable cells were detected by addition of 75 .mu.L/well of CellTiter-Glo.RTM. reagent (Promega), followed by mixing on a plate shaker for 2 minutes, and incubation in the dark at room temperature for a further 10 minutes. Plates were read on a SpectraMax M5 plate reader (Molecular Devices) set to luminescence mode, with a read time of 0.5 seconds/well against a blank of medium only. Data were plotted as a percentage of PBS-treated cell growth against the logarithm of FN3 domain molar concentration. IC.sub.50 values were determined by fitting data to the equation for a sigmoidal dose response with variable slope using GraphPad Prism 4 (GraphPad Software). Table 5 shows IC.sub.50 values ranging from 5.9 nM to 1.15 .mu.M and 9.2 nM to >3.1 .mu.M, using the NCI-H292 and NCI-H322 cells respectively. Table 5 shows the summary of biological properties of EGFR-binding FN3 domains for each assay.
TABLE-US-00014 TABLE 5 A431 NCI- NCI- EGFR- Cell A431 Phospho- H292 H322 FN3 SEQ Fc Binding Cell EGF EGFR Growth Growth Domain ID Affinity EC.sub.50 Competition IC.sub.50 IC.sub.50 IC.sub.50 Clone ID NO: (nM) (nM) IC.sub.50 (nM) (nM) (nM) (nM) P53A1R5-17 18 1.89 4.0 9.8 >2500 86 65 P54AR4-17 19 9.62 16 21 184 ND ND P54AR4-47 20 2.51 8.6 7.1 295 44 39 P54AR4-48 21 7.78 12 9.8 170 ND ND P54AR4-73 22 0.197 9.4 4.6 141 83 73 P54AR4-74 23 ND 77 ND ND ND ND P54AR4-81 24 ND 84 121 ND ND ND P54AR4-83 25 0.255 2.2 1.8 18 5.9 9.2 P54CR4-31 26 0.383 >20000 55 179 1150 >3073
Example 4: Engineering of EGFR-Binding FN3 Domains
[0586] A subset of the EGFR binding FN3 domains was engineered to increase the conformational stability of each molecule. The mutations L17A, N46V, and E861 which have been shown to improve FN3 domain stability (described in US Pat. Publ. No. US2011/0274623) were incorporated into clones P54AR4-83, P54CR4-31, and P54AR4-37 by DNA synthesis. The new mutants, P54AR5-83v2, P54CR431-v2, and P54AR4-37v2 were expressed and purified as described above. Differential scanning calorimetry in PBS was used to assess the stability of each mutant in order to compare it to that of the corresponding parent molecule. Table 6 shows that each variant molecule was stabilized significantly, with an average increase in the T.sub.m of 18.5.degree. C.
TABLE-US-00015 TABLE 6 FN3 domain Clone SEQID NO: T.sub.m (.degree. C.) P54AR4-83 25 50.6 P54AR4-83v2 27 69.8 P54CR4-31 26 60.9 P54CR4-31v2 28 78.9 P54AR4-37 22 45.9 P54AR4-37v2 29 64.2
Example 5: Selection of Fibronectin Type III (FN3) Domains that Bind c-Met and Inhibit HGF Binding
[0587] Panning on Human c-Met
[0588] The TCL14 library was screened against biotinylated-human c-Met extracellular domain (bt-c-Met) to identify FN3 domains capable of specifically binding c-Met. For selections, 3 .mu.g of TCL14 library was in vitro transcribed and translated (IVTT) in E. Coli S30 Linear Extract (Promega, Madison, Wis.) and the expressed library blocked with Cis Block (2% BSA (Sigma-Aldrich, St. Louis, Mo.), 100 .mu.g/ml Herring Sperm DNA (Promega), 1 mg/mL heparin (Sigma-Aldrich)). For selections, bt-c-Met was added at concentrations of 400 nM (Round 1), 200 nM (Rounds 2 and 3) and 100 nM (Rounds 4 and 5). Bound library members were recovered using neutravidin magnetic beads (Thermo Fisher, Rockford, Ill.) (Rounds 1, 3, and 5) or streptavidin magnetic beads (Promega) (Rounds 2 and 4) and unbound library members were removed by washing the beads 5-14 times with 500 uL PBS-T followed by 2 washes with 500 .mu.L PBS.
[0589] Additional selection rounds were performed to identify FN3 domains molecules with improved affinities. Briefly, outputs from round 5 were prepared as described above and subjected to additional iterative rounds of selection with the following changes: incubation with bt-c-Met was decreased from 1 hour to 15 minutes and bead capture was decreased from 20 minutes to 15 minutes, bt-c-Met decreased to 25 nM (Rounds 6 and 7) or 2.5 nM (Rounds 8 and 9), and an additional 1 hour wash was performed in the presence of an excess of non-biotinylated c-Met. The goal of these changes was to simultaneously select for binders with a potentially faster on-rate and a slower off-rate yielding a substantially lower K.sub.D.
[0590] Rounds 5, 7 and 9 outputs were PCR cloned into a modified pET15 vector (EMD Biosciences, Gibbstown, N.J.) containing a ligase independent cloning site (pET15-LIC) using TCON6 (SEQ ID No. 30) and TCONS E86I short (SEQ ID No. 31) primers, and the proteins were expressed as C-terminal His6-tagged proteins after transformations and IPTG induction (1 mM final, 30.degree. C. for 16 hours) using standard protocols. The cells were harvested by centrifugation and subsequently lysed with Bugbuster HT (EMD Biosciences) supplemented with 0.2 mg/mL Chicken Egg White Lysozyme (Sigma-Aldrich). The bacterial lysates were clarified by centrifugation and the supernatants were transferred to new 96 deep-well plates.
Screening for FN3 Domains that Inhibit HGF Binding to c-Met
[0591] FN3 domains present in E. coli lysates were screened for their ability to inhibit HGF binding to purified c-Met extracellular domain in a biochemical format. Recombinant human c-Met Fc chimera (0.5 .mu.g/mL in PBS, 100 .mu.L/well) was coated on 96-well White Maxisorp Plates (Nunc) and incubated overnight at 4.degree. C. The plates were washed two times with 300 .mu.l/well of Tris-buffered saline with 0.05% Tween 20 (TBS-T, Sigma-Aldrich) on a Biotek plate washer. Assay plates were blocked with StartingBlock T20 (200 .mu.L/well, Thermo Fisher Scientific, Rockland, Ill.) for 1 hour at room temperature (RT) with shaking and again washed twice with 300 .mu.l of TBS-T. FN3 domain lysates were diluted in StartingBlock T20 (from 1:10 to 1:100,000) using the Hamilton STARplus robotics system. Lysates (50 .mu.L/well) were incubated on assay plates for 1 hour at RT with shaking. Without washing the plates, bt-HGF (1 .mu.g/mL in StartingBlock T20, 50 .mu.L/well, biotinylated) was added to the plate for 30 min at RT while shaking. Control wells containing Tencon27 lysates received either Starting Block T20 or diluted bt-HGF. Plates were then washed four times with 300 .mu.l/well of TBS-T and incubated with 100 .mu.l/well of Streptavidin-HRP (1:2000 in TBS-T, Jackson Immunoresearch, West Grove, Pa.) for 30-40 minutes at RT with shaking. Again the plates were washed four times with TBS-T. To develop signal, POD Chemiluminescence Substrate (50 .mu.L/well, Roche Diagnostics, Indianapolis, Ind.), prepared according to manufacturer's instructions, was added to the plate and within approximately 3 minutes luminescence was read on the Molecular Devices MS using SoftMax Pro. Percent inhibition was determined using the following calculation: 100-((RLU.sub.sample-Mean RLU.sub.NO bt-HGF control)/(Mean RLU.sub.bt-HGF control-Mean RLU.sub.No bt-HGF control)*100). Percent inhibition values of 50% or greater were considered hits.
High-Throughput Expression and Purification of FN3 Domains
[0592] His-tagged FN3 domains were purified from clarified E. coli lysates with His MultiTrap.TM. HP plates (GE Healthcare) and eluted in buffer containing 20 mM sodium phosphate, 500 mM sodium chloride, and 250 mM imidazole at pH 7.4. Purified samples were exchanged into PBS pH 7.4 for analysis using PD MultiTrap.TM. G-25 plates (GE Healthcare).
IC.sub.50 Determination of Inhibition of HGF Binding to c-Met
[0593] Select FN3 domains were further characterized in the HGF competition assay. Dose response curves for purified FN3 domains were generated utilizing the assay described above (starting concentrations of 5 .mu.M). Percent inhibition values were calculated. The data were plotted as % inhibition against the logarithm of FN3 domain molar concentrations and IC.sub.50 values were determined by fitting data to a sigmoidal dose response with variable slope using GraphPad Prism 4.
[0594] 35 unique sequences were identified from Round 5 to exhibit activity at dilutions of 1:10, with IC.sub.50 values ranging from 0.5 to 1500 nM. Round 7 yielded 39 unique sequences with activity at dilutions of 1:100 and IC.sub.50 values ranging from 0.16 to 2.9 nM. 66 unique sequences were identified from Round 9, where hits were defined as being active at dilutions of 1:1000. IC.sub.50 values as low as 0.2 nM were observed in Round 9 (Table 8).
Affinity Determination of Selected c-Met-Binding FN3 Domains to c-Met-Fc (EGFR-Fc Affinity)
[0595] Affinities were determined for select c-Met binding FN3 domains as is described in Example 3 for affinity determination fo selected EGFR-binding FN3 domains, except that c-Met-Fc was used in the assays.
Example 6: Characterization of FN3 Domains that Bind c-Met and Inhibit HGF Binding
[0596] FN3 domains were expressed and purified as described above in Example 2. Size exclusion chromatography and kinetic analysis was done as described above in Examples 1 and 2, respectively. Table 7 shows the sequences of the C-strand, CD loop, F-strand, and FG loop, and a SEQ ID NO: for the entire amino acid sequence for each domain.
TABLE-US-00016 TABLE 7 Clone SEQ ID CD FG Name NO: C loop strand F loop strand P114AR5P74-A5 32 FDSFWIRYDE VVVGGE FEYYVNILGV KGGSISV P114AR5P75-E9 33 FDSFFIRYDE FLRSGE ILYWVTILGV KGGLVST P114AR7P92-F3 34 FDSFWIRYFE FLGSGE ILYIVNIMGV KGGSISH P114AR7P92-F6 35 FDSFWIRYFE FLGSGE ILYVVNILGV KGGGLSV P114AR7P92-G8 36 FDSFVIRYFE FLGSGE ILYVVQILGV KGGYISI P114AR7P92-H5 37 FDSFWIRYLE FLLGGE ILYVVQIMGV KGGTVSP P114AR7P93-D11 38 FDSFWIRYFE FLGSGE ILYVVGINGV KGGYISY P114AR7P93-G8 39 FDSFWIRYFE FLGSGE ILYGVTINGV KGGRVST P114AR7P93-H9 40 FDSFWIRYFE FLGSGE ILYVVQIIGV KGGHISL P114AR7P94-A3 41 FDSFWIRYFE FLGSGE ILYVVNIMGV KGGKISP P114AR7P94-E5 42 FDSFWIRYFE FLGSGE ILYAVNIMGV KGGRVSV P114AR7P95-B9 43 FDSFWIRYFE FLGSGE ILYVVQILGV KGGSISV P114AR7P95-D3 44 FDSFWIRYFE FLGSGE ILYVVNIMGV KGGSISY P114AR7P95-D4 45 FDSFWIRYFE FLGSGE ILYVVQILGV KGGYISI P114AR7P95-E3 46 FDSFWIRYFE FLGSGE ILYVVQIMGV KGGTVSP P114AR7P95-F10 47 FDSFWIRYFE FTTAGE ILYVVNIMGV KGGSISP P114AR7P95-G7 48 FDSFWIRYFE LLSTGE ILYVVNIMGV KGGSISP P114AR7P95-H8 49 FDSFWIRYFE FVSKGE ILYVVNIMGV KGGSISP C loop residues correspond to residues 28-37 of indicated SEQ ID NO CD strand residues correspond to residues 38-43 of indicated SEQ ID NO F loop residues correspond to residues 65-74 of indicated SEQ ID NO FG strand residues correspond to residues 75-81 of indicated SEQ ID NO
Binding of Selected c-Met-Binding FN3 Domains to c-Met on Cells ("H441 Cell Binding Assay")
[0597] NCI-H441 cells (Cat # HTB-174, American Type Culture Collection, Manassas, Va.) were plated at 20,000 cells per well in Poly-D-lysine coated black clear bottom 96-well plates (BD Biosciences, San Jose, Calif.) and allowed to attach overnight at 37.degree. C., 5% CO.sub.2. Purified FN3 domains (50 .mu.L/well; 0 to 1000 nM) were added to the cells for 1 hour at 4.degree. C. in duplicate plates. Supernatant was removed and cells were washed three times with FACS stain buffer (150 .mu.L/well, BD Biosciences, cat #554657). Cells were incubated with biotinylated-anti HIS antibody (diluted 1:160 in FACS stain buffer, 50 .mu.L/well, R&D Systems, cat # BAM050) for 30 minutes at 4.degree. C. Cells were washed three times with FACS stain buffer (150 .mu.L/well), after which wells were incubated with anti-mouse IgG1-Alexa 488 conjugated antibody (diluted 1:80 in FACS stain buffer, 50 .mu.L/well, Life Technologies, cat # A21121) for 30 minutes at 4.degree. C. Cells were washed three times with FACS stain buffer (150 .mu.L/well) and left in FACS stain buffer (50 .mu.L/well). Total fluorescence was determined using an Acumen eX3 reader. Data were plotted as raw fluorescence signal against the logarithm of the FN3 domain molar concentration and fitted to a sigmoidal dose-response curve with variable slope using GraphPad Prism 4 (GraphPad Software) to calculate EC.sub.50 values. FN3 domains were found to exhibit a range of binding activities, with EC.sub.50 values between 1.4 nM and 22.0 nM, as shown in Table 8.
Inhibition of HGF-Stimulated c-Met Phosphorylation
[0598] Purified FN3 domains were tested for their ability to inhibit HGF-stimulated phosphorylation of c-Met in NCI-H441, using the c-Met phospho(Tyr1349) kit from Meso Scale Discovery (Gaithersburg, Md.). Cells were plated at 20,000/well in clear 96-well tissue culture-treated plates in 100 .mu.L/well of RPMI medium (containing Glutamax and HEPES, Life Technologies) with 10% fetal bovine serum (FBS; Life Technologies) and allowed to attach overnight at 37.degree. C., 5% CO.sub.2. Culture medium was removed completely and cells were starved overnight in serum-free RPMI medium (100 .mu.L/well) at 37.degree. C., 5% CO.sub.2. Cells were then replenished with fresh serum-free RPMI medium (100 .mu.L/well) containing FN3 domains at a concentration of 20 .mu.M and below for 1 hour at 37.degree. C., 5% CO.sub.2. Controls were treated with medium only. Cells were stimulated with 100 ng/mL recombinant human HGF (100 .mu.L/well, R&D Systems cat #294-HGN) and incubated at 37.degree. C., 5% CO.sub.2 for 15 minutes. One set of control wells was left un-stimulated as negative controls. Medium was then completely removed and cells were lysed with Complete Lysis Buffer (50 .mu.L/well, Meso Scale Discovery) for 10 minutes at RT with shaking, as per manufacturer's instructions. Assay plates configured for measuring phosphorylated c-Met were blocked with the provided blocking solution as per the manufacturer's instructions at room temperature for 1 hour. Plates were then washed three times with Tris Wash Buffer (200 .mu.L/well, Meso Scale Discovery). Cell lysates (30 .mu.L/well) were transferred to assay plates, and incubated at RT with shaking for 1 hour. Assay plates were then washed four times with Tris Wash Buffer, after which ice-cold Detection Antibody Solution (25 .mu.L/well, Meso Scale Discovery) was added to each well for lhr at RT with shaking. Plates were again rinsed four times with Tris Wash Buffer. Signals were detected by addition of 150 Read Buffer (150 .mu.L/well, Meso Scale Discovery) and reading on a SECTOR.RTM. Imager 6000 instrument (Meso Scale Discovery) using manufacturer-installed assay-specific default settings. Data were plotted as electrochemiluminescence signal against the logarithm of FN3 domain molar concentration and IC.sub.50 values were determined by fitting data to a sigmoidal dose response with variable slope using GraphPad Prism 4. FN3 domains were found to inhibit phosphorylated c-Met with IC.sub.50 values ranging from 4.6 nM to 1415 nM as shown in Table 8.
Inhibition of Human Tumor Cell Growth or Viability
[0599] Inhibition of c-Met-dependent cell growth was assessed by measuring viability of U87-MG cells (American Type Culture Collection, cat # HTB-14), following exposure to c-Met-binding FN3 domains. Cells were plated at 8000 cells per well in opaque white 96-well tissue culture-treated plates (Nunc) in 100 .mu.L/well of RPMI medium, supplemented with 10% FBS and allowed to attach overnight at 37.degree. C., 5% CO.sub.2. Twenty-four hours after plating, medium was aspirated and cells were replenished with serum-free RPMI medium. Twenty-four hours after serum starvation, cells were treated by addition of serum-free medium containing c-Met-binding FN3 domains (30 .mu.L/well). Cells were incubated at 37.degree. C., 5% CO.sub.2 for 72 hours. Viable cells were detected by addition of 100 .mu.L/well of CellTiter-Glo.RTM. reagent (Promega), followed by mixing on a plate shaker for 10 minutes. Plates were read on a SpectraMax M5 plate reader (Molecular Devices) set to luminescence mode, with a read time of 0.5 seconds/well. Data were plotted as raw luminescence units (RLU) against the logarithm of FN3 domain molar concentration. IC.sub.50 values were determined by fitting data to an equation for a sigmoidal dose response with variable slope using GraphPad Prism 4. Table 8 reports IC.sub.50 values ranging from 1 nM to >1000 nM. Characteristics of the c-Met binding FN3 domains are summarized in Table 8.
TABLE-US-00017 TABLE 8 Inhbibition pMet of inhibition Proliferation SEQ Affinity HGF H441 Cell in H441 of U87-MG Clone ID (Kd, competition binding cells cells (IC.sub.50, Name NO: nM) IC.sub.50 (nM) (EC.sub.50, nM) (IC.sub.50, nM) nM) P114AR5P74-A5 32 10.1 5.2 18.7 1078 464.4 P114AR5P75-E9 33 45.8 51.9 ND 1415 1193.9 P114AR7P92-F3 34 0.4 0.2 1.5 8.3 2.7 P114AR7P92-F6 35 3.1 2.2 4.9 165.3 350.5 P114AR7P92-G8 36 1.0 1.6 5.9 155.3 123.9 P114AR7P92-H5 37 11.6 ND 22.0 766.4 672.3 P114AR7P93-D11 38 ND ND 2.3 16 14.4 P114AR7P93-G8 39 6.9 1 3.8 459.5 103.5 P114AR7P93-H9 40 3.3 2.9 12.9 288.2 269.9 P114AR7P94-A3 41 0.4 0.2 1.4 5 9.3 P114AR7P94-E5 42 4.2 0.7 3.4 124.3 195.6 P114AR7P95-B9 43 0.5 0.3 ND 9.8 17.4 P114AR7P95-D3 44 0.3 0.2 1.5 4.6 1.7 P114AR7P95-D4 45 0.4 ND 1.4 19.5 19.4 P114AR7P95-E3 46 1.5 ND 3.2 204.6 209.2 P114AR7P95-F10 47 4.2 1.4 4.4 187.6 129.7 P114AR7P95-G7 48 20.0 ND 11.3 659.3 692 P114AR7P95-H8 49 3.7 ND 4.1 209.8 280.7
Thermal Stability of c-Met--Binding FN3 Domains Differential Scanning Calorimetry in PBS was Used to Assess the Stability of Each FN3 Domain. Results of the Experiment are Shown in Table 9.
TABLE-US-00018 TABLE 9 Clone Thermal SEQ ID Stability Name NO: (Tm, C) P114AR5P74-A5 32 74.1 P114AR5P75-E9 33 ND P114AR7P92-F3 34 81.5 P114AR7P92-F6 35 76.8 P114AR7P92-G8 36 90.9 P114AR7P92-H5 37 87 P114AR7P93-D11 38 ND P114AR7P93-G8 39 76.8 P114AR7P93-H9 40 88.2 P114AR7P94-A3 41 86.2 P114AR7P94-E5 42 80 P114AR7P95-89 43 86.3 P114AR7P95-D3 44 82 P114AR7P95-D4 45 85.3 P114AR7P95-E3 46 94.2 P114AR7P95-F10 47 85.2 P114AR7P95-G7 48 87.2 P114AR7P95-H8 49 83
Example 7. Generation and Characterization of Bispecific Anti-EGFR/c-Met Molecules
Generation of Bispecific EGFR/c-Met Molecules
[0600] Numerous combinations of the EGFR and c-Met-binding FN3 domains described in Examples 1-6 were joined into bispecific molecules capable of binding to both EGFR and c-Met. Additionally, EGFR-binding FN3 domains having amino acid sequences shown in SEQ ID NOs: 107-110 and c-Met binding FN3 domains having amino acid sequences shown in SEQ ID NOs: 111-114 were made and joined into bispecific molecules. Synthetic genes were created to encode for the amino acid sequences described in SEQ ID NOs: 50-72 and 106 (Table 10) such that the following format was maintained: EGFR-binding FN3 domain followed by a peptide linker followed by a c-Met-binding FN3 domain. A poly-histidine tag was incorporated at the C-terminus to aid purification. In addition to those molecules described in Table 10, the linker between the two FN3 domains was varied according to length, sequence composition and structure according to those listed in Table 11. It is envisioned that a number of other linkers could be used to link such FN3 domains Bispecific EGFR/c-Met molecules were expressed and purified from E. coli as described for monospecific EGFR or c-Met FN3 domains using IMAC and gel filtration chromatography steps.
TABLE-US-00019 TABLE 10 Bispecifcic EGFR/c- EGFR-binding FN3 Met molecule comain cMET-binding FN3 domain Linker SEQ ID SEQ ID SEQ ID SEQ ID Clone ID NO: Clone ID NO: Clone ID NO: Sequence NO: ECB1 50 P54AR4-83v2 27 P114AR5P74-A5 32 (GGGGS).sub.4 79 ECB2 51 P54AR4-83v2 27 P114AR7P94-A3 41 (GGGGS).sub.4 79 ECB3 52 P54AR4-83v2 27 P114AR7P93-H9 40 (GGGGS).sub.4 79 ECB4 53 P54AR4-83v2 27 P114AR5P75-E9 33 (GGGGS).sub.4 79 ECB5 54 P53A1R5-17v2 107 P114AR7P94-A3 41 (GGGGS).sub.4 79 ECB6 55 P53A1R5-17v2 107 P114AR7P93-H9 40 (GGGGS).sub.4 79 ECB7 56 P53A1R5-17v2 107 P114AR5P75-E9 33 (GGGGS).sub.4 79 ECB15 57 P54AR4-83v2 27 P114AR7P94-A3 41 (AP).sub.5 81 ECB27 58 P54AR4-83v2 27 P114AR5P74-A5 32 (AP).sub.5 81 ECB60 59 P53A1R5-17v2 107 P114AR7P94-A3 41 (AP).sub.5 81 ECB37 60 P53A1R5-17v2 107 P114AR5P74-A5 32 (AP).sub.5 81 ECB94 61 P54AR4-83v22 108 P114AR7P94-A3v22 111 (AP).sub.5 81 ECB95 62 P54AR4-83v22 108 P114AR9P121-A6v2 112 (AP).sub.5 81 ECB96 63 P54AR4-83v22 108 P114AR9P122-A7v2 113 (AP).sub.5 81 ECB97 64 P54AR4-83v22 108 P114AR7P95-C5v2 114 (AP).sub.5 81 ECB106 65 P54AR4-83v23 109 P114AR7P94-A3v22 111 (AP).sub.5 81 ECB107 66 P54AR4-83v23 109 P114AR9P121-A6v2 112 (AP).sub.5 81 ECB108 67 P54AR4-83v23 109 P114AR9P122-A7v2 113 (AP).sub.5 81 ECB109 68 P54AR4-83v23 109 P114AR7P95-C5v2 114 (AP).sub.5 81 ECB118 69 P53A1R5-17v22 110 P114AR7P94-A3v22 111 (AP).sub.5 81 ECB119 70 P53A1R5-17v22 110 P114AR9P121-A6v2 112 (AP).sub.5 81 ECB120 71 P53A1R5-17v22 110 P114AR9P122-A7v2 113 (AP).sub.5 81 ECB121 72 P53A1R5-17v22 110 P114AR7P95-C5v2 114 (AP).sub.5 81 ECB91 106 P54AR4-83v22 108 P114AR7P95-C5v2 114 (AP).sub.5 81 ECB18 118 P54AR4-83v2 27 P114AR5P74-A5 32 (AP).sub.5 81 ECB28 119 P53A1R5-17v2 107 P114AR5P74-A5 32 (AP).sub.5 81 ECB38 120 P54AR4-83v2 27 P114AR7P94-A3 41 (AP).sub.5 81 ECB39 121 P53A1R5-17v2 107 P114AR7P94-A3 41 (AP).sub.5 81
TABLE-US-00020 TABLE 11 SEQ ID Linker length in Linker NO: amino acids Structure GS 78 2 Disordered GGGGS 105 5 Disordered (GGGGS).sub.4 79 20 Disordered (AP).sub.2 80 4 Rigid (AP).sub.5 81 5 Rigid (AP).sub.10 82 20 Rigid (AP).sub.20 83 40 Rigid A(EAAAK).sub.5AAA 84 29 .alpha.-helical
Bispecific EGFR/c-Met Molecules Enhance Potency Compared to Monospecific Molecules Alone, Suggesting Avidity
[0601] NCI-H292 cells were plated in 96 well plates in RPMI medium containing 10% FBS. 24 hours later, medium was replaced with serum free RPMI. 24 hours after serum starvation, cells were treated with varying concentrations of FN3 domains: either a high affinity monospecific EGFR FN3 domain (P54AR4-83v2), a weak affinity monospecific c-Met FN3 domain (P114AR5P74-A5), the mixture of the two monospecific EGFR and c-Met FN3 domains, or a bispecific EGFR/c-Met molecules comprised of the low affinity c-Met FN3 domain linked to the high affinity EGFR FN3 domain (ECB1). Cells were treated for 1 h with the monospecific or bispecific molecules and then stimulated with EGF, HGF, or a combination of EGF and HGF for 15 minutes at 37.degree. C., 5% CO.sub.2. Cells were lysed with MSD Lysis Buffer and cell signaling was assessed using appropriate MSD Assay plates, according to manufacturer's instructions, as described above.
[0602] The low affinity c-Met FN3 domain inhibited phosphorylation of c-Met with an IC.sub.50 of 610 nM (FIG. 4). As expected the EGFR FN3 domain was not able to inhibit c-Met phosphorylation and the mixture of the mono-specific molecules looked identical to the c-Met FN3 domain alone. However, the bi-specific EGFR/c-Met molecule inhibited phosphorylation of c-Met with an IC.sub.50 of 1 nM (FIG. 4), providing more than a 2-log shift in improving potency relative to the c-Met monospecific alone.
[0603] The potential for the bispecific EGFR/c-Met molecule to enhance the inhibition of c-Met and/or EGFR phosphorylation through an avidity effect was evaluated in multiple cell types with variable c-Met and EGFR densities and ratios (FIG. 5). NCI-H292, NCI-H441, or NCI-H596 cells were plated in 96 well plates in RPMI medium containing 10% FBS. 24 hours later, medium was replaced with serum free RPMI. 24 hours after serum starvation, cells were treated with varying concentrations of either monospecific EGFR-binding FN3 domain, monospecific c-Met FN3 domain, or a bispecific EGFR/c-Met molecule (ECB5, comprised of P53A1R5-17v2 and P114AR7P94-A3). Cells were treated for 1 h with the monospecific or bispecific molecules and then stimulated with EGF, HGF, or a combination of EGF and HGF for 15 minutes at 37.degree. C., 5% CO.sub.2. Cells were lysed with MSD Lysis Buffer and cell signaling was assessed using appropriate MSD Assay plates, according to manufacturer's instructions, as described above.
[0604] FIG. 5 (A-C) shows the inhibition of EGFR using a monospecific EGFR-binding FN3 domain compared to a bispecific EGFR/cMet molecule in three different cell lines. To assess avidity in an EGFR phosphorylation assay, a medium affinity EGFR-binding FN3 domain (1.9 nM) (P53A1R5-17v2) was compared to a bispecific EGFR/c-Met molecule containing the same EGFR-binding FN3 domain linked to a high-affinity c-Met-binding FN3 domain (0.4 nM) (P114AR7P94-A3). In NCI-H292 and H596 cells, inhibition of phosphorylation of EGFR was comparable for the monospecific and bispecific molecules (FIGS. 5A and 5B), likely because these cell lines have a high ratio of EGFR to c-Met receptors. To test this theory, inhibition of EGFR phosphorylation was evaluated in NCI-H441 cells which exhibit more c-Met receptors than EGFR. Treatment of NCI-H441 cells with the bispecific EGFR/c-Met molecule decreased the IC.sub.50 for inhibition of EGFR phosphorylation compared to the monospecific EGFR-binding FN3 domain by 30-fold (FIG. 5C).
[0605] The potential for enhanced potency with a bi-specific EGFR/c-Met molecule was evaluated in a c-Met phosphorylation assay using a molecule with a high affinity to EGFR (0.26 nM) and medium affinity to c-Met (10.1 nM). In both NCI-H292 and NCI-H596 cells, the inhibition of phosphorylation of c-Met was enhanced with the bispecific molecule compared to the monospecific c-Met-binding FN3 domain, by 134 and 1012 fold, respectively (FIGS. 3D and 3E).
[0606] It was verified that the enhanced potency for inhibition of EGFR and c-Met phosphorylation with the bispecific EGFR/c-Met molecules translated into an enhanced inhibition of signaling and proliferation. For these experiments, the mixture of FN3 EGFR-binding and c-Met-binding FN3 domains was compared to a bispecific EGFR/c-Met molecule. As described in Tables 12 and 13, the IC.sub.50 values for ERK phosphorylation (Table 12) and proliferation of NCI-H292 cells (Table 13) were decreased when cells were treated with the bispecific EGFR/c-Met molecule compared to the mixture of the monospecific binders. The IC.sub.50 for inhibition of ERK phosphorylation for the bi-specific EGFR/c-Met molecule was 143-fold lower relative to the mixture of the two monospecific EGFR and c-Met FN3 domains, showing the effect of avidity to the potency of the molecules in this assay. In Table 12, the monospecific EGFR- and c-Met binding FN3 domains do not fully inhibit activity and therefore the IC.sub.50 values shown should be considered lower limits. The proliferation assay was completed using different combinations EGFR and c-Met binding FN3 domains either as a mixture or linked in a bispecific format. The IC.sub.50 for inhibition of proliferation for the bispecific EGFR/c-Met molecule was 34-236-fold lower relative to the mixture of the monospecific parent EGFR or c-Met binding FN3 domains. This confirmed that the avidity effect observed at the level of the receptors (FIG. 4 and FIG. 5) translates into an improvement in inhibiting cell signaling (Table 12) and cell proliferation (Table 13).
TABLE-US-00021 TABLE 12 Specificity of the IC.sub.50 (nM) FN3-domain (ERK molecule Clone name Type phosphorylation) EGFR P54AR4-83v2 monospecific >10,000 c-Met P114AR5P74-A5 monospecific 2366 EGFR or c-Met P54AR4-83v2 + mixture of 798.4 P114AR5P74-A5 monospecific molecules EGFR and c-Met ECB1 bispecific 5.6
TABLE-US-00022 TABLE 13 EGFR- IC.sub.50 for Fold increase binding mixture of IC.sub.50 for in IC.sub.50 for FN3 c-Met binding monospecific bispecific mixture of domain FN3 domain molecules molecule monospecific/ (affinity) (affinity) (nM) (nM) bispecific P54AR4- P114ARP94- 36.5 1.04 35 83v2 A3 (0.4 nM) (0.26 nM) P54AR4- P114AR7P93- 274.5 8.05 34 83v2 H9 (3.3 nM) (0.26 nM) P54AR4- P114AR5P74- 1719 7.29 236 83v2 A5 (10.1 nM) (0.26 nM)
In Vivo Tumor Xenografts: PK/PD
[0607] In order to determine efficacy of the monospecific and bispecific FN3 domain molecules in vivo, tumor cells were engineered to secrete human HGF (murine HGF does not bind to human c-Met). Human HGF was stably expressed in NCI-H292 cells using lentiviral infection (Lentiviral DNA vector expressing human HGF (Accession # X16322) and lentiviral packaging kit from Genecopoeia). After infection, HGF-expressing cells were selected with 4 .mu.g/mL puromycin (Invitrogen). Human HGF protein was detected in the conditioned medium of pooled cells using assay plates from MesoScale Discovery.
[0608] SCID Beige mice were subcutaneously inoculated with NCI-H292 cells expressing human HGF (2.0.times.10.sup.6 cells in Cultrex (Trevigen) in a volume of 200 .mu.L) on the dorsal flank of each animal. Tumor measurements were taken twice weekly until tumor volumes ranged between 150-250 mm.sup.3. Mice were then given a single i.p. dose of bispecific EGFR/c-Met molecules (linked to an albumin binding domain to increase half-life) or PBS vehicle. At 6 h or 72 h after dosing, tumors were extracted and immediately frozen in liquid nitrogen. Blood samples were collected via cardiac puncture into 3.8% citrate containing protease inhibitors Immediately after collection, the blood samples were centrifuged and the resulting plasma was transferred to sample tubes and stored at -80.degree. C. Tumors were weighed, cut into small pieces, and lysed in Lysing Matrix A tubes (LMA) containing RIPA buffer with HALT protease/phosphatase inhibitors (Pierce), 50 mM sodium fluoride (Sigma), 2 mM activated sodium orthovanadate (Sigma), and 1 mM PMSF (MesoScale Discovery). Lysates were removed from LMA matrix and centrifuged to remove insoluble protein. The soluble tumor protein was quantified with a BCA protein assay and diluted to equivalent protein levels in tumor lysis buffer. Phosphorylated c-Met, EGFR and ERK were measured using assay plates from MesoScale Discovery (according to Manufacturer's protocol and as described above).
[0609] FIG. 6 shows the results of the experiments. Each bispecific EGFR/c-Met molecule significantly reduced the levels of phosphorylated c-Met, EGFR, and ERK at both 6 h and 72 h. The data presented in FIG. 6 show the importance of inhibiting both c-Met and EGFR simultaneously and how the affinity of the bispecific EGFR/c-Met molecule for each receptor plays a role in inhibition of downstream ERK. The molecules containing the high affinity EGFR-binding FN3 domains (P54AR4-83v2; shown as "8" in the Figure, K.sub.D=0.26 nM) inhibited phosphorylation of EGFR to a larger extent compared to those containing the medium affinity EGFR-binding FN3 domains (P53A1R5-17v2; shown as "17" in the figure K.sub.D=1.9 nM) at both 6 h and 72 h. All four bispecific molecules tested completely inhibited phosphorylation of ERK at the 6 hour time point, regardless of affinity. At the 72 hour time point, the molecules containing the tight affinity c-Met-binding domain (P114AR7P94-A3; shown as "A3" in the figure K.sub.D=0.4 nM) significantly inhibited phosphorylation of ERK compared to the medium affinity c-Met-binding FN3 domain (P114AR5P74-A5; shown as "A5" in the Figure; K.sub.D=10.1 nM; FIG. 6).
[0610] The concentration of each bispecific EGFR/c-Met molecule was measured at 6 and 72 hours after dosing in the blood and in the tumor (FIG. 7). Interestingly, the bispecific molecule with the medium affinity EGFR-binding domain (P53A1R5-17v2; K.sub.D=1.9 nM) but high affinity c-Met-binding FN3 domain (P114AR7P94-A3; K.sub.D=0.4 nM) had significantly more tumor accumulation at 6 hours relative to the other molecules, while the difference is diminished by 72 hours. It can be hypothesized that cells outside the tumor have lower levels of both EGFR and c-Met surface expression and therefore the medium affinity EGFR molecule doesn't bind to normal tissue as tightly compared to the higher affinity EGFR-binding FN3 domain. Therefore there is more free medium affinity EGFR-binding FN3 domain available to bind in the tumor. Therefore, identifying the appropriate affinities to each receptor may allow for identification of a therapeutic with decreased systemic toxicities and increased tumor accumulation.
Tumor Efficacy Studies with Bispecific EGFR/c-Met Molecules
[0611] SCID Beige mice were subcutaneously inoculated with NCI-H292 cells expressing human HGF (2.0.times.10.sup.6 cells in Cultrex (Trevigen) in 200 .mu.L) in the dorsal flank of each animal. One week after implantation, mice were stratified into groups with equivalent tumor volumes (mean tumor volume=77.9+/-1.7 mm.sup.3). Mice were dosed three times per week with the bispecific molecules and tumor volumes were recorded twice weekly. Tumor growth inhibition (TGI) was observed with four different bispecific molecules, with variable affinities for c-Met and EGFR. FIG. 8 shows the benefit of inhibiting both c-Met and EGFR as a delay in tumor growth was observed in the mice treated with molecules containing the high affinity EGFR-binding FN3 domain compared to the medium affinity EGFR-binding FN3 domain when the c-Met-binding FN3 domain was medium affinity (open vs. closed triangles, P54AR4-83v2-P114AR5P74-A5 compared to P53A1R5-17-P114AR5P74-A5). In addition, the data shows the importance of having a high affinity c-Met-binding FN3 domain as bispecific molecules containing either the high or medium affinity EGFR-binding FN3 domain but high affinity c-Met-binding FN3 domain showed the most efficacy (dotted gray and black lines, P54AR4-83v2-P114AR7P94-A3 and P53A1R5-17v2-P114AR7P94-A3).
Efficacy of Bispecific Molecule and Other Inhibitors of EGFR and c-Met
[0612] The in vivo therapeutic efficacies of a bispecific EGFR/c-Met molecule (ECB38) and the small molecule inhibitors crizotinib (c-Met inhibitor) and erlotinib (EGFR inhibitor), cetuximab (anti-EGFR antibody), each as a single agent, and the combination of crizotinib and erlotinib were evaluated in the treatment of subcutaneous H292-HGF human lung cancer xenograft model in SCID/Beige mice.
[0613] The H292-HGF cells were maintained in vitro in RPMI1640 medium supplemented with fetal bovine serum (10% v/v), and L-glutamine (2 mM) at 37.degree. C. in an atmosphere of 5% CO.sub.2 in air. The cells were routinely subcultured twice weekly by trypsin-EDTA treatment. The cells growing in an exponential growth phase were harvested and counted for tumor inoculation.
[0614] Each mouse was inoculated subcutaneously at the right flank region with H292-HGF tumor cells (2.times.10.sup.6) in 0.1 ml of PBS with Cultrex (1:1) for tumor development. The treatments were started when the mean tumor size reached 139 mm.sup.3. The test article administration and the animal numbers in each study group were shown in the following experimental design table. The date of tumor cell inoculation was denoted as day 0. Table 14 shows the treatment groups.
TABLE-US-00023 TABLE 14 Dose Dosing Planned Actual Group N Treatment (mg/kg) Route Schedule Schedule 1 10 Vehicle 0 i.p. QD .times. 3 QD .times. 3 Control weeks weeks 2 10 bispecific 25 i.p. 3 times/ 3 times/ EGFR/c-Met week .times. week .times. molecule 3 weeks 3 weeks 3 10 crizotinib 50 p.o. QD .times. 3 QD .times. 17 weeks days 4 10 erlotinib 50 p.o. QD .times. 2 QD .times. 3 weeks weeks 5 10 crizotinib 50 p.o. QD .times. 3 QD .times. 3 weeks weeks 6 10 cetuximab 1 mg/mouse i.p. Q4d*6 Q4d*6 N: animal number; p.o.: oral administration; i.p.: intraperitoneal injection 3 times/week: doses were given on days 1, 3 and 5 of the week. QD: once daily Q4d: once every four days; the interval of the combination of crizotinib and erlotinib was 0.5 hrs; dosing volume was adjusted based on body weight (10 l/g); a: dosing was not given on day 14 post grouping.
Before commencement of treatment, all animals were weighed and the tumor volumes were measured. Since the tumor volume can affect the effectiveness of any given treatment, mice were assigned into groups using randomized block design based upon their tumor volumes. This ensures that all the groups are comparable at the baseline. The randomized block design was used to assign experimental animals to groups. First, the experimental animals were divided into homogeneous blocks according to their initial tumor volume. Secondly, within each block, randomization of experimental animals to treatments was conducted. Using randomized block design to assign experimental animals ensured that each animal had the same probability of being assigned to a given treatment and therefore systematic error was reduced.
[0615] At the time of routine monitoring, the animals were checked for any effects of tumor growth and treatments on normal behavior, such as mobility, visual estimation of food and water consumption, body weight gain/loss (body weights were measured twice weekly), eye/hair matting and any other abnormal effect.
[0616] The endpoint was whether tumor growth can be delayed or tumor bearing mice can be cured. Tumor size was measured twice weekly in two dimensions using a caliper, and the volume was expressed in mm.sup.3 using the formula: V=0.5 a.times.b.sup.2 where a and b are the long and short diameters of the tumor, respectively. The tumor size was then used for calculations of both T-C and T/C values. T-C was calculated with T as the time (in days) required for the mean tumor size of the treatment group to reach 1000 mm.sup.3, and C was the time (in days) for the mean tumor size of the control group to reach the same size. The T/C value (in percent) was an indication of antitumor efficacy; T and C were the mean tumor volume of the treated and control groups, respectively, on a given day. Complete tumor regression (CR) is defined as tumors that are reduced to below the limit of palpation (62.5 mm.sup.3). Partial tumor regression (PR) is defined as tumors that are reduced from initial tumor volume. A minimum duration of CR or PR in 3 or more successive tumor measurements is required for a CP or PR to be considered durable.
[0617] Animals for which the body weight loss exceeded 20%, or for which the mean tumor size of the group exceeds 2000 mm.sup.3 were euthanized. The study was terminated after two weeks of observation after the final dose.
[0618] Summary statistics, including mean and the standard error of the mean (SEM), are provided for the tumor volume of each group at each time point are shown in Table 15. Statistical analyses of difference in tumor volume among the groups were evaluated using a one-way ANOVA followed by individual comparisons using Games-Howell (equal variance not assumed). All data were analyzed using SPSS 18.0. p<0.05 was considered to be statistically significant.
TABLE-US-00024 TABLE 15 Tumor volume (mm.sup.3)a bispecific crizotinib; EGFR/c-Met erlotinib molecule at crizotinib at erlotinib at at 50 mg/kg; cetuximab at Days Vehicle 25 mg/kg 50 mg/kg 50 mg/kg 50 mg/kg 1 mg/mouse 7 139 .+-. 7 137 .+-. 7 140 .+-. 9 141 .+-. 8 139 .+-. 8 139 .+-. 10 9 230 .+-. 20 142 .+-. 7 217 .+-. 20 201 .+-. 19 134 .+-. 9 168 .+-. 13 13 516 .+-. 45 83 .+-. 6 547 .+-. 43 392 .+-. 46 109 .+-. 10 212 .+-. 20 16 808 .+-. 104 44 .+-. 7 914 .+-. 92 560 .+-. 70 127 .+-. 15 252 .+-. 28 20 1280 .+-. 209 30 .+-. 6 1438 .+-. 239 872 .+-. 136 214 .+-. 30 371 .+-. 48 23 1758 .+-. 259 23 .+-. 7 2102 .+-. 298 1122 .+-. 202 265 .+-. 40 485 .+-. 61 27 2264 .+-. 318 21 .+-. 5 -- 1419 .+-. 577 266 .+-. 42 640 .+-. 82 30 -- 23 .+-. 6 -- 1516 .+-. 623 482 .+-. 61 869 .+-. 100
[0619] The mean tumor size of the vehicle treated group (Group 1) reached 1,758 mm.sup.3 at day 23 after tumor inoculation. Treatment with the bispecific EGFR/c-Met molecule at 25 mg/kg dose level (Group 2) led to complete tumor regression (CR) in all mice which were durable in >3 successive tumor measurements (average TV=23 mm.sup.3, T/C value=1%, p=0.004 compared with the vehicle group at day 23).
[0620] Treatment with crizotinib as a single agent at 50 mg/kg dose level (Group 3) showed no antitumor activity; the mean tumor size was 2,102 mm.sup.3 at day 23 (T/C value=120%, p=0.944 compared with the vehicle group).
[0621] Treatment with erlotinib as a single agent at 50 mg/kg dosing level (Group 4) showed minor antitumor activity, but no significant difference was found compared with the vehicle group; the mean tumor size was 1,122 mm.sup.3 at day 23 (T/C value=64%, p=0.429 compared with the vehicle group), with 4 days of tumor growth delay at tumor size of 1,000 mm.sup.3 compared with the vehicle group.
[0622] The combination of crizotinib (50 mg/kg, Group 5) and erlotinib (50 mg/kg, Group 5) showed significant antitumor activity; the mean tumor size was 265 mm.sup.3 at day 23 (T/C=15%; p=0.008), with 17 days of tumor growth delay at tumor size of 1,000 mm.sup.3 compared with the vehicle group.
[0623] Cetuximab at 1 mg/mouse dosing level as a single agent (Group 6) showed significant antitumor activities; the mean tumor size was 485 mm.sup.3 at day 23 (T/C=28%; p=0.018), with 17 days of tumor growth delay at tumor size of 1,000 mm.sup.3 compared with the vehicle group. FIG. 15 and Table 16 show the anti-tumor activities of the various therapies.
TABLE-US-00025 TABLE 16 Tumor Size T-C (mm.sup.3) at T/C (days) at P Treatment day 23 (%) 1000 mm.sup.3 value Vehicle 1758 .+-. 259 -- -- -- bispecific EGFR/c-Met 23 .+-. 7 1 -- 0.004 molecule (25 mg/kg) crizotinib (50 mg/kg) 2102 .+-. 298 120 -1 0.944 erlotinib (50 mg/kg) 1122 .+-. 202 64 4 0.429 crizotinib + erlotinib 265 .+-. 40 15 17 0.008 (50 mg/kg + 50 mg/kg) cetuximab (1 485 .+-. 61 28 17 0.018 mg/mouse)
[0624] Medium to severe body weight loss was observed in the vehicle group which might be due to the increasing tumor burden; 3 mice died and 1 mouse were euthanized when BWL>20% by day 23. Slight toxicity of the bispecific EGFR/c-Met molecule was observed in Group 2; 3 mice were euthanized when BWL>20% during the treatment period; the body weight was gradually recovered when the treatment was withdrawn during the 2 weeks of observation period. More severe body weight loss was observed in the crizotinib or erlotinib monotherapy group compared to the vehicle group, suggesting the treatment related toxicity. The combination of crizotinib and erlotinib was generally tolerated during the dosing phase, but severe body weight loss was observed at the end of the study, which might be due to the resumption of the fast tumor growth during the non-treatment period. The monotherapy of cetuximab was well tolerated in the study; body weight loss was only observed at the end of the study due to the resume of the tumor growth.
[0625] In summary, the bispecific EGFR/c-Met molecule at 25 mg/kg (3 times/week x 3 weeks) produced a complete response in H292-HGF human lung cancer xenograft model in SCID/Beige mice. The treatment was tolerated in 7 out of 10 mice, and resulted in severe body weight loss in 3 out of 10 mice. FIG. 9 shows the impact of the various therapies on tumor size during the time points after treatment.
Example 8: Half-Life Extension of the Bispecific EGFR/c-Met Molecules
[0626] Numerous methods have been described to reduce kidney filtration and thus extend the serum half-life of proteins including modification with polyethylene glycol (PEG) or other polymers, binding to albumin, fusion to protein domains which bind to albumin or other serum proteins, genetic fusion to albumin, fusion to IgG Fc domains, and fusion to long, unstructured amino acid sequences.
[0627] Bispecific EGFR/c-Met molecules were modified with PEG in order to increase the hydrodynamic radius by incorporating a free cysteine at the C-terminus of the molecule. Most commonly, the free thiol group of the cysteine residue is used to attach PEG molecules that are functionalized with maleimide or iodoacetemide groups using standard methods. Various forms of PEG can be used to modify the protein including linear PEG of 1000, 2000, 5000, 10,000, 20,000, or 40,000 kDa. Branched PEG molecules of these molecular weights can also be used for modification. PEG groups may also be attached through primary amines in the bispecific EGFR/c-Met molecules in some instances.
[0628] In addition to PEGylation, the half-life of bispecific EGFR/c-Met molecules was extended by producing these proteins as fusion molecules with a naturally occurring 3-helix bundle serum albumin binding domain (ABD) or a consensus albumin binding domain (ABDCon). These protein domains were linked to the C-terminus of the c-Met-binding FN3 domain via any of the linkers described in Table 12. The ABD or ABDCon domain may also be placed between the EGFR-binding FN3 domain and the c-Met binding FN3 domain in the primary sequence.
Example 9: Characterization of Select Bispecific EGFR/c-Met Molecules
[0629] Select bispecific EGFR/c-Met molecules were characterized for their affinity to both EGFR and c-Met, their ability to inhibit EGFR and c-Met autophosphorylation, and their effect on proliferation of HGF cells. Binding affinity of the bispecific EGFR/c-Met molecules to recombinant EGFR and/or c-Met extracellular domain was further evaluated by surface Plasmon resonance methods using a Proteon Instrument (BioRad) according to protocol described in Example 3. Results of the characterization are shown in Table 17.
TABLE-US-00026 TABLE 17 H292-HGF H292 Proliferation pMet pEGFR inhibition inhibition inhibition in HGF- K.sub.D K.sub.D in H441 in induced (EGFR, (c-Met, cells H292 cells H292 cells nM) nM) (IC.sub.50, nM) (IC.sub.50, nM) (IC.sub.50, nM) ECB15 0.2 2.6 n/a 4.2 23 ECB94 1 4.3 53.8 5.1 29.6 ECB95 1.1 6.2 178.8 13.6 383.4 ECB96 1.6 22.1 835.4 24.7 9480 ECB97 1.3 1.7 24.2 16.6 31.0 ECB106 16.7 5.1 53.3 367.4 484.5 ECB107 16.9 9 29.9 812.3 2637 ECB108 15.3 25.5 126.2 814.4 11372 ECB109 17.3 2.1 26 432 573.6
Example 10. Generation of Bispecific EGFR/cMet Antibodies
[0630] Several monospecific EGFR and c-Met antibodies were expressed as IgG1, kappa, having Fc substitutions K409R or F405L (numbering according to the EU index) in their Fc regions. The monospecific antibodies were expressed in two CHO cell lines, one cell line having reduced fucosylation ability resulting in antibodies with 1-15% fucose content in the antibody polysaccharide chain.
[0631] The monospecific antibodies were purified using standard methods using a Protein A column (HiTrap MabSelect SuRe column). After elution, the pools were dialyzed into D-PBS, pH 7.2
[0632] Bispecific EGFR/c-Met antibodies were generated by combining a monospecific EGFR mAb and a monospecific c-Met mAb in in vitro Fab arm exchange (as described in WO2011/131746). Briefly, at about 1-20 mg/ml at a molar ratio of 1:1 of each antibody in PBS, pH 7-7.4 and 75 mM 2-mercaptoethanolamine (2-MEA) was mixed together and incubated at 25-37.degree. C. for 2-6 h, followed by removal of the 2-MEA via dialysis, diafiltration, tangential flow filtration and/or spinned cell filtration using standard methods.
[0633] Several monospecific anti-EGFR antibodies and anti-c-Met antibodies were combined in matrix in in vitro Fab arm exchange to generate bispecific antibodies that were subsequently characterized further. The generated bispecific antibodies were ranked using a four step strategy using assays as follows: Step 1: binding to NCI-H441, NCI-H1975 and A549 cells in a FACS assay. Step 2: inhibition of pMet phosphorylation in A549 cells. Step 3: inhibition of proliferation in NCI-H1975, KP4 and NCI-H441 cells. Step 4: inhibition of EGFR phosphorylation in A549 and SNU-5 cells. Noteworthy, the characteristics of the parental antibodies were not preserved in the bispecific antibody. For example, the presence of certain EGFR binding arms in the bispecific antibody resulted in a loss or reduced inhibition, or enhanced c-Met phosphorylation. Based on the characterization studies select pairs were chosen.
[0634] A monospecific bivalent anti-EGFR antibody E1-K409R was generated comprising the VH and VL regions of an anti-EGFR antibody 2F8 having the VH of SEQ ID NO: 189 and the VL of SEQ ID NO: 190 (antibody 2F8 is described in Int. Pat. Publ. No. WO2002/100348) and an IgG1 constant region with a K409R substitution.
[0635] A monospecific bivalent anti-EGFR antibody E1-F405L was generated comprising the VH and VL regions of an anti-EGFR antibody 2F8 having the VH of SEQ ID NO: 189 and the VL of SEQ ID NO: 190 (antibody 2F8 is described in Int. Pat. Publ. No. WO2002/100348) and an IgG1 constant region with a F405L substitution.
[0636] A monospecific bivalent anti-EGFR antibody E2-K409R was generated comprising the VH and VL regions of an anti-EGFR antibody 018 having the VH of SEQ ID NO: 191 and the VL of SEQ ID NO: 192 (antibody 018 is described in Int. Pat. Publ. No. WO2009/030239) and an IgG1 constant region with a K409R substitution.
[0637] A monospecific bivalent anti-EGFR antibody E2-F405L was generated comprising the VH and VL regions of an anti-EGFR antibody 018 having the VH of SEQ ID NO: 191 and the VL of SEQ ID NO: 192 (antibody 018 is described in Int. Pat. Publ. No. WO2009/030239) and an IgG1 constant region with a F405L substitution.
[0638] A monospecific bivalent anti-c-Met antibody M1-K409R was generated comprising the VH and VL regions of an anti-c-Met antibody 069 having the VH of SEQ ID NO: 193 and the VL of SEQ ID NO: 194 (antibody 069 is described in WO2011/110642) and an IgG1 constant region with a K409R substitution.
[0639] A monospecific bivalent anti-c-Met antibody M1-F405L was generated comprising the VH and VL regions of an anti-c-Met antibody 069 having the VH of SEQ ID NO: 193 and the VL of SEQ ID NO: 194 (antibody 069 is described in WO2011/110642) and an IgG1 constant region with a F405L substitution.
[0640] A monospecific anti-c-Met antibody M2-K409R was generated comprising the VH and VL regions of an anti-c-Met antibody 058 having the VH of SEQ ID NO: 195 and the VL of SEQ ID NO: 196 (antibody 058 is described in WO2011/110642) and an IgG1 constant region with a K409R substitution.
[0641] A monospecific anti-c-Met antibody M2-F405L was generated comprising the VH and VL regions of an anti-c-Met antibody 058 having the VH of SEQ ID NO: 195 and the VL of SEQ ID NO: 196 (antibody 058 is described in WO2011/110642) and an IgG1 constant region with a F405L substitution.
[0642] The VH, VL, HC and LC sequences of the antibodies are shown below:
TABLE-US-00027 >EGFR mAb E1 VH SEQ ID NO: 189 QVQLVESGGGVVQPGRSLRLSCAASGFTFSTYGMHWVRQAPGKGLEWVAV IWDDGSYKYYGDSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARDG ITMVRGVMKDYFDYWGQGTLVTVSS >EGFR mAb E1 VL SEQ ID NO: 190 AIQLTQSPSSLSASVGDRVTITCRASQDISSALVWYQQKPGKAPKLLIYD ASSLESGVPSRFSGSESGTDFTLTISSLQPEDFATYYCQQFNSYPLTFGG GTKVEIK >EGFR mAb E2 VH SEQ ID NO: 191 EVQLVESGGG LVQPGGSLRL SCAASGFTFS SYWMNWVRQA PGKGLEWVAN IKKDGSEKYY VDSVKGRFTI SRDNAKNSLY LQMNSLRAED TAVYYCARDL GWGWGWYFDL WGRGTLVTVSS >EGFR mAb E2 VL SEQ ID NO: 192 EIVLTQSPAT LSLSPGERAT LSCRASQSVS SYLAWYQQKP GQAPRLLIYD ASNRATGIPARFSGSGSGTD FTLTISSLEP EDFAVYYCQQ RSNWPPTFGQ GTKVEIK >cMet mAb M1 VH SEQ ID NO: 193 QVQLVQSGAEVKKPGASVKVSCETSGYTFTSYGISWVRQAPGHGLEWMGW ISAYNGYTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDL RGTNYFDYWGQGTLVTVSS >cMet mAb M1VL SEQ ID NO: 194 DIQMTQSPSSVSASVGDRVTITCRASQGISNWLAWFQHKPGKAPKLLIYA ASSLLSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQANSFP-ITFG QGTRLEIK >cMet mAb M2 VH SEQ ID NO: 195 EVQLVESGGGLVKPGGSLKLSCAASGFTFSDYYMYWVRQTPEKRLEWVAT ISDDGSYTYYPDSVKGRFTISRDNAKNNLYLQMSSLKSEDTAMYYCAREG LYYYGSGSYYNQDYWGQGTLVTVSS >cMet mAb M2 VL SEQ ID NO: 196 QLTQSPSSLSASVGDRVTITCRASQGLSSALAWYRQKPGKAPKLLIYDAS SLESGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQFTSYPQITFGQG TRLEIK >EM1-mAb H1 (anti-EGFR, 405L) SEQ ID NO: 199 QVQLVESGGGVVQPGRSLRLSCAASGFTFSTYGMHWVRQAPGKGLEWVAV IWDDGSYKYYGDSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARDG ITMVRGVMKDYFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAAL GCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSS LGTQTYICNVNHKPSNTKVDKRVEPKSCDKTHTCPPCPAPELLGGPSVFL FPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPR EEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQ PREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYK TTPPVLDSDGSFLLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLS LSPGK >EM-1 mAb L1 SEQ ID NO: 200 AIQLTQSPSSLSASVGDRVTITCRASQDISSALVWYQQKPGKAPKLLIYD ASSLESGVPSRFSGSESGTDFTLTISSLQPEDFATYYCQQFNSYPLTFGG GTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKV DNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQG LSSPVTKSFNRGEC >EM-1 mAb H2 (K409R, anti-cMet) SEQ ID NO: 201 QVQLVQSGAEVKKPGASVKVSCETSGYTFTSYGISWVRQAPGHGLEWMGW ISAYNGYTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDL RGTNYFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKD YFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTY ICNVNHKPSNTKVDKRVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPK DTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNS TYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQV YTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVL DSDGSFFLYSRLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK > EM-1 mAb L2 SEQ ID NO: 202 DIQMTQSPSSVSASVGDRVTITCRASQGISNWLAWFQHKPGKAPKLLIYA ASSLLSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQANSFPITFGQ GTRLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKV DNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQG LSSPVTKSFNRGEC >E2 mAb HC1 (EGFR-F405L) SEQ ID NO: 234 EVQLVESGGG LVQPGGSLRL SCAASGFTFS SYWMNWVRQA PGKGLEWVAN IKKDGSEKYY VDSVKGRFTI SRDNAKNSLY LQMNSLRAED TAVYYCARDL GWGWGWYFDLWGRGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLV KDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQ TYICNVNHKPSNTKVDKRVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPK PKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQY NSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREP QVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPP VLDSDGSFLLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG K >E2 mAb LC1 (EGFR) SEQ ID NO: 235 EIVLTQSPAT LSLSPGERAT LSCRASQSVS SYLAWYQQKP GQAPRLLIYD ASNRATGIPARFSGSGSGTD FTLTISSLEP EDFAVYYCQQ RSNWPPTFGQ GTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKV DNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQG LSSPVTKSFNRGEC >E2 mAb HC2 (c-Met-K409R) SEQ ID NO: 236 EVQLVESGGGLVKPGGSLKLSCAASGETFSDYYMYWVRQTPEKRLEWVAT ISDDGSYTYYPDSVKGRFTISRDNAKNNLYLQMSSLKSEDTAMYYCAREG LYYYGSGSYYNQDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAAL GCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSS LGTQTYICNVNHKPSNTKVDKRVEPKSCDKTHTCPPCPAPELLGGPSVFL FPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPR EEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQ PREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYK TTPPVLDSDGSFFLYSRLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLS LSPGK >E2 mAb LC2 (cMet) SEQ ID NO: 237 QLTQSPSSLSASVGDRVTITCRASQGLSSALAWYRQKPGKAPKLLIYDAS SLESGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQFTSYPQITFGQG TRLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVD NALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGL SSPVTKSFNRGEC
[0643] The generated monospecific anti-EGFR and c-Met antibodies were mixed for in vitro Fab arm exchange in matrix and characterized in various assays. The bispecific antibody EM1-mAb comprises the EGFR binding arm of mAb E1-F405L and the c-Met binding arm of mAb M1-K409R. The bispecific antibody EM2-mAb comprises the EGFR binding arm of mAb E2-F405L and the c-Met binding arm of mAb M2-K409R. The bispecific antibody EM3-mAb comprises the EGFR binding arm of mAb E1-K409R and the c-Met binding arm of mAb M1-F405L. The bispecific antibody EM4-mAb comprises the EGFR binding arm of mAb E2-K409R and the c-Met binding arm of mAb M2-F405L. EM1-mAb and EM3-mAb had comparable characteristics.
[0644] The bispecific EM1-mAb was cultured in a CHO cell line having reduced fucosylation ability of glycoproteins, and hence have a fucosyl content of about 1-15%. The removal of the core fucose from the biantennary complex-type oligosaccharides attached to the Fc regions significantly enhances the ADCC of antibodies via improved Fc.gamma.RIIIa binding without altering antigen binding or CDC activity. Such mAbs can be achieved using different methods reported to lead to the successful expression of relatively high defucosylated therapeutic antibodies bearing the biantennary complex-type of Fc oligosaccharides and are described supra.
Example 11. Purification of Bispecific EGFR/c-Met Antibodies
[0645] The bispecific EM1-mAb was further purified after the in vitro Fab-arm exchange using hydrophobic interaction chromatography to minimize residual parental c-Met and EGFR antibodies using standard methods.
Example 12. Characterization of Bispecific EGFR/c-Met Antibodies
[0646] The EGFR/c-Met bispecific antibody EM1-mAb was tested in various assays for its characteristics including inhibition of EGF-stimulated EGFR phosphorylation, HGF-stimulated c-Met phosphorylation, ERK1/2 phosphorylation, AKT phosphorylation, inhibition of ligand binding and cell viability. The characteristics of the EM1-mAb was compared to control monovalent EGFR- or c-Met binding antibodies, and to known EGFR inhibitors such as erlotinib (CAS 183321-74-6; tyrosine kinase inhibitor) and cetuximab (CAS 205923-56-4).
[0647] As the parent antibodies of the EM-1 mAb antibodies are bivalent, control monovalent EGFR and c-Met antibodies were generated in a bispecific format combined with a Fab arm that binds to an unrelated/irrelevant antigen to accurately compare the synergy and avidity of a bispecific EM-1 mAb in comparison to a mixture of corresponding control monovalent molecules.
[0648] To generate the control monovalent EGFR and c-Met antibodies, a monospecific anti-HIV gp120 antibody gp120-K409R was generated comprising heavy chain of SEQ ID NO: 198 and a light chain of SEQ ID NO: 209. A monospecific anti-HIV gp120 antibody gp120-F405L was generated comprising the heavy chain of SEQ ID NO: 197 and the light chain of SEQ ID NO: 209.
[0649] The control monovalent anti-EGFR mAb E1-F405L-gp120-K409R was generated by in vitro Fab arm exchange between E1-F405L and gp120-K409R, and the control monovalent anti-cMet mAb M1-K409R-gp120-F405L was generated by in vitro Fab-arm exchange between M1-K409R and gp120-F405L and purified as described earlier.
[0650] The following cell lines were used in characterization of the bispecific antibodies: NCI-H292 (American Type Culture Collection (ATCC), Cat. No. CRL-1848), NCI-H1975 (ATCC Cat. No. CRL-5908), SKMES-1 (ATCC Cat. No. HTB-58), A431 (ATCC Cat. No. CRL-1555), NCI-H441 (ATCC Cat. No. HTB-174), NCI-H3255 (DCTD tumor/cell line repository, NCI, Frederick, NCI-Navy Medical oncology Cell Line supplement. J Cell Biochem suppl 24, 1996; Tracy S. cancer Res 64:7241-4, 2004; Shimamura T. cancer Res 65:6401-8, 2005) and HCC-827 (ATCC Cat. No. CRL-2868). NCI-H292 and SKMES-1 cells express both wild type EGFR and wild type c-Met. NCI-3255 expresses mutant L858R EGFR and displays EGFR and c-Met amplification. H1975 expresses mutant L858R/T790M EGFR and wild type c-Met. HCC-827 expresses A (E746, A750) EGFR and displays EGFR amplification. Cell line NCI-H292, NCI-H975, NCI-H441 and NCI-H3255 are interchangeably referred to as H292, H975, H441 and H3255, respectively, in the specification.
Binding of Bispecific EGFR/cMet Antibodies to EGFR and c-Met on Cells (A431 Cell Binding Assay)
[0651] The bispecific EGFR/c-Met antibody EM1-mAb was tested for binding to EGFR and c-Met on cells using protocol described in Example 3 ("A431 Cell Binding Assay") and Example 6 ("H441 Cell Binding Assay"). Cetuximab and a control antibody monovalent towards EGFR E1-F405L-gp120-K409R were used as controls for the A431 cells. Cetuximab had an EC.sub.50 value of 5.0 nM. Table 18 shows the EC.sub.50 values for binding. EM1-mAb demonstrated a 1.9-fold (A431 cells) and 2.3-fold (H441 cells) decrease in binding when compared to the bivalent monospecific parental control antibodies. Cetuximab was comparable to the bivalent parental control antibodies. EM1-mAb displays higher EC.sub.50 binding values than the values for the parental mAbs due to the monovalent binding of EGFR and c-Met. EM1-mAb has similar binding EC.sub.50 values as the single arm E1/inert arm and E2/inert arm bispecific monovalent mAbs.
TABLE-US-00028 TABLE 18 EC.sub.50 (nM) binding to cells E1-F405L-gp120-K409R (A431 cells) or M1- Parental K409R-gp120-F405L EM1-mAb mAbs (H441 cells) A431 (assay for 9.6 .+-. 3 5.1 .+-. 0.3 10.1 .+-. 0.6 EGFR binding) H441 (assay for c- 1.5 .+-. 0.7 0.65 .+-. 0.1 1.0 .+-. 0.3 Met binding)
Inhibition of Ligand Binding to the Receptor
[0652] The bispecific antibodies were tested for their ability to block binding of EGF to EGFR extracellular domain and HGF to c-Met extracellular domain in an ELISA assay. Recombinant human EGF R-Fc (R&D Systems, Cat #: 344-ER-050) or human HGF (R&D Systems, Cat #: 294-HGN-025/CF) was coated onto MSD HighBind plates (Meso Scale Discovery, Gaithersburg, Md.) for 2 hr at room temperature. MSD Blocker A buffer (Meso Scale Discovery, Gaithersburg, Md.) was added to each well and incubated for 2 hr at room temperature. Plates were washed three times with 0.1 M HEPES buffer, pH 7.4, followed by the addition of a mixture of either flurescently labeled (MSD) EGF or biotinylated HGF proteins with different concentrations of antibodies. Ruthenium-labeled EGF protein was incubated for 30 min at RT with increasing concentrations of different antibodies, from 1 nM to 4 .mu.M. After 2-hour incubation with gentle shaking at room temperature, the plates were washed 3 times with 0.1M HEPES buffer (pH 7.4). MSD Read Buffer T was diluted and dispensed and the signals were analyzed with a SECTOR Imager 6000. The HGF inhibition assays were performed as the EGF/EGFR inhibition assays except that 10 nM of biotinylated HGF was incubated for 30 min at RT with increasing concentrations of different antibodies, from 1 nM to 2 .mu.M.
[0653] EM1-mAb inhibited EGF binding to EGFR with an IC.sub.50 value of 10.7 nM+1.2 in the presence of 50 nM EGF and with an IC.sub.50 value of 10.6.+-.1.5 nM in the presence of 80 nM EGF. The parental bivalent antibody inhibited EGF binding to EGFR with an IC.sub.50 value of 0.14.+-.1.5 nM in the presence of 50 nM EGF and with an IC.sub.50 value of 1.7.+-.1.4 nM in the presence of 80 nM EGF. EM1 mAb had a weaker inhibition of EGF binding to the EGFR extracellular domain because of the monovalent binding of EM1 mAb as compared to the parental bivalent mAb.
[0654] EM1-mAb inhibited HGF binding to c-Met with an IC.sub.50 value of 29.9.+-.1.5 nM. The parental bivalent antibody inhibited HGF binding to c-Met with and IC.sub.50 of 14.8.+-.1.6 nM. EM1 mAb had a weaker inhibition of HGF binding to the cMet extracellular domain because of the monovalent binding of EM1-mAb as compared to the parental bivalent mAb.
Inhibition of EGF-Stimulated EGFR Phosphorylation and HGF-Stimulated c-Met Phosphorylation
[0655] Antibodies were tested to determine IC.sub.50 values for inhibition of EGFR and c-Met phosphorylation. Inhibition of EGF-stimulated EGFR phosphorylation and HGF-stimulated c-Met phosphorylation were assessed at varying antibody concentrations (0.035-700 nM final) as described in Example 2 ("Inhibition of EGF-Stimulated EGFR Phosphorylation") and Example 6 ("Inhibition of HGF-Stimulated c-Met Phosphorylation"). In some experiments, both EGF and HGF were added to the cells so the same cell lysate could be used to detect both EGFR and c-Met phosphorylation.
[0656] The control anti-EGFR mAb E1-F405L-gp120-K409R monovalent for EGFR and the parental bivalent anti-EGFR antibody with low fucose content were used as control antibodies. Table 19 shows the IC.sub.50 values of the assays.
TABLE-US-00029 TABLE 19 pEGFR (IC.sub.50, nM) pMet (IC.sub.50, nM) Cell line H292 H1975 H292 H1975 Antibody EM1-mAb 8.6-29 1.5 0.55-0.83 0.64 E1-F405L-gp120-K409R 10.9-13.1 ND 0.7-4 ND Parental EGFR 1.5 ND No effect ND (F405L)mAb* *Antibody had low fucose content
Enhanced Inhibition of pERK and pAKT with EM1-mAb Compared to Mixture of Monovalent Antibodies (mAb pERK Assay) (mAb pAKT Assay)
[0657] The potential for enhanced potency with a bispecific EGFR/c-Met antibody was evaluated by assessing mAb effects on pERK and pAKT downstream signaling. For these experiments, the mixture of monovalent control EGFR and monovalent control c-Met antibodies was compared to the bispecific EM1-mAb. Cells were plated in clear 96-well tissue culture-treated plates (Nunc) in 100 .mu.L/well of RPMI medium containing GlutaMAX and 25 mM Hepes (Invitrogen), supplemented with 1 mM sodium pyruvate (Gibco), 0.1 mM NEAA (Gibco), 10% heat inactivated fetal bovine serum (Gibco), and 7.5 ng/mL HGF (R&D Systems cat #294-HGN) and allowed to attach overnight at 37.degree. C. in a humidified 5% CO.sub.2 atmosphere. Cells were not serum-starved. Cells were treated for 30 mM (pERK assay) or 1 hour (pAkt assay) with varying concentrations (0.11-700 nM final) of monovalent control antibodies or EM1-mAb.
[0658] Cells were assessed for pERK or pAKT levels using the following kits and according to manufacturer's instructions from Meso Scale Discovery: Phospho-ERK1/2 (Thr202/Tyr204; Thr185/Tyr187) Assay Whole Cell Lysate Kit (cat # K151DWD, Phospho-Akt (Ser473) Assay Whole Cell Lysate Kit (cat # K151CAD), Phospho-Akt (Thr308) Assay Whole Cell Lysate Kit (cat # K151DYD). For the pERK assay, cells were lysed, and whole cell lysates were added to plates coated with anti-phospho-ERK1/2 antibody (recognizing ERK1 phosphorylated at residues Thr202 and Tyr204 and ERK2 phosphorylated at residues Thr185 and Tyr187), and phosphorylated ERK1/2 was detected with anti-total ERK1/2 antibody conjugated with MSD SULFO-TAG.TM. reagent. For the pAKT Ser473 assay, the capture antibody was anti-totalAKT antibody and the detection antibody anti-pAKT Ser473 antibody conjugated with MSD SULFO-TAG.TM. reagent. For the pAKT Thr308 assay, the capture antibody was anti-totalAKT antibody and the detection antibody anti-pAKT Thr308 antibody conjugated with MSD SULFO-TAG.TM. reagent.
[0659] Plates were read on a SECTOR.RTM. Imager 6000 instrument (Meso Scale Discovery) using manufacturer-installed assay-specific default settings. Data were plotted as electrochemiluminescence signal against the logarithm of antibody concentration and IC.sub.50 values were determined by fitting the data to a sigmoidal dose response with variable slope using GraphPad Prism 5 software. NCI-H292, H1975 and SKMES-1 cell lines were used in these assays.
[0660] The IC.sub.50 for inhibition of ERK phosphorylation by the bispecific EM1-mAb was about 14-63 fold lower relative to the mixture of the two monovalent control antibodies, depending on a cell line tested (Table 20). The improved potency of the EM1-mAb compared to the mixture of two monovalent control antibodies suggests a cooperative or avidity effect due to enhanced binding of EM1-mAb to these cell lines. The IC.sub.50 for inhibition of Ser475 (pAKTS475) and Thr308 (pAKTT308) AKT phosphorylation in NCI-H1975 cell line was about 75-fold and 122-fold lower, respectively, when compared to the mixture of the two monovalent control antibodies (Table 21). The improved potency of the EM1-mAb compared to the mixture of two monovalent control antibodies suggests a cooperative or avidity effect due to enhanced binding of EM1-mAb to these cell lines. Thus, the bispecific nature of the EM1-mAb resulted in an enhanced effect on downstream signaling effectors.
TABLE-US-00030 TABLE 20 IC.sub.50 (nM) pERK Antibody Fold change Mixture of E1- bispecific vs. F405L-gp120- mixture of two K409R and M1- monovalent Bispecific K409R-gp120- control Cell line EM1-mAb F405L antibodies H292 0.64 34.94 55 H1975 1.67 106 63 SKMES-1 0.54 7.63 14
TABLE-US-00031 TABLE 21 IC.sub.50 (nM) IC.sub.50 (nM) Antibody pAKTS473 pAKTT308 Bispecific EM1-mAb 0.87 0.96 Mixture of E1-F405L-gp120- 65 117 K409R and M1-K409R-gp120- F405L Fold change mixture of two 75 122 monovalent vs. bispecific
Inhibition of Human Tumor Cell Growth or Viability by Antibodies
[0661] Inhibition of c-Met-dependent cell growth was assessed by measuring viability of various tumor cells following exposure to the bispecific EM1-mAb. NCI-H292, SKMES-1, NCI-H1975 and NCI-H3255 cells were used in the studies.
[0662] Cells were cultured in standard 2D and low attachment formats. Erlotinib and cetuximab were used as controls. Table 22 summarizes the IC.sub.50 values for the assay.
Inhibition of Human Tumor Cell Growth or Viability by Antibodies--Standard 2D Format
[0663] The inhibition of cell growth was assessed by measuring the viability of NCI-H292 and NCI-H1975 following exposure to antibodies in two formats. For the standard 2D format cells were plated in opaque white 96-well tissue culture-treated plates (PerkinElmer) in RPMI medium containing GlutaMAX and 25 mM Hepes (Invitrogen), supplemented with 1 mM sodium pyruvate (Gibco), 0.1 mM NEAA (Gibco), and 10% heat inactivated fetal bovine serum (Gibco), and allowed to attach overnight at 37.degree. C., 5% CO.sub.2. Cells were treated with varying concentrations of antibodies (0.035-700 nM final), along with HGF (7.5 ng/mL, R&D Systems cat #294-HGF), then incubated at 37.degree. C., 5% CO.sub.2 for 72 hours. Some wells were left untreated with either HGF or antibodies as controls. Viable cells were detected using CellTiter-Glo.RTM. reagent (Promega), and data were analyzed as described in Example 3 in "Inhibition of Human Tumor Cell Growth (NCI-H292 growth and NCI-H322 growth assay)".
Inhibition of Human Tumor Cell Growth or Viability by Antibodies--Low Attachment Format
[0664] To assess survival in low attachment conditions, cells were plated in Ultra Low Attachment 96-well plates (Corning Costar) in 50 .mu.L/well of RPMI medium (Invitrogen) containing GlutaMAX and 25 mM Hepes, supplemented with 1 mM sodium pyruvate (Gibco), 0.1 mM NEAA (Gibco), and 10% heat inactivated fetal bovine serum (Gibco), and allowed to attach overnight at 37.degree. C., 5% CO.sub.2. Cells were treated with varying concentrations of antibodies (0.035-700 nM final), along with HGF (7.5 ng/mL, R&D Systems cat #294-HGN), then incubated at 37.degree. C., 5% CO.sub.2 for 72 hours. Some wells were left untreated with either HGF or antibodies as controls. Viable cells were detected using CellTiter-Glo.RTM. reagent (Promega), and data were analyzed as described above in "Inhibition of Human Tumor Cell Growth (NCI-H292 growth and NCI-H322 growth assay)" in Example 3, except that lysates were transferred to opaque white 96-well tissue culture-treated plates (PerkinElmer) prior to reading luminescence.
[0665] In the standard 2D culture, EM1-mAb inhibited NCI-H292 growth with an IC.sub.50 of 31 nM, and in low attachment conditions with an IC.sub.50 of 0.64 nM. EM-1 mAb inhibited NCI-H1975 cell growth with an IC.sub.50 of >700 nM and 0.8-1.35 nM in standard 2D and low attachment culture, respectively. In NCI-H292 cells expressing both wild type EGFR and cMet, EM1-mAb had over 22 fold improved potency in the standard 2D and about 330-fold improved potency in the low attachment culture conditions when compared to cetuximab. In NCI-H1975 cell, which express L858R, T790M EGFR mutant and wild type cMet, EM-1 mAb had at least a 518-fold improved potency when compared to cetuximab in low attachment culture conditions. Table 22 shows the summary of the assays.
TABLE-US-00032 TABLE 22 EM1-mAb EM1-mAb Cetuximab Cetuximab Standard Low Standard Low 2D culture attachment 2D culture attachment Cell line EGFR state cMet state IC.sub.50 (nM) IC.sub.50 (nM) IC.sub.50 (nM) IC.sub.50 (nM) NCI-H292 WT WT 31 0.64 >700 212 NCI-H1975 L858R, T790M WT >700 0.8-1.35 >700 >700
Combination of Erlotinib and EM1-mAb is Efficient in Inhibition of Growth of EGFR Mutant Cell Lines
[0666] The inhibition of cell growth by the combination of erlotinib plus EM1-mAb was evaluated in both standard 2D culture conditions and in the low attachment format. NCI-H3255 and HCC-827 cells were plated as described above in "Inhibition of Human Tumor Cell Growth or Viability by Antibodies". HGF (7.5 ng/mL, R&D Systems cat #294-HGN) was added to cells along with treatment with antibodies. Cells were treated with varying concentrations of antibodies (0.11-700 nM final), or erlotinib (0.46-3000 nM final), or the combination of erlotinib plus antibody, using increasing amounts of each in a fixed ratio (e.g. lowest concentration of combination=lowest concentration of antibody (0.11 nM)+lowest concentration of erlotinib (0.46 nM)). Some wells were left untreated with either HGF or antibodies as controls. Cells were incubated at 37.degree. C., 5% CO.sub.2 for 72 hours, then viable cells were detected using CellTiter-Glo.RTM. reagent (Promega), and data were analyzed as described above in "Inhibition of Human Tumor Cell Growth (NCI-H292 growth and NCI-H322 growth assay)". Table 23 summarizes the results of the experiment. In the table, the IC.sub.50 values for the combinations are relative to either the antibody, or erlotinib, depending on what is indicated in parentheses.
[0667] In both NCI-H3255 and HCC-827 cells (EGFR mutant cell lines) the addition of EM1-mAb to erlotinib both increased the potency of inhibition of cell viability and was more effective resulting in fewer viable cells overall. In the NCI-H3255 cells using standard 2D conditions, the IC.sub.50 for erlotinib alone was 122 nM, whereas the combination was 49 nM. Similarly, in HCC-827 cells, the IC.sub.50 for erlotinib alone was 27 nM, whereas the combination was 15 nM. Also, the combination of erlotinib plus EM1-mAb was more effective than the combination of erlotinib plus cetuximab. Thus, in the presence of HGF, addition of EM1-mAb increased the effectiveness of erlotinib in this assay.
[0668] NCI-H3255 cells express L858R mutant EGFR and amplified cMet. HCC-827 cells express EGFR mutants with deletions at amino acid positions 746 and 750 and wild type c-Met. EM1-mAb has stronger effects in the viability of HCC-827 and NCI-3255 in the presence of erlotinib than erlotinib alone in either standard or low attachment cultures.
TABLE-US-00033 TABLE 23 EM1 mAb + erlotinib Samples erlotinib IC.sub.50 (nM) IC.sub.50 (nM) NCI-H3255, standard 2D culture 49.0 122 NCI-H3255, low attachment culture 10.6 47.1 HCC-827, standard 2D culture 14.6 27.4 HCC-827, low attachment culture 3.5 9.5
Example 13. Antibody Mediated Cellular Cytotoxicity (ADCC) of EM1-mAb in In Vitro Cell Lines
[0669] ADCC assays were performed as previously described (Scallon et al., Mol Immunol 44:1524-1534 2007). Briefly, PBMCs were purified from human blood by Ficoll gradients and used as effector cells for ADCC assays. NCI-H292, NCI-H1975 or NCI-H441 cells were used as target cells with a ratio of 1 target cell to 50 effector cells. Target cells were pre-labeled with BATDA (PerkinElmer) for 20 minutes at 37.degree. C., washed twice and resuspended in DMEM, 10% heat-inactivated FBS, 2 mM L-glutamine (all from Invitrogen). Target (1.times.10.sup.4 cells) and effector cells (0.5.times.10.sup.6 cells) were combined and 100 .mu.l of cells were added to the wells of 96-well U-bottom plates. An additional 100 .mu.l was added with or without wild type and protease-resistant mAb constructs. All samples were performed in duplicate. The plates were centrifuged at 200 g for 3 minutes, incubated at 37.degree. C. for 2 hours, and then centrifuged again at 200 g for 3 minutes. A total of 20 .mu.l of supernatant was removed per well and cell lysis was measured by the addition of 200 .mu.l of the DELPHIA Europium-based reagent (PerkinElmer). Fluorescence was measured using an Envision 2101 Multilabel Reader (PerkinElmer). Data were normalized to maximal cytotoxicity with 0.67% Triton X-100 (Sigma Aldrich) and minimal control determined by spontaneous release of BATDA from target cells in the absence of any antibody using the following equation: (experimental release-spontaneous release)/(maximal release-spontaneous release).times.100%. Data were fit to a sigmoidal dose-response model using GraphPad Prism v5.
[0670] The ADCC results for the EM1 mAbs and comparators are summarized in the Table 24 (NCI-H292 cells), Table 25 (NCI-H1975 cells) and Table 26 (NCI-H441 cells) and Table 27 (NCI-H1993 cells) list the EC.sub.50 values and maximum lysis achieved. NCI-H292 cells express wild type (WT) EGFR, WT c-Met, and WT KRAS; NCI-H1975 cells express mutant EGFR (L858R T790M), WT cMet and WT KRAS; NCI-H441 express WT EGFR, WT cMet, and mutant KRAS (G12V), and NCI-H1993 cells express WT EGFR, amplified cMet, WT KRAS. KRAS is also known as GTPase KRas and as V-Ki-ras2 Kirsten rat sarcoma viral oncogene homolog.
[0671] The EM1-mAb has higher potency of ADCC responses than cetuximab and the normal fucose version of EM1-mAb as indicated by having lower EC.sub.50 values. The EM1 mAb has higher efficacy in terms of maximum lysis achieved than cetuximab and the normal fucose bispecific mAb. From profiles of on Tables 24-27, the EM-1 mAb has ADCC activity on cells that have mutant and WT EGFR, WT with normal and amplified levels of cMet, and WT and mutant KRAS.
TABLE-US-00034 TABLE 24 Potency (EC.sub.50 Efficacy (maximum mAb .mu.g/ml) R.sup.2 lysis achieved) EM1 mAb 0.0058 0.93 19% Anti-EGFR .times. cMet 0.22 0.85 13% normal fucose bispecific mAb Cetuximab 0.0064 0.94 12%
TABLE-US-00035 TABLE 25 Potency (EC.sub.50 Efficacy (maximum mAb .mu.g/ml) R.sup.2 lysis achieved) EM1 mAb 0.022 0.91 19% Anti-EGFR .times. cMet 1.8 0.79 13% normal fucose bispecific mAb Cetuximab 0.029 0.70 11%
TABLE-US-00036 TABLE 26 Potency (EC.sub.50 Efficacy (maximum mAb .mu.g/ml) R.sup.2 lysis achieved) EM1 mAb 0.022 0.97 24% Anti-EGFR .times. cMet 0.52 0.87 7.9% normal fucose bispecific mAb Cetuximab 0.013 0.85 15%
TABLE-US-00037 TABLE 27 Potency (EC.sub.50 Efficacy (maximum mAb .mu.g/ml) R.sup.2 lysis achieved) EM1 mAb 0.0013 0.95 27% Anti-EGFR .times. cMet 0.054 0.87 17% normal fucose bispecific mAb Cetuximab 0.0042 0.76 21%
Example 14. Tumor Efficacy Studies with the EM1-mAb
[0672] The efficacy of the EM1 mAb against tumor growth was conducted as described in Example 7 "Tumor efficacy studies with bispecific EGFR/c-Met molecules". In brief, NCI-H292-HGF cells were implanted subcutaneously (s.c.) with Cultrex at 2.times.10.sup.6 into female SCID Beige mice. The mice were stratified by tumor volume 7 days after implant into 5 Groups with 10 mice per group. The dosing began after the starting mean tumor volume per group ranged from 62-66 mm.sup.3 (small tumors). PBS or therapeutic were dosed intraperitoneally (i.p.) 2 times per week.
[0673] The evaluation of the efficacy also employed SKMES-HGF, a human squamous cell carcinoma that was transfected with human HGF (hepatic growth factor). These cells were implanted s.c. at 10.times.10.sup.6 into female SCID Beige mice. These mice were stratified by tumor volume 12 days after implant into 5 groups with 8 mice per group. The first study began with starting mean tumor volume per group ranged from 98-101 mm.sup.3 (large tumors). PBS or therapeutic mAbs were--dosed i.p. 2.times./week for 4 weeks. In the larger sized tumor study, the mice that were stratified after the tumor volumes were about 200-300 mm.sup.3 by splitting into 2 groups. These mice were then treated with either cetuximab (20 mg/kg) or EM1-mAb (20 mg/kg), i.p., 2.times./week (3 weeks).
[0674] The summary of the data is shown in Table 28. FIG. 10 shows the efficacy of the molecules over time. EM1-mAb has an improved tumor suppression profile when compared to cetuximab in H292-HGF small tumor model and in SKMES-HGF small and large tumor models.
TABLE-US-00038 TABLE 28 Dosing at Partial Complete Sample and time Cell line mg per kg regression regression EM1 at day 35 H292-HGF small 20 10/10 10/10 tumor 5 10/10 10/10 1 0/10 0/10 Cetuximab at day H292-HGF small 20 0/10 0/10 35 tumor EM1 at day 67 SKMES-HGF 20 0/8 8/8 small tumor 5 1/8 6/8 1 2/8 4/8 Cetuximab at day 20 0/8 6/8 67 EM1 at day 70 SKMES-HGF 20 4/7 3/7 Cetuximab at day large tumor 20 0/7 0/7 35
Table 29 shows the tumor sizes in treatment groups from the SKMES-HGF tumors, and table 30 shows the anti-tumor activity.
[0675] EM1-mAb inhibited tumor growth in the SKMES-HGF model 97% or more at multiple doses down to 1 mg/kg. While initially cetuximab was very effective (88% TGI at 20 mg/kg), after dosing ended the cetuximab-treated tumors grew back. In contrast, the tumors treated with EM1-mAb at either 5 or 20 mg/kg did not grow back over the course of the study (>2 months).
TABLE-US-00039 TABLE 29 Tumor volume (mm.sup.3) bispecific bispecific bispecific Cetuximab EM1 at EM1 at EM1 at at Days Vehicle 20 mg/kg 5 mg/kg 1 mg/kg 20 mg/kg 1 99 .+-. 6 99 .+-. 7 101 .+-. 6 101 .+-. 6 98 .+-. 5 8 146 .+-. 14 48 .+-. 10 49 .+-. 9 49 .+-. 10 60 .+-. 8 15 192 .+-. 21 9 .+-. 1 22 .+-. 10 41 .+-. 13 44 .+-. 23 22 326 .+-. 43 3 .+-. 2 17 .+-. 12 33 .+-. 15 42 .+-. 23 29 577 .+-. 55 2 .+-. 1 15 .+-. 9 38 .+-. 17 85 .+-. 60 36 994 .+-. 114 0.2 .+-. 0.1 13 .+-. 9 26 .+-. 14 125 .+-. 62 50 -- 0.04 .+-. 0.04 10 .+-. 7 18 .+-. 9 423 .+-. 115 57 -- 0.1 .+-. 0.2 3 .+-. 2 21 .+-. 10 650 .+-. 116 67 -- 0 .+-. 0 8 .+-. 7 34 .+-. 22 1257 .+-. 151
TABLE-US-00040 TABLE 30 Tumor T-C Size (days) at (mm.sup.3)a T/C 1000 P Treatment at day 36 (%) mm.sup.3 value Vehicle 994 .+-. 114 -- -- -- bispecific EM1 at 0.19 .+-. 0.12 0.02 -- 20 mg/kg bispecific EM1 at 13 .+-. 9 1.3 -- 5 mg/kg bispecific EM1 at 26 .+-. 14 2.6 -- 1 mg/kg Cetuximab 125 .+-. 62 13 31 (20 mg/kg)
Example 15. Inhibition of Cell Migration with EM1-mAb In Vitro Method
[0676] Effect of the EM-mAb and the control monovalent antibodies on inhibition of tumor cell migration was assessed in NIH-1650 cells. EGFR mutant cell line H1650 (Lung Bronchioloalveolar carcinoma cells harboring an exon 19 mutation [deletion E746, A750]) was cultured in tissue culture flasks under normal culture conditions (37.degree. C., 5% CO.sub.2, 95% humidity). All media and supplementation were as suggested by the supplier of the cells (American Type Culture Collection, Manassas, Va., USA).
[0677] Spheroids were generated by plating H1650 lung tumor cells at 10,000 cells/well into "U" bottom Ultra Low Adherence (ULA) 96-well plates (Corning, Tewksbury, USA) at 200 .mu.l/well. These plates stimulate spontaneous formation of a single spheroid of cells within 24 hours (upon incubation at 37.degree. C., 5% CO.sub.2) and the spheroids were grown for four days under normal culture conditions.
[0678] Round bottom 96-well plates (BD Bioscience) were coated with 0.1% gelatin (EMD Millipore, Billerica, USA) in sterile water for 1 h at 37.degree. C. For compound evaluation studies, day 4 10,000 cell tumor spheroids (H1650 and NCI-H1975) were transferred to the coated round bottom plates and treated with the EM1-mAb, the control monovalent anti-EGFR mAb E1-F405L-gp120-K409R having low fucose content, the control monovalent anti-cMet mAb M1-K409R-gp120-F405L having low fucose content, and a combination of the two monovalent antibodies E1-F405L-gp120-K409R and M1-K409R-gp120-F405L (produced in low fucose) in a dilution series with 20 ng/ml of HGF (R&D systems). Controls were treated with vehicle which was IgG.sub.1 kappa isotype control (concentration equal to highest drug-treated cells). Effects of compounds were analyzed at 48 hrs by measuring the area covered by migrating cells using bright field images in a fully automated Operetta high content imaging system (Perkin Elmer) with a 2.times. objective. Inhibition of cell migration (total area) due to treatment effect was assessed by normalizing data by dividing by media only control to create a percentage cell migration to control. Thus, a value less than 1 would be inhibitory to cell migration.
Results
[0679] The EM1-mAb demonstrated potent synergistic inhibition of cell migration in H1650 (L858R EGFR mutant) and H1975 (L858R/T790M EGFR mutant) cells when compared to a combination of the control monovalent anti-EGRF and anti-c-Met antibodies E1-F405L-gp120-K409R and M1-K409R-gp120-F405L. In H1650 cells, the six highest concentrations of the EM1-mAb significantly inhibited cell migration (p<0.001) compared to the isotype control. The EC.sub.50 value for the EM1-mAb was 0.23 nM, whereas the EC.sub.50 value for the combination of the monospecific control antibodies was 4.39 nM. The EM1-mAb therefore was about 19 fold more efficient in inhibiting H1650 cell migration when compared to the combination of the monovalent control antibodies. The level of cell migration inhibition of EM1-mAb was superior to the combination of monospecific control mAbs for H1650 and H1975 cells. Table 31 shows the EC.sub.50 values for the assay.
TABLE-US-00041 TABLE 31 H1650 H1975 EC.sub.50 Inhibition at Inhibition at Samples (nM) 30 nM 30 nM EM1-mAb 0.23 64% 38% Mixture of E1-F405L- 4.39 59% 20% gp120-K409R* and M1-K409R-gp120-F405L* E1-F405L-gp120-K409R* 5.44 15% 7% M1-K409R-gp120-F405L* 7.36 43 10% *antibodies have low fucose content
Example 16. Epitope Mapping of Anti-c-Met Antibody 069 and 5D5
[0680] The anti-c-Met mAb 069 binding epitope was mapped using the linear and constrained CLIPS peptide technology. The peptides scanned the SEMA, PSI, and Ig domains of human cMet. The linear and CLIPS peptides were synthesized using the amino acid sequence of the aforementioned cMet using standard Fmoc chemistry and deprotected using trifluoric acid with scavengers. The constrained peptides were synthesized on chemical scaffolds in order to reconstruct conformational epitopes using Chemically linked Peptides on Scaffolds (CLIPS) Technology (Timmerman et al., J Mol Recognition 20:283, 2007). The linear and constrained peptides were coupled to PEPSCAN cards and screened using a PEPSCAN based ELISA (Slootstra et al., Molecular Diversity 1, 87-96, 1996). The anti-c-Met mab 069 binding epitope is a discontinuous epitope consisting of c-Met amino acids 239-253 PEFRDSYPIKYVHAF (SEQ ID NO: 238) and 346-361 FAQSKPDSAEPMDRSA (SEQ ID NO: 239). c-Met amino acid sequence is shown in SEQ ID NO: 201.
[0681] Similar methods were used to map mAb 5D5 (MetMab, Onartuzumab) epitope. mAb 5D5 binds c-Met residues 325-340 PGAQLARQIGASLNDD (SEQ ID NO: 240).
Example 17. In Vivo Tumor Efficacy Studies with EM1-mAb
[0682] The efficacy of EM1 mAb against tumor growth was conducted as described in Example 7 "Tumor efficacy studies with bispecific EGFR/c-Met molecules" and Example 14 employing additional tumor cell lines with EGFR mutation or EGFR and/or c-Met amplifications. In brief, SNU-5, H1975, HCC827 cells, H1975 cells expressing human HGF, or a clone of HCC827 cells selected for its increased resistance to erlotinib (HCC827-ER1 cells) were implanted subcutaneously (s.c.) into female nude mice, except that SNU-5 cells were implanted in CR17/SCID mice. Mice were dosed intraperitoneally with PBS or EM1-mAb, cetuximab (CAS 205923-56-4), erlotinib (CAS 183321-74-6), afatinib (CAS 439081-18-2), or a combination of EM-1 mAb and afatinib and EM-1 mAb and erlotinib at indicated dosage and schedule shown in Table 32. Antitumor efficacy was measured as % TGI (tumor growth inhibition) calculated as 100-% T/C (T=mean tumor size of the treatment group; C=mean tumor size of the control group on a given day as described in Example 7).
[0683] In tumors with primary EGFR activating mutations (no resistance to EGFR TKIs): (HCC827 tumor, EGFR del(E746, A750)), EM1-mAb dosed 10 mg/kg inhibited tumor growth by 82%. Erlotinib was similarly effective in this model, as was the combination of erlotinib and EM1-mAb. FIG. 11 shows efficacy of the therapeutics over time in the HCC827 tumor model.
[0684] In tumors with wild type EGFR and c-Met gene amplification (gastric cancer model SNU-5), EM1-mAb showed antitumor activity with full tumor regression (98% TGI, at day 34 p<0.01, compared to vehicle using one-way ANOVA followed by individual comparisons using Games-Howell). Antitumor activity of anti-EGFR mAb cetuximab was less, 49% at day 34, in this model. FIG. 12 shows the efficacy of the therapeutics over time in the SNU-5 model.
[0685] EM1-mAb was tested in a NSCLC model containing primary EGFR activating mutation and the T790M EGFR mutation which renders tumors resistant to l' generation EGFR TKIs (H1975 model). EM1-mAb inhibited tumor growth with a 57% TGI in the H1975 cell line model implanted in nude mice (p<0.0001, compared to PBS vehicle using Logrank analysis with Prism 3.03). As expected, erlotinib was not effective in this model with the T790M mutation. Afatinib was equally effective as the EM1-mAb (57% TGI). Cetuximab and the combination of EM1-mAb with afatinib were the most effective, regressing tumors with 91% and 96% tumor growth inhibition, respectively, (p<0.0001 for both cetuximab compared to PBS and EM1-mAb+afatinib compared to the PBS+afatinib vehicles group using Logrank analysis with Prism 3.03). c-Met signaling pathways are not activated in this model as the mouse HGF does not bind to human c-Met.
[0686] EM1-mAb was tested in several models that were engineered to express human HGF using a lentiviral transduction system. This allows modeling of ligand activation of the c-Met pathway in vivo because mouse HGF does not activate the human c-Met on the implanted human tumor cells. Results with SKMES-HGF model are shown in Example 14 and FIG. 10, and the % TGI summarized in Table 32. EM1-mAb inhibited tumor growth in the H1975-HGF model 71% (p<0.0001, compared to PBS vehicle using Logrank analysis with Prism 3.03). Afatinib, erlotinib and cetuximab were less efficacious in this model. The combination of EM1-mAb and afatinib was very effective (96% TGI, p<0.0001, compared to the PBS+afatinib vehicles group using Logrank analysis with Prism 3.03). FIG. 13 shows the efficacy of the molecules over time in the H1975-HGF model. Erlotinib, afatinib and cetuximab thus lose their antitumor efficacy in tumor models in which c-Met pathway is activated.
[0687] EM1-mAb was tested in a tumor model characterized by primary EGFR activating mutation and increased resistance to 1.sup.st generation EGFR TKI (erlotinib) due to c-Met gene amplification (HCC827-ER1 model). EM1-mAb dosed at 10 mg/kg partially regressed HCC827-ER1 tumors implanted with 86% TGI at day 25, and was more efficacious than erlotinib alone (65% TGI at day 25). Combination of EM1-mAb and erlotinib did not further improve efficacy. FIG. 14 shows the efficacy of the molecules over time.
[0688] EM1-mAb thus demonstrates efficacy in tumor models with wild type EGFR, with primary activating EGFR mutations, with the EGFR mutation T790M associated with resistance to EGFR therapeutics, as well as in models where c-Met is activated in either a ligand-dependent (autocrine HGF expression) or ligand-independent (c-Met gene amplification) manner. Combination of EM1-mAb with erlotinib or afatinib may improve efficacy in some tumor models.
TABLE-US-00042 TABLE 32 % TGI (day Treatment of study); (dose in mg/kg), compared to Tumor Type EGFR cMet schedule PBS vehicle SKMES- WT WT EM1-mAb (20), 100 (36) HGF BIW .times. 4 wk lung cetuximab (20), 88 (36) squamous BIW .times. 4 wk SNU-5 WT AMP EM1-mAb (10), 98 (34) gastric BIW .times. 4 wk cetuximab (10), 49 (34) BIW .times. 4 wk H1975 L858R; WT EM1-mAb (10), 57 (18) NSCLC T790M BIW .times. 3 wk cetuximab (10), 91 (18) BIW .times. 3 wk erlotinib (50), 9 (18) QD .times. 21 d afatinib (15), 57 (18) QD .times. 21 d EM1-mAb (10), 96 (18) BIW .times. 3 wk + afatinib (15), QD .times. 21 d H1975- L858R; WT EM1-mAb (10), 71 (16) HGF T790M BIW .times. 3 wk NSCLC cetuximab (10), 42 (16) BIW .times. 3 wk erlotinib (50), 20 (16) QD .times. 21 d afatinib (15), 29 (16) QD .times. 21 d EM1-mAb (10), 96 (16) BIW .times. 3 wk + afatinib (15), QD .times. 21 d HCC827 del WT EM1-mAb (10), 82 (35) NSCLC (E746, BIW .times. 4 wk A750); erlotinib (25), 79 (35) AMP QD .times. 28 d EM1-mAb (10), 78 (35) BIW .times. 3 wk + erlotinib (25), QD .times. 28 d HCC827- del AMP EM1-mAb (10), 86 (25) ER1 (E746, BIW .times. 4 wk NSCLC A750); erlotinib (25), 65 (25) AMP QD .times. 28 d EM1-mAb (10), 87 (25) BIW .times. 3 wk + erlotinib (25), QD .times. 28 d BIW = biweekly QD = once per day WT = wild type AMP = amplified
Example 18. EM1 mAb Induced Degradation of EGFR and c-Met In Vivo
[0689] To demonstrate engagement of both EGFR and c-Met by EM1-mAb in the tumor, samples were taken from H1975-HGF tumors at various times after a single dose of 20 mg/kg EM1-mAb Tumor lysates were prepared, normalized to total protein, and samples run on SDS-PAGE gels. Gels were transferred to nitrocellulose and Western blotted for either EGFR (Mouse (mAb) Anti-human EGFR (EGF-R2); Santa Cruz Biotechnology, Cat # sc-73511) or c-Met (Mouse (mAb) Anti-human Met (L41G3); Cell Signaling Technology, Cat #3148). EGFR levels were normalized to GAPDH; c-Met levels were normalized to actin. The levels of receptors from EM1-mAb treated tumors were compared to those of PBS-treated tumors to get % total receptor. EM1-mAb treatment decreased the total EGFR and cMet receptor levels in H1975-HGF tumors to between 20% to 60% of control, depending on the time point analyzed. FIG. 15 shows the average receptor levels compared to PBS over time. pEGFR, pc-Met and pAKT were also decreased at 72 hours after the single dose of EM1.
Example 19. Anti-Tumor Activity Comparing IgG.sub.1 and IgG.sub.2.sigma. Variant Isoforms of EGFR/c-Met Bispecific Antibodies
[0690] To better understand the contribution of effector function to the efficacy observed in the H1975-HGF model, a comparison was performed between EM1-mAb and a variant of EM1-mAb having an IgG2 Fc with effector silencing substitutions V234A/G237A/P238S/H268AN309L/A330S/P331S on IgG2 (substitutions described in Intl. Pat. Appl. No. WO2011/066501) (numbering according to the EU index). An IgG2 antibody with V234A/G237A/P238S/H268A/V309L/A330S/P331S substitutions does not interact with Fc receptors or effector cells (such as NK cells and macrophages). Any loss of activity observed with the IgG2 V234A/G237A/P238S/H268A/V309L/A330S/P331S variant of the EM1-mAb may thus represent antitumor activity contributed by effector-mediated mechanisms such as ADCC and/or ADCP. After 32 day post tumor cell implant in the H1975-HGF model described above, there is an indication of loss of antitumor activity with the IgG2 V234A/G237A/P238S/H268A/V309L/A330S/P331S variant of the EM1-mAb when compared to the parental EM1-mAb, suggesting that effector-mediated mechanisms contribute to the function of EM-1 mAb. FIG. 16 shows the antitumor activity of the molecules.
Example 20. EGFR and c-Met Receptor Densities on Cells Correlate with EM1-mAb Efficacy In Vitro
[0691] The role of EGFR and c-Met cell-surface densities in in vitro efficacy of EM-1 mAb, particularly in the context of heterogeneous cancer cell populations, was studied. In vitro efficacy of EM-1 mAb was assessed by its ability to inhibit EGFR and/or c-Met receptor phosphorylation as described in Example 12 "Inhibition of EGF-Stimulated EGFR Phosphorylation and HGF-Stimulated c-Met Phosphorylation".
[0692] EGFR and c-Met expression levels were quantified in a panel of tumor cell lines using Quantitative Flow Cytometry (QFCM) using the control monovalent antibodies E1-F405L-gp120-K409R or M1-K409R-gp120-F405L, EM-1 mAb or commercially available anti-EGFR and c-Met antibodies. Tumor cell lines and their EGFR and c-Met genotypes used in the study are shown in Table 33.
TABLE-US-00043 TABLE 33 EGFR cMet Cell line Origin Genotype Amplified* Genotype Amplified* H292 Lung WT N WT N mucoepidermoid SKMES-1 Lung squamous WT N WT N HCC827 Lung adeno del (E746, A750) Y (19) WT N H1975 Lung adeno L858R, T790M N WT N H3255 Lung adeno L858R Y (12) WT N H1650 Lung del (E746, A750) Y (4) WT N Bronchioloalveolar HCC4006 Lung adeno del (L747, S752) Y (5) WT N HCC2935 Lung adeno del (E746, N WT N A750) H820 Lung adeno del (E746, UNK WT Y (4-6) A750), T790M H1993 Lung adeno WT UNK WT Y (UNK) SNU-5 Stomach adeno WT N WT Y (9) WT: wild-type; del: deletion mutant; Lung adeno: Lung adenocarcinoma, UNK: Unknown, Y: Yes, N: No *Value in parenthesis indicates the number of gene copies per cell
Cell Culture
[0693] Tumor cell lines were cultured in 150 cm.sup.2 tissue culture flasks under standard culture conditions (37.degree. C., 5% CO.sub.2, 95% humidity). Media formulations used were as recommended by the supplier of the cells (ATCC, Manassas, Va., USA). All cells were supplemented with 0.1 mM Non-Essential Amino Acids (Life Technologies) and 1 mM Sodium Pyruvate (Life Technologies) unless otherwise indicated. Media were routinely changed two to three times weekly. When subconfluent, cell monolayers were passaged using Accutase (Sigma-Aldrich, St Louis, USA). Cells were harvested for receptor quantitation at approximately 80% confluence and passage number was never allowed to exceed p15. To dissociate cells for quantitation analysis, enzyme-free Cellstripper (Corning) was used to avoid proteolysis of surface antigens.
Conjugation of Antibodies with R-Phycoerythrin (RPE)
[0694] Antibodies were conjugated to RPE (Prozyme) using heterobifunctional chemistry. The R-PE was activated using sulfo-SMCC (Pierce) for 60 minutes. Activated R-PE was separated from free sulfo-SMCC by gel filtration chromatography. The antibodies were reduced using dithiothreitol (DTT, Sigma) for 30 minutes and separated from free DTT by gel filtration chromatography. The activated R-PE was covalently coupled to the reduced antibodies for 90 minutes. The reaction was quenched with N-Ethylmaleimide (Fluka) for 20 minutes. The RPE-conjugated antibodies were purified using a size-exclusion Tosoh TSKgel G3000SW column in 100 mM sodium phosphate, 100 mM sodium sulfate, 0.05% sodium azide, pH 6.5 on an AKTA explorer FPLC system. The RPE conjugated antibodies were stored at 2-8.degree. C. in light occlusive containers. For analytical size-exclusion chromatography the pooled fractions were applied to a Tosoh TSKgel G3000SW.times.1 column in 100 mM sodium phosphate, 100 mM sodium sulfate, 0.05% sodium azide, pH6.5. The samples were quantified by UV absorbance at 280 nm.
Flow Cytometry Staining
[0695] Immediately following dissociation the cells were assessed for viability by exclusion of 0.4% Trypan Blue (Life Technologies, Carlsbad, USA). Cell number and viability were calculated using a C-chip hemocytometer (Incyto, Covington, USA). Cells were resuspended at 1.times.10.sup.6 cells/mL in BSA Stain Buffer (BD Biosciences, San Jose, USA), at which point strict adherence to 4.degree. C. was followed to minimize receptor internalization. Fixation of cells was avoided due to the potential effects of paraformaldehyde on antibody dissociation rates and/or epitope integrity. FcRs (though not present on these cells) were blocked with 5 .mu.l per test of Human TruStain FcX.TM. (BioLegend, San Diego, USA) for 30 minutes at 4.degree. C. 1.times.10.sup.5 cells per well were transferred to 96-well V-bottom microplates (Greiner, Monroe, USA). Cells were incubated on ice for 1-2 hours with serial dilutions of either in-house R-phycoerythrin labeled control monovalent anti-EGFR mAb E1-F405L-gp120-K409R or control monovalent anti-c-Met mAb M1-K409R-gp120-F405L or the bispecific EM1 mAb at empirically determined saturating concentrations. Commercial antibodies used in these studies included RPE-conjugated anti-human HGFR/cMet (R&D systems, Minneapolis, USA), and anti-human EGFR (BioLegend, San Diego, USA). Isotype matched controls included RPE-conjugated mouse IgG1 Isotype control, (R&D systems, Minneapolis, USA) and mouse IgG2bk isotype control (BioLegend, San Diego, USA). These were included at the highest concentration used for the respective receptor specific mAb. Cells were washed two times with 150 .mu.L of BSA Stain Buffer and resuspended in 250 .mu.L of BSA Stain Buffer containing 1:50 diluted DRAQ7 live/dead stain (Cell Signaling Technology, Danvers, USA). Single stain controls for RPE and DRAQ7 were included. Samples were read on either a BD FACSCalibur (BD Biosciences) or Miltenyi MACSQuant flow cytometer (Miltenyi Biotec, Auburn, USA). The data collection channels for PE/DRAQ7 were: FL2/FL4 (FACSCalibur) and B2/B4 (MACSQuant). Live cells were gated according to DRAQ7 exclusion and the geometric mean fluorescence intensity was determined for >5,000 live events, collected at a low flow rate. FlowJo vX software (FlowJo, Ashland, USA) was used for analysis.
Determination of Antibody Binding Capacity (ABC)
[0696] Receptor density values were reported as antibodies bound per cell (ABC). ABC values were derived from standard curves generated with commercial microspheres coated with defined amounts of a spectrally-matched fluorochrome, QuantiBRITE.TM. PE Beads.
[0697] To calculate ABC values, geometric means for the four QuantiBRITE singlet bead populations were derived in FlowJo. Linear regression was used to create a standard curve in GraphPad Prism (GraphPad Software, La Jolla, USA) of the log [# of RPE molecules/bead] versus the log[MFI]. R.sup.2 values were typically .gtoreq.0.99, ABC values for the RPE-mAbs were then interpolated from the QuantiBRITE standard curve. ABC values presented here are the specific antibody binding sites (sABC) calculated by subtracting the ABC values from an anti-RSV isotype-matched control from the ABC of the EGFR or cMet mAbs.
[0698] The observed EGFR and c-Met densities using the antibodies E1-F405L-gp120-K409R or M1-K409R-gp120-F405L in a panel of tumor cell lines is shown in Table 34. The experiments were done in duplicates in cell lines where S.E.M. is indicated in table, and as single point in cell lines with no S.E.M. indicated.
TABLE-US-00044 TABLE 34 c-Met density EGFR density Cell Line Mean S.E.M. Mean S.E.M. H1975 90867 5649 65523 1954 HCC827 204217 9413 419194 5874 H1650 58618 5374 101889 n.d. H3255 126692 46221 714987 n.d. H820 192226 4738 111323 1409 HCC2935 44768 n.d. 78669 n.d. HCC4006 59616 n.d. 49339 n.d. H1993 564704 n.d. 340404 n.d. H292 63915 n.d. 365617 n.d. SKMES-1 47469 n.d. 174532 n.d. SNU-5 504140 n.d. 118721 n.d.
[0699] The IC.sub.50 values of the EM-1 mAb for inhibition of phosphorylation of EGFR and c-Met was determined as described in Example 12. The potency of inhibition of receptor phosphorylation by the EM-1 mAb was inversely proportional to the receptor density for both EGFR (FIG. 17A) and c-Met (FIG. 17B). The correlation of inhibition of receptor phosphorylation to receptor quantitation was statistically significant with Pearson correlation p=0.00082, (r.sup.2=0.7820) for EGFR and p<0.0001 (r.sup.2=0.9336) for c-Met.
Example 21. EM-1 Bispecific EGFR/c-Met mAb Heterodimerizes EGFR and c-Met on the Cell Surface
[0700] The effect of the EM-1 mAb on EGFR and c-Met heterodimerization was assessed as a mechanism for the observed synergistic inhibition with the mAb.
Heterodimerization Assay Using Enzyme Fragment Complementation
[0701] For the EGFR/c-Met heterodimerization assay, complementary inactive fragments of .beta.-galactosidase (.beta.-gal) were fused to the C-terminus of c-Met and EGFR. Stably transfected human osteosarcoma U-2 OS cell line (ATCC HTB-96.TM.) was generated expressing both constructs, and enzyme fragment complementation was carried out essentially as described in Wehrman et al., Proc Natl Acad Sci USA 103: 19063-8, 2006. Induced heterodimerization of EGFR and c-Met leads to association of the complementary .beta.-gal fragments regenerating fully functional enzyme whose activity can be monitored using a chemiluminescent substrate.
[0702] The generated cells were incubated with 12 point 1:3 serially diluted titrations of the control monovalent anti-EGFR antibody E1-F405L-gp120-K409R, the control monovalent anti-c-Met antibody M1-K409R-gp120-F405L, the combination of E1-F405L-gp120-K409R and M1-K409R-gp120-F405L, EM-1 mAb, anti-cMet parental mAb 069 or anti-EGFR parental mAb 2F8.
[0703] EM1 mAb produced a dose-dependent increase in chemiluminescence in the engineered U-2 OS cells indicating the EM1 mAb induced heterodimerization of EGFR and c-Met (FIG. 18). Neither the control monovalent antibodies nor parental bivalent antibodies induced heterodimerization of EGFR and c-Met. Thus, the synergism observed with the EM1 mAb may result from its ability to heterodimerize EGFR and c-Met, and inhibit downstream signaling of the heterodimer.
Example 22. The EM-1 Bispecific EGFR/c-Met mAb Synergistically Inhibits EGFR or c-Met Phosphorylation
[0704] The potency of the bispecific anti-EGFR/c-Met EM-1 mAb to block ligand-induced receptor phosphorylation relative to the control monovalent anti-EGFR mAb E1-F405L-gp120-K409R or anti-c-Met mAb M1-K409R-gp120-F405L, or a combination of the two was evaluated in H292, H1993 and SNU-5 cell lines having different ratios of EGFR to c-Met surface expression. The specific ABC values for EGFR and c-Met in H292, H1993, and SNU-5 cells are reported in Table 33. Inhibition of EGFR and c-Met phosphorylation was assessed as described in Example 12.
[0705] The ratio of surface EGFR to c-Met was 5.72 in H292, 0.6 in H1993 and 0.2 in SNU-5 cell line.
[0706] FIG. 19 shows the inhibition of ligand-induced c-Met (FIG. 19A) or EGFR (FIG. 19B) phosphorylation in H292 cells. The EM-1 mAb inhibited c-Met phosphorylation with a 5-fold lower IC.sub.50 value (IC.sub.50 0.80.+-.0.11 nM) when compared to the control M1-K409R-gp120-F405L antibody (IC.sub.50 4.05.+-.0.21 nM) or a combination of M1-K409R-gp120-F405L and E1-F405L-gp120-K409R. The enhanced potency of the EM-1 mAb in inhibiting c-Met phosphorylation may be a result of induced heterodimerization of EGFR and c-Met due to high EGFR expression in this cell line. The IC.sub.50 values for pEGFR inhibition were similar with the EM-1 mAb and the control monovalent E1-F405L-gp120-K409R. Thus, the higher surface EGFR levels in this cell line drive synergistic inhibition of c-Met signaling by the EM1 mAb. There was no change in the potency of the EM-1 mAb in blocking EGFR receptor phosphorylation relative to the E1-F405L-gp120-K409R alone.
[0707] FIG. 20 shows the inhibition of ligand-induced EGFR phosphorylation in SNU-5 cells. The ratio of EGFR to c-Met ABC in SNU-5 was 0.2.
[0708] EM1 mAb inhibited EGFR phosphorylation more potently (IC.sub.50 12.2 nM) when compared to the E1-F405L-gp120-K409R (no IC.sub.50 values was obtainable from the titration curve). Thus, the higher surface c-Met expression levels in this cell line drive synergistic inhibition of EGFR signaling by the EM1 mAb. There was no change in the potency of the EM1 mAb in blocking c-Met phosphorylation relative to M1-K409R-gp120-F405L alone.
[0709] FIG. 21 shows the inhibition of ligand-induced EGFR phosphorylation in H1993 cells. The ratio of EGFR to c-Met ABC in H1993 was 0.6.
[0710] EM1 mAb inhibited EGFR phosphorylation more potently when compared to E1-F405L-gp120-K409R. Thus, the higher surface c-Met expression levels also in this cell line drive synergistic inhibition of EGFR signaling by the EM1 mAb. As in SNU-5 cell line, there was no change in the potency of the EM-1 mAb in blocking c-Met phosphorylation relative to M1-K409R-gp120-F405L alone.
Sequence CWU 1
1
240189PRTArtificial SequenceTencon FN3 scaffold 1Leu Pro Ala Pro
Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp Ser1 5 10 15Leu Arg Leu
Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Leu 20 25 30Ile Gln
Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Asn Leu Thr 35 40 45Val
Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55
60Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val Lys Gly Gly His Arg Ser65
70 75 80Asn Pro Leu Ser Ala Glu Phe Thr Thr 85258DNAArtificial
Sequenceprimer 2ggaaacagga tctaccatgc tgccggcgcc gaaaaacctg
gttgtttctg aagttacc 58321DNAArtificial Sequenceprimer 3aacaccgtag
atagaaacgg t 214131DNAArtificial Sequenceprimer 4cggcggttag
aacgcggcta caattaatac ataaccccat ccccctgttg acaattaatc 60atcggctcgt
ataatgtgtg gaattgtgag cggataacaa tttcacacag gaaacaggat
120ctaccatgct g 131521DNAArtificial Sequenceprimer 5cggcggttag
aacgcggcta c 21666DNAArtificial
Sequenceprimermisc_feature(26)..(27)n is a, c, g, or
tmisc_feature(29)..(30)n is a, c, g, or tmisc_feature(32)..(33)n is
a, c, g, or tmisc_feature(35)..(36)n is a, c, g, or
tmisc_feature(38)..(39)n is a, c, g, or tmisc_feature(41)..(42)n is
a, c, g, or tmisc_feature(44)..(45)n is a, c, g, or t 6ggtggtgaat
tccgcagaca gcggsnnsnn snnsnnsnns nnsnnaacac cgtagataga 60aacggt
66769DNAArtificial Sequenceprimermisc_feature(26)..(27)n is a, c,
g, or tmisc_feature(29)..(30)n is a, c, g, or
tmisc_feature(32)..(33)n is a, c, g, or tmisc_feature(35)..(36)n is
a, c, g, or tmisc_feature(38)..(39)n is a, c, g, or
tmisc_feature(41)..(42)n is a, c, g, or tmisc_feature(44)..(45)n is
a, c, g, or tmisc_feature(47)..(48)n is a, c, g, or t 7ggtggtgaat
tccgcagaca gcggsnnsnn snnsnnsnns nnsnnsnnaa caccgtagat 60agaaacggt
69872DNAArtificial Sequenceprimermisc_feature(26)..(27)n is a, c,
g, or tmisc_feature(29)..(30)n is a, c, g, or
tmisc_feature(32)..(33)n is a, c, g, or tmisc_feature(35)..(36)n is
a, c, g, or tmisc_feature(38)..(39)n is a, c, g, or
tmisc_feature(41)..(42)n is a, c, g, or tmisc_feature(44)..(45)n is
a, c, g, or tmisc_feature(47)..(48)n is a, c, g, or
tmisc_feature(50)..(51)n is a, c, g, or t 8ggtggtgaat tccgcagaca
gcggsnnsnn snnsnnsnns nnsnnsnnsn naacaccgta 60gatagaaacg gt
72975DNAArtificial Sequenceprimermisc_feature(26)..(27)n is a, c,
g, or tmisc_feature(29)..(30)n is a, c, g, or
tmisc_feature(32)..(33)n is a, c, g, or tmisc_feature(35)..(36)n is
a, c, g, or tmisc_feature(38)..(39)n is a, c, g, or
tmisc_feature(41)..(42)n is a, c, g, or tmisc_feature(44)..(45)n is
a, c, g, or tmisc_feature(47)..(48)n is a, c, g, or
tmisc_feature(50)..(51)n is a, c, g, or tmisc_feature(53)..(54)n is
a, c, g, or t 9ggtggtgaat tccgcagaca gcggsnnsnn snnsnnsnns
nnsnnsnnsn nsnnaacacc 60gtagatagaa acggt 751080DNAArtificial
Sequenceprimermisc_feature(28)..(29)n is a, c, g, or
tmisc_feature(31)..(32)n is a, c, g, or tmisc_feature(34)..(35)n is
a, c, g, or tmisc_feature(37)..(38)n is a, c, g, or
tmisc_feature(40)..(41)n is a, c, g, or tmisc_feature(43)..(44)n is
a, c, g, or tmisc_feature(46)..(47)n is a, c, g, or
tmisc_feature(49)..(50)n is a, c, g, or tmisc_feature(52)..(53)n is
a, c, g, or tmisc_feature(55)..(56)n is a, c, g, or
tmisc_feature(58)..(59)n is a, c, g, or t 10rmggtggtga attccgcaga
cagcggsnns nnsnnsnnsn nsnnsnnsnn snnsnnsnna 60acaccgtaga tagaaacggt
801181DNAArtificial Sequenceprimermisc_feature(26)..(27)n is a, c,
g, or tmisc_feature(29)..(30)n is a, c, g, or
tmisc_feature(32)..(33)n is a, c, g, or tmisc_feature(35)..(36)n is
a, c, g, or tmisc_feature(38)..(39)n is a, c, g, or
tmisc_feature(41)..(42)n is a, c, g, or tmisc_feature(44)..(45)n is
a, c, g, or tmisc_feature(47)..(48)n is a, c, g, or
tmisc_feature(50)..(51)n is a, c, g, or tmisc_feature(53)..(54)n is
a, c, g, or tmisc_feature(56)..(57)n is a, c, g, or
tmisc_feature(59)..(60)n is a, c, g, or t 11ggtggtgaat tccgcagaca
gcggsnnsnn snnsnnsnns nnsnnsnnsn nsnnsnnsnn 60aacaccgtag atagaaacgg
t 811281DNAArtificial Sequenceprimer 12aagatcagtt gcggccgcta
gactagaacc gctgccatgg tgatggtgat ggtgaccgcc 60ggtggtgaat tccgcagaca
g 811330DNAArtificial Sequenceprimer 13cggcggttag aacgcggcta
caattaatac 301422DNAArtificial Sequenceprimer 14catgattacg
ccaagctcag aa 221565DNAArtificial Sequenceprimer 15gagccgccgc
caccggttta atggtgatgg tgatggtgac caccggtggt gaattccgca 60gacag
651637DNAArtificial Sequenceprimer 16aagaaggaga accggtatgc
tgccggcgcc gaaaaac 371789DNAArtificial Sequenceprimer 17tttgggaagc
ttctaggtct cggcggtcac catcaccatc accatggcag cggttctagt 60ctagcggccc
caactgatct tcaccaaac 891894PRTArtificial SequenceEGFR binding FN3
domain 18Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu
Asp Ser1 5 10 15Leu Arg Leu Ser Trp Ala Asp Pro His Gly Phe Tyr Asp
Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala
Ile Asn Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr
Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val
His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Met Arg Gly Leu Pro Leu
Ser Ala Glu Phe Thr Thr 85 901994PRTArtificial SequenceEGFR binding
FN3 domain 19Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr
Glu Asp Ser1 5 10 15Leu Arg Leu Ser Trp Thr Tyr Asp Arg Asp Gly Tyr
Asp Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu
Ala Ile Asn Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu
Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly
Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Met Arg Gly Leu Pro
Leu Ser Ala Glu Phe Thr Thr 85 902094PRTArtificial SequenceEGFR
binding FN3 domain 20Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu
Val Thr Glu Asp Ser1 5 10 15Leu Arg Leu Ser Trp Gly Tyr Asn Gly Asp
His Phe Asp Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val
Gly Glu Ala Ile Asn Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr
Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile
Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Met Arg Gly
Leu Pro Leu Ser Ala Glu Phe Thr Thr 85 902194PRTArtificial
SequenceEGFR binding FN3 domain 21Leu Pro Ala Pro Lys Asn Leu Val
Val Ser Glu Val Thr Glu Asp Ser1 5 10 15Leu Arg Leu Ser Trp Asp Asp
Pro Arg Gly Phe Tyr Glu Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser
Glu Lys Val Gly Glu Ala Ile Asn Leu Thr 35 40 45Val Pro Gly Ser Glu
Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr
Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn
Met Arg Gly Leu Pro Leu Ser Ala Glu Phe Thr Thr 85
902294PRTArtificial SequenceEGFR binding FN3 domain 22Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp Ser1 5 10 15Leu Arg
Leu Ser Trp Thr Trp Pro Tyr Ala Asp Leu Asp Ser Phe Leu 20 25 30Ile
Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Asn Leu Thr 35 40
45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly
50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys
Asp65 70 75 80Thr Asn Met Arg Gly Leu Pro Leu Ser Ala Glu Phe Thr
Thr 85 902394PRTArtificial SequenceEGFR binding FN3 domain 23Leu
Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp Ser1 5 10
15Leu Arg Leu Ser Trp Gly Tyr Asn Gly Asp His Phe Asp Ser Phe Leu
20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Asn Leu
Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys
Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val
Tyr Lys Asp65 70 75 80Thr Asn Met Arg Gly Leu Pro Leu Ser Ala Glu
Phe Thr Thr 85 902495PRTArtificial SequenceEGFR binding FN3 domain
24Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp Ser1
5 10 15Leu Arg Leu Ser Trp Asp Tyr Asp Leu Gly Val Tyr Phe Asp Ser
Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile
Asn Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly
Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His
Asn Val Tyr Lys65 70 75 80Asp Thr Asn Met Arg Gly Leu Pro Leu Ser
Ala Glu Phe Thr Thr 85 90 952594PRTArtificial SequenceEGFR binding
FN3 domain 25Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr
Glu Asp Ser1 5 10 15Leu Arg Leu Ser Trp Asp Asp Pro Trp Ala Phe Tyr
Glu Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu
Ala Ile Asn Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu
Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly
Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Met Arg Gly Leu Pro
Leu Ser Ala Glu Phe Thr Thr 85 902694PRTArtificial SequenceEGFR
binding FN3 domain 26Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu
Val Thr Glu Asp Ser1 5 10 15Leu Arg Leu Ser Trp Thr Ala Pro Asp Ala
Ala Phe Asp Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val
Gly Glu Ala Ile Asn Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr
Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile
Tyr Gly Val Leu Gly Ser Tyr Val Phe65 70 75 80Glu His Asp Val Met
Leu Pro Leu Ser Ala Glu Phe Thr Thr 85 902794PRTArtificial
SequenceEGFR binding FN3 domain 27Leu Pro Ala Pro Lys Asn Leu Val
Val Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser Trp Asp Asp
Pro Trp Ala Phe Tyr Glu Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser
Glu Lys Val Gly Glu Ala Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu
Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr
Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn
Met Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr 85
902894PRTArtificial SequenceEGFR binding FN3 domain 28Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg
Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Leu 20 25 30Ile
Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Val Leu Thr 35 40
45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly
50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val Leu Gly Ser Tyr Val
Phe65 70 75 80Glu His Asp Val Met Leu Pro Leu Ser Ala Ile Phe Thr
Thr 85 902994PRTArtificial SequenceEGFR binding FN3 domain 29Leu
Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp Ser1 5 10
15Leu Arg Leu Ser Trp Thr Trp Pro Tyr Ala Asp Leu Asp Ser Phe Leu
20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Asn Leu
Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys
Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val
Tyr Lys Asp65 70 75 80Thr Asn Met Arg Gly Leu Pro Leu Ser Ala Glu
Phe Thr Thr 85 903037PRTArtificial Sequenceprimer 30Ala Ala Gly Ala
Ala Gly Gly Ala Gly Ala Ala Cys Cys Gly Gly Thr1 5 10 15Ala Thr Gly
Cys Thr Gly Cys Cys Gly Gly Cys Gly Cys Cys Gly Ala 20 25 30Ala Ala
Ala Ala Cys 353165PRTArtificial Sequenceprimer 31Gly Ala Gly Cys
Cys Gly Cys Cys Gly Cys Cys Ala Cys Cys Gly Gly1 5 10 15Thr Thr Thr
Ala Ala Thr Gly Gly Thr Gly Ala Thr Gly Gly Thr Gly 20 25 30Ala Thr
Gly Gly Thr Gly Ala Cys Cys Ala Cys Cys Gly Gly Thr Gly 35 40 45Gly
Thr Gly Ala Ala Gly Ala Thr Cys Gly Cys Ala Gly Ala Cys Ala 50 55
60Gly653289PRTArtificial Sequencec-Met binding FN3 domain 32Leu Pro
Ala Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala
Arg Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp 20 25
30Ile Arg Tyr Asp Glu Val Val Val Gly Gly Glu Ala Ile Val Leu Thr
35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro
Gly 50 55 60Thr Glu Tyr Tyr Val Asn Ile Leu Gly Val Lys Gly Gly Ser
Ile Ser65 70 75 80Val Pro Leu Ser Ala Ile Phe Thr Thr
853389PRTArtificial Sequencec-Met binding FN3 domain 33Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala Arg
Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Phe 20 25 30Ile
Arg Tyr Asp Glu Phe Leu Arg Ser Gly Glu Ala Ile Val Leu Thr 35 40
45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly
50 55 60Thr Glu Tyr Trp Val Thr Ile Leu Gly Val Lys Gly Gly Leu Val
Ser65 70 75 80Thr Pro Leu Ser Ala Ile Phe Thr Thr
853489PRTArtificial Sequencec-Met binding FN3 domain 34Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala Arg
Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp 20 25 30Ile
Arg Tyr Phe Glu Phe Leu Gly Ser Gly Glu Ala Ile Val Leu Thr 35 40
45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly
50 55 60Thr Glu Tyr Ile Val Asn Ile Met Gly Val Lys Gly Gly Ser Ile
Ser65 70 75 80His Pro Leu Ser Ala Ile Phe Thr Thr
853589PRTArtificial Sequencec-Met binding FN3 domain 35Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala Arg
Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp 20 25 30Ile
Arg Tyr Phe Glu Phe Leu Gly Ser Gly Glu Ala Ile Val Leu Thr 35 40
45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly
50 55 60Thr Glu Tyr Val Val Asn Ile Leu Gly Val Lys Gly Gly Gly Leu
Ser65 70 75 80Val Pro Leu Ser Ala Ile Phe Thr Thr
853689PRTArtificial Sequencec-Met binding FN3 domain 36Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5
10 15Ala Arg Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe
Val 20 25 30Ile Arg Tyr Phe Glu Phe Leu Gly Ser Gly Glu Ala Ile Val
Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu
Lys Pro Gly 50 55 60Thr Glu Tyr Val Val Gln Ile Leu Gly Val Lys Gly
Gly Tyr Ile Ser65 70 75 80Ile Pro Leu Ser Ala Ile Phe Thr Thr
853789PRTArtificial Sequencec-Met binding FN3 domain 37Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala Arg
Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp 20 25 30Ile
Arg Tyr Leu Glu Phe Leu Leu Gly Gly Glu Ala Ile Val Leu Thr 35 40
45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly
50 55 60Thr Glu Tyr Val Val Gln Ile Met Gly Val Lys Gly Gly Thr Val
Ser65 70 75 80Pro Pro Leu Ser Ala Ile Phe Thr Thr
853889PRTArtificial Sequencec-Met binding FN3 domain 38Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala Arg
Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp 20 25 30Ile
Arg Tyr Phe Glu Phe Leu Gly Ser Gly Glu Ala Ile Val Leu Thr 35 40
45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly
50 55 60Thr Glu Tyr Val Val Gly Ile Asn Gly Val Lys Gly Gly Tyr Ile
Ser65 70 75 80Tyr Pro Leu Ser Ala Ile Phe Thr Thr
853989PRTArtificial Sequencec-Met binding FN3 domain 39Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala Arg
Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp 20 25 30Ile
Arg Tyr Phe Glu Phe Leu Gly Ser Gly Glu Ala Ile Val Leu Thr 35 40
45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Asp Leu Lys Pro Gly
50 55 60Thr Glu Tyr Gly Val Thr Ile Asn Gly Val Lys Gly Gly Arg Val
Ser65 70 75 80Thr Pro Leu Ser Ala Ile Phe Thr Thr
854089PRTArtificial Sequencec-Met binding FN3 domain 40Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala Arg
Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp 20 25 30Ile
Arg Tyr Phe Glu Phe Leu Gly Ser Gly Glu Ala Ile Val Leu Thr 35 40
45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly
50 55 60Thr Glu Tyr Val Val Gln Ile Ile Gly Val Lys Gly Gly His Ile
Ser65 70 75 80Leu Pro Leu Ser Ala Ile Phe Thr Thr
854189PRTArtificial Sequencec-Met binding FN3 domain 41Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala Arg
Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp 20 25 30Ile
Arg Tyr Phe Glu Phe Leu Gly Ser Gly Glu Ala Ile Val Leu Thr 35 40
45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly
50 55 60Thr Glu Tyr Val Val Asn Ile Met Gly Val Lys Gly Gly Lys Ile
Ser65 70 75 80Pro Pro Leu Ser Ala Ile Phe Thr Thr
854289PRTArtificial Sequencec-Met binding FN3 domain 42Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala Arg
Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp 20 25 30Ile
Arg Tyr Phe Glu Phe Leu Gly Ser Gly Glu Ala Ile Val Leu Thr 35 40
45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly
50 55 60Thr Glu Tyr Ala Val Asn Ile Met Gly Val Lys Gly Gly Arg Val
Ser65 70 75 80Val Pro Leu Ser Ala Ile Phe Thr Thr
854389PRTArtificial Sequencec-Met binding FN3 domain 43Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala Arg
Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp 20 25 30Ile
Arg Tyr Phe Glu Phe Leu Gly Ser Gly Glu Ala Ile Val Leu Thr 35 40
45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly
50 55 60Thr Glu Tyr Val Val Gln Ile Leu Gly Val Lys Gly Gly Ser Ile
Ser65 70 75 80Val Pro Leu Ser Ala Ile Phe Thr Thr
854489PRTArtificial Sequencec-Met binding FN3 domain 44Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala Arg
Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp 20 25 30Ile
Arg Tyr Phe Glu Phe Leu Gly Ser Gly Glu Ala Ile Val Leu Thr 35 40
45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly
50 55 60Thr Glu Tyr Val Val Asn Ile Met Gly Val Lys Gly Gly Ser Ile
Ser65 70 75 80Tyr Pro Leu Ser Ala Ile Phe Thr Thr
854589PRTArtificial Sequencec-Met binding FN3 domain 45Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala Arg
Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp 20 25 30Ile
Arg Tyr Phe Glu Phe Leu Gly Ser Gly Glu Ala Ile Val Leu Thr 35 40
45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly
50 55 60Thr Glu Tyr Val Val Gln Ile Leu Gly Val Lys Gly Gly Tyr Ile
Ser65 70 75 80Ile Pro Leu Ser Ala Ile Phe Thr Thr
854689PRTArtificial Sequencec-Met binding FN3 domain 46Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala Arg
Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp 20 25 30Ile
Arg Tyr Phe Glu Phe Leu Gly Ser Gly Glu Ala Ile Val Leu Thr 35 40
45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly
50 55 60Thr Glu Tyr Val Val Gln Ile Met Gly Val Lys Gly Gly Thr Val
Ser65 70 75 80Pro Pro Leu Ser Ala Ile Phe Thr Thr
854789PRTArtificial Sequencec-Met binding FN3 domain 47Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala Arg
Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp 20 25 30Ile
Arg Tyr Phe Glu Phe Thr Thr Ala Gly Glu Ala Ile Val Leu Thr 35 40
45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly
50 55 60Thr Glu Tyr Val Val Asn Ile Met Gly Val Lys Gly Gly Ser Ile
Ser65 70 75 80Pro Pro Leu Ser Ala Ile Phe Thr Thr
854889PRTArtificial Sequencec-Met binding FN3 domain 48Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala Arg
Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp 20 25 30Ile
Arg Tyr Phe Glu Leu Leu Ser Thr Gly Glu Ala Ile Val Leu Thr 35 40
45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly
50 55 60Thr Glu Tyr Val Val Asn Ile Met Gly Val Lys Gly Gly Ser Ile
Ser65 70 75 80Pro Pro Leu Ser Ala Ile Phe Thr Thr
854989PRTArtificial Sequencec-Met binding FN3 domain 49Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala Arg
Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp 20 25 30Ile
Arg Tyr Phe Glu Phe Val Ser Lys Gly Glu Ala Ile Val Leu Thr 35 40
45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly
50 55 60Thr Glu Tyr Val Val Asn Ile Met Gly Val Lys Gly Gly Ser Ile
Ser65 70 75 80Pro Pro Leu Ser Ala Ile Phe Thr Thr
8550205PRTArtificial Sequencebispecific EGFR/cMet binding molecule
50Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1
5 10 15Ser Ala Arg Leu Ser Trp Asp Asp Pro Trp Ala Phe Tyr Glu Ser
Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile
Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly
Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His
Asn Val Tyr Lys65 70 75 80Asp Thr Asn Met Arg Gly Leu Pro Leu Ser
Ala Ile Phe Thr Thr Gly 85 90 95Gly Gly Gly Ser Gly Gly Gly Gly Ser
Gly Gly Gly Gly Ser Gly Gly 100 105 110Gly Gly Ser Met Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Arg Val 115 120 125Thr Glu Asp Ser Ala
Arg Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe 130 135 140Asp Ser Phe
Trp Ile Arg Tyr Asp Glu Val Val Val Gly Gly Glu Ala145 150 155
160Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly
165 170 175Leu Lys Pro Gly Thr Glu Tyr Tyr Val Asn Ile Leu Gly Val
Lys Gly 180 185 190Gly Ser Ile Ser Val Pro Leu Ser Ala Ile Phe Thr
Thr 195 200 20551204PRTArtificial Sequencebispecific EGFR/cMet
binding molecule 51Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu
Val Thr Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Asp Asp Pro Trp Ala
Phe Tyr Glu Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val
Gly Glu Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr
Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile
Tyr Gly Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn Met Arg Gly
Leu Pro Leu Ser Ala Ile Phe Thr Thr Gly 85 90 95Gly Gly Gly Ser Gly
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly 100 105 110Gly Gly Ser
Leu Pro Ala Pro Lys Asn Leu Val Val Ser Arg Val Thr 115 120 125Glu
Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp 130 135
140Ser Phe Trp Ile Arg Tyr Phe Glu Phe Leu Gly Ser Gly Glu Ala
Ile145 150 155 160Val Leu Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp
Leu Thr Gly Leu 165 170 175Lys Pro Gly Thr Glu Tyr Val Val Asn Ile
Met Gly Val Lys Gly Gly 180 185 190Lys Ile Ser Pro Pro Leu Ser Ala
Ile Phe Thr Thr 195 20052205PRTArtificial Sequencebispecific
EGFR/cMet binding molecule 52Met Leu Pro Ala Pro Lys Asn Leu Val
Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Asp Asp
Pro Trp Ala Phe Tyr Glu Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser
Glu Lys Val Gly Glu Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser Glu
Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr
Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn
Met Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Gly 85 90 95Gly Gly
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly 100 105
110Gly Gly Ser Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Arg Val
115 120 125Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro Asp Ala
Ala Phe 130 135 140Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Leu Gly
Ser Gly Glu Ala145 150 155 160Ile Val Leu Thr Val Pro Gly Ser Glu
Arg Ser Tyr Asp Leu Thr Gly 165 170 175Leu Lys Pro Gly Thr Glu Tyr
Val Val Gln Ile Ile Gly Val Lys Gly 180 185 190Gly His Ile Ser Leu
Pro Leu Ser Ala Ile Phe Thr Thr 195 200 20553205PRTArtificial
Sequencebispecific EGFR/cMet binding molecule 53Met Leu Pro Ala Pro
Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Ala Arg Leu
Ser Trp Asp Asp Pro Trp Ala Phe Tyr Glu Ser Phe 20 25 30Leu Ile Gln
Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Val Leu 35 40 45Thr Val
Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly
Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys65 70 75
80Asp Thr Asn Met Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Gly
85 90 95Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
Gly 100 105 110Gly Gly Ser Met Leu Pro Ala Pro Lys Asn Leu Val Val
Ser Arg Val 115 120 125Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala
Pro Asp Ala Ala Phe 130 135 140Asp Ser Phe Phe Ile Arg Tyr Asp Glu
Phe Leu Arg Ser Gly Glu Ala145 150 155 160Ile Val Leu Thr Val Pro
Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly 165 170 175Leu Lys Pro Gly
Thr Glu Tyr Trp Val Thr Ile Leu Gly Val Lys Gly 180 185 190Gly Leu
Val Ser Thr Pro Leu Ser Ala Ile Phe Thr Thr 195 200
20554205PRTArtificial Sequencebispecific EGFR/cMet binding molecule
54Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1
5 10 15Ser Ala Arg Leu Ser Trp Ala Asp Pro His Gly Phe Tyr Asp Ser
Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile
Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly
Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His
Asn Val Tyr Lys65 70 75 80Asp Thr Asn Met Arg Gly Leu Pro Leu Ser
Ala Ile Phe Thr Thr Gly 85 90 95Gly Gly Gly Ser Gly Gly Gly Gly Ser
Gly Gly Gly Gly Ser Gly Gly 100 105 110Gly Gly Ser Met Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Arg Val 115 120 125Thr Glu Asp Ser Ala
Arg Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe 130 135 140Asp Ser Phe
Trp Ile Arg Tyr Phe Glu Phe Leu Gly Ser Gly Glu Ala145 150 155
160Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly
165 170 175Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile Met Gly Val
Lys Gly 180 185 190Gly Lys Ile Ser Pro Pro Leu Ser Ala Ile Phe Thr
Thr 195 200 20555205PRTArtificial Sequencebispecific EGFR/cMet
binding molecule 55Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu
Val Thr Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Ala Asp Pro His Gly
Phe Tyr Asp Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val
Gly Glu Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr
Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile
Tyr Gly Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn Met Arg Gly
Leu Pro Leu Ser Ala Ile Phe Thr Thr Gly 85 90 95Gly Gly Gly Ser Gly
Gly
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly 100 105 110Gly Gly Ser Met
Leu Pro Ala Pro Lys Asn Leu Val Val Ser Arg Val 115 120 125Thr Glu
Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe 130 135
140Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Leu Gly Ser Gly Glu
Ala145 150 155 160Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser Tyr
Asp Leu Thr Gly 165 170 175Leu Lys Pro Gly Thr Glu Tyr Val Val Gln
Ile Ile Gly Val Lys Gly 180 185 190Gly His Ile Ser Leu Pro Leu Ser
Ala Ile Phe Thr Thr 195 200 20556205PRTArtificial
Sequencebispecific EGFR/cMet binding molecule 56Met Leu Pro Ala Pro
Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Ala Arg Leu
Ser Trp Ala Asp Pro His Gly Phe Tyr Asp Ser Phe 20 25 30Leu Ile Gln
Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Val Leu 35 40 45Thr Val
Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly
Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys65 70 75
80Asp Thr Asn Met Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Gly
85 90 95Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
Gly 100 105 110Gly Gly Ser Met Leu Pro Ala Pro Lys Asn Leu Val Val
Ser Arg Val 115 120 125Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala
Pro Asp Ala Ala Phe 130 135 140Asp Ser Phe Trp Ile Arg Tyr Phe Glu
Phe Leu Gly Ser Gly Glu Ala145 150 155 160Ile Val Leu Thr Val Pro
Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly 165 170 175Leu Lys Pro Gly
Thr Glu Tyr Val Val Gln Ile Ile Gly Val Lys Gly 180 185 190Gly His
Ile Ser Leu Pro Leu Ser Ala Ile Phe Thr Thr 195 200
20557194PRTArtificial Sequencebispecific EGFR/cMet binding molecule
57Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1
5 10 15Ser Ala Arg Leu Ser Trp Asp Asp Pro Trp Ala Phe Tyr Glu Ser
Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile
Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly
Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His
Asn Val Tyr Lys65 70 75 80Asp Thr Asn Met Arg Gly Leu Pro Leu Ser
Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala Pro Ala Pro Ala Pro Ala Pro
Leu Pro Ala Pro Lys Asn Leu 100 105 110Val Val Ser Arg Val Thr Glu
Asp Ser Ala Arg Leu Ser Trp Thr Ala 115 120 125Pro Asp Ala Ala Phe
Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Leu 130 135 140Gly Ser Gly
Glu Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser145 150 155
160Tyr Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile
165 170 175Met Gly Val Lys Gly Gly Lys Ile Ser Pro Pro Leu Ser Ala
Ile Phe 180 185 190Thr Thr58194PRTArtificial Sequencebispecific
EGFR/cMet binding molecule 58Met Leu Pro Ala Pro Lys Asn Leu Val
Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Asp Asp
Pro Trp Ala Phe Tyr Glu Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser
Glu Lys Val Gly Glu Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser Glu
Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr
Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn
Met Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala
Pro Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys Asn Leu 100 105
110Val Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala
115 120 125Pro Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg Tyr Asp Glu
Val Val 130 135 140Val Gly Gly Glu Ala Ile Val Leu Thr Val Pro Gly
Ser Glu Arg Ser145 150 155 160Tyr Asp Leu Thr Gly Leu Lys Pro Gly
Thr Glu Tyr Tyr Val Asn Ile 165 170 175Leu Gly Val Lys Gly Gly Ser
Ile Ser Val Pro Leu Ser Ala Ile Phe 180 185 190Thr
Thr59195PRTArtificial Sequencebispecific EGFR/cMet binding molecule
59Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1
5 10 15Ser Ala Arg Leu Ser Trp Ala Asp Pro His Gly Phe Tyr Asp Ser
Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile
Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly
Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His
Asn Val Tyr Lys65 70 75 80Asp Thr Asn Met Arg Gly Leu Pro Leu Ser
Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala Pro Ala Pro Ala Pro Ala Pro
Met Leu Pro Ala Pro Lys Asn 100 105 110Leu Val Val Ser Arg Val Thr
Glu Asp Ser Ala Arg Leu Ser Trp Thr 115 120 125Ala Pro Asp Ala Ala
Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe 130 135 140Leu Gly Ser
Gly Glu Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg145 150 155
160Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn
165 170 175Ile Met Gly Val Lys Gly Gly Lys Ile Ser Pro Pro Leu Ser
Ala Ile 180 185 190Phe Thr Thr 19560194PRTArtificial
Sequencebispecific EGFR/cMet binding molecule 60Met Leu Pro Ala Pro
Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Ala Arg Leu
Ser Trp Ala Asp Pro His Gly Phe Tyr Asp Ser Phe 20 25 30Leu Ile Gln
Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Val Leu 35 40 45Thr Val
Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly
Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys65 70 75
80Asp Thr Asn Met Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala
85 90 95Pro Ala Pro Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys Asn
Leu 100 105 110Val Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu Ser
Trp Thr Ala 115 120 125Pro Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg
Tyr Asp Glu Val Val 130 135 140Val Gly Gly Glu Ala Ile Val Leu Thr
Val Pro Gly Ser Glu Arg Ser145 150 155 160Tyr Asp Leu Thr Gly Leu
Lys Pro Gly Thr Glu Tyr Tyr Val Asn Ile 165 170 175Leu Gly Val Lys
Gly Gly Ser Ile Ser Val Pro Leu Ser Ala Ile Phe 180 185 190Thr
Thr61194PRTArtificial Sequencebispecific EGFR/cMet binding molecule
61Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1
5 10 15Ser Ala Arg Leu Ser Trp Asp Asp Pro Trp Ala Phe Tyr Glu Ser
Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile
Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly
Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His
Asn Val Tyr Lys65 70 75 80Asp Thr Asn Ile Arg Gly Leu Pro Leu Ser
Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala Pro Ala Pro Ala Pro Ala Pro
Leu Pro Ala Pro Lys Asn Leu 100 105 110Val Val Ser Arg Val Thr Glu
Asp Ser Ala Arg Leu Ser Trp Thr Ala 115 120 125Pro Asp Ala Ala Phe
Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Leu 130 135 140Gly Ser Gly
Glu Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser145 150 155
160Tyr Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile
165 170 175Leu Gly Val Lys Gly Gly Lys Ile Ser Pro Pro Leu Ser Ala
Ile Phe 180 185 190Thr Thr62194PRTArtificial Sequencebispecific
EGFR/cMet binding molecule 62Met Leu Pro Ala Pro Lys Asn Leu Val
Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Asp Asp
Pro Trp Ala Phe Tyr Glu Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser
Glu Lys Val Gly Glu Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser Glu
Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr
Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn
Ile Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala
Pro Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys Asn Leu 100 105
110Val Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala
115 120 125Pro Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu
Phe Val 130 135 140Gly Ser Gly Glu Ala Ile Val Leu Thr Val Pro Gly
Ser Glu Arg Ser145 150 155 160Tyr Asp Leu Thr Gly Leu Lys Pro Gly
Thr Glu Tyr Val Val Asn Ile 165 170 175Leu Gly Val Lys Gly Gly Ser
Ile Ser Pro Pro Leu Ser Ala Ile Phe 180 185 190Thr
Thr63194PRTArtificial Sequencebispecific EGFR/cMet binding molecule
63Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1
5 10 15Ser Ala Arg Leu Ser Trp Asp Asp Pro Trp Ala Phe Tyr Glu Ser
Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile
Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly
Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His
Asn Val Tyr Lys65 70 75 80Asp Thr Asn Ile Arg Gly Leu Pro Leu Ser
Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala Pro Ala Pro Ala Pro Ala Pro
Leu Pro Ala Pro Lys Asn Leu 100 105 110Val Val Ser Arg Val Thr Glu
Asp Ser Ala Arg Leu Ser Trp Thr Ala 115 120 125Pro Asp Ala Ala Phe
Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Val 130 135 140Ser Lys Gly
Asp Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser145 150 155
160Tyr Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile
165 170 175Leu Gly Val Lys Gly Gly Ser Ile Ser Pro Pro Leu Ser Ala
Ile Phe 180 185 190Thr Thr64194PRTArtificial Sequencebispecific
EGFR/cMet binding molecule 64Met Leu Pro Ala Pro Lys Asn Leu Val
Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Asp Asp
Pro Trp Ala Phe Tyr Glu Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser
Glu Lys Val Gly Glu Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser Glu
Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr
Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn
Ile Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala
Pro Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys Asn Leu 100 105
110Val Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala
115 120 125Pro Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu
Phe Leu 130 135 140Gly Ser Gly Glu Ala Ile Val Leu Thr Val Pro Gly
Ser Glu Arg Ser145 150 155 160Tyr Asp Leu Thr Gly Leu Lys Pro Gly
Thr Glu Tyr Val Val Asn Ile 165 170 175Leu Ser Val Lys Gly Gly Ser
Ile Ser Pro Pro Leu Ser Ala Ile Phe 180 185 190Thr
Thr65194PRTArtificial Sequencebispecific EGFR/cMet binding molecule
65Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1
5 10 15Ser Ala Arg Leu Ser Trp Asp Asp Pro His Ala Phe Tyr Glu Ser
Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile
Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly
Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His
Asn Val Tyr Lys65 70 75 80Asp Thr Asn Ile Arg Gly Leu Pro Leu Ser
Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala Pro Ala Pro Ala Pro Ala Pro
Leu Pro Ala Pro Lys Asn Leu 100 105 110Val Val Ser Arg Val Thr Glu
Asp Ser Ala Arg Leu Ser Trp Thr Ala 115 120 125Pro Asp Ala Ala Phe
Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Leu 130 135 140Gly Ser Gly
Glu Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser145 150 155
160Tyr Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile
165 170 175Leu Gly Val Lys Gly Gly Lys Ile Ser Pro Pro Leu Ser Ala
Ile Phe 180 185 190Thr Thr66194PRTArtificial Sequencebispecific
EGFR/cMet binding molecule 66Met Leu Pro Ala Pro Lys Asn Leu Val
Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Asp Asp
Pro His Ala Phe Tyr Glu Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser
Glu Lys Val Gly Glu Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser Glu
Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr
Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn
Ile Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala
Pro Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys Asn Leu 100 105
110Val Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala
115 120 125Pro Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu
Phe Val 130 135 140Gly Ser Gly Glu Ala Ile Val Leu Thr Val Pro Gly
Ser Glu Arg Ser145 150 155 160Tyr Asp Leu Thr Gly Leu Lys Pro Gly
Thr Glu Tyr Val Val Asn Ile 165 170 175Leu Gly Val Lys Gly Gly Ser
Ile Ser Pro Pro Leu Ser Ala Ile Phe 180 185 190Thr
Thr67194PRTArtificial Sequencebispecific EGFR/cMet binding molecule
67Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1
5 10 15Ser Ala Arg Leu Ser Trp Asp Asp Pro His Ala Phe Tyr Glu Ser
Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile
Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly
Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His
Asn Val Tyr Lys65 70 75 80Asp Thr Asn Ile Arg Gly Leu Pro Leu Ser
Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala Pro Ala Pro Ala Pro Ala Pro
Leu Pro Ala Pro Lys Asn Leu 100 105 110Val Val Ser Arg Val Thr
Glu
Asp Ser Ala Arg Leu Ser Trp Thr Ala 115 120 125Pro Asp Ala Ala Phe
Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Val 130 135 140Ser Lys Gly
Asp Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser145 150 155
160Tyr Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile
165 170 175Leu Gly Val Lys Gly Gly Ser Ile Ser Pro Pro Leu Ser Ala
Ile Phe 180 185 190Thr Thr68194PRTArtificial Sequencebispecific
EGFR/cMet binding molecule 68Met Leu Pro Ala Pro Lys Asn Leu Val
Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Asp Asp
Pro His Ala Phe Tyr Glu Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser
Glu Lys Val Gly Glu Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser Glu
Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr
Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn
Ile Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala
Pro Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys Asn Leu 100 105
110Val Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala
115 120 125Pro Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu
Phe Leu 130 135 140Gly Ser Gly Glu Ala Ile Val Leu Thr Val Pro Gly
Ser Glu Arg Ser145 150 155 160Tyr Asp Leu Thr Gly Leu Lys Pro Gly
Thr Glu Tyr Val Val Asn Ile 165 170 175Leu Ser Val Lys Gly Gly Ser
Ile Ser Pro Pro Leu Ser Ala Ile Phe 180 185 190Thr
Thr69194PRTArtificial Sequencebispecific EGFR/cMet binding molecule
69Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1
5 10 15Ser Ala Arg Leu Ser Trp Ala Asp Pro His Gly Phe Tyr Asp Ser
Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile
Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly
Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His
Asn Val Tyr Lys65 70 75 80Asp Thr Asn Ile Arg Gly Leu Pro Leu Ser
Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala Pro Ala Pro Ala Pro Ala Pro
Leu Pro Ala Pro Lys Asn Leu 100 105 110Val Val Ser Arg Val Thr Glu
Asp Ser Ala Arg Leu Ser Trp Thr Ala 115 120 125Pro Asp Ala Ala Phe
Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Leu 130 135 140Gly Ser Gly
Glu Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser145 150 155
160Tyr Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile
165 170 175Leu Gly Val Lys Gly Gly Lys Ile Ser Pro Pro Leu Ser Ala
Ile Phe 180 185 190Thr Thr70194PRTArtificial Sequencebispecific
EGFR/cMet binding molecule 70Met Leu Pro Ala Pro Lys Asn Leu Val
Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Ala Asp
Pro His Gly Phe Tyr Asp Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser
Glu Lys Val Gly Glu Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser Glu
Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr
Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn
Ile Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala
Pro Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys Asn Leu 100 105
110Val Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala
115 120 125Pro Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu
Phe Val 130 135 140Gly Ser Gly Glu Ala Ile Val Leu Thr Val Pro Gly
Ser Glu Arg Ser145 150 155 160Tyr Asp Leu Thr Gly Leu Lys Pro Gly
Thr Glu Tyr Val Val Asn Ile 165 170 175Leu Gly Val Lys Gly Gly Ser
Ile Ser Pro Pro Leu Ser Ala Ile Phe 180 185 190Thr
Thr71194PRTArtificial Sequencebispecific EGFR/cMet binding molecule
71Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1
5 10 15Ser Ala Arg Leu Ser Trp Ala Asp Pro His Gly Phe Tyr Asp Ser
Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile
Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly
Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His
Asn Val Tyr Lys65 70 75 80Asp Thr Asn Ile Arg Gly Leu Pro Leu Ser
Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala Pro Ala Pro Ala Pro Ala Pro
Leu Pro Ala Pro Lys Asn Leu 100 105 110Val Val Ser Arg Val Thr Glu
Asp Ser Ala Arg Leu Ser Trp Thr Ala 115 120 125Pro Asp Ala Ala Phe
Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Val 130 135 140Ser Lys Gly
Asp Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser145 150 155
160Tyr Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile
165 170 175Leu Gly Val Lys Gly Gly Ser Ile Ser Pro Pro Leu Ser Ala
Ile Phe 180 185 190Thr Thr72194PRTArtificial Sequencebispecific
EGFR/cMet binding molecule 72Met Leu Pro Ala Pro Lys Asn Leu Val
Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Ala Asp
Pro His Gly Phe Tyr Asp Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser
Glu Lys Val Gly Glu Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser Glu
Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr
Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn
Ile Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala
Pro Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys Asn Leu 100 105
110Val Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala
115 120 125Pro Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu
Phe Leu 130 135 140Gly Ser Gly Glu Ala Ile Val Leu Thr Val Pro Gly
Ser Glu Arg Ser145 150 155 160Tyr Asp Leu Thr Gly Leu Lys Pro Gly
Thr Glu Tyr Val Val Asn Ile 165 170 175Leu Ser Val Lys Gly Gly Ser
Ile Ser Pro Pro Leu Ser Ala Ile Phe 180 185 190Thr Thr731210PRTHOMO
SAPIENS 73Met Arg Pro Ser Gly Thr Ala Gly Ala Ala Leu Leu Ala Leu
Leu Ala1 5 10 15Ala Leu Cys Pro Ala Ser Arg Ala Leu Glu Glu Lys Lys
Val Cys Gln 20 25 30Gly Thr Ser Asn Lys Leu Thr Gln Leu Gly Thr Phe
Glu Asp His Phe 35 40 45Leu Ser Leu Gln Arg Met Phe Asn Asn Cys Glu
Val Val Leu Gly Asn 50 55 60Leu Glu Ile Thr Tyr Val Gln Arg Asn Tyr
Asp Leu Ser Phe Leu Lys65 70 75 80Thr Ile Gln Glu Val Ala Gly Tyr
Val Leu Ile Ala Leu Asn Thr Val 85 90 95Glu Arg Ile Pro Leu Glu Asn
Leu Gln Ile Ile Arg Gly Asn Met Tyr 100 105 110Tyr Glu Asn Ser Tyr
Ala Leu Ala Val Leu Ser Asn Tyr Asp Ala Asn 115 120 125Lys Thr Gly
Leu Lys Glu Leu Pro Met Arg Asn Leu Gln Glu Ile Leu 130 135 140His
Gly Ala Val Arg Phe Ser Asn Asn Pro Ala Leu Cys Asn Val Glu145 150
155 160Ser Ile Gln Trp Arg Asp Ile Val Ser Ser Asp Phe Leu Ser Asn
Met 165 170 175Ser Met Asp Phe Gln Asn His Leu Gly Ser Cys Gln Lys
Cys Asp Pro 180 185 190Ser Cys Pro Asn Gly Ser Cys Trp Gly Ala Gly
Glu Glu Asn Cys Gln 195 200 205Lys Leu Thr Lys Ile Ile Cys Ala Gln
Gln Cys Ser Gly Arg Cys Arg 210 215 220Gly Lys Ser Pro Ser Asp Cys
Cys His Asn Gln Cys Ala Ala Gly Cys225 230 235 240Thr Gly Pro Arg
Glu Ser Asp Cys Leu Val Cys Arg Lys Phe Arg Asp 245 250 255Glu Ala
Thr Cys Lys Asp Thr Cys Pro Pro Leu Met Leu Tyr Asn Pro 260 265
270Thr Thr Tyr Gln Met Asp Val Asn Pro Glu Gly Lys Tyr Ser Phe Gly
275 280 285Ala Thr Cys Val Lys Lys Cys Pro Arg Asn Tyr Val Val Thr
Asp His 290 295 300Gly Ser Cys Val Arg Ala Cys Gly Ala Asp Ser Tyr
Glu Met Glu Glu305 310 315 320Asp Gly Val Arg Lys Cys Lys Lys Cys
Glu Gly Pro Cys Arg Lys Val 325 330 335Cys Asn Gly Ile Gly Ile Gly
Glu Phe Lys Asp Ser Leu Ser Ile Asn 340 345 350Ala Thr Asn Ile Lys
His Phe Lys Asn Cys Thr Ser Ile Ser Gly Asp 355 360 365Leu His Ile
Leu Pro Val Ala Phe Arg Gly Asp Ser Phe Thr His Thr 370 375 380Pro
Pro Leu Asp Pro Gln Glu Leu Asp Ile Leu Lys Thr Val Lys Glu385 390
395 400Ile Thr Gly Phe Leu Leu Ile Gln Ala Trp Pro Glu Asn Arg Thr
Asp 405 410 415Leu His Ala Phe Glu Asn Leu Glu Ile Ile Arg Gly Arg
Thr Lys Gln 420 425 430His Gly Gln Phe Ser Leu Ala Val Val Ser Leu
Asn Ile Thr Ser Leu 435 440 445Gly Leu Arg Ser Leu Lys Glu Ile Ser
Asp Gly Asp Val Ile Ile Ser 450 455 460Gly Asn Lys Asn Leu Cys Tyr
Ala Asn Thr Ile Asn Trp Lys Lys Leu465 470 475 480Phe Gly Thr Ser
Gly Gln Lys Thr Lys Ile Ile Ser Asn Arg Gly Glu 485 490 495Asn Ser
Cys Lys Ala Thr Gly Gln Val Cys His Ala Leu Cys Ser Pro 500 505
510Glu Gly Cys Trp Gly Pro Glu Pro Arg Asp Cys Val Ser Cys Arg Asn
515 520 525Val Ser Arg Gly Arg Glu Cys Val Asp Lys Cys Asn Leu Leu
Glu Gly 530 535 540Glu Pro Arg Glu Phe Val Glu Asn Ser Glu Cys Ile
Gln Cys His Pro545 550 555 560Glu Cys Leu Pro Gln Ala Met Asn Ile
Thr Cys Thr Gly Arg Gly Pro 565 570 575Asp Asn Cys Ile Gln Cys Ala
His Tyr Ile Asp Gly Pro His Cys Val 580 585 590Lys Thr Cys Pro Ala
Gly Val Met Gly Glu Asn Asn Thr Leu Val Trp 595 600 605Lys Tyr Ala
Asp Ala Gly His Val Cys His Leu Cys His Pro Asn Cys 610 615 620Thr
Tyr Gly Cys Thr Gly Pro Gly Leu Glu Gly Cys Pro Thr Asn Gly625 630
635 640Pro Lys Ile Pro Ser Ile Ala Thr Gly Met Val Gly Ala Leu Leu
Leu 645 650 655Leu Leu Val Val Ala Leu Gly Ile Gly Leu Phe Met Arg
Arg Arg His 660 665 670Ile Val Arg Lys Arg Thr Leu Arg Arg Leu Leu
Gln Glu Arg Glu Leu 675 680 685Val Glu Pro Leu Thr Pro Ser Gly Glu
Ala Pro Asn Gln Ala Leu Leu 690 695 700Arg Ile Leu Lys Glu Thr Glu
Phe Lys Lys Ile Lys Val Leu Gly Ser705 710 715 720Gly Ala Phe Gly
Thr Val Tyr Lys Gly Leu Trp Ile Pro Glu Gly Glu 725 730 735Lys Val
Lys Ile Pro Val Ala Ile Lys Glu Leu Arg Glu Ala Thr Ser 740 745
750Pro Lys Ala Asn Lys Glu Ile Leu Asp Glu Ala Tyr Val Met Ala Ser
755 760 765Val Asp Asn Pro His Val Cys Arg Leu Leu Gly Ile Cys Leu
Thr Ser 770 775 780Thr Val Gln Leu Ile Thr Gln Leu Met Pro Phe Gly
Cys Leu Leu Asp785 790 795 800Tyr Val Arg Glu His Lys Asp Asn Ile
Gly Ser Gln Tyr Leu Leu Asn 805 810 815Trp Cys Val Gln Ile Ala Lys
Gly Met Asn Tyr Leu Glu Asp Arg Arg 820 825 830Leu Val His Arg Asp
Leu Ala Ala Arg Asn Val Leu Val Lys Thr Pro 835 840 845Gln His Val
Lys Ile Thr Asp Phe Gly Leu Ala Lys Leu Leu Gly Ala 850 855 860Glu
Glu Lys Glu Tyr His Ala Glu Gly Gly Lys Val Pro Ile Lys Trp865 870
875 880Met Ala Leu Glu Ser Ile Leu His Arg Ile Tyr Thr His Gln Ser
Asp 885 890 895Val Trp Ser Tyr Gly Val Thr Val Trp Glu Leu Met Thr
Phe Gly Ser 900 905 910Lys Pro Tyr Asp Gly Ile Pro Ala Ser Glu Ile
Ser Ser Ile Leu Glu 915 920 925Lys Gly Glu Arg Leu Pro Gln Pro Pro
Ile Cys Thr Ile Asp Val Tyr 930 935 940Met Ile Met Val Lys Cys Trp
Met Ile Asp Ala Asp Ser Arg Pro Lys945 950 955 960Phe Arg Glu Leu
Ile Ile Glu Phe Ser Lys Met Ala Arg Asp Pro Gln 965 970 975Arg Tyr
Leu Val Ile Gln Gly Asp Glu Arg Met His Leu Pro Ser Pro 980 985
990Thr Asp Ser Asn Phe Tyr Arg Ala Leu Met Asp Glu Glu Asp Met Asp
995 1000 1005Asp Val Val Asp Ala Asp Glu Tyr Leu Ile Pro Gln Gln
Gly Phe 1010 1015 1020Phe Ser Ser Pro Ser Thr Ser Arg Thr Pro Leu
Leu Ser Ser Leu 1025 1030 1035Ser Ala Thr Ser Asn Asn Ser Thr Val
Ala Cys Ile Asp Arg Asn 1040 1045 1050Gly Leu Gln Ser Cys Pro Ile
Lys Glu Asp Ser Phe Leu Gln Arg 1055 1060 1065Tyr Ser Ser Asp Pro
Thr Gly Ala Leu Thr Glu Asp Ser Ile Asp 1070 1075 1080Asp Thr Phe
Leu Pro Val Pro Glu Tyr Ile Asn Gln Ser Val Pro 1085 1090 1095Lys
Arg Pro Ala Gly Ser Val Gln Asn Pro Val Tyr His Asn Gln 1100 1105
1110Pro Leu Asn Pro Ala Pro Ser Arg Asp Pro His Tyr Gln Asp Pro
1115 1120 1125His Ser Thr Ala Val Gly Asn Pro Glu Tyr Leu Asn Thr
Val Gln 1130 1135 1140Pro Thr Cys Val Asn Ser Thr Phe Asp Ser Pro
Ala His Trp Ala 1145 1150 1155Gln Lys Gly Ser His Gln Ile Ser Leu
Asp Asn Pro Asp Tyr Gln 1160 1165 1170Gln Asp Phe Phe Pro Lys Glu
Ala Lys Pro Asn Gly Ile Phe Lys 1175 1180 1185Gly Ser Thr Ala Glu
Asn Ala Glu Tyr Leu Arg Val Ala Pro Gln 1190 1195 1200Ser Ser Glu
Phe Ile Gly Ala 1205 12107453PRTHOMO SAPIENS 74Asn Ser Asp Ser Glu
Cys Pro Leu Ser His Asp Gly Tyr Cys Leu His1 5 10 15Asp Gly Val Cys
Met Tyr Ile Glu Ala Leu Asp Lys Tyr Ala Cys Asn 20 25 30Cys Val Val
Gly Tyr Ile Gly Glu Arg Cys Gln Tyr Arg Asp Leu Lys 35 40 45Trp Trp
Glu Leu Arg 50752201PRTHomo sapiens 75Met Gly Ala Met Thr Gln Leu
Leu Ala Gly Val Phe Leu Ala Phe Leu1 5 10 15Ala Leu Ala Thr Glu Gly
Gly Val Leu Lys Lys Val Ile Arg His Lys 20 25 30Arg Gln Ser Gly Val
Asn Ala Thr Leu Pro Glu Glu Asn Gln Pro Val 35 40 45Val Phe Asn His
Val Tyr Asn Ile Lys Leu Pro Val Gly Ser Gln Cys 50 55 60Ser Val Asp
Leu Glu Ser Ala Ser Gly Glu Lys Asp Leu Ala Pro Pro65 70 75 80Ser
Glu Pro Ser Glu Ser Phe Gln Glu His Thr Val Asp Gly Glu Asn 85 90
95Gln Ile Val Phe Thr His Arg Ile Asn Ile Pro Arg Arg Ala Cys Gly
100 105
110Cys Ala Ala Ala Pro Asp Val Lys Glu Leu Leu Ser Arg Leu Glu Glu
115 120 125Leu Glu Asn Leu Val Ser Ser Leu Arg Glu Gln Cys Thr Ala
Gly Ala 130 135 140Gly Cys Cys Leu Gln Pro Ala Thr Gly Arg Leu Asp
Thr Arg Pro Phe145 150 155 160Cys Ser Gly Arg Gly Asn Phe Ser Thr
Glu Gly Cys Gly Cys Val Cys 165 170 175Glu Pro Gly Trp Lys Gly Pro
Asn Cys Ser Glu Pro Glu Cys Pro Gly 180 185 190Asn Cys His Leu Arg
Gly Arg Cys Ile Asp Gly Gln Cys Ile Cys Asp 195 200 205Asp Gly Phe
Thr Gly Glu Asp Cys Ser Gln Leu Ala Cys Pro Ser Asp 210 215 220Cys
Asn Asp Gln Gly Lys Cys Val Asn Gly Val Cys Ile Cys Phe Glu225 230
235 240Gly Tyr Ala Gly Ala Asp Cys Ser Arg Glu Ile Cys Pro Val Pro
Cys 245 250 255Ser Glu Glu His Gly Thr Cys Val Asp Gly Leu Cys Val
Cys His Asp 260 265 270Gly Phe Ala Gly Asp Asp Cys Asn Lys Pro Leu
Cys Leu Asn Asn Cys 275 280 285Tyr Asn Arg Gly Arg Cys Val Glu Asn
Glu Cys Val Cys Asp Glu Gly 290 295 300Phe Thr Gly Glu Asp Cys Ser
Glu Leu Ile Cys Pro Asn Asp Cys Phe305 310 315 320Asp Arg Gly Arg
Cys Ile Asn Gly Thr Cys Tyr Cys Glu Glu Gly Phe 325 330 335Thr Gly
Glu Asp Cys Gly Lys Pro Thr Cys Pro His Ala Cys His Thr 340 345
350Gln Gly Arg Cys Glu Glu Gly Gln Cys Val Cys Asp Glu Gly Phe Ala
355 360 365Gly Val Asp Cys Ser Glu Lys Arg Cys Pro Ala Asp Cys His
Asn Arg 370 375 380Gly Arg Cys Val Asp Gly Arg Cys Glu Cys Asp Asp
Gly Phe Thr Gly385 390 395 400Ala Asp Cys Gly Glu Leu Lys Cys Pro
Asn Gly Cys Ser Gly His Gly 405 410 415Arg Cys Val Asn Gly Gln Cys
Val Cys Asp Glu Gly Tyr Thr Gly Glu 420 425 430Asp Cys Ser Gln Leu
Arg Cys Pro Asn Asp Cys His Ser Arg Gly Arg 435 440 445Cys Val Glu
Gly Lys Cys Val Cys Glu Gln Gly Phe Lys Gly Tyr Asp 450 455 460Cys
Ser Asp Met Ser Cys Pro Asn Asp Cys His Gln His Gly Arg Cys465 470
475 480Val Asn Gly Met Cys Val Cys Asp Asp Gly Tyr Thr Gly Glu Asp
Cys 485 490 495Arg Asp Arg Gln Cys Pro Arg Asp Cys Ser Asn Arg Gly
Leu Cys Val 500 505 510Asp Gly Gln Cys Val Cys Glu Asp Gly Phe Thr
Gly Pro Asp Cys Ala 515 520 525Glu Leu Ser Cys Pro Asn Asp Cys His
Gly Gln Gly Arg Cys Val Asn 530 535 540Gly Gln Cys Val Cys His Glu
Gly Phe Met Gly Lys Asp Cys Lys Glu545 550 555 560Gln Arg Cys Pro
Ser Asp Cys His Gly Gln Gly Arg Cys Val Asp Gly 565 570 575Gln Cys
Ile Cys His Glu Gly Phe Thr Gly Leu Asp Cys Gly Gln His 580 585
590Ser Cys Pro Ser Asp Cys Asn Asn Leu Gly Gln Cys Val Ser Gly Arg
595 600 605Cys Ile Cys Asn Glu Gly Tyr Ser Gly Glu Asp Cys Ser Glu
Val Ser 610 615 620Pro Pro Lys Asp Leu Val Val Thr Glu Val Thr Glu
Glu Thr Val Asn625 630 635 640Leu Ala Trp Asp Asn Glu Met Arg Val
Thr Glu Tyr Leu Val Val Tyr 645 650 655Thr Pro Thr His Glu Gly Gly
Leu Glu Met Gln Phe Arg Val Pro Gly 660 665 670Asp Gln Thr Ser Thr
Ile Ile Gln Glu Leu Glu Pro Gly Val Glu Tyr 675 680 685Phe Ile Arg
Val Phe Ala Ile Leu Glu Asn Lys Lys Ser Ile Pro Val 690 695 700Ser
Ala Arg Val Ala Thr Tyr Leu Pro Ala Pro Glu Gly Leu Lys Phe705 710
715 720Lys Ser Ile Lys Glu Thr Ser Val Glu Val Glu Trp Asp Pro Leu
Asp 725 730 735Ile Ala Phe Glu Thr Trp Glu Ile Ile Phe Arg Asn Met
Asn Lys Glu 740 745 750Asp Glu Gly Glu Ile Thr Lys Ser Leu Arg Arg
Pro Glu Thr Ser Tyr 755 760 765Arg Gln Thr Gly Leu Ala Pro Gly Gln
Glu Tyr Glu Ile Ser Leu His 770 775 780Ile Val Lys Asn Asn Thr Arg
Gly Pro Gly Leu Lys Arg Val Thr Thr785 790 795 800Thr Arg Leu Asp
Ala Pro Ser Gln Ile Glu Val Lys Asp Val Thr Asp 805 810 815Thr Thr
Ala Leu Ile Thr Trp Phe Lys Pro Leu Ala Glu Ile Asp Gly 820 825
830Ile Glu Leu Thr Tyr Gly Ile Lys Asp Val Pro Gly Asp Arg Thr Thr
835 840 845Ile Asp Leu Thr Glu Asp Glu Asn Gln Tyr Ser Ile Gly Asn
Leu Lys 850 855 860Pro Asp Thr Glu Tyr Glu Val Ser Leu Ile Ser Arg
Arg Gly Asp Met865 870 875 880Ser Ser Asn Pro Ala Lys Glu Thr Phe
Thr Thr Gly Leu Asp Ala Pro 885 890 895Arg Asn Leu Arg Arg Val Ser
Gln Thr Asp Asn Ser Ile Thr Leu Glu 900 905 910Trp Arg Asn Gly Lys
Ala Ala Ile Asp Ser Tyr Arg Ile Lys Tyr Ala 915 920 925Pro Ile Ser
Gly Gly Asp His Ala Glu Val Asp Val Pro Lys Ser Gln 930 935 940Gln
Ala Thr Thr Lys Thr Thr Leu Thr Gly Leu Arg Pro Gly Thr Glu945 950
955 960Tyr Gly Ile Gly Val Ser Ala Val Lys Glu Asp Lys Glu Ser Asn
Pro 965 970 975Ala Thr Ile Asn Ala Ala Thr Glu Leu Asp Thr Pro Lys
Asp Leu Gln 980 985 990Val Ser Glu Thr Ala Glu Thr Ser Leu Thr Leu
Leu Trp Lys Thr Pro 995 1000 1005Leu Ala Lys Phe Asp Arg Tyr Arg
Leu Asn Tyr Ser Leu Pro Thr 1010 1015 1020Gly Gln Trp Val Gly Val
Gln Leu Pro Arg Asn Thr Thr Ser Tyr 1025 1030 1035Val Leu Arg Gly
Leu Glu Pro Gly Gln Glu Tyr Asn Val Leu Leu 1040 1045 1050Thr Ala
Glu Lys Gly Arg His Lys Ser Lys Pro Ala Arg Val Lys 1055 1060
1065Ala Ser Thr Glu Gln Ala Pro Glu Leu Glu Asn Leu Thr Val Thr
1070 1075 1080Glu Val Gly Trp Asp Gly Leu Arg Leu Asn Trp Thr Ala
Ala Asp 1085 1090 1095Gln Ala Tyr Glu His Phe Ile Ile Gln Val Gln
Glu Ala Asn Lys 1100 1105 1110Val Glu Ala Ala Arg Asn Leu Thr Val
Pro Gly Ser Leu Arg Ala 1115 1120 1125Val Asp Ile Pro Gly Leu Lys
Ala Ala Thr Pro Tyr Thr Val Ser 1130 1135 1140Ile Tyr Gly Val Ile
Gln Gly Tyr Arg Thr Pro Val Leu Ser Ala 1145 1150 1155Glu Ala Ser
Thr Gly Glu Thr Pro Asn Leu Gly Glu Val Val Val 1160 1165 1170Ala
Glu Val Gly Trp Asp Ala Leu Lys Leu Asn Trp Thr Ala Pro 1175 1180
1185Glu Gly Ala Tyr Glu Tyr Phe Phe Ile Gln Val Gln Glu Ala Asp
1190 1195 1200Thr Val Glu Ala Ala Gln Asn Leu Thr Val Pro Gly Gly
Leu Arg 1205 1210 1215Ser Thr Asp Leu Pro Gly Leu Lys Ala Ala Thr
His Tyr Thr Ile 1220 1225 1230Thr Ile Arg Gly Val Thr Gln Asp Phe
Ser Thr Thr Pro Leu Ser 1235 1240 1245Val Glu Val Leu Thr Glu Glu
Val Pro Asp Met Gly Asn Leu Thr 1250 1255 1260Val Thr Glu Val Ser
Trp Asp Ala Leu Arg Leu Asn Trp Thr Thr 1265 1270 1275Pro Asp Gly
Thr Tyr Asp Gln Phe Thr Ile Gln Val Gln Glu Ala 1280 1285 1290Asp
Gln Val Glu Glu Ala His Asn Leu Thr Val Pro Gly Ser Leu 1295 1300
1305Arg Ser Met Glu Ile Pro Gly Leu Arg Ala Gly Thr Pro Tyr Thr
1310 1315 1320Val Thr Leu His Gly Glu Val Arg Gly His Ser Thr Arg
Pro Leu 1325 1330 1335Ala Val Glu Val Val Thr Glu Asp Leu Pro Gln
Leu Gly Asp Leu 1340 1345 1350Ala Val Ser Glu Val Gly Trp Asp Gly
Leu Arg Leu Asn Trp Thr 1355 1360 1365Ala Ala Asp Asn Ala Tyr Glu
His Phe Val Ile Gln Val Gln Glu 1370 1375 1380Val Asn Lys Val Glu
Ala Ala Gln Asn Leu Thr Leu Pro Gly Ser 1385 1390 1395Leu Arg Ala
Val Asp Ile Pro Gly Leu Glu Ala Ala Thr Pro Tyr 1400 1405 1410Arg
Val Ser Ile Tyr Gly Val Ile Arg Gly Tyr Arg Thr Pro Val 1415 1420
1425Leu Ser Ala Glu Ala Ser Thr Ala Lys Glu Pro Glu Ile Gly Asn
1430 1435 1440Leu Asn Val Ser Asp Ile Thr Pro Glu Ser Phe Asn Leu
Ser Trp 1445 1450 1455Met Ala Thr Asp Gly Ile Phe Glu Thr Phe Thr
Ile Glu Ile Ile 1460 1465 1470Asp Ser Asn Arg Leu Leu Glu Thr Val
Glu Tyr Asn Ile Ser Gly 1475 1480 1485Ala Glu Arg Thr Ala His Ile
Ser Gly Leu Pro Pro Ser Thr Asp 1490 1495 1500Phe Ile Val Tyr Leu
Ser Gly Leu Ala Pro Ser Ile Arg Thr Lys 1505 1510 1515Thr Ile Ser
Ala Thr Ala Thr Thr Glu Ala Leu Pro Leu Leu Glu 1520 1525 1530Asn
Leu Thr Ile Ser Asp Ile Asn Pro Tyr Gly Phe Thr Val Ser 1535 1540
1545Trp Met Ala Ser Glu Asn Ala Phe Asp Ser Phe Leu Val Thr Val
1550 1555 1560Val Asp Ser Gly Lys Leu Leu Asp Pro Gln Glu Phe Thr
Leu Ser 1565 1570 1575Gly Thr Gln Arg Lys Leu Glu Leu Arg Gly Leu
Ile Thr Gly Ile 1580 1585 1590Gly Tyr Glu Val Met Val Ser Gly Phe
Thr Gln Gly His Gln Thr 1595 1600 1605Lys Pro Leu Arg Ala Glu Ile
Val Thr Glu Ala Glu Pro Glu Val 1610 1615 1620Asp Asn Leu Leu Val
Ser Asp Ala Thr Pro Asp Gly Phe Arg Leu 1625 1630 1635Ser Trp Thr
Ala Asp Glu Gly Val Phe Asp Asn Phe Val Leu Lys 1640 1645 1650Ile
Arg Asp Thr Lys Lys Gln Ser Glu Pro Leu Glu Ile Thr Leu 1655 1660
1665Leu Ala Pro Glu Arg Thr Arg Asp Ile Thr Gly Leu Arg Glu Ala
1670 1675 1680Thr Glu Tyr Glu Ile Glu Leu Tyr Gly Ile Ser Lys Gly
Arg Arg 1685 1690 1695Ser Gln Thr Val Ser Ala Ile Ala Thr Thr Ala
Met Gly Ser Pro 1700 1705 1710Lys Glu Val Ile Phe Ser Asp Ile Thr
Glu Asn Ser Ala Thr Val 1715 1720 1725Ser Trp Arg Ala Pro Thr Ala
Gln Val Glu Ser Phe Arg Ile Thr 1730 1735 1740Tyr Val Pro Ile Thr
Gly Gly Thr Pro Ser Met Val Thr Val Asp 1745 1750 1755Gly Thr Lys
Thr Gln Thr Arg Leu Val Lys Leu Ile Pro Gly Val 1760 1765 1770Glu
Tyr Leu Val Ser Ile Ile Ala Met Lys Gly Phe Glu Glu Ser 1775 1780
1785Glu Pro Val Ser Gly Ser Phe Thr Thr Ala Leu Asp Gly Pro Ser
1790 1795 1800Gly Leu Val Thr Ala Asn Ile Thr Asp Ser Glu Ala Leu
Ala Arg 1805 1810 1815Trp Gln Pro Ala Ile Ala Thr Val Asp Ser Tyr
Val Ile Ser Tyr 1820 1825 1830Thr Gly Glu Lys Val Pro Glu Ile Thr
Arg Thr Val Ser Gly Asn 1835 1840 1845Thr Val Glu Tyr Ala Leu Thr
Asp Leu Glu Pro Ala Thr Glu Tyr 1850 1855 1860Thr Leu Arg Ile Phe
Ala Glu Lys Gly Pro Gln Lys Ser Ser Thr 1865 1870 1875Ile Thr Ala
Lys Phe Thr Thr Asp Leu Asp Ser Pro Arg Asp Leu 1880 1885 1890Thr
Ala Thr Glu Val Gln Ser Glu Thr Ala Leu Leu Thr Trp Arg 1895 1900
1905Pro Pro Arg Ala Ser Val Thr Gly Tyr Leu Leu Val Tyr Glu Ser
1910 1915 1920Val Asp Gly Thr Val Lys Glu Val Ile Val Gly Pro Asp
Thr Thr 1925 1930 1935Ser Tyr Ser Leu Ala Asp Leu Ser Pro Ser Thr
His Tyr Thr Ala 1940 1945 1950Lys Ile Gln Ala Leu Asn Gly Pro Leu
Arg Ser Asn Met Ile Gln 1955 1960 1965Thr Ile Phe Thr Thr Ile Gly
Leu Leu Tyr Pro Phe Pro Lys Asp 1970 1975 1980Cys Ser Gln Ala Met
Leu Asn Gly Asp Thr Thr Ser Gly Leu Tyr 1985 1990 1995Thr Ile Tyr
Leu Asn Gly Asp Lys Ala Glu Ala Leu Glu Val Phe 2000 2005 2010Cys
Asp Met Thr Ser Asp Gly Gly Gly Trp Ile Val Phe Leu Arg 2015 2020
2025Arg Lys Asn Gly Arg Glu Asn Phe Tyr Gln Asn Trp Lys Ala Tyr
2030 2035 2040Ala Ala Gly Phe Gly Asp Arg Arg Glu Glu Phe Trp Leu
Gly Leu 2045 2050 2055Asp Asn Leu Asn Lys Ile Thr Ala Gln Gly Gln
Tyr Glu Leu Arg 2060 2065 2070Val Asp Leu Arg Asp His Gly Glu Thr
Ala Phe Ala Val Tyr Asp 2075 2080 2085Lys Phe Ser Val Gly Asp Ala
Lys Thr Arg Tyr Lys Leu Lys Val 2090 2095 2100Glu Gly Tyr Ser Gly
Thr Ala Gly Asp Ser Met Ala Tyr His Asn 2105 2110 2115Gly Arg Ser
Phe Ser Thr Phe Asp Lys Asp Thr Asp Ser Ala Ile 2120 2125 2130Thr
Asn Cys Ala Leu Ser Tyr Lys Gly Ala Phe Trp Tyr Arg Asn 2135 2140
2145Cys His Arg Val Asn Leu Met Gly Arg Tyr Gly Asp Asn Asn His
2150 2155 2160Ser Gln Gly Val Asn Trp Phe His Trp Lys Gly His Glu
His Ser 2165 2170 2175Ile Gln Phe Ala Glu Met Lys Leu Arg Pro Ser
Asn Phe Arg Asn 2180 2185 2190Leu Glu Gly Arg Arg Lys Arg Ala 2195
22007689PRTArtificial SequenceFibcon FN3 domain 76Leu Asp Ala Pro
Thr Asp Leu Gln Val Thr Asn Val Thr Asp Thr Ser1 5 10 15Ile Thr Val
Ser Trp Thr Pro Pro Ser Ala Thr Ile Thr Gly Tyr Arg 20 25 30Ile Thr
Tyr Thr Pro Ser Asn Gly Pro Gly Glu Pro Lys Glu Leu Thr 35 40 45Val
Pro Pro Ser Ser Thr Ser Val Thr Ile Thr Gly Leu Thr Pro Gly 50 55
60Val Glu Tyr Val Val Ser Leu Tyr Ala Leu Lys Asp Asn Gln Glu Ser65
70 75 80Pro Pro Leu Val Gly Thr Gln Thr Thr 857794PRTHomo sapiens
77Val Ser Asp Val Pro Arg Asp Leu Glu Val Val Ala Ala Thr Pro Thr1
5 10 15Ser Leu Leu Ile Ser Trp Asp Ala Pro Ala Val Thr Val Arg Tyr
Tyr 20 25 30Arg Ile Thr Tyr Gly Glu Thr Gly Gly Asn Ser Pro Val Gln
Glu Phe 35 40 45Thr Val Pro Gly Ser Lys Ser Thr Ala Thr Ile Ser Gly
Leu Lys Pro 50 55 60Gly Val Asp Tyr Thr Ile Thr Val Tyr Ala Val Thr
Gly Arg Gly Asp65 70 75 80Ser Pro Ala Ser Ser Lys Pro Ile Ser Ile
Asn Tyr Arg Thr 85 90784PRTArtificial Sequencelinker 78Gly Ser Gly
Ser17925PRTArtificial Sequencelinker 79Gly Gly Gly Gly Ser Gly Gly
Gly Gly Ser Gly Gly Gly Gly Ser Gly1 5 10 15Gly Gly Gly Ser Gly Gly
Gly Gly Ser 20 25804PRTArtificial Sequencelinker 80Ala Pro Ala
Pro18110PRTArtificial Sequencelinker 81Ala Pro Ala Pro Ala Pro Ala
Pro Ala Pro1 5 108220PRTArtificial Sequencelinker 82Ala Pro Ala Pro
Ala Pro Ala Pro Ala Pro Ala Pro Ala Pro Ala Pro1 5 10 15Ala Pro Ala
Pro 208340PRTArtificial Sequencelinker 83Ala Pro Ala Pro Ala Pro
Ala Pro Ala Pro Ala Pro Ala Pro Ala Pro1 5 10 15Ala Pro Ala Pro Ala
Pro Ala Pro Ala Pro Ala Pro Ala Pro Ala Pro 20 25 30Ala Pro Ala Pro
Ala Pro Ala Pro 35 408429PRTArtificial Sequencelinker 84Ala Glu Ala
Ala Ala Lys Glu Ala Ala Ala Lys Glu Ala Ala Ala Lys1 5 10 15Glu Ala
Ala Ala Lys Glu Ala Ala Ala Lys Ala Ala Ala 20 25858PRTArtificial
Sequencetencon BC loop 85Thr Ala Pro Asp Ala Ala Phe
Asp1 5867PRTArtificial Sequencetencon FG loop 86Lys Gly Gly His Arg
Ser Asn1 5878PRTArtificial SequenceEGFR binding FN3 domain BC loop
87Ala Asp Pro His Gly Phe Tyr Asp1 5888PRTArtificial SequenceEGFR
binding FN3 domain BC loop 88Thr Tyr Asp Arg Asp Gly Tyr Asp1
5898PRTArtificial SequenceEGFR binding FN3 domain BC loop 89Trp Asp
Pro Phe Ser Phe Tyr Asp1 5908PRTArtificial SequenceEGFR binding FN3
domain BC loop 90Asp Asp Pro Arg Gly Phe Tyr Glu1 5918PRTArtificial
SequenceEGFR binding FN3 domain BC loop 91Thr Trp Pro Tyr Ala Asp
Leu Asp1 5928PRTArtificial SequenceEGFR binding FN3 domain BC loop
92Gly Tyr Asn Gly Asp His Phe Asp1 5938PRTArtificial SequenceEGFR
binding FN3 domain BC loop 93Asp Tyr Asp Leu Gly Val Tyr Asp1
5948PRTArtificial SequenceEGFR binding FN3 domain BC loop 94Asp Asp
Pro Trp Asp Phe Tyr Glu1 59512PRTArtificial SequenceEGFR binding
FN3 domain FG loop 95His Asn Val Tyr Lys Asp Thr Asn Met Arg Gly
Leu1 5 109611PRTArtificial SequenceEGFR binding FN3 domain FG loop
96Leu Gly Ser Tyr Val Phe Glu His Asp Val Met1 5
1097285DNAArtificial SequenceEGFR binding FN3 domain from ECB97;
P54AR4-83V22 97atgttgccag cgccgaagaa cctggtagtt agcgaggtta
ctgaggacag cgcgcgtctg 60agctgggacg atccgtgggc gttctacgag agctttctga
tccagtatca agagagcgag 120aaagtcggtg aagcgattgt gctgaccgtc
ccgggctccg agcgttccta cgacctgacc 180ggtttgaagc cgggtaccga
gtatacggtg agcatctacg gtgttcacaa tgtctataag 240gacactaata
tccgcggtct gcctctgagc gccattttca ccacc 28598285DNAArtificial
SequenceEGFR binding FN3 domain from ECB15; P54AR4-83V2
98atgctgccag cccctaagaa tctggtcgtg agcgaagtaa ccgaggacag cgcccgcctg
60agctgggacg acccgtgggc gttctatgag tctttcctga ttcagtatca agaaagcgaa
120aaagttggcg aagcgatcgt cctgaccgtc ccgggtagcg agcgctccta
cgatctgacc 180ggcctgaaac cgggtacgga gtacacggtg tccatttacg
gtgttcacaa tgtgtataaa 240gacaccaaca tgcgtggcct gccgctgtcg
gcgattttca ccacc 2859989PRTArtificial Sequencetencon27 FN3 domain
99Leu Pro Ala Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1
5 10 15Ala Arg Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe
Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Val
Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu
Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val Lys Gly
Gly His Arg Ser65 70 75 80Asn Pro Leu Ser Ala Ile Phe Thr Thr
8510089PRTArtificial SequenceTCL14 libraryMISC_FEATURE(32)..(32)Xaa
is any amino acidMISC_FEATURE(34)..(34)Xaa is any amino
acidMISC_FEATURE(36)..(36)Xaa is any amino
acidMISC_FEATURE(38)..(38)Xaa is any amino
acidMISC_FEATURE(39)..(39)Xaa is any amino
acidMISC_FEATURE(40)..(40)Xaa is any amino
acidMISC_FEATURE(41)..(41)Xaa is any amino
acidMISC_FEATURE(68)..(68)Xaa is any amino
acidMISC_FEATURE(70)..(70)Xaa is any amino
acidMISC_FEATURE(72)..(72)Xaa is any amino
acidMISC_FEATURE(78)..(78)Xaa is any amino
acidMISC_FEATURE(79)..(79)Xaa is any amino
acidMISC_FEATURE(81)..(81)Xaa is any amino acid 100Leu Pro Ala Pro
Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu
Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Xaa 20 25 30Ile Xaa
Tyr Xaa Glu Xaa Xaa Xaa Xaa Gly Glu Ala Ile Val Leu Thr 35 40 45Val
Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55
60Thr Glu Tyr Xaa Val Xaa Ile Xaa Gly Val Lys Gly Gly Xaa Xaa Ser65
70 75 80Xaa Pro Leu Ser Ala Ile Phe Thr Thr 851011408PRTHOMO
SAPIENS 101Met Lys Ala Pro Ala Val Leu Ala Pro Gly Ile Leu Val Leu
Leu Phe1 5 10 15Thr Leu Val Gln Arg Ser Asn Gly Glu Cys Lys Glu Ala
Leu Ala Lys 20 25 30Ser Glu Met Asn Val Asn Met Lys Tyr Gln Leu Pro
Asn Phe Thr Ala 35 40 45Glu Thr Pro Ile Gln Asn Val Ile Leu His Glu
His His Ile Phe Leu 50 55 60Gly Ala Thr Asn Tyr Ile Tyr Val Leu Asn
Glu Glu Asp Leu Gln Lys65 70 75 80Val Ala Glu Tyr Lys Thr Gly Pro
Val Leu Glu His Pro Asp Cys Phe 85 90 95Pro Cys Gln Asp Cys Ser Ser
Lys Ala Asn Leu Ser Gly Gly Val Trp 100 105 110Lys Asp Asn Ile Asn
Met Ala Leu Val Val Asp Thr Tyr Tyr Asp Asp 115 120 125Gln Leu Ile
Ser Cys Gly Ser Val Asn Arg Gly Thr Cys Gln Arg His 130 135 140Val
Phe Pro His Asn His Thr Ala Asp Ile Gln Ser Glu Val His Cys145 150
155 160Ile Phe Ser Pro Gln Ile Glu Glu Pro Ser Gln Cys Pro Asp Cys
Val 165 170 175Val Ser Ala Leu Gly Ala Lys Val Leu Ser Ser Val Lys
Asp Arg Phe 180 185 190Ile Asn Phe Phe Val Gly Asn Thr Ile Asn Ser
Ser Tyr Phe Pro Asp 195 200 205His Pro Leu His Ser Ile Ser Val Arg
Arg Leu Lys Glu Thr Lys Asp 210 215 220Gly Phe Met Phe Leu Thr Asp
Gln Ser Tyr Ile Asp Val Leu Pro Glu225 230 235 240Phe Arg Asp Ser
Tyr Pro Ile Lys Tyr Val His Ala Phe Glu Ser Asn 245 250 255Asn Phe
Ile Tyr Phe Leu Thr Val Gln Arg Glu Thr Leu Asp Ala Gln 260 265
270Thr Phe His Thr Arg Ile Ile Arg Phe Cys Ser Ile Asn Ser Gly Leu
275 280 285His Ser Tyr Met Glu Met Pro Leu Glu Cys Ile Leu Thr Glu
Lys Arg 290 295 300Lys Lys Arg Ser Thr Lys Lys Glu Val Phe Asn Ile
Leu Gln Ala Ala305 310 315 320Tyr Val Ser Lys Pro Gly Ala Gln Leu
Ala Arg Gln Ile Gly Ala Ser 325 330 335Leu Asn Asp Asp Ile Leu Phe
Gly Val Phe Ala Gln Ser Lys Pro Asp 340 345 350Ser Ala Glu Pro Met
Asp Arg Ser Ala Met Cys Ala Phe Pro Ile Lys 355 360 365Tyr Val Asn
Asp Phe Phe Asn Lys Ile Val Asn Lys Asn Asn Val Arg 370 375 380Cys
Leu Gln His Phe Tyr Gly Pro Asn His Glu His Cys Phe Asn Arg385 390
395 400Thr Leu Leu Arg Asn Ser Ser Gly Cys Glu Ala Arg Arg Asp Glu
Tyr 405 410 415Arg Thr Glu Phe Thr Thr Ala Leu Gln Arg Val Asp Leu
Phe Met Gly 420 425 430Gln Phe Ser Glu Val Leu Leu Thr Ser Ile Ser
Thr Phe Ile Lys Gly 435 440 445Asp Leu Thr Ile Ala Asn Leu Gly Thr
Ser Glu Gly Arg Phe Met Gln 450 455 460Val Val Val Ser Arg Ser Gly
Pro Ser Thr Pro His Val Asn Phe Leu465 470 475 480Leu Asp Ser His
Pro Val Ser Pro Glu Val Ile Val Glu His Thr Leu 485 490 495Asn Gln
Asn Gly Tyr Thr Leu Val Ile Thr Gly Lys Lys Ile Thr Lys 500 505
510Ile Pro Leu Asn Gly Leu Gly Cys Arg His Phe Gln Ser Cys Ser Gln
515 520 525Cys Leu Ser Ala Pro Pro Phe Val Gln Cys Gly Trp Cys His
Asp Lys 530 535 540Cys Val Arg Ser Glu Glu Cys Leu Ser Gly Thr Trp
Thr Gln Gln Ile545 550 555 560Cys Leu Pro Ala Ile Tyr Lys Val Phe
Pro Asn Ser Ala Pro Leu Glu 565 570 575Gly Gly Thr Arg Leu Thr Ile
Cys Gly Trp Asp Phe Gly Phe Arg Arg 580 585 590Asn Asn Lys Phe Asp
Leu Lys Lys Thr Arg Val Leu Leu Gly Asn Glu 595 600 605Ser Cys Thr
Leu Thr Leu Ser Glu Ser Thr Met Asn Thr Leu Lys Cys 610 615 620Thr
Val Gly Pro Ala Met Asn Lys His Phe Asn Met Ser Ile Ile Ile625 630
635 640Ser Asn Gly His Gly Thr Thr Gln Tyr Ser Thr Phe Ser Tyr Val
Asp 645 650 655Pro Val Ile Thr Ser Ile Ser Pro Lys Tyr Gly Pro Met
Ala Gly Gly 660 665 670Thr Leu Leu Thr Leu Thr Gly Asn Tyr Leu Asn
Ser Gly Asn Ser Arg 675 680 685His Ile Ser Ile Gly Gly Lys Thr Cys
Thr Leu Lys Ser Val Ser Asn 690 695 700Ser Ile Leu Glu Cys Tyr Thr
Pro Ala Gln Thr Ile Ser Thr Glu Phe705 710 715 720Ala Val Lys Leu
Lys Ile Asp Leu Ala Asn Arg Glu Thr Ser Ile Phe 725 730 735Ser Tyr
Arg Glu Asp Pro Ile Val Tyr Glu Ile His Pro Thr Lys Ser 740 745
750Phe Ile Ser Thr Trp Trp Lys Glu Pro Leu Asn Ile Val Ser Phe Leu
755 760 765Phe Cys Phe Ala Ser Gly Gly Ser Thr Ile Thr Gly Val Gly
Lys Asn 770 775 780Leu Asn Ser Val Ser Val Pro Arg Met Val Ile Asn
Val His Glu Ala785 790 795 800Gly Arg Asn Phe Thr Val Ala Cys Gln
His Arg Ser Asn Ser Glu Ile 805 810 815Ile Cys Cys Thr Thr Pro Ser
Leu Gln Gln Leu Asn Leu Gln Leu Pro 820 825 830Leu Lys Thr Lys Ala
Phe Phe Met Leu Asp Gly Ile Leu Ser Lys Tyr 835 840 845Phe Asp Leu
Ile Tyr Val His Asn Pro Val Phe Lys Pro Phe Glu Lys 850 855 860Pro
Val Met Ile Ser Met Gly Asn Glu Asn Val Leu Glu Ile Lys Gly865 870
875 880Asn Asp Ile Asp Pro Glu Ala Val Lys Gly Glu Val Leu Lys Val
Gly 885 890 895Asn Lys Ser Cys Glu Asn Ile His Leu His Ser Glu Ala
Val Leu Cys 900 905 910Thr Val Pro Asn Asp Leu Leu Lys Leu Asn Ser
Glu Leu Asn Ile Glu 915 920 925Trp Lys Gln Ala Ile Ser Ser Thr Val
Leu Gly Lys Val Ile Val Gln 930 935 940Pro Asp Gln Asn Phe Thr Gly
Leu Ile Ala Gly Val Val Ser Ile Ser945 950 955 960Thr Ala Leu Leu
Leu Leu Leu Gly Phe Phe Leu Trp Leu Lys Lys Arg 965 970 975Lys Gln
Ile Lys Asp Leu Gly Ser Glu Leu Val Arg Tyr Asp Ala Arg 980 985
990Val His Thr Pro His Leu Asp Arg Leu Val Ser Ala Arg Ser Val Ser
995 1000 1005Pro Thr Thr Glu Met Val Ser Asn Glu Ser Val Asp Tyr
Arg Ala 1010 1015 1020Thr Phe Pro Glu Asp Gln Phe Pro Asn Ser Ser
Gln Asn Gly Ser 1025 1030 1035Cys Arg Gln Val Gln Tyr Pro Leu Thr
Asp Met Ser Pro Ile Leu 1040 1045 1050Thr Ser Gly Asp Ser Asp Ile
Ser Ser Pro Leu Leu Gln Asn Thr 1055 1060 1065Val His Ile Asp Leu
Ser Ala Leu Asn Pro Glu Leu Val Gln Ala 1070 1075 1080Val Gln His
Val Val Ile Gly Pro Ser Ser Leu Ile Val His Phe 1085 1090 1095Asn
Glu Val Ile Gly Arg Gly His Phe Gly Cys Val Tyr His Gly 1100 1105
1110Thr Leu Leu Asp Asn Asp Gly Lys Lys Ile His Cys Ala Val Lys
1115 1120 1125Ser Leu Asn Arg Ile Thr Asp Ile Gly Glu Val Ser Gln
Phe Leu 1130 1135 1140Thr Glu Gly Ile Ile Met Lys Asp Phe Ser His
Pro Asn Val Leu 1145 1150 1155Ser Leu Leu Gly Ile Cys Leu Arg Ser
Glu Gly Ser Pro Leu Val 1160 1165 1170Val Leu Pro Tyr Met Lys His
Gly Asp Leu Arg Asn Phe Ile Arg 1175 1180 1185Asn Glu Thr His Asn
Pro Thr Val Lys Asp Leu Ile Gly Phe Gly 1190 1195 1200Leu Gln Val
Ala Lys Gly Met Lys Tyr Leu Ala Ser Lys Lys Phe 1205 1210 1215Val
His Arg Asp Leu Ala Ala Arg Asn Cys Met Leu Asp Glu Lys 1220 1225
1230Phe Thr Val Lys Val Ala Asp Phe Gly Leu Ala Arg Asp Met Tyr
1235 1240 1245Asp Lys Glu Tyr Tyr Ser Val His Asn Lys Thr Gly Ala
Lys Leu 1250 1255 1260Pro Val Lys Trp Met Ala Leu Glu Ser Leu Gln
Thr Gln Lys Phe 1265 1270 1275Thr Thr Lys Ser Asp Val Trp Ser Phe
Gly Val Leu Leu Trp Glu 1280 1285 1290Leu Met Thr Arg Gly Ala Pro
Pro Tyr Pro Asp Val Asn Thr Phe 1295 1300 1305Asp Ile Thr Val Tyr
Leu Leu Gln Gly Arg Arg Leu Leu Gln Pro 1310 1315 1320Glu Tyr Cys
Pro Asp Pro Leu Tyr Glu Val Met Leu Lys Cys Trp 1325 1330 1335His
Pro Lys Ala Glu Met Arg Pro Ser Phe Ser Glu Leu Val Ser 1340 1345
1350Arg Ile Ser Ala Ile Phe Ser Thr Phe Ile Gly Glu His Tyr Val
1355 1360 1365His Val Asn Ala Thr Tyr Val Asn Val Lys Cys Val Ala
Pro Tyr 1370 1375 1380Pro Ser Leu Leu Ser Ser Glu Asp Asn Ala Asp
Asp Glu Val Asp 1385 1390 1395Thr Arg Pro Ala Ser Phe Trp Glu Thr
Ser 1400 1405102697PRTHOMO SAPIENS 102Gln Arg Lys Arg Arg Asn Thr
Ile His Glu Phe Lys Lys Ser Ala Lys1 5 10 15Thr Thr Leu Ile Lys Ile
Asp Pro Ala Leu Lys Ile Lys Thr Lys Lys 20 25 30Val Asn Thr Ala Asp
Gln Cys Ala Asn Arg Cys Thr Arg Asn Lys Gly 35 40 45Leu Pro Phe Thr
Cys Lys Ala Phe Val Phe Asp Lys Ala Arg Lys Gln 50 55 60Cys Leu Trp
Phe Pro Phe Asn Ser Met Ser Ser Gly Val Lys Lys Glu65 70 75 80Phe
Gly His Glu Phe Asp Leu Tyr Glu Asn Lys Asp Tyr Ile Arg Asn 85 90
95Cys Ile Ile Gly Lys Gly Arg Ser Tyr Lys Gly Thr Val Ser Ile Thr
100 105 110Lys Ser Gly Ile Lys Cys Gln Pro Trp Ser Ser Met Ile Pro
His Glu 115 120 125His Ser Phe Leu Pro Ser Ser Tyr Arg Gly Lys Asp
Leu Gln Glu Asn 130 135 140Tyr Cys Arg Asn Pro Arg Gly Glu Glu Gly
Gly Pro Trp Cys Phe Thr145 150 155 160Ser Asn Pro Glu Val Arg Tyr
Glu Val Cys Asp Ile Pro Gln Cys Ser 165 170 175Glu Val Glu Cys Met
Thr Cys Asn Gly Glu Ser Tyr Arg Gly Leu Met 180 185 190Asp His Thr
Glu Ser Gly Lys Ile Cys Gln Arg Trp Asp His Gln Thr 195 200 205Pro
His Arg His Lys Phe Leu Pro Glu Arg Tyr Pro Asp Lys Gly Phe 210 215
220Asp Asp Asn Tyr Cys Arg Asn Pro Asp Gly Gln Pro Arg Pro Trp
Cys225 230 235 240Tyr Thr Leu Asp Pro His Thr Arg Trp Glu Tyr Cys
Ala Ile Lys Thr 245 250 255Cys Ala Asp Asn Thr Met Asn Asp Thr Asp
Val Pro Leu Glu Thr Thr 260 265 270Glu Cys Ile Gln Gly Gln Gly Glu
Gly Tyr Arg Gly Thr Val Asn Thr 275 280 285Ile Trp Asn Gly Ile Pro
Cys Gln Arg Trp Asp Ser Gln Tyr Pro His 290 295 300Glu His Asp Met
Thr Pro Glu Asn Phe Lys Cys Lys Asp Leu Arg Glu305 310 315 320Asn
Tyr Cys Arg Asn Pro Asp Gly Ser Glu Ser Pro Trp Cys Phe Thr 325 330
335Thr Asp Pro Asn Ile Arg Val Gly Tyr Cys Ser Gln Ile Pro Asn Cys
340 345 350Asp Met Ser His Gly Gln Asp Cys Tyr Arg Gly Asn Gly Lys
Asn Tyr 355 360 365Met Gly Asn Leu Ser Gln Thr Arg Ser Gly Leu Thr
Cys Ser Met Trp 370 375 380Asp Lys Asn Met Glu Asp Leu His Arg His
Ile Phe Trp Glu Pro Asp385 390 395 400Ala Ser Lys Leu Asn Glu Asn
Tyr Cys Arg Asn Pro Asp Asp Asp Ala 405 410 415His Gly Pro Trp Cys
Tyr Thr Gly Asn Pro Leu Ile Pro Trp Asp Tyr 420 425 430Cys Pro Ile
Ser Arg Cys Glu Gly Asp Thr Thr Pro Thr Ile Val Asn 435 440 445Leu
Asp His Pro Val Ile Ser Cys Ala Lys Thr Lys Gln Leu Arg Val 450 455
460Val Asn Gly Ile Pro Thr Arg Thr Asn Ile Gly Trp Met Val Ser
Leu465 470 475 480Arg Tyr Arg Asn Lys His Ile Cys Gly Gly Ser
Leu
Ile Lys Glu Ser 485 490 495Trp Val Leu Thr Ala Arg Gln Cys Phe Pro
Ser Arg Asp Leu Lys Asp 500 505 510Tyr Glu Ala Trp Leu Gly Ile His
Asp Val His Gly Arg Gly Asp Glu 515 520 525Lys Cys Lys Gln Val Leu
Asn Val Ser Gln Leu Val Tyr Gly Pro Glu 530 535 540Gly Ser Asp Leu
Val Leu Met Lys Leu Ala Arg Pro Ala Val Leu Asp545 550 555 560Asp
Phe Val Ser Thr Ile Asp Leu Pro Asn Tyr Gly Cys Thr Ile Pro 565 570
575Glu Lys Thr Ser Cys Ser Val Tyr Gly Trp Gly Tyr Thr Gly Leu Ile
580 585 590Asn Tyr Asp Gly Leu Leu Arg Val Ala His Leu Tyr Ile Met
Gly Asn 595 600 605Glu Lys Cys Ser Gln His His Arg Gly Lys Val Thr
Leu Asn Glu Ser 610 615 620Glu Ile Cys Ala Gly Ala Glu Lys Ile Gly
Ser Gly Pro Cys Glu Gly625 630 635 640Asp Tyr Gly Gly Pro Leu Val
Cys Glu Gln His Lys Met Arg Met Val 645 650 655Leu Gly Val Ile Val
Pro Gly Arg Gly Cys Ala Ile Pro Asn Arg Pro 660 665 670Gly Ile Phe
Val Arg Val Ala Tyr Tyr Ala Lys Trp Ile His Lys Ile 675 680 685Ile
Leu Thr Tyr Lys Val Pro Gln Ser 690 695103276DNAArtificial
Sequencec-Met binding FN3 domain 103ctgccggctc cgaagaactt
ggtggtgagc cgtgttaccg aagatagcgc acgcctgagc 60tggacggcac cggatgcggc
gttcgatagc ttctggattc gctattttga gtttctgggt 120agcggtgagg
caattgttct gacggtgccg ggctctgaac gctcctacga tttgaccggt
180ctgaaaccgg gcaccgagta tgtggtgaac attctgagcg ttaagggcgg
tagcatcagc 240ccaccgctga gcgcgatctt cacgactggt ggttgc
276104267DNAArtificial Sequencec-Met binding FN3 domain
104ctgccggcac cgaagaacct ggttgtcagc cgtgtgaccg aggatagcgc
acgtttgagc 60tggaccgctc cggatgcagc ctttgacagc ttctggattc gttactttga
atttctgggt 120agcggtgagg cgatcgttct gacggtgccg ggctctgaac
gcagctatga tttgacgggc 180ctgaagccgg gtactgagta cgtggttaac
atcatgggcg ttaagggtgg taaaatcagc 240ccgccattgt ccgcgatctt taccacg
2671055PRTArtificial Sequencelinker 105Gly Gly Gly Gly Ser1
5106194PRTArtificial SequenceBispecific EGFR/c-Met binding molecule
106Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1
5 10 15Ser Ala Arg Leu Ser Trp Asp Asp Pro Trp Ala Phe Tyr Glu Ser
Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile
Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly
Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His
Asn Val Tyr Lys65 70 75 80Asp Thr Asn Ile Arg Gly Leu Pro Leu Ser
Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala Pro Ala Pro Ala Pro Ala Pro
Leu Pro Ala Pro Lys Asn Leu 100 105 110Val Val Ser Arg Val Thr Glu
Asp Ser Ala Arg Leu Ser Trp Thr Ala 115 120 125Pro Asp Ala Ala Phe
Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Leu 130 135 140Gly Ser Gly
Glu Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser145 150 155
160Tyr Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile
165 170 175Leu Ser Val Lys Gly Gly Ser Ile Ser Pro Pro Leu Ser Ala
Ile Phe 180 185 190Thr Thr10794PRTArtificial SequenceEGFR binding
FN3 domain 107Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr
Glu Asp Ser1 5 10 15Ala Arg Leu Ser Trp Ala Asp Pro His Gly Phe Tyr
Asp Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu
Ala Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu
Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly
Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Met Arg Gly Leu Pro
Leu Ser Ala Ile Phe Thr Thr 85 9010894PRTArtificial SequenceEGFR
binding FN3 domain 108Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu
Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser Trp Asp Asp Pro Trp Ala
Phe Tyr Glu Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val
Gly Glu Ala Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr
Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile
Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Ile Arg Gly
Leu Pro Leu Ser Ala Ile Phe Thr Thr 85 9010994PRTArtificial
SequenceEGFR binding FN3 domain 109Leu Pro Ala Pro Lys Asn Leu Val
Val Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser Trp Asp Asp
Pro His Ala Phe Tyr Glu Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser
Glu Lys Val Gly Glu Ala Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu
Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr
Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn
Ile Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr 85
9011094PRTArtificial SequenceEGFR binding FN3 domain 110Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg
Leu Ser Trp Ala Asp Pro His Gly Phe Tyr Asp Ser Phe Leu 20 25 30Ile
Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Val Leu Thr 35 40
45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly
50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys
Asp65 70 75 80Thr Asn Ile Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr
Thr 85 9011189PRTArtificial Sequencec-Met binding FN3 domain 111Leu
Pro Ala Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10
15Ala Arg Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp
20 25 30Ile Arg Tyr Phe Glu Phe Leu Gly Ser Gly Glu Ala Ile Val Leu
Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys
Pro Gly 50 55 60Thr Glu Tyr Val Val Asn Ile Leu Gly Val Lys Gly Gly
Lys Ile Ser65 70 75 80Pro Pro Leu Ser Ala Ile Phe Thr Thr
8511289PRTArtificial Sequencec-Met binding FN3 domain 112Leu Pro
Ala Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala
Arg Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp 20 25
30Ile Arg Tyr Phe Glu Phe Val Gly Ser Gly Glu Ala Ile Val Leu Thr
35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro
Gly 50 55 60Thr Glu Tyr Val Val Asn Ile Leu Gly Val Lys Gly Gly Ser
Ile Ser65 70 75 80Pro Pro Leu Ser Ala Ile Phe Thr Thr
8511389PRTArtificial Sequencec-Met binding FN3 domain 113Leu Pro
Ala Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala
Arg Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp 20 25
30Ile Arg Tyr Phe Glu Phe Val Ser Lys Gly Asp Ala Ile Val Leu Thr
35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro
Gly 50 55 60Thr Glu Tyr Val Val Asn Ile Leu Gly Val Lys Gly Gly Ser
Ile Ser65 70 75 80Pro Pro Leu Ser Ala Ile Phe Thr Thr
8511489PRTArtificial Sequencec-Met binding FN3 domain 114Leu Pro
Ala Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala
Arg Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp 20 25
30Ile Arg Tyr Phe Glu Phe Leu Gly Ser Gly Glu Ala Ile Val Leu Thr
35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro
Gly 50 55 60Thr Glu Tyr Val Val Asn Ile Leu Ser Val Lys Gly Gly Ser
Ile Ser65 70 75 80Pro Pro Leu Ser Ala Ile Phe Thr Thr
85115591DNAArtificial SequenceBispecific EGFR/c-Met binding
molecule 115atgttgccag cgccgaagaa cctggtagtt agcgaggtta ctgaggacag
cgcgcgtctg 60agctgggacg atccgtgggc gttctacgag agctttctga tccagtatca
agagagcgag 120aaagtcggtg aagcgattgt gctgaccgtc ccgggctccg
agcgttccta cgacctgacc 180ggtttgaagc cgggtaccga gtatacggtg
agcatctacg gtgttcacaa tgtctataag 240gacactaata tccgcggtct
gcctctgagc gccattttca ccaccgcacc ggcaccggct 300ccggctcctg
ccccgctgcc ggctccgaag aacttggtgg tgagccgtgt taccgaagat
360agcgcacgcc tgagctggac ggcaccggat gcggcgttcg atagcttctg
gattcgctat 420tttgagtttc tgggtagcgg tgaggcaatt gttctgacgg
tgccgggctc tgaacgctcc 480tacgatttga ccggtctgaa accgggcacc
gagtatgtgg tgaacattct gagcgttaag 540ggcggtagca tcagcccacc
gctgagcgcg atcttcacga ctggtggttg c 591116582DNAArtificial
SequenceBispecific EGFR/c-Met binding molecule 116atgctgccag
cccctaagaa tctggtcgtg agcgaagtaa ccgaggacag cgcccgcctg 60agctgggacg
acccgtgggc gttctatgag tctttcctga ttcagtatca agaaagcgaa
120aaagttggcg aagcgatcgt cctgaccgtc ccgggtagcg agcgctccta
cgatctgacc 180ggcctgaaac cgggtacgga gtacacggtg tccatttacg
gtgttcacaa tgtgtataaa 240gacaccaaca tgcgtggcct gccgctgtcg
gcgattttca ccaccgcgcc tgcgccagcg 300cctgcaccgg ctccgctgcc
ggcaccgaag aacctggttg tcagccgtgt gaccgaggat 360agcgcacgtt
tgagctggac cgctccggat gcagcctttg acagcttctg gattcgttac
420tttgaatttc tgggtagcgg tgaggcgatc gttctgacgg tgccgggctc
tgaacgcagc 480tatgatttga cgggcctgaa gccgggtact gagtacgtgg
ttaacatcat gggcgttaag 540ggtggtaaaa tcagcccgcc attgtccgcg
atctttacca cg 58211751PRTArtificial Sequencealbumin binding domain
117Thr Ile Asp Glu Trp Leu Leu Lys Glu Ala Lys Glu Lys Ala Ile Glu1
5 10 15Glu Leu Lys Lys Ala Gly Ile Thr Ser Asp Tyr Tyr Phe Asp Leu
Ile 20 25 30Asn Lys Ala Lys Thr Val Glu Gly Val Asn Ala Leu Lys Asp
Glu Ile 35 40 45Leu Lys Ala 50118250PRTArtificial
SequenceBispecific EGFR/c-Met binding molecule 118Met Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Ala Arg
Leu Ser Trp Asp Asp Pro Trp Ala Phe Tyr Glu Ser Phe 20 25 30Leu Ile
Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Val Leu 35 40 45Thr
Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55
60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys65
70 75 80Asp Thr Asn Met Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr
Ala 85 90 95Pro Ala Pro Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys
Asn Leu 100 105 110Val Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu
Ser Trp Thr Ala 115 120 125Pro Asp Ala Ala Phe Asp Ser Phe Trp Ile
Arg Tyr Asp Glu Val Val 130 135 140Val Gly Gly Glu Ala Ile Val Leu
Thr Val Pro Gly Ser Glu Arg Ser145 150 155 160Tyr Asp Leu Thr Gly
Leu Lys Pro Gly Thr Glu Tyr Tyr Val Asn Ile 165 170 175Leu Gly Val
Lys Gly Gly Ser Ile Ser Val Pro Leu Ser Ala Ile Phe 180 185 190Thr
Thr Ala Pro Ala Pro Ala Pro Ala Pro Ala Pro Leu Ala Glu Ala 195 200
205Lys Val Leu Ala Asn Arg Glu Leu Asp Lys Tyr Gly Val Ser Asp Tyr
210 215 220Tyr Lys Asn Leu Ile Asn Asn Ala Lys Thr Val Glu Gly Val
Lys Ala225 230 235 240Leu Leu Asp Glu Ile Leu Ala Ala Leu Pro 245
250119250PRTArtificial SequenceBispecific EGFR/c-Met binding
molecule 119Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr
Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Ala Asp Pro His Gly Phe Tyr
Asp Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu
Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu
Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly
Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn Met Arg Gly Leu Pro
Leu Ser Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala Pro Ala Pro Ala Pro
Ala Pro Leu Pro Ala Pro Lys Asn Leu 100 105 110Val Val Ser Arg Val
Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala 115 120 125Pro Asp Ala
Ala Phe Asp Ser Phe Trp Ile Arg Tyr Asp Glu Val Val 130 135 140Val
Gly Gly Glu Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser145 150
155 160Tyr Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Tyr Val Asn
Ile 165 170 175Leu Gly Val Lys Gly Gly Ser Ile Ser Val Pro Leu Ser
Ala Ile Phe 180 185 190Thr Thr Ala Pro Ala Pro Ala Pro Ala Pro Ala
Pro Leu Ala Glu Ala 195 200 205Lys Val Leu Ala Asn Arg Glu Leu Asp
Lys Tyr Gly Val Ser Asp Tyr 210 215 220Tyr Lys Asn Leu Ile Asn Asn
Ala Lys Thr Val Glu Gly Val Lys Ala225 230 235 240Leu Leu Asp Glu
Ile Leu Ala Ala Leu Pro 245 250120250PRTArtificial
SequenceBispecific EGFR/c-Met binding molecule 120Met Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Ala Arg
Leu Ser Trp Asp Asp Pro Trp Ala Phe Tyr Glu Ser Phe 20 25 30Leu Ile
Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Val Leu 35 40 45Thr
Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55
60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys65
70 75 80Asp Thr Asn Met Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr
Ala 85 90 95Pro Ala Pro Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys
Asn Leu 100 105 110Val Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu
Ser Trp Thr Ala 115 120 125Pro Asp Ala Ala Phe Asp Ser Phe Trp Ile
Arg Tyr Phe Glu Phe Leu 130 135 140Gly Ser Gly Glu Ala Ile Val Leu
Thr Val Pro Gly Ser Glu Arg Ser145 150 155 160Tyr Asp Leu Thr Gly
Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile 165 170 175Met Gly Val
Lys Gly Gly Lys Ile Ser Pro Pro Leu Ser Ala Ile Phe 180 185 190Thr
Thr Ala Pro Ala Pro Ala Pro Ala Pro Ala Pro Leu Ala Glu Ala 195 200
205Lys Val Leu Ala Asn Arg Glu Leu Asp Lys Tyr Gly Val Ser Asp Tyr
210 215 220Tyr Lys Asn Leu Ile Asn Asn Ala Lys Thr Val Glu Gly Val
Lys Ala225 230 235 240Leu Leu Asp Glu Ile Leu Ala Ala Leu Pro 245
250121250PRTArtificial SequenceBispecific EGFR/c-Met binding
molecule 121Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr
Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Ala Asp Pro His Gly Phe Tyr
Asp Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu
Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu
Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly
Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn Met Arg Gly Leu Pro
Leu Ser Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala Pro Ala Pro Ala Pro
Ala Pro Leu Pro
Ala Pro Lys Asn Leu 100 105 110Val Val Ser Arg Val Thr Glu Asp Ser
Ala Arg Leu Ser Trp Thr Ala 115 120 125Pro Asp Ala Ala Phe Asp Ser
Phe Trp Ile Arg Tyr Phe Glu Phe Leu 130 135 140Gly Ser Gly Glu Ala
Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser145 150 155 160Tyr Asp
Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile 165 170
175Met Gly Val Lys Gly Gly Lys Ile Ser Pro Pro Leu Ser Ala Ile Phe
180 185 190Thr Thr Ala Pro Ala Pro Ala Pro Ala Pro Ala Pro Leu Ala
Glu Ala 195 200 205Lys Val Leu Ala Asn Arg Glu Leu Asp Lys Tyr Gly
Val Ser Asp Tyr 210 215 220Tyr Lys Asn Leu Ile Asn Asn Ala Lys Thr
Val Glu Gly Val Lys Ala225 230 235 240Leu Leu Asp Glu Ile Leu Ala
Ala Leu Pro 245 25012295PRTArtificial SequenceEGFR binding FN3
domain 122Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr
Glu Asp1 5 10 15Ser Leu Arg Leu Ser Trp Ala Asp Pro His Gly Phe Tyr
Asp Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu
Ala Ile Asn Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu
Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly
Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn Met Arg Gly Leu Pro
Leu Ser Ala Glu Phe Thr Thr 85 90 9512395PRTArtificial SequenceEGFR
binding FN3 domain 123Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser
Glu Val Thr Glu Asp1 5 10 15Ser Leu Arg Leu Ser Trp Thr Tyr Asp Arg
Asp Gly Tyr Asp Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys
Val Gly Glu Ala Ile Asn Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser
Tyr Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser
Ile Tyr Gly Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn Met Arg
Gly Leu Pro Leu Ser Ala Glu Phe Thr Thr 85 90 9512495PRTArtificial
SequenceEGFR binding FN3 domain 124Met Leu Pro Ala Pro Lys Asn Leu
Val Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Leu Arg Leu Ser Trp Gly
Tyr Asn Gly Asp His Phe Asp Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu
Ser Glu Lys Val Gly Glu Ala Ile Asn Leu 35 40 45Thr Val Pro Gly Ser
Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr
Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys65 70 75 80Asp Thr
Asn Met Arg Gly Leu Pro Leu Ser Ala Glu Phe Thr Thr 85 90
9512595PRTArtificial SequenceEGFR binding FN3 domain 125Met Leu Pro
Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Leu
Arg Leu Ser Trp Asp Asp Pro Arg Gly Phe Tyr Glu Ser Phe 20 25 30Leu
Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Asn Leu 35 40
45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro
50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr
Lys65 70 75 80Asp Thr Asn Met Arg Gly Leu Pro Leu Ser Ala Glu Phe
Thr Thr 85 90 9512695PRTArtificial SequenceEGFR binding FN3 domain
126Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1
5 10 15Ser Leu Arg Leu Ser Trp Thr Trp Pro Tyr Ala Asp Leu Asp Ser
Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile
Asn Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly
Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His
Asn Val Tyr Lys65 70 75 80Asp Thr Asn Met Arg Gly Leu Pro Leu Ser
Ala Glu Phe Thr Thr 85 90 9512795PRTArtificial SequenceEGFR binding
FN3 domain 127Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val
Thr Glu Asp1 5 10 15Ser Leu Arg Leu Ser Trp Gly Tyr Asn Gly Asp His
Phe Asp Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly
Glu Ala Ile Asn Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp
Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr
Gly Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn Met Arg Gly Leu
Pro Leu Ser Ala Glu Phe Thr Thr 85 90 9512896PRTArtificial
SequenceEGFR binding FN3 domain 128Met Leu Pro Ala Pro Lys Asn Leu
Val Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Leu Arg Leu Ser Trp Asp
Tyr Asp Leu Gly Val Tyr Phe Asp Ser 20 25 30Phe Leu Ile Gln Tyr Gln
Glu Ser Glu Lys Val Gly Glu Ala Ile Asn 35 40 45Leu Thr Val Pro Gly
Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys 50 55 60Pro Gly Thr Glu
Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr65 70 75 80Lys Asp
Thr Asn Met Arg Gly Leu Pro Leu Ser Ala Glu Phe Thr Thr 85 90
9512995PRTArtificial SequenceEGFR binding FN3 domain 129Met Leu Pro
Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Leu
Arg Leu Ser Trp Asp Asp Pro Trp Ala Phe Tyr Glu Ser Phe 20 25 30Leu
Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Asn Leu 35 40
45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro
50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr
Lys65 70 75 80Asp Thr Asn Met Arg Gly Leu Pro Leu Ser Ala Glu Phe
Thr Thr 85 90 9513095PRTArtificial SequenceEGFR binding FN3 domain
130Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1
5 10 15Ser Leu Arg Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser
Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile
Asn Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly
Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val Leu
Gly Ser Tyr Val65 70 75 80Phe Glu His Asp Val Met Leu Pro Leu Ser
Ala Glu Phe Thr Thr 85 90 9513195PRTArtificial SequenceEGFR binding
FN3 domain 131Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val
Thr Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Asp Asp Pro Trp Ala Phe
Tyr Glu Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly
Glu Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp
Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr
Gly Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn Met Arg Gly Leu
Pro Leu Ser Ala Ile Phe Thr Thr 85 90 9513295PRTArtificial
SequenceEGFR binding FN3 domain 132Met Leu Pro Ala Pro Lys Asn Leu
Val Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Thr
Ala Pro Asp Ala Ala Phe Asp Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu
Ser Glu Lys Val Gly Glu Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser
Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr
Thr Val Ser Ile Tyr Gly Val Leu Gly Ser Tyr Val65 70 75 80Phe Glu
His Asp Val Met Leu Pro Leu Ser Ala Ile Phe Thr Thr 85 90
9513395PRTArtificial SequenceEGFR binding FN3 domain 133Met Leu Pro
Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Leu
Arg Leu Ser Trp Thr Trp Pro Tyr Ala Asp Leu Asp Ser Phe 20 25 30Leu
Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Asn Leu 35 40
45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro
50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr
Lys65 70 75 80Asp Thr Asn Met Arg Gly Leu Pro Leu Ser Ala Glu Phe
Thr Thr 85 90 9513495PRTArtificial SequenceEGFR binding FN3 domain
134Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1
5 10 15Ser Ala Arg Leu Ser Trp Ala Asp Pro His Gly Phe Tyr Asp Ser
Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile
Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly
Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His
Asn Val Tyr Lys65 70 75 80Asp Thr Asn Met Arg Gly Leu Pro Leu Ser
Ala Ile Phe Thr Thr 85 90 9513595PRTArtificial SequenceEGFR binding
FN3 domain 135Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val
Thr Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Asp Asp Pro Trp Ala Phe
Tyr Glu Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly
Glu Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp
Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr
Gly Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn Ile Arg Gly Leu
Pro Leu Ser Ala Ile Phe Thr Thr 85 90 9513695PRTArtificial
SequenceEGFR binding FN3 domain 136Met Leu Pro Ala Pro Lys Asn Leu
Val Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Asp
Asp Pro His Ala Phe Tyr Glu Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu
Ser Glu Lys Val Gly Glu Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser
Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr
Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys65 70 75 80Asp Thr
Asn Ile Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr 85 90
9513795PRTArtificial SequenceEGFR binding FN3 domain 137Met Leu Pro
Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Ala
Arg Leu Ser Trp Ala Asp Pro His Gly Phe Tyr Asp Ser Phe 20 25 30Leu
Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Val Leu 35 40
45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro
50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr
Lys65 70 75 80Asp Thr Asn Ile Arg Gly Leu Pro Leu Ser Ala Ile Phe
Thr Thr 85 90 95138204PRTArtificial SequenceBispecific EGFR/c-Met
binding domain 138Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val
Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser Trp Asp Asp Pro Trp Ala Phe
Tyr Glu Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly
Glu Ala Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp
Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr
Gly Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Met Arg Gly Leu
Pro Leu Ser Ala Ile Phe Thr Thr Gly Gly 85 90 95Gly Gly Ser Gly Gly
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly 100 105 110Gly Ser Met
Leu Pro Ala Pro Lys Asn Leu Val Val Ser Arg Val Thr 115 120 125Glu
Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp 130 135
140Ser Phe Trp Ile Arg Tyr Asp Glu Val Val Val Gly Gly Glu Ala
Ile145 150 155 160Val Leu Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp
Leu Thr Gly Leu 165 170 175Lys Pro Gly Thr Glu Tyr Tyr Val Asn Ile
Leu Gly Val Lys Gly Gly 180 185 190Ser Ile Ser Val Pro Leu Ser Ala
Ile Phe Thr Thr 195 200139203PRTArtificial SequenceBispecific
EGFR/c-Met binding domain 139Leu Pro Ala Pro Lys Asn Leu Val Val
Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser Trp Asp Asp Pro
Trp Ala Phe Tyr Glu Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu
Lys Val Gly Glu Ala Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg
Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val
Ser Ile Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Met
Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Gly Gly 85 90 95Gly Gly
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly 100 105
110Gly Ser Leu Pro Ala Pro Lys Asn Leu Val Val Ser Arg Val Thr Glu
115 120 125Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe
Asp Ser 130 135 140Phe Trp Ile Arg Tyr Phe Glu Phe Leu Gly Ser Gly
Glu Ala Ile Val145 150 155 160Leu Thr Val Pro Gly Ser Glu Arg Ser
Tyr Asp Leu Thr Gly Leu Lys 165 170 175Pro Gly Thr Glu Tyr Val Val
Asn Ile Met Gly Val Lys Gly Gly Lys 180 185 190Ile Ser Pro Pro Leu
Ser Ala Ile Phe Thr Thr 195 200140204PRTArtificial
SequenceBispecific EGFR/c-Met binding domain 140Leu Pro Ala Pro Lys
Asn Leu Val Val Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser
Trp Asp Asp Pro Trp Ala Phe Tyr Glu Ser Phe Leu 20 25 30Ile Gln Tyr
Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Val Leu Thr 35 40 45Val Pro
Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr
Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75
80Thr Asn Met Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Gly Gly
85 90 95Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
Gly 100 105 110Gly Ser Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser
Arg Val Thr 115 120 125Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro
Asp Ala Ala Phe Asp 130 135 140Ser Phe Trp Ile Arg Tyr Phe Glu Phe
Leu Gly Ser Gly Glu Ala Ile145 150 155 160Val Leu Thr Val Pro Gly
Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu 165 170 175Lys Pro Gly Thr
Glu Tyr Val Val Gln Ile Ile Gly Val Lys Gly Gly 180 185 190His Ile
Ser Leu Pro Leu Ser Ala Ile Phe Thr Thr 195 200141204PRTArtificial
SequenceBispecific EGFR/c-Met binding domain 141Leu Pro Ala Pro Lys
Asn Leu Val Val Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser
Trp Asp Asp Pro Trp Ala Phe Tyr Glu Ser Phe Leu 20 25 30Ile Gln Tyr
Gln Glu Ser Glu Lys Val
Gly Glu Ala Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr
Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile
Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Met Arg Gly
Leu Pro Leu Ser Ala Ile Phe Thr Thr Gly Gly 85 90 95Gly Gly Ser Gly
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly 100 105 110Gly Ser
Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Arg Val Thr 115 120
125Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp
130 135 140Ser Phe Phe Ile Arg Tyr Asp Glu Phe Leu Arg Ser Gly Glu
Ala Ile145 150 155 160Val Leu Thr Val Pro Gly Ser Glu Arg Ser Tyr
Asp Leu Thr Gly Leu 165 170 175Lys Pro Gly Thr Glu Tyr Trp Val Thr
Ile Leu Gly Val Lys Gly Gly 180 185 190Leu Val Ser Thr Pro Leu Ser
Ala Ile Phe Thr Thr 195 200142204PRTArtificial SequenceBispecific
EGFR/c-Met binding domain 142Leu Pro Ala Pro Lys Asn Leu Val Val
Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser Trp Ala Asp Pro
His Gly Phe Tyr Asp Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu
Lys Val Gly Glu Ala Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg
Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val
Ser Ile Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Met
Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Gly Gly 85 90 95Gly Gly
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly 100 105
110Gly Ser Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Arg Val Thr
115 120 125Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro Asp Ala Ala
Phe Asp 130 135 140Ser Phe Trp Ile Arg Tyr Phe Glu Phe Leu Gly Ser
Gly Glu Ala Ile145 150 155 160Val Leu Thr Val Pro Gly Ser Glu Arg
Ser Tyr Asp Leu Thr Gly Leu 165 170 175Lys Pro Gly Thr Glu Tyr Val
Val Asn Ile Met Gly Val Lys Gly Gly 180 185 190Lys Ile Ser Pro Pro
Leu Ser Ala Ile Phe Thr Thr 195 200143204PRTArtificial
SequenceBispecific EGFR/c-Met binding domain 143Leu Pro Ala Pro Lys
Asn Leu Val Val Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser
Trp Ala Asp Pro His Gly Phe Tyr Asp Ser Phe Leu 20 25 30Ile Gln Tyr
Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Val Leu Thr 35 40 45Val Pro
Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr
Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75
80Thr Asn Met Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Gly Gly
85 90 95Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
Gly 100 105 110Gly Ser Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser
Arg Val Thr 115 120 125Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro
Asp Ala Ala Phe Asp 130 135 140Ser Phe Trp Ile Arg Tyr Phe Glu Phe
Leu Gly Ser Gly Glu Ala Ile145 150 155 160Val Leu Thr Val Pro Gly
Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu 165 170 175Lys Pro Gly Thr
Glu Tyr Val Val Gln Ile Ile Gly Val Lys Gly Gly 180 185 190His Ile
Ser Leu Pro Leu Ser Ala Ile Phe Thr Thr 195 200144204PRTArtificial
SequenceBispecific EGFR/c-Met binding domain 144Leu Pro Ala Pro Lys
Asn Leu Val Val Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser
Trp Ala Asp Pro His Gly Phe Tyr Asp Ser Phe Leu 20 25 30Ile Gln Tyr
Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Val Leu Thr 35 40 45Val Pro
Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr
Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75
80Thr Asn Met Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Gly Gly
85 90 95Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
Gly 100 105 110Gly Ser Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser
Arg Val Thr 115 120 125Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro
Asp Ala Ala Phe Asp 130 135 140Ser Phe Trp Ile Arg Tyr Phe Glu Phe
Leu Gly Ser Gly Glu Ala Ile145 150 155 160Val Leu Thr Val Pro Gly
Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu 165 170 175Lys Pro Gly Thr
Glu Tyr Val Val Gln Ile Ile Gly Val Lys Gly Gly 180 185 190His Ile
Ser Leu Pro Leu Ser Ala Ile Phe Thr Thr 195 200145193PRTArtificial
SequenceBispecific EGFR/c-Met binding domain 145Leu Pro Ala Pro Lys
Asn Leu Val Val Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser
Trp Asp Asp Pro Trp Ala Phe Tyr Glu Ser Phe Leu 20 25 30Ile Gln Tyr
Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Val Leu Thr 35 40 45Val Pro
Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr
Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75
80Thr Asn Met Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala Pro
85 90 95Ala Pro Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys Asn Leu
Val 100 105 110Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu Ser Trp
Thr Ala Pro 115 120 125Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg Tyr
Phe Glu Phe Leu Gly 130 135 140Ser Gly Glu Ala Ile Val Leu Thr Val
Pro Gly Ser Glu Arg Ser Tyr145 150 155 160Asp Leu Thr Gly Leu Lys
Pro Gly Thr Glu Tyr Val Val Asn Ile Met 165 170 175Gly Val Lys Gly
Gly Lys Ile Ser Pro Pro Leu Ser Ala Ile Phe Thr 180 185
190Thr146193PRTArtificial SequenceBispecific EGFR/c-Met binding
domain 146Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu
Asp Ser1 5 10 15Ala Arg Leu Ser Trp Asp Asp Pro Trp Ala Phe Tyr Glu
Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala
Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr
Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val
His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Met Arg Gly Leu Pro Leu
Ser Ala Ile Phe Thr Thr Ala Pro 85 90 95Ala Pro Ala Pro Ala Pro Ala
Pro Leu Pro Ala Pro Lys Asn Leu Val 100 105 110Val Ser Arg Val Thr
Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro 115 120 125Asp Ala Ala
Phe Asp Ser Phe Trp Ile Arg Tyr Asp Glu Val Val Val 130 135 140Gly
Gly Glu Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser Tyr145 150
155 160Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Tyr Val Asn Ile
Leu 165 170 175Gly Val Lys Gly Gly Ser Ile Ser Val Pro Leu Ser Ala
Ile Phe Thr 180 185 190Thr147194PRTArtificial SequenceBispecific
EGFR/c-Met binding domain 147Leu Pro Ala Pro Lys Asn Leu Val Val
Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser Trp Ala Asp Pro
His Gly Phe Tyr Asp Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu
Lys Val Gly Glu Ala Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg
Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val
Ser Ile Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Met
Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala Pro 85 90 95Ala Pro
Ala Pro Ala Pro Ala Pro Met Leu Pro Ala Pro Lys Asn Leu 100 105
110Val Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala
115 120 125Pro Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu
Phe Leu 130 135 140Gly Ser Gly Glu Ala Ile Val Leu Thr Val Pro Gly
Ser Glu Arg Ser145 150 155 160Tyr Asp Leu Thr Gly Leu Lys Pro Gly
Thr Glu Tyr Val Val Asn Ile 165 170 175Met Gly Val Lys Gly Gly Lys
Ile Ser Pro Pro Leu Ser Ala Ile Phe 180 185 190Thr
Thr148193PRTArtificial SequenceBispecific EGFR/c-Met binding domain
148Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp Ser1
5 10 15Ala Arg Leu Ser Trp Ala Asp Pro His Gly Phe Tyr Asp Ser Phe
Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Val
Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu
Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn
Val Tyr Lys Asp65 70 75 80Thr Asn Met Arg Gly Leu Pro Leu Ser Ala
Ile Phe Thr Thr Ala Pro 85 90 95Ala Pro Ala Pro Ala Pro Ala Pro Leu
Pro Ala Pro Lys Asn Leu Val 100 105 110Val Ser Arg Val Thr Glu Asp
Ser Ala Arg Leu Ser Trp Thr Ala Pro 115 120 125Asp Ala Ala Phe Asp
Ser Phe Trp Ile Arg Tyr Asp Glu Val Val Val 130 135 140Gly Gly Glu
Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser Tyr145 150 155
160Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Tyr Val Asn Ile Leu
165 170 175Gly Val Lys Gly Gly Ser Ile Ser Val Pro Leu Ser Ala Ile
Phe Thr 180 185 190Thr149193PRTArtificial SequenceBispecific
EGFR/c-Met binding domain 149Leu Pro Ala Pro Lys Asn Leu Val Val
Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser Trp Asp Asp Pro
Trp Ala Phe Tyr Glu Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu
Lys Val Gly Glu Ala Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg
Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val
Ser Ile Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Ile
Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala Pro 85 90 95Ala Pro
Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys Asn Leu Val 100 105
110Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro
115 120 125Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe
Leu Gly 130 135 140Ser Gly Glu Ala Ile Val Leu Thr Val Pro Gly Ser
Glu Arg Ser Tyr145 150 155 160Asp Leu Thr Gly Leu Lys Pro Gly Thr
Glu Tyr Val Val Asn Ile Leu 165 170 175Gly Val Lys Gly Gly Lys Ile
Ser Pro Pro Leu Ser Ala Ile Phe Thr 180 185
190Thr150193PRTArtificial SequenceBispecific EGFR/c-Met binding
domain 150Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu
Asp Ser1 5 10 15Ala Arg Leu Ser Trp Asp Asp Pro Trp Ala Phe Tyr Glu
Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala
Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr
Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val
His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Ile Arg Gly Leu Pro Leu
Ser Ala Ile Phe Thr Thr Ala Pro 85 90 95Ala Pro Ala Pro Ala Pro Ala
Pro Leu Pro Ala Pro Lys Asn Leu Val 100 105 110Val Ser Arg Val Thr
Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro 115 120 125Asp Ala Ala
Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Val Gly 130 135 140Ser
Gly Glu Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser Tyr145 150
155 160Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile
Leu 165 170 175Gly Val Lys Gly Gly Ser Ile Ser Pro Pro Leu Ser Ala
Ile Phe Thr 180 185 190Thr151193PRTArtificial SequenceBispecific
EGFR/c-Met binding domain 151Leu Pro Ala Pro Lys Asn Leu Val Val
Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser Trp Asp Asp Pro
Trp Ala Phe Tyr Glu Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu
Lys Val Gly Glu Ala Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg
Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val
Ser Ile Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Ile
Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala Pro 85 90 95Ala Pro
Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys Asn Leu Val 100 105
110Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro
115 120 125Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe
Val Ser 130 135 140Lys Gly Asp Ala Ile Val Leu Thr Val Pro Gly Ser
Glu Arg Ser Tyr145 150 155 160Asp Leu Thr Gly Leu Lys Pro Gly Thr
Glu Tyr Val Val Asn Ile Leu 165 170 175Gly Val Lys Gly Gly Ser Ile
Ser Pro Pro Leu Ser Ala Ile Phe Thr 180 185
190Thr152193PRTArtificial SequenceBispecific EGFR/c-Met binding
domain 152Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu
Asp Ser1 5 10 15Ala Arg Leu Ser Trp Asp Asp Pro Trp Ala Phe Tyr Glu
Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala
Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr
Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val
His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Ile Arg Gly Leu Pro Leu
Ser Ala Ile Phe Thr Thr Ala Pro 85 90 95Ala Pro Ala Pro Ala Pro Ala
Pro Leu Pro Ala Pro Lys Asn Leu Val 100 105 110Val Ser Arg Val Thr
Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro 115 120 125Asp Ala Ala
Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Leu Gly 130 135 140Ser
Gly Glu Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser Tyr145 150
155 160Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile
Leu 165 170 175Ser Val Lys Gly Gly Ser Ile Ser Pro Pro Leu Ser Ala
Ile Phe Thr 180 185 190Thr153193PRTArtificial SequenceBispecific
EGFR/c-Met binding domain 153Leu Pro Ala Pro Lys Asn Leu Val Val
Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser Trp Asp Asp Pro
His Ala Phe Tyr Glu Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu
Lys Val Gly Glu Ala Ile Val Leu Thr 35 40
45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly
50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys
Asp65 70 75 80Thr Asn Ile Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr
Thr Ala Pro 85 90 95Ala Pro Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro
Lys Asn Leu Val 100 105 110Val Ser Arg Val Thr Glu Asp Ser Ala Arg
Leu Ser Trp Thr Ala Pro 115 120 125Asp Ala Ala Phe Asp Ser Phe Trp
Ile Arg Tyr Phe Glu Phe Leu Gly 130 135 140Ser Gly Glu Ala Ile Val
Leu Thr Val Pro Gly Ser Glu Arg Ser Tyr145 150 155 160Asp Leu Thr
Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile Leu 165 170 175Gly
Val Lys Gly Gly Lys Ile Ser Pro Pro Leu Ser Ala Ile Phe Thr 180 185
190Thr154193PRTArtificial SequenceBispecific EGFR/c-Met binding
domain 154Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu
Asp Ser1 5 10 15Ala Arg Leu Ser Trp Asp Asp Pro His Ala Phe Tyr Glu
Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala
Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr
Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val
His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Ile Arg Gly Leu Pro Leu
Ser Ala Ile Phe Thr Thr Ala Pro 85 90 95Ala Pro Ala Pro Ala Pro Ala
Pro Leu Pro Ala Pro Lys Asn Leu Val 100 105 110Val Ser Arg Val Thr
Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro 115 120 125Asp Ala Ala
Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Val Gly 130 135 140Ser
Gly Glu Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser Tyr145 150
155 160Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile
Leu 165 170 175Gly Val Lys Gly Gly Ser Ile Ser Pro Pro Leu Ser Ala
Ile Phe Thr 180 185 190Thr155193PRTArtificial SequenceBispecific
EGFR/c-Met binding domain 155Leu Pro Ala Pro Lys Asn Leu Val Val
Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser Trp Asp Asp Pro
His Ala Phe Tyr Glu Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu
Lys Val Gly Glu Ala Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg
Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val
Ser Ile Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Ile
Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala Pro 85 90 95Ala Pro
Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys Asn Leu Val 100 105
110Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro
115 120 125Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe
Val Ser 130 135 140Lys Gly Asp Ala Ile Val Leu Thr Val Pro Gly Ser
Glu Arg Ser Tyr145 150 155 160Asp Leu Thr Gly Leu Lys Pro Gly Thr
Glu Tyr Val Val Asn Ile Leu 165 170 175Gly Val Lys Gly Gly Ser Ile
Ser Pro Pro Leu Ser Ala Ile Phe Thr 180 185
190Thr156193PRTArtificial SequenceBispecific EGFR/c-Met binding
domain 156Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu
Asp Ser1 5 10 15Ala Arg Leu Ser Trp Asp Asp Pro His Ala Phe Tyr Glu
Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala
Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr
Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val
His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Ile Arg Gly Leu Pro Leu
Ser Ala Ile Phe Thr Thr Ala Pro 85 90 95Ala Pro Ala Pro Ala Pro Ala
Pro Leu Pro Ala Pro Lys Asn Leu Val 100 105 110Val Ser Arg Val Thr
Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro 115 120 125Asp Ala Ala
Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Leu Gly 130 135 140Ser
Gly Glu Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser Tyr145 150
155 160Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile
Leu 165 170 175Ser Val Lys Gly Gly Ser Ile Ser Pro Pro Leu Ser Ala
Ile Phe Thr 180 185 190Thr157193PRTArtificial SequenceBispecific
EGFR/c-Met binding domain 157Leu Pro Ala Pro Lys Asn Leu Val Val
Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser Trp Ala Asp Pro
His Gly Phe Tyr Asp Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu
Lys Val Gly Glu Ala Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg
Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val
Ser Ile Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Ile
Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala Pro 85 90 95Ala Pro
Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys Asn Leu Val 100 105
110Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro
115 120 125Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe
Leu Gly 130 135 140Ser Gly Glu Ala Ile Val Leu Thr Val Pro Gly Ser
Glu Arg Ser Tyr145 150 155 160Asp Leu Thr Gly Leu Lys Pro Gly Thr
Glu Tyr Val Val Asn Ile Leu 165 170 175Gly Val Lys Gly Gly Lys Ile
Ser Pro Pro Leu Ser Ala Ile Phe Thr 180 185
190Thr158193PRTArtificial SequenceBispecific EGFR/c-Met binding
domain 158Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu
Asp Ser1 5 10 15Ala Arg Leu Ser Trp Ala Asp Pro His Gly Phe Tyr Asp
Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala
Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr
Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val
His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Ile Arg Gly Leu Pro Leu
Ser Ala Ile Phe Thr Thr Ala Pro 85 90 95Ala Pro Ala Pro Ala Pro Ala
Pro Leu Pro Ala Pro Lys Asn Leu Val 100 105 110Val Ser Arg Val Thr
Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro 115 120 125Asp Ala Ala
Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Val Gly 130 135 140Ser
Gly Glu Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser Tyr145 150
155 160Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile
Leu 165 170 175Gly Val Lys Gly Gly Ser Ile Ser Pro Pro Leu Ser Ala
Ile Phe Thr 180 185 190Thr159193PRTArtificial SequenceBispecific
EGFR/c-Met binding domain 159Leu Pro Ala Pro Lys Asn Leu Val Val
Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser Trp Ala Asp Pro
His Gly Phe Tyr Asp Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu
Lys Val Gly Glu Ala Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg
Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val
Ser Ile Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Ile
Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala Pro 85 90 95Ala Pro
Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys Asn Leu Val 100 105
110Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro
115 120 125Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe
Val Ser 130 135 140Lys Gly Asp Ala Ile Val Leu Thr Val Pro Gly Ser
Glu Arg Ser Tyr145 150 155 160Asp Leu Thr Gly Leu Lys Pro Gly Thr
Glu Tyr Val Val Asn Ile Leu 165 170 175Gly Val Lys Gly Gly Ser Ile
Ser Pro Pro Leu Ser Ala Ile Phe Thr 180 185
190Thr160193PRTArtificial SequenceBispecific EGFR/c-Met binding
domain 160Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu
Asp Ser1 5 10 15Ala Arg Leu Ser Trp Ala Asp Pro His Gly Phe Tyr Asp
Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala
Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr
Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val
His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Ile Arg Gly Leu Pro Leu
Ser Ala Ile Phe Thr Thr Ala Pro 85 90 95Ala Pro Ala Pro Ala Pro Ala
Pro Leu Pro Ala Pro Lys Asn Leu Val 100 105 110Val Ser Arg Val Thr
Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro 115 120 125Asp Ala Ala
Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Leu Gly 130 135 140Ser
Gly Glu Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser Tyr145 150
155 160Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile
Leu 165 170 175Ser Val Lys Gly Gly Ser Ile Ser Pro Pro Leu Ser Ala
Ile Phe Thr 180 185 190Thr161193PRTArtificial SequenceBispecific
EGFR/c-Met binding domain 161Leu Pro Ala Pro Lys Asn Leu Val Val
Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser Trp Asp Asp Pro
Trp Ala Phe Tyr Glu Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu
Lys Val Gly Glu Ala Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg
Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val
Ser Ile Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Ile
Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala Pro 85 90 95Ala Pro
Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys Asn Leu Val 100 105
110Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro
115 120 125Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe
Leu Gly 130 135 140Ser Gly Glu Ala Ile Val Leu Thr Val Pro Gly Ser
Glu Arg Ser Tyr145 150 155 160Asp Leu Thr Gly Leu Lys Pro Gly Thr
Glu Tyr Val Val Asn Ile Leu 165 170 175Ser Val Lys Gly Gly Ser Ile
Ser Pro Pro Leu Ser Ala Ile Phe Thr 180 185
190Thr162249PRTArtificial SequenceBispecific EGFR/c-Met binding
domain 162Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu
Asp Ser1 5 10 15Ala Arg Leu Ser Trp Asp Asp Pro Trp Ala Phe Tyr Glu
Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala
Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr
Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val
His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Met Arg Gly Leu Pro Leu
Ser Ala Ile Phe Thr Thr Ala Pro 85 90 95Ala Pro Ala Pro Ala Pro Ala
Pro Leu Pro Ala Pro Lys Asn Leu Val 100 105 110Val Ser Arg Val Thr
Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro 115 120 125Asp Ala Ala
Phe Asp Ser Phe Trp Ile Arg Tyr Asp Glu Val Val Val 130 135 140Gly
Gly Glu Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser Tyr145 150
155 160Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Tyr Val Asn Ile
Leu 165 170 175Gly Val Lys Gly Gly Ser Ile Ser Val Pro Leu Ser Ala
Ile Phe Thr 180 185 190Thr Ala Pro Ala Pro Ala Pro Ala Pro Ala Pro
Leu Ala Glu Ala Lys 195 200 205Val Leu Ala Asn Arg Glu Leu Asp Lys
Tyr Gly Val Ser Asp Tyr Tyr 210 215 220Lys Asn Leu Ile Asn Asn Ala
Lys Thr Val Glu Gly Val Lys Ala Leu225 230 235 240Leu Asp Glu Ile
Leu Ala Ala Leu Pro 245163249PRTArtificial SequenceBispecific
EGFR/c-Met binding domain 163Leu Pro Ala Pro Lys Asn Leu Val Val
Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser Trp Ala Asp Pro
His Gly Phe Tyr Asp Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu
Lys Val Gly Glu Ala Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg
Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val
Ser Ile Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Met
Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala Pro 85 90 95Ala Pro
Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys Asn Leu Val 100 105
110Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro
115 120 125Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg Tyr Asp Glu Val
Val Val 130 135 140Gly Gly Glu Ala Ile Val Leu Thr Val Pro Gly Ser
Glu Arg Ser Tyr145 150 155 160Asp Leu Thr Gly Leu Lys Pro Gly Thr
Glu Tyr Tyr Val Asn Ile Leu 165 170 175Gly Val Lys Gly Gly Ser Ile
Ser Val Pro Leu Ser Ala Ile Phe Thr 180 185 190Thr Ala Pro Ala Pro
Ala Pro Ala Pro Ala Pro Leu Ala Glu Ala Lys 195 200 205Val Leu Ala
Asn Arg Glu Leu Asp Lys Tyr Gly Val Ser Asp Tyr Tyr 210 215 220Lys
Asn Leu Ile Asn Asn Ala Lys Thr Val Glu Gly Val Lys Ala Leu225 230
235 240Leu Asp Glu Ile Leu Ala Ala Leu Pro 245164249PRTArtificial
SequenceBispecific EGFR/c-Met binding domain 164Leu Pro Ala Pro Lys
Asn Leu Val Val Ser Glu Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser
Trp Asp Asp Pro Trp Ala Phe Tyr Glu Ser Phe Leu 20 25 30Ile Gln Tyr
Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Val Leu Thr 35 40 45Val Pro
Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr
Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys Asp65 70 75
80Thr Asn Met Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala Pro
85 90 95Ala Pro Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys Asn Leu
Val 100 105 110Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu Ser Trp
Thr Ala Pro 115 120 125Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg Tyr
Phe Glu Phe Leu Gly 130 135 140Ser Gly Glu Ala Ile Val Leu Thr Val
Pro Gly Ser Glu Arg Ser Tyr145 150 155 160Asp Leu Thr Gly Leu Lys
Pro Gly Thr Glu Tyr Val Val Asn Ile Met 165 170 175Gly Val Lys Gly
Gly Lys Ile Ser Pro Pro Leu Ser Ala Ile Phe Thr 180
185 190Thr Ala Pro Ala Pro Ala Pro Ala Pro Ala Pro Leu Ala Glu Ala
Lys 195 200 205Val Leu Ala Asn Arg Glu Leu Asp Lys Tyr Gly Val Ser
Asp Tyr Tyr 210 215 220Lys Asn Leu Ile Asn Asn Ala Lys Thr Val Glu
Gly Val Lys Ala Leu225 230 235 240Leu Asp Glu Ile Leu Ala Ala Leu
Pro 245165249PRTArtificial SequenceBispecific EGFR/c-Met binding
domain 165Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu
Asp Ser1 5 10 15Ala Arg Leu Ser Trp Ala Asp Pro His Gly Phe Tyr Asp
Ser Phe Leu 20 25 30Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala
Ile Val Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr
Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val
His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Met Arg Gly Leu Pro Leu
Ser Ala Ile Phe Thr Thr Ala Pro 85 90 95Ala Pro Ala Pro Ala Pro Ala
Pro Leu Pro Ala Pro Lys Asn Leu Val 100 105 110Val Ser Arg Val Thr
Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala Pro 115 120 125Asp Ala Ala
Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Leu Gly 130 135 140Ser
Gly Glu Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser Tyr145 150
155 160Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile
Met 165 170 175Gly Val Lys Gly Gly Lys Ile Ser Pro Pro Leu Ser Ala
Ile Phe Thr 180 185 190Thr Ala Pro Ala Pro Ala Pro Ala Pro Ala Pro
Leu Ala Glu Ala Lys 195 200 205Val Leu Ala Asn Arg Glu Leu Asp Lys
Tyr Gly Val Ser Asp Tyr Tyr 210 215 220Lys Asn Leu Ile Asn Asn Ala
Lys Thr Val Glu Gly Val Lys Ala Leu225 230 235 240Leu Asp Glu Ile
Leu Ala Ala Leu Pro 245166588DNAArtificial SequenceBispecific
EGFR/c-Met binding domain 166ttgccagcgc cgaagaacct ggtagttagc
gaggttactg aggacagcgc gcgtctgagc 60tgggacgatc cgtgggcgtt ctacgagagc
tttctgatcc agtatcaaga gagcgagaaa 120gtcggtgaag cgattgtgct
gaccgtcccg ggctccgagc gttcctacga cctgaccggt 180ttgaagccgg
gtaccgagta tacggtgagc atctacggtg ttcacaatgt ctataaggac
240actaatatcc gcggtctgcc tctgagcgcc attttcacca ccgcaccggc
accggctccg 300gctcctgccc cgctgccggc tccgaagaac ttggtggtga
gccgtgttac cgaagatagc 360gcacgcctga gctggacggc accggatgcg
gcgttcgata gcttctggat tcgctatttt 420gagtttctgg gtagcggtga
ggcaattgtt ctgacggtgc cgggctctga acgctcctac 480gatttgaccg
gtctgaaacc gggcaccgag tatgtggtga acattctgag cgttaagggc
540ggtagcatca gcccaccgct gagcgcgatc ttcacgactg gtggttgc
588167579DNAArtificial SequenceBispecific EGFR/c-Met binding domain
167ctgccagccc ctaagaatct ggtcgtgagc gaagtaaccg aggacagcgc
ccgcctgagc 60tgggacgacc cgtgggcgtt ctatgagtct ttcctgattc agtatcaaga
aagcgaaaaa 120gttggcgaag cgatcgtcct gaccgtcccg ggtagcgagc
gctcctacga tctgaccggc 180ctgaaaccgg gtacggagta cacggtgtcc
atttacggtg ttcacaatgt gtataaagac 240accaacatgc gtggcctgcc
gctgtcggcg attttcacca ccgcgcctgc gccagcgcct 300gcaccggctc
cgctgccggc accgaagaac ctggttgtca gccgtgtgac cgaggatagc
360gcacgtttga gctggaccgc tccggatgca gcctttgaca gcttctggat
tcgttacttt 420gaatttctgg gtagcggtga ggcgatcgtt ctgacggtgc
cgggctctga acgcagctat 480gatttgacgg gcctgaagcc gggtactgag
tacgtggtta acatcatggg cgttaagggt 540ggtaaaatca gcccgccatt
gtccgcgatc tttaccacg 579168282DNAArtificial SequenceEGFR binding
FN3 domain from ECB97 168ttgccagcgc cgaagaacct ggtagttagc
gaggttactg aggacagcgc gcgtctgagc 60tgggacgatc cgtgggcgtt ctacgagagc
tttctgatcc agtatcaaga gagcgagaaa 120gtcggtgaag cgattgtgct
gaccgtcccg ggctccgagc gttcctacga cctgaccggt 180ttgaagccgg
gtaccgagta tacggtgagc atctacggtg ttcacaatgt ctataaggac
240actaatatcc gcggtctgcc tctgagcgcc attttcacca cc
282169282DNAArtificial SequenceEGFR binding FN3 domain from ECB15
169ctgccagccc ctaagaatct ggtcgtgagc gaagtaaccg aggacagcgc
ccgcctgagc 60tgggacgacc cgtgggcgtt ctatgagtct ttcctgattc agtatcaaga
aagcgaaaaa 120gttggcgaag cgatcgtcct gaccgtcccg ggtagcgagc
gctcctacga tctgaccggc 180ctgaaaccgg gtacggagta cacggtgtcc
atttacggtg ttcacaatgt gtataaagac 240accaacatgc gtggcctgcc
gctgtcggcg attttcacca cc 282170195PRTArtificial SequenceBispecific
EGFR/c-Met binding molecule 170Met Leu Pro Ala Pro Lys Asn Leu Val
Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Asp Asp
Pro Trp Ala Phe Tyr Glu Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser
Glu Lys Val Gly Glu Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser Glu
Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr
Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn
Ile Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala
Pro Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys Asn Leu 100 105
110Val Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala
115 120 125Pro Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu
Phe Leu 130 135 140Gly Ser Gly Glu Ala Ile Val Leu Thr Val Pro Gly
Ser Glu Arg Ser145 150 155 160Tyr Asp Leu Thr Gly Leu Lys Pro Gly
Thr Glu Tyr Val Val Asn Ile 165 170 175Leu Gly Val Lys Gly Gly Lys
Ile Ser Pro Pro Leu Ser Ala Ile Phe 180 185 190Thr Thr Cys
195171195PRTArtificial SequenceBispecific EGFR/c-Met binding
molecule 171Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr
Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Asp Asp Pro Trp Ala Phe Tyr
Glu Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu
Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu
Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly
Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn Ile Arg Gly Leu Pro
Leu Ser Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala Pro Ala Pro Ala Pro
Ala Pro Leu Pro Ala Pro Lys Asn Leu 100 105 110Val Val Ser Arg Val
Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala 115 120 125Pro Asp Ala
Ala Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Val 130 135 140Gly
Ser Gly Glu Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser145 150
155 160Tyr Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn
Ile 165 170 175Leu Gly Val Lys Gly Gly Ser Ile Ser Pro Pro Leu Ser
Ala Ile Phe 180 185 190Thr Thr Cys 195172195PRTArtificial
SequenceBispecific EGFR/c-Met binding molecule 172Met Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Ala Arg
Leu Ser Trp Asp Asp Pro Trp Ala Phe Tyr Glu Ser Phe 20 25 30Leu Ile
Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Val Leu 35 40 45Thr
Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55
60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys65
70 75 80Asp Thr Asn Ile Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr
Ala 85 90 95Pro Ala Pro Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys
Asn Leu 100 105 110Val Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu
Ser Trp Thr Ala 115 120 125Pro Asp Ala Ala Phe Asp Ser Phe Trp Ile
Arg Tyr Phe Glu Phe Val 130 135 140Ser Lys Gly Asp Ala Ile Val Leu
Thr Val Pro Gly Ser Glu Arg Ser145 150 155 160Tyr Asp Leu Thr Gly
Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile 165 170 175Leu Gly Val
Lys Gly Gly Ser Ile Ser Pro Pro Leu Ser Ala Ile Phe 180 185 190Thr
Thr Cys 195173195PRTArtificial SequenceBispecific EGFR/c-Met
binding molecule 173Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu
Val Thr Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Asp Asp Pro Trp Ala
Phe Tyr Glu Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val
Gly Glu Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr
Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile
Tyr Gly Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn Ile Arg Gly
Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala Pro Ala Pro
Ala Pro Ala Pro Leu Pro Ala Pro Lys Asn Leu 100 105 110Val Val Ser
Arg Val Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala 115 120 125Pro
Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Leu 130 135
140Gly Ser Gly Glu Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg
Ser145 150 155 160Tyr Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr
Val Val Asn Ile 165 170 175Leu Ser Val Lys Gly Gly Ser Ile Ser Pro
Pro Leu Ser Ala Ile Phe 180 185 190Thr Thr Cys
195174195PRTArtificial SequenceBispecific EGFR/c-Met binding
molecule 174Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr
Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Asp Asp Pro His Ala Phe Tyr
Glu Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu
Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu
Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly
Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn Ile Arg Gly Leu Pro
Leu Ser Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala Pro Ala Pro Ala Pro
Ala Pro Leu Pro Ala Pro Lys Asn Leu 100 105 110Val Val Ser Arg Val
Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala 115 120 125Pro Asp Ala
Ala Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Leu 130 135 140Gly
Ser Gly Glu Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser145 150
155 160Tyr Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn
Ile 165 170 175Leu Gly Val Lys Gly Gly Lys Ile Ser Pro Pro Leu Ser
Ala Ile Phe 180 185 190Thr Thr Cys 195175195PRTArtificial
SequenceBispecific EGFR/c-Met binding molecule 175Met Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Ala Arg
Leu Ser Trp Asp Asp Pro His Ala Phe Tyr Glu Ser Phe 20 25 30Leu Ile
Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Val Leu 35 40 45Thr
Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55
60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys65
70 75 80Asp Thr Asn Ile Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr
Ala 85 90 95Pro Ala Pro Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys
Asn Leu 100 105 110Val Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu
Ser Trp Thr Ala 115 120 125Pro Asp Ala Ala Phe Asp Ser Phe Trp Ile
Arg Tyr Phe Glu Phe Val 130 135 140Gly Ser Gly Glu Ala Ile Val Leu
Thr Val Pro Gly Ser Glu Arg Ser145 150 155 160Tyr Asp Leu Thr Gly
Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile 165 170 175Leu Gly Val
Lys Gly Gly Ser Ile Ser Pro Pro Leu Ser Ala Ile Phe 180 185 190Thr
Thr Cys 195176195PRTArtificial SequenceBispecific EGFR/c-Met
binding molecule 176Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu
Val Thr Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Asp Asp Pro His Ala
Phe Tyr Glu Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val
Gly Glu Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr
Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile
Tyr Gly Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn Ile Arg Gly
Leu Pro Leu Ser Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala Pro Ala Pro
Ala Pro Ala Pro Leu Pro Ala Pro Lys Asn Leu 100 105 110Val Val Ser
Arg Val Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala 115 120 125Pro
Asp Ala Ala Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Val 130 135
140Ser Lys Gly Asp Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg
Ser145 150 155 160Tyr Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr
Val Val Asn Ile 165 170 175Leu Gly Val Lys Gly Gly Ser Ile Ser Pro
Pro Leu Ser Ala Ile Phe 180 185 190Thr Thr Cys
195177195PRTArtificial SequenceBispecific EGFR/c-Met binding
molecule 177Met Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu Val Thr
Glu Asp1 5 10 15Ser Ala Arg Leu Ser Trp Asp Asp Pro His Ala Phe Tyr
Glu Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu
Ala Ile Val Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu
Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly
Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn Ile Arg Gly Leu Pro
Leu Ser Ala Ile Phe Thr Thr Ala 85 90 95Pro Ala Pro Ala Pro Ala Pro
Ala Pro Leu Pro Ala Pro Lys Asn Leu 100 105 110Val Val Ser Arg Val
Thr Glu Asp Ser Ala Arg Leu Ser Trp Thr Ala 115 120 125Pro Asp Ala
Ala Phe Asp Ser Phe Trp Ile Arg Tyr Phe Glu Phe Leu 130 135 140Gly
Ser Gly Glu Ala Ile Val Leu Thr Val Pro Gly Ser Glu Arg Ser145 150
155 160Tyr Asp Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn
Ile 165 170 175Leu Ser Val Lys Gly Gly Ser Ile Ser Pro Pro Leu Ser
Ala Ile Phe 180 185 190Thr Thr Cys 195178195PRTArtificial
SequenceBispecific EGFR/c-Met binding molecule 178Met Leu Pro Ala
Pro Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp1 5 10 15Ser Ala Arg
Leu Ser Trp Asp Asp Pro Trp Ala Phe Tyr Glu Ser Phe 20 25 30Leu Ile
Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Val Leu 35 40 45Thr
Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55
60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val His Asn Val Tyr Lys65
70 75 80Asp Thr Asn Ile Arg Gly Leu Pro Leu Ser Ala Ile Phe Thr Thr
Ala 85 90 95Pro Ala Pro Ala Pro Ala Pro Ala Pro Leu Pro Ala Pro Lys
Asn Leu 100 105 110Val Val Ser Arg Val Thr Glu Asp Ser Ala Arg Leu
Ser Trp Thr Ala 115 120 125Pro Asp Ala Ala Phe Asp Ser Phe Trp Ile
Arg Tyr Phe Glu Phe Leu 130 135 140Gly Ser Gly Glu Ala Ile Val Leu
Thr Val Pro Gly Ser Glu Arg Ser145 150 155 160Tyr Asp
Leu Thr Gly Leu Lys Pro Gly Thr Glu Tyr Val Val Asn Ile 165 170
175Leu Ser Val Lys Gly Gly Ser Ile Ser Pro Pro Leu Ser Ala Ile Phe
180 185 190Thr Thr Cys 19517912PRTArtificial SequenceConsensus FG
loop of EGFR bindiing FN3 domainVARIANT(9)..(9)Xaa may be Met or
Ile 179His Asn Val Tyr Lys Asp Thr Asn Xaa Arg Gly Leu1 5
1018012PRTArtificial SequenceGF loop of EGFR binding FN3 domain
180Leu Gly Ser Tyr Val Phe Glu His Asp Val Met Leu1 5
101818PRTArtificial SequenceConsensus BC loop of EGFR binding FN3
domainVARIANT(1)..(1)Xaa may be Ala, Thr, Gly or
AspVARIANT(2)..(2)Xaa may be Ala, Asp, Tyr or TrpVARIANT(3)..(3)Xaa
may be Pro, Asp or AsnVARIANT(4)..(4)Xaa may be Leu or
absentVARIANT(5)..(5)Xaa may be Asp, His, Arg, Gly, Tyr or
TrpVARIANT(6)..(6)Xaa may be Glu, Asp or AlaVARIANT(7)..(7)Xaa may
be Ala, Phe, Gly, His or AspVARIANT(8)..(8)Xaa may be Tyr, Phe or
Leu 181Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa1 518295PRTArtificial
SequenceEGFR binding FN3 domain consensus
sequenceVARIANT(22)..(22)Xaa may be Ala, Thr, Gly or
AspVARIANT(23)..(23)Xaa may be Ala, Asp, Tyr or
TrpVARIANT(24)..(24)Xaa may be Pro, Asp or AsnVARIANT(25)..(25)Xaa
may be Leu or absentVARIANT(26)..(26)Xaa may be Asp, His, Arg, Gly,
Tyr or TrpVARIANT(27)..(27)Xaa may be Gly, Asp or
AlaVARIANT(28)..(28)Xaa may be Ala, Phe, Gly, His or
AspVARIANT(29)..(29)Xaa may be Tyr, Phe or LeuVARIANT(84)..(84)Xaa
may be Met or Ile 182Leu Pro Ala Pro Lys Asn Leu Val Val Ser Glu
Val Thr Glu Asp Ser1 5 10 15Leu Arg Leu Ser Trp Xaa Xaa Xaa Xaa Xaa
Xaa Xaa Xaa Asp Ser Phe 20 25 30Leu Ile Gln Tyr Gln Glu Ser Glu Lys
Val Gly Glu Ala Ile Asn Leu 35 40 45Thr Val Pro Gly Ser Glu Arg Ser
Tyr Asp Leu Thr Gly Leu Lys Pro 50 55 60Gly Thr Glu Tyr Thr Val Ser
Ile Tyr Gly Val His Asn Val Tyr Lys65 70 75 80Asp Thr Asn Xaa Arg
Gly Leu Pro Leu Ser Ala Glu Phe Thr Thr 85 90 9518395PRTArtificial
SequenceEGFR binding FN3 domain consensus
sequenceVARIANT(22)..(22)Xaa may be Ala, Thr, Gly or
AspVARIANT(23)..(23)Xaa may be Ala, Asp, Tyr or
TrpVARIANT(24)..(24)Xaa may be Pro, Asp or AsnVARIANT(25)..(25)Xaa
may be Leu or absentVARIANT(26)..(26)Xaa may be Asp, His, Arg, Gly,
Tyr or TrpVARIANT(27)..(27)Xaa may be Gly, Asp or
AlaVARIANT(28)..(28)Xaa may be Ala, Phe, Gly, His or
AspVARIANT(29)..(29)Xaa may be Tyr, Phe or Leu 183Leu Pro Ala Pro
Lys Asn Leu Val Val Ser Glu Val Thr Glu Asp Ser1 5 10 15Leu Arg Leu
Ser Trp Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Asp Ser Phe 20 25 30Leu Ile
Gln Tyr Gln Glu Ser Glu Lys Val Gly Glu Ala Ile Asn Leu 35 40 45Thr
Val Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro 50 55
60Gly Thr Glu Tyr Thr Val Ser Ile Tyr Gly Val Leu Gly Ser Tyr Val65
70 75 80Phe Glu His Asp Val Met Leu Pro Leu Ser Ala Glu Phe Thr Thr
85 90 9518415PRTArtificial Sequencec-Met binding FN3 domain C
strand and CD loop consensus sequenceVARIANT(4)..(4)Xaa is Trp, Phe
or ValVARIANT(8)..(8)Xaa is Asp, Phe or LeuVARIANT(10)..(10)Xaa is
Val, Phe or LeuVARIANT(11)..(11)Xaa is Val, Leu or
ThrVARIANT(12)..(12)Xaa is Val, Arg, Gly, Leu, Thr or
SerVARIANT(13)..(13)Xaa is Gly, Ser, Ala, Thr or
LysVARIANT(15)..(15)Xaa is Glu or Asp 184Asp Ser Phe Xaa Ile Arg
Tyr Xaa Glu Xaa Xaa Xaa Xaa Gly Xaa1 5 10 1518517PRTArtificial
Sequencec-Met binding FN3 domain F strand and FG loop consensus
sequenceVARIANT(4)..(4)Xaa is Tyr, Trp, Ile, Val, Gly or
AlaVARIANT(6)..(6)Xaa is Asn, Thr, Qln or GlyVARIANT(8)..(8)Xaa is
Leu, Met, Asn or IleVARIANT(9)..(9)Xaa is Gly or
SerVARIANT(14)..(14)Xaa is Ser, Leu, Gly, Tyr, Thr, Arg, His or
LysVARIANT(15)..(15)Xaa is Ile, Val or LeuVARIANT(17)..(17)Xaa is
Val, Thr, His, Ile, Pro, Tyr or Leu 185Thr Glu Tyr Xaa Val Xaa Ile
Xaa Xaa Val Lys Gly Gly Xaa Xaa Ser1 5 10 15Xaa18689PRTArtificial
Sequencec-Met binding FN3 domain consensus
sequenceVARIANT(32)..(32)Xaa is Trp, Phe or ValVARIANT(36)..(36)Xaa
is Asp, Phe or LeuVARIANT(38)..(38)Xaa is Val, Phe or
LeuVARIANT(39)..(39)Xaa is Val, Leu or ThrVARIANT(40)..(40)Xaa is
Val, Arg, Gly, Leu, Thr or SerVARIANT(41)..(41)Xaa is Gly, Ser,
Ala, Thr or LysVARIANT(43)..(43)Xaa is Glu or
AspVARIANT(68)..(68)Xaa is Tyr, Trp, Ile, Val, Gly or
AlaVARIANT(70)..(70)Xaa is Asn, Thr, Qln or GlyVARIANT(72)..(72)Xaa
is Leu, Met, Asn or IleVARIANT(73)..(73)Xaa is Gly or
SerVARIANT(78)..(78)Xaa is Ser, Leu, Gly, Tyr, Thr, Arg, His or
LysVARIANT(79)..(79)Xaa is Ile, Val or LeuVARIANT(81)..(81)Xaa is
Val, Thr, His, Ile, Pro, Tyr or Leu 186Leu Pro Ala Pro Lys Asn Leu
Val Val Ser Arg Val Thr Glu Asp Ser1 5 10 15Ala Arg Leu Ser Trp Thr
Ala Pro Asp Ala Ala Phe Asp Ser Phe Xaa 20 25 30Ile Arg Tyr Xaa Glu
Xaa Xaa Xaa Xaa Gly Xaa Ala Ile Val Leu Thr 35 40 45Val Pro Gly Ser
Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr
Xaa Val Xaa Ile Xaa Xaa Val Lys Gly Gly Xaa Xaa Ser65 70 75 80Xaa
Pro Leu Ser Ala Glu Phe Thr Thr 8518794PRTArtificial SequenceEGFR
consensus FN3 domain of bispecific EGFR/c-Met
moleculeVARIANT(11)..(11)Xaa is Glu, Asn or ArgVARIANT(14)..(14)Xaa
is Glu or ProVARIANT(17)..(17)Xaa is Leu or AlaVARIANT(25)..(25)Xaa
is His or TrpVARIANT(29)..(29)Xaa is Glu or AspVARIANT(37)..(37)Xaa
is Glu or ProVARIANT(46)..(46)Xaa is Asn or ValVARIANT(73)..(73)Xaa
is Gly or TyrVARIANT(83)..(83)Xaa is Met or IleVARIANT(91)..(91)Xaa
is Glu or Ile 187Leu Pro Ala Pro Lys Asn Leu Val Val Ser Xaa Val
Thr Xaa Asp Ser1 5 10 15Xaa Arg Leu Ser Trp Asp Asp Pro Xaa Ala Phe
Tyr Xaa Ser Phe Leu 20 25 30Ile Gln Tyr Gln Xaa Ser Glu Lys Val Gly
Glu Ala Ile Xaa Leu Thr 35 40 45Val Pro Gly Ser Glu Arg Ser Tyr Asp
Leu Thr Gly Leu Lys Pro Gly 50 55 60Thr Glu Tyr Thr Val Ser Ile Tyr
Xaa Val His Asn Val Tyr Lys Asp65 70 75 80Thr Asn Xaa Arg Gly Leu
Pro Leu Ser Ala Xaa Phe Thr Thr 85 9018889PRTArtificial
Sequencec-Met consensus FN3 domain of bispecific EGFR/c-Met
moleculeVARIANT(11)..(11)Xaa is Glu, Asn or ArgVARIANT(14)..(14)Xaa
is Glu or ProVARIANT(17)..(17)Xaa is Leu or AlaVARIANT(37)..(37)Xaa
is Glu or ProVARIANT(39)..(39)Xaa is Val or LeuVARIANT(40)..(40)Xaa
is Gly or SerVARIANT(41)..(41)Xaa is Ser or LysVARIANT(43)..(43)Xaa
is Glu or AspVARIANT(46)..(46)Xaa is Asn or ValVARIANT(72)..(72)Xaa
is Leu or MetVARIANT(73)..(73)Xaa is Gly or SerVARIANT(78)..(78)Xaa
is Ser or LysVARIANT(86)..(86)Xaa is Glu or Ile 188Leu Pro Ala Pro
Lys Asn Leu Val Val Ser Xaa Val Thr Xaa Asp Ser1 5 10 15Xaa Arg Leu
Ser Trp Thr Ala Pro Asp Ala Ala Phe Asp Ser Phe Trp 20 25 30Ile Arg
Tyr Phe Xaa Phe Xaa Xaa Xaa Gly Xaa Ala Ile Xaa Leu Thr 35 40 45Val
Pro Gly Ser Glu Arg Ser Tyr Asp Leu Thr Gly Leu Lys Pro Gly 50 55
60Thr Glu Tyr Val Val Asn Ile Xaa Xaa Val Lys Gly Gly Xaa Ile Ser65
70 75 80Pro Pro Leu Ser Ala Xaa Phe Thr Thr 85189125PRTHomo Sapiens
189Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg1
5 10 15Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr
Tyr 20 25 30Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu
Trp Val 35 40 45Ala Val Ile Trp Asp Asp Gly Ser Tyr Lys Tyr Tyr Gly
Asp Ser Val 50 55 60Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys
Asn Thr Leu Tyr65 70 75 80Leu Gln Met Asn Ser Leu Arg Ala Glu Asp
Thr Ala Val Tyr Tyr Cys 85 90 95Ala Arg Asp Gly Ile Thr Met Val Arg
Gly Val Met Lys Asp Tyr Phe 100 105 110Asp Tyr Trp Gly Gln Gly Thr
Leu Val Thr Val Ser Ser 115 120 125190107PRTHomo Sapiens 190Ala Ile
Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly1 5 10 15Asp
Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Ser Ala 20 25
30Leu Val Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45Tyr Asp Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser
Gly 50 55 60Ser Glu Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
Gln Pro65 70 75 80Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Phe Asn
Ser Tyr Pro Leu 85 90 95Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105191121PRTHomo Sapiens 191Glu Val Gln Leu Val Glu Ser Gly Gly
Gly Leu Val Gln Pro Gly Gly1 5 10 15Ser Leu Arg Leu Ser Cys Ala Ala
Ser Gly Phe Thr Phe Ser Ser Tyr 20 25 30Trp Met Asn Trp Val Arg Gln
Ala Pro Gly Lys Gly Leu Glu Trp Val 35 40 45Ala Asn Ile Lys Lys Asp
Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val 50 55 60Lys Gly Arg Phe Thr
Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr65 70 75 80Leu Gln Met
Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys 85 90 95Ala Arg
Asp Leu Gly Trp Gly Trp Gly Trp Tyr Phe Asp Leu Trp Gly 100 105
110Arg Gly Thr Leu Val Thr Val Ser Ser 115 120192107PRTHomo Sapiens
192Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly1
5 10 15Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser
Tyr 20 25 30Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu
Leu Ile 35 40 45Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg
Phe Ser Gly 50 55 60Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
Ser Leu Glu Pro65 70 75 80Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln
Arg Ser Asn Trp Pro Pro 85 90 95Thr Phe Gly Gln Gly Thr Lys Val Glu
Ile Lys 100 105193119PRTHomo Sapiens 193Gln Val Gln Leu Val Gln Ser
Gly Ala Glu Val Lys Lys Pro Gly Ala1 5 10 15Ser Val Lys Val Ser Cys
Glu Thr Ser Gly Tyr Thr Phe Thr Ser Tyr 20 25 30Gly Ile Ser Trp Val
Arg Gln Ala Pro Gly His Gly Leu Glu Trp Met 35 40 45Gly Trp Ile Ser
Ala Tyr Asn Gly Tyr Thr Asn Tyr Ala Gln Lys Leu 50 55 60Gln Gly Arg
Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr65 70 75 80Met
Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys 85 90
95Ala Arg Asp Leu Arg Gly Thr Asn Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110Thr Leu Val Thr Val Ser Ser 115194107PRTHomo Sapiens
194Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly1
5 10 15Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn
Trp 20 25 30Leu Ala Trp Phe Gln His Lys Pro Gly Lys Ala Pro Lys Leu
Leu Ile 35 40 45Tyr Ala Ala Ser Ser Leu Leu Ser Gly Val Pro Ser Arg
Phe Ser Gly 50 55 60Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
Ser Leu Gln Pro65 70 75 80Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln
Ala Asn Ser Phe Pro Ile 85 90 95Thr Phe Gly Gln Gly Thr Arg Leu Glu
Ile Lys 100 105195125PRTHomo Sapiens 195Glu Val Gln Leu Val Glu Ser
Gly Gly Gly Leu Val Lys Pro Gly Gly1 5 10 15Ser Leu Lys Leu Ser Cys
Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr 20 25 30Tyr Met Tyr Trp Val
Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val 35 40 45Ala Thr Ile Ser
Asp Asp Gly Ser Tyr Thr Tyr Tyr Pro Asp Ser Val 50 55 60Lys Gly Arg
Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Asn Leu Tyr65 70 75 80Leu
Gln Met Ser Ser Leu Lys Ser Glu Asp Thr Ala Met Tyr Tyr Cys 85 90
95Ala Arg Glu Gly Leu Tyr Tyr Tyr Gly Ser Gly Ser Tyr Tyr Asn Gln
100 105 110Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser 115
120 125196106PRTHomo Sapiens 196Gln Leu Thr Gln Ser Pro Ser Ser Leu
Ser Ala Ser Val Gly Asp Arg1 5 10 15Val Thr Ile Thr Cys Arg Ala Ser
Gln Gly Leu Ser Ser Ala Leu Ala 20 25 30Trp Tyr Arg Gln Lys Pro Gly
Lys Ala Pro Lys Leu Leu Ile Tyr Asp 35 40 45Ala Ser Ser Leu Glu Ser
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly 50 55 60Ser Gly Thr Asp Phe
Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp65 70 75 80Phe Ala Thr
Tyr Tyr Cys Gln Gln Phe Thr Ser Tyr Pro Gln Ile Thr 85 90 95Phe Gly
Gln Gly Thr Arg Leu Glu Ile Lys 100 105197457PRTHomo Sapiens 197Gln
Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala1 5 10
15Ser Val Lys Val Ser Cys Gln Ala Ser Gly Tyr Arg Phe Ser Asn Phe
20 25 30Val Ile His Trp Val Arg Gln Ala Pro Gly Gln Arg Phe Glu Trp
Met 35 40 45Gly Trp Ile Asn Pro Tyr Asn Gly Asn Lys Glu Phe Ser Ala
Lys Phe 50 55 60Gln Asp Arg Val Thr Phe Thr Ala Asp Thr Ser Ala Asn
Thr Ala Tyr65 70 75 80Met Glu Leu Arg Ser Leu Arg Ser Ala Asp Thr
Ala Val Tyr Tyr Cys 85 90 95Ala Arg Val Gly Pro Tyr Ser Trp Asp Asp
Ser Pro Gln Asp Asn Tyr 100 105 110Tyr Met Asp Val Trp Gly Lys Gly
Thr Thr Val Ile Val Ser Ser Ala 115 120 125Ser Thr Lys Gly Pro Ser
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser 130 135 140Thr Ser Gly Gly
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe145 150 155 160Pro
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly 165 170
175Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu
180 185 190Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln
Thr Tyr 195 200 205Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
Val Asp Lys Arg 210 215 220Val Glu Pro Lys Ser Cys Asp Lys Thr His
Thr Cys Pro Pro Cys Pro225 230 235 240Ala Pro Glu Leu Leu Gly Gly
Pro Ser Val Phe Leu Phe Pro Pro Lys 245 250 255Pro Lys Asp Thr Leu
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val 260 265 270Val Val Asp
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr 275 280 285Val
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu 290 295
300Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
His305 310 315 320Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
Val Ser Asn Lys 325 330 335Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
Ser Lys Ala Lys Gly Gln 340 345 350Pro Arg Glu Pro Gln Val Tyr Thr
Leu Pro Pro Ser Arg Glu Glu Met 355 360 365Thr Lys Asn Gln Val Ser
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro 370 375 380Ser Asp Ile Ala
Val Glu Trp Glu Ser Asn Gly Gln
Pro Glu Asn Asn385 390 395 400Tyr Lys Thr Thr Pro Pro Val Leu Asp
Ser Asp Gly Ser Phe Leu Leu 405 410 415Tyr Ser Lys Leu Thr Val Asp
Lys Ser Arg Trp Gln Gln Gly Asn Val 420 425 430Phe Ser Cys Ser Val
Met His Glu Ala Leu His Asn His Tyr Thr Gln 435 440 445Lys Ser Leu
Ser Leu Ser Pro Gly Lys 450 455198457PRTHomo Sapiens 198Gln Val Gln
Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala1 5 10 15Ser Val
Lys Val Ser Cys Gln Ala Ser Gly Tyr Arg Phe Ser Asn Phe 20 25 30Val
Ile His Trp Val Arg Gln Ala Pro Gly Gln Arg Phe Glu Trp Met 35 40
45Gly Trp Ile Asn Pro Tyr Asn Gly Asn Lys Glu Phe Ser Ala Lys Phe
50 55 60Gln Asp Arg Val Thr Phe Thr Ala Asp Thr Ser Ala Asn Thr Ala
Tyr65 70 75 80Met Glu Leu Arg Ser Leu Arg Ser Ala Asp Thr Ala Val
Tyr Tyr Cys 85 90 95Ala Arg Val Gly Pro Tyr Ser Trp Asp Asp Ser Pro
Gln Asp Asn Tyr 100 105 110Tyr Met Asp Val Trp Gly Lys Gly Thr Thr
Val Ile Val Ser Ser Ala 115 120 125Ser Thr Lys Gly Pro Ser Val Phe
Pro Leu Ala Pro Ser Ser Lys Ser 130 135 140Thr Ser Gly Gly Thr Ala
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe145 150 155 160Pro Glu Pro
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly 165 170 175Val
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu 180 185
190Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr
195 200 205Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp
Lys Arg 210 215 220Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
Pro Pro Cys Pro225 230 235 240Ala Pro Glu Leu Leu Gly Gly Pro Ser
Val Phe Leu Phe Pro Pro Lys 245 250 255Pro Lys Asp Thr Leu Met Ile
Ser Arg Thr Pro Glu Val Thr Cys Val 260 265 270Val Val Asp Val Ser
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr 275 280 285Val Asp Gly
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu 290 295 300Gln
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His305 310
315 320Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
Lys 325 330 335Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
Lys Gly Gln 340 345 350Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
Ser Arg Glu Glu Met 355 360 365Thr Lys Asn Gln Val Ser Leu Thr Cys
Leu Val Lys Gly Phe Tyr Pro 370 375 380Ser Asp Ile Ala Val Glu Trp
Glu Ser Asn Gly Gln Pro Glu Asn Asn385 390 395 400Tyr Lys Thr Thr
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu 405 410 415Tyr Ser
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val 420 425
430Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
435 440 445Lys Ser Leu Ser Leu Ser Pro Gly Lys 450 455199455PRTHomo
Sapiens 199Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro
Gly Arg1 5 10 15Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
Ser Thr Tyr 20 25 30Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly
Leu Glu Trp Val 35 40 45Ala Val Ile Trp Asp Asp Gly Ser Tyr Lys Tyr
Tyr Gly Asp Ser Val 50 55 60Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn
Ser Lys Asn Thr Leu Tyr65 70 75 80Leu Gln Met Asn Ser Leu Arg Ala
Glu Asp Thr Ala Val Tyr Tyr Cys 85 90 95Ala Arg Asp Gly Ile Thr Met
Val Arg Gly Val Met Lys Asp Tyr Phe 100 105 110Asp Tyr Trp Gly Gln
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr 115 120 125Lys Gly Pro
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser 130 135 140Gly
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu145 150
155 160Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
His 165 170 175Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
Leu Ser Ser 180 185 190Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
Gln Thr Tyr Ile Cys 195 200 205Asn Val Asn His Lys Pro Ser Asn Thr
Lys Val Asp Lys Arg Val Glu 210 215 220Pro Lys Ser Cys Asp Lys Thr
His Thr Cys Pro Pro Cys Pro Ala Pro225 230 235 240Glu Leu Leu Gly
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys 245 250 255Asp Thr
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val 260 265
270Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
Gln Tyr 290 295 300Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
Leu His Gln Asp305 310 315 320Trp Leu Asn Gly Lys Glu Tyr Lys Cys
Lys Val Ser Asn Lys Ala Leu 325 330 335Pro Ala Pro Ile Glu Lys Thr
Ile Ser Lys Ala Lys Gly Gln Pro Arg 340 345 350Glu Pro Gln Val Tyr
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys 355 360 365Asn Gln Val
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp 370 375 380Ile
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys385 390
395 400Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Leu Leu Tyr
Ser 405 410 415Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
Val Phe Ser 420 425 430Cys Ser Val Met His Glu Ala Leu His Asn His
Tyr Thr Gln Lys Ser 435 440 445Leu Ser Leu Ser Pro Gly Lys 450
455200214PRTHomo Sapiens 200Ala Ile Gln Leu Thr Gln Ser Pro Ser Ser
Leu Ser Ala Ser Val Gly1 5 10 15Asp Arg Val Thr Ile Thr Cys Arg Ala
Ser Gln Asp Ile Ser Ser Ala 20 25 30Leu Val Trp Tyr Gln Gln Lys Pro
Gly Lys Ala Pro Lys Leu Leu Ile 35 40 45Tyr Asp Ala Ser Ser Leu Glu
Ser Gly Val Pro Ser Arg Phe Ser Gly 50 55 60Ser Glu Ser Gly Thr Asp
Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro65 70 75 80Glu Asp Phe Ala
Thr Tyr Tyr Cys Gln Gln Phe Asn Ser Tyr Pro Leu 85 90 95Thr Phe Gly
Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala 100 105 110Pro
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly 115 120
125Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
Ser Gln145 150 155 160Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
Thr Tyr Ser Leu Ser 165 170 175Ser Thr Leu Thr Leu Ser Lys Ala Asp
Tyr Glu Lys His Lys Val Tyr 180 185 190Ala Cys Glu Val Thr His Gln
Gly Leu Ser Ser Pro Val Thr Lys Ser 195 200 205Phe Asn Arg Gly Glu
Cys 210201449PRTHomo Sapiens 201Gln Val Gln Leu Val Gln Ser Gly Ala
Glu Val Lys Lys Pro Gly Ala1 5 10 15Ser Val Lys Val Ser Cys Glu Thr
Ser Gly Tyr Thr Phe Thr Ser Tyr 20 25 30Gly Ile Ser Trp Val Arg Gln
Ala Pro Gly His Gly Leu Glu Trp Met 35 40 45Gly Trp Ile Ser Ala Tyr
Asn Gly Tyr Thr Asn Tyr Ala Gln Lys Leu 50 55 60Gln Gly Arg Val Thr
Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr65 70 75 80Met Glu Leu
Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys 85 90 95Ala Arg
Asp Leu Arg Gly Thr Asn Tyr Phe Asp Tyr Trp Gly Gln Gly 100 105
110Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
Ala Leu 130 135 140Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
Thr Val Ser Trp145 150 155 160Asn Ser Gly Ala Leu Thr Ser Gly Val
His Thr Phe Pro Ala Val Leu 165 170 175Gln Ser Ser Gly Leu Tyr Ser
Leu Ser Ser Val Val Thr Val Pro Ser 180 185 190Ser Ser Leu Gly Thr
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro 195 200 205Ser Asn Thr
Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys 210 215 220Thr
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro225 230
235 240Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
Ser 245 250 255Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
His Glu Asp 260 265 270Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
Val Glu Val His Asn 275 280 285Ala Lys Thr Lys Pro Arg Glu Glu Gln
Tyr Asn Ser Thr Tyr Arg Val 290 295 300Val Ser Val Leu Thr Val Leu
His Gln Asp Trp Leu Asn Gly Lys Glu305 310 315 320Tyr Lys Cys Lys
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys 325 330 335Thr Ile
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr 340 345
350Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
Trp Glu 370 375 380Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
Pro Pro Val Leu385 390 395 400Asp Ser Asp Gly Ser Phe Phe Leu Tyr
Ser Arg Leu Thr Val Asp Lys 405 410 415Ser Arg Trp Gln Gln Gly Asn
Val Phe Ser Cys Ser Val Met His Glu 420 425 430Ala Leu His Asn His
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly 435 440
445Lys202214PRTHomo Sapiens 202Asp Ile Gln Met Thr Gln Ser Pro Ser
Ser Val Ser Ala Ser Val Gly1 5 10 15Asp Arg Val Thr Ile Thr Cys Arg
Ala Ser Gln Gly Ile Ser Asn Trp 20 25 30Leu Ala Trp Phe Gln His Lys
Pro Gly Lys Ala Pro Lys Leu Leu Ile 35 40 45Tyr Ala Ala Ser Ser Leu
Leu Ser Gly Val Pro Ser Arg Phe Ser Gly 50 55 60Ser Gly Ser Gly Thr
Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro65 70 75 80Glu Asp Phe
Ala Thr Tyr Tyr Cys Gln Gln Ala Asn Ser Phe Pro Ile 85 90 95Thr Phe
Gly Gln Gly Thr Arg Leu Glu Ile Lys Arg Thr Val Ala Ala 100 105
110Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
Glu Ala 130 135 140Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
Gly Asn Ser Gln145 150 155 160Glu Ser Val Thr Glu Gln Asp Ser Lys
Asp Ser Thr Tyr Ser Leu Ser 165 170 175Ser Thr Leu Thr Leu Ser Lys
Ala Asp Tyr Glu Lys His Lys Val Tyr 180 185 190Ala Cys Glu Val Thr
His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser 195 200 205Phe Asn Arg
Gly Glu Cys 210203330PRTHomo Sapiens 203Ala Ser Thr Lys Gly Pro Ser
Val Phe Pro Leu Ala Pro Ser Ser Lys1 5 10 15Ser Thr Ser Gly Gly Thr
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr 20 25 30Phe Pro Glu Pro Val
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser 35 40 45Gly Val His Thr
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser 50 55 60Leu Ser Ser
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr65 70 75 80Tyr
Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys 85 90
95Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
Pro Pro 115 120 125Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
Glu Val Thr Cys 130 135 140Val Val Val Asp Val Ser His Glu Asp Pro
Glu Val Lys Phe Asn Trp145 150 155 160Tyr Val Asp Gly Val Glu Val
His Asn Ala Lys Thr Lys Pro Arg Glu 165 170 175Glu Gln Tyr Asn Ser
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu 180 185 190His Gln Asp
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn 195 200 205Lys
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly 210 215
220Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu
Glu225 230 235 240Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
Lys Gly Phe Tyr 245 250 255Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
Asn Gly Gln Pro Glu Asn 260 265 270Asn Tyr Lys Thr Thr Pro Pro Val
Leu Asp Ser Asp Gly Ser Phe Phe 275 280 285Leu Tyr Ser Arg Leu Thr
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn 290 295 300Val Phe Ser Cys
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr305 310 315 320Gln
Lys Ser Leu Ser Leu Ser Pro Gly Lys 325 330204330PRTHomo Sapiens
204Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys1
5 10 15Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
Tyr 20 25 30Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
Thr Ser 35 40 45Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
Leu Tyr Ser 50 55 60Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
Gly Thr Gln Thr65 70 75 80Tyr Ile Cys Asn Val Asn His Lys Pro Ser
Asn Thr Lys Val Asp Lys 85 90 95Arg Val Glu Pro Lys Ser Cys Asp Lys
Thr His Thr Cys Pro Pro Cys 100 105 110Pro Ala Pro Glu Leu Leu Gly
Gly Pro Ser Val Phe Leu Phe Pro Pro 115 120 125Lys Pro Lys Asp Thr
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys 130 135 140Val Val Val
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp145 150 155
160Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
Val Leu 180 185 190His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
Lys Val Ser Asn 195 200 205Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
Ile Ser Lys Ala Lys Gly 210 215 220Gln Pro Arg Glu Pro Gln Val Tyr
Thr Leu Pro Pro Ser Arg Glu Glu225 230 235 240Met Thr Lys Asn Gln
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr 245 250 255Pro Ser Asp
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn 260 265
270Asn
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Leu 275 280
285Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
Tyr Thr305 310 315 320Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys 325
3302051368DNAHomo Sapiens 205caggtgcagc tggtcgagag cggcggaggg
gtggtgcagc ccggcagaag cctgaggctg 60tcctgcgccg ccagcggctt caccttcagc
acctacggca tgcactgggt gcggcaggcc 120ccaggcaagg gcctggagtg
ggtggccgtg atctgggacg acggcagcta caagtactac 180ggcgacagcg
tgaagggcag gttcaccatc agcagggaca acagcaagaa caccctgtac
240ctgcagatga acagcctgag ggccgaggac accgccgtgt actactgcgc
cagggacggc 300atcaccatgg tgcggggcgt gatgaaggac tacttcgact
actggggcca gggcaccctg 360gtgaccgtga gcagcgccag caccaagggc
ccaagcgtgt tccccctggc ccccagcagc 420aagagcacca gcggcggcac
agccgccctg ggctgcctgg tgaaggacta cttccccgag 480ccagtgaccg
tgtcctggaa ctctggcgcc ctgacctccg gcgtgcacac cttccccgcc
540gtgctgcaga gcagcggcct gtacagcctg agcagcgtgg tgaccgtgcc
cagcagcagc 600ctgggcaccc agacctacat ctgcaacgtg aaccacaagc
ccagcaacac caaggtggac 660aagagagtgg agcccaagag ctgcgacaag
acccacacct gccccccctg cccagcccca 720gagctgctgg gcggacccag
cgtgttcctg ttccccccca agcccaagga caccctgatg 780atcagcagga
cccccgaggt gacctgcgtg gtggtggacg tgagccacga ggacccagag
840gtgaagttca actggtacgt ggacggcgtg gaggtgcaca acgccaagac
caagcccaga 900gaggagcagt acaacagcac ctacagggtg gtgtccgtgc
tgaccgtgct gcaccaggac 960tggctgaacg gcaaggaata caagtgcaag
gtctccaaca aggccctgcc agcccccatc 1020gaaaagacca tcagcaaggc
caagggccag ccacgggagc cccaggtgta caccctgccc 1080cccagccggg
aggagatgac caagaaccag gtgtccctga cctgtctggt gaagggcttc
1140taccccagcg acatcgccgt ggagtgggag agcaacggcc agcccgagaa
caactacaag 1200accacccccc cagtgctgga cagcgacggc agcttcctcc
tgtacagcaa gctgaccgtg 1260gacaagtcca ggtggcagca gggcaacgtg
ttcagctgca gcgtgatgca cgaggccctg 1320cacaaccact acacccagaa
gtccctgagc ctgagccccg gcaaatga 1368206642DNAHomo Sapiens
206atccagctga cccagagccc cagcagcctg agcgccagcg tgggcgaccg
ggtgaccatc 60acctgccggg ccagccagga catcagcagc gccctggtct ggtatcagca
gaagcccggc 120aaggccccca agctgctgat ctacgacgcc agctccctgg
aaagcggcgt gcccagccgg 180ttcagcggca gcgagagcgg caccgacttc
accctgacca tcagcagcct gcagcccgag 240gacttcgcca cctactactg
ccagcagttc aacagctacc ccctgacctt tggcggcgga 300acaaaggtgg
agatcaagcg tacggtggcc gctcccagcg tgttcatctt cccccccagc
360gacgagcagc tgaagagcgg caccgccagc gtggtgtgcc tgctgaacaa
cttctacccc 420cgggaggcca aggtgcagtg gaaggtggac aacgccctgc
agagcggcaa cagccaggag 480agcgtcaccg agcaggacag caaggactcc
acctacagcc tgagcagcac cctgaccctg 540tccaaggccg actacgagaa
gcacaaggtg tacgcctgcg aggtgaccca ccagggcctg 600tccagccccg
tgaccaagag cttcaacagg ggcgagtgct ga 6422071350DNAHomo Sapiens
207caggttcagc tggtgcagtc tggagctgag gtgaagaagc ctggggcctc
agtgaaggtc 60tcctgcgaga cttctggtta cacctttacc agctatggta tcagctgggt
gcgacaggcc 120cctggacacg ggcttgagtg gatgggatgg atcagcgctt
acaatggtta cacaaactat 180gcacagaagc tccagggcag ggtcaccatg
accacagaca catccacgag cacagcctac 240atggagctga ggagcctgag
atctgacgac acggccgtgt attactgtgc gagagatctg 300agaggaacta
actactttga ctactggggc cagggaaccc tggtcaccgt ctcctcagcc
360tccaccaagg gcccaagcgt gttccctctg gcccccagca gcaagagcac
atctggcgga 420acagccgccc tgggctgcct ggtgaaagac tacttccccg
agcccgtgac cgtgtcctgg 480aactctggcg ccctgaccag cggcgtgcac
acctttccag ccgtgctgca gagcagcggc 540ctgtacagcc tgtccagcgt
ggtgaccgtg cccagcagct ccctgggcac ccagacctac 600atctgcaacg
tgaaccacaa gcccagcaac accaaggtgg acaagcgggt ggaacccaag
660agctgcgaca agacccacac ctgtcccccc tgccctgccc ctgaactgct
gggcggaccc 720tccgtgttcc tgttcccccc aaagcccaag gacaccctga
tgatcagccg gacccccgaa 780gtgacctgcg tggtggtgga cgtgtcccac
gaggaccctg aagtgaagtt caattggtac 840gtggacggcg tggaagtgca
caacgccaag accaagccca gagaggaaca gtacaacagc 900acctaccggg
tggtgtccgt gctgacagtg ctgcaccagg actggctgaa cggcaaagag
960tacaagtgca aggtctccaa caaggccctg cctgctccca tcgagaaaac
catcagcaag 1020gccaagggcc agccccgcga gcctcaggtg tacacactgc
ctcccagccg ggaagagatg 1080accaagaacc aggtgtccct gacctgtctg
gtgaaaggct tctaccccag cgatatcgcc 1140gtggaatggg agagcaacgg
acagcccgag aacaactaca agaccacccc ccctgtgctg 1200gacagcgacg
gctccttctt cctgtactct cggctgaccg tggacaagag ccggtggcag
1260cagggaaacg tgttcagctg cagcgtgatg cacgaggccc tgcacaacca
ctacacccag 1320aagtccctga gcctgagccc cgggaagtga 1350208645DNAHomo
Sapiens 208gacatccaga tgacccagtc cccctcctcc gtgtccgcct ctgtgggcga
cagagtgacc 60atcacctgtc gggcctccca gggcatctcc aactggctgg cctggttcca
gcacaagccc 120ggcaaggccc ccaagctgct gatctacgcc gcctcctccc
tgctgtccgg cgtgccctcc 180agattctccg gctctggctc cggcaccgac
ttcaccctga ccatctccag cctgcagccc 240gaggacttcg ccacctacta
ctgccagcag gccaactcct tccccatcac cttcggccag 300ggcacccggc
tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttccccccc
360agcgacgagc agctgaagag cggcaccgcc agcgtggtgt gcctgctgaa
caacttctac 420ccccgggagg ccaaggtgca gtggaaggtg gacaacgccc
tgcagagcgg caacagccag 480gagagcgtca ccgagcagga cagcaaggac
tccacctaca gcctgagcag caccctgacc 540ctgtccaagg ccgactacga
gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 600ctgtccagcc
ccgtgaccaa gagcttcaac aggggcgagt gctga 645209215PRTHomo Sapiens
209Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly1
5 10 15Glu Arg Ala Thr Phe Ser Cys Arg Ser Ser His Ser Ile Arg Ser
Arg 20 25 30Arg Val Ala Trp Tyr Gln His Lys Pro Gly Gln Ala Pro Arg
Leu Val 35 40 45Ile His Gly Val Ser Asn Arg Ala Ser Gly Ile Ser Asp
Arg Phe Ser 50 55 60Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
Thr Arg Val Glu65 70 75 80Pro Glu Asp Phe Ala Leu Tyr Tyr Cys Gln
Val Tyr Gly Ala Ser Ser 85 90 95Tyr Thr Phe Gly Gln Gly Thr Lys Leu
Glu Arg Lys Arg Thr Val Ala 100 105 110Ala Pro Ser Val Phe Ile Phe
Pro Pro Ser Asp Glu Gln Leu Lys Ser 115 120 125Gly Thr Ala Ser Val
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu 130 135 140Ala Lys Val
Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser145 150 155
160Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
Lys Val 180 185 190Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
Pro Val Thr Lys 195 200 205Ser Phe Asn Arg Gly Glu Cys 210
2152105PRTHomo Sapiens 210Thr Tyr Gly Met His1 521117PRTHomo
Sapiens 211Val Ile Trp Asp Asp Gly Ser Tyr Lys Tyr Tyr Gly Asp Ser
Val Lys1 5 10 15Gly21216PRTHomo Sapiens 212Asp Gly Ile Thr Met Val
Arg Gly Val Met Lys Asp Tyr Phe Asp Tyr1 5 10 1521311PRTHomo
Sapiens 213Arg Ala Ser Gln Asp Ile Ser Ser Ala Leu Val1 5
102147PRTHomo Sapiens 214Asp Ala Ser Ser Leu Glu Ser1 52159PRTHomo
Sapiens 215Gln Gln Phe Asn Ser Tyr Pro Leu Thr1 52165PRTHomo
Sapiens 216Ser Tyr Gly Ile Ser1 521717PRTHomo Sapiens 217Trp Ile
Ser Ala Tyr Asn Gly Tyr Thr Asn Tyr Ala Gln Lys Leu Gln1 5 10
15Gly21810PRTHomo Sapiens 218Asp Leu Arg Gly Thr Asn Tyr Phe Asp
Tyr1 5 1021911PRTHomo Sapiens 219Arg Ala Ser Gln Gly Ile Ser Asn
Trp Leu Ala1 5 102207PRTHomo Sapiens 220Ala Ala Ser Ser Leu Leu
Ser1 52219PRTHomo Sapiens 221Gln Gln Ala Asn Ser Phe Pro Ile Thr1
52225PRTHomo Sapiens 222Ser Tyr Trp Met Asn1 522317PRTHomo Sapiens
223Asn Ile Lys Lys Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val Lys1
5 10 15Gly22412PRTHomo Sapiens 224Asp Leu Gly Trp Gly Trp Gly Trp
Tyr Phe Asp Leu1 5 1022511PRTHomo Sapiens 225Arg Ala Ser Gln Ser
Val Ser Ser Tyr Leu Ala1 5 102267PRTHomo Sapiens 226Asp Ala Ser Asn
Arg Ala Thr1 52279PRTHomo Sapiens 227Gln Gln Arg Ser Asn Trp Pro
Pro Thr1 52285PRTHomo Sapiens 228Asp Tyr Tyr Met Tyr1 522917PRTHomo
Sapiens 229Thr Ile Ser Asp Asp Gly Ser Tyr Thr Tyr Tyr Pro Asp Ser
Val Lys1 5 10 15Gly23016PRTHomo Sapiens 230Glu Gly Leu Tyr Tyr Tyr
Gly Ser Gly Ser Tyr Tyr Asn Gln Asp Tyr1 5 10 1523111PRTHomo
Sapiens 231Arg Ala Ser Gln Gly Leu Ser Ser Ala Leu Ala1 5
102327PRTHomo Sapiens 232Asp Ala Ser Ser Leu Glu Ser1 523310PRTHomo
Sapiens 233Gln Gln Phe Thr Ser Tyr Pro Gln Ile Thr1 5
10234451PRTHomo Sapiens 234Glu Val Gln Leu Val Glu Ser Gly Gly Gly
Leu Val Gln Pro Gly Gly1 5 10 15Ser Leu Arg Leu Ser Cys Ala Ala Ser
Gly Phe Thr Phe Ser Ser Tyr 20 25 30Trp Met Asn Trp Val Arg Gln Ala
Pro Gly Lys Gly Leu Glu Trp Val 35 40 45Ala Asn Ile Lys Lys Asp Gly
Ser Glu Lys Tyr Tyr Val Asp Ser Val 50 55 60Lys Gly Arg Phe Thr Ile
Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr65 70 75 80Leu Gln Met Asn
Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys 85 90 95Ala Arg Asp
Leu Gly Trp Gly Trp Gly Trp Tyr Phe Asp Leu Trp Gly 100 105 110Arg
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser 115 120
125Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
Thr Val145 150 155 160Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
His Thr Phe Pro Ala 165 170 175Val Leu Gln Ser Ser Gly Leu Tyr Ser
Leu Ser Ser Val Val Thr Val 180 185 190Pro Ser Ser Ser Leu Gly Thr
Gln Thr Tyr Ile Cys Asn Val Asn His 195 200 205Lys Pro Ser Asn Thr
Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys 210 215 220Asp Lys Thr
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly225 230 235
240Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
Ser His 260 265 270Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
Gly Val Glu Val 275 280 285His Asn Ala Lys Thr Lys Pro Arg Glu Glu
Gln Tyr Asn Ser Thr Tyr 290 295 300Arg Val Val Ser Val Leu Thr Val
Leu His Gln Asp Trp Leu Asn Gly305 310 315 320Lys Glu Tyr Lys Cys
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile 325 330 335Glu Lys Thr
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val 340 345 350Tyr
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser 355 360
365Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
Pro Pro385 390 395 400Val Leu Asp Ser Asp Gly Ser Phe Leu Leu Tyr
Ser Lys Leu Thr Val 405 410 415Asp Lys Ser Arg Trp Gln Gln Gly Asn
Val Phe Ser Cys Ser Val Met 420 425 430His Glu Ala Leu His Asn His
Tyr Thr Gln Lys Ser Leu Ser Leu Ser 435 440 445Pro Gly Lys
450235214PRTHomo Sapiens 235Glu Ile Val Leu Thr Gln Ser Pro Ala Thr
Leu Ser Leu Ser Pro Gly1 5 10 15Glu Arg Ala Thr Leu Ser Cys Arg Ala
Ser Gln Ser Val Ser Ser Tyr 20 25 30Leu Ala Trp Tyr Gln Gln Lys Pro
Gly Gln Ala Pro Arg Leu Leu Ile 35 40 45Tyr Asp Ala Ser Asn Arg Ala
Thr Gly Ile Pro Ala Arg Phe Ser Gly 50 55 60Ser Gly Ser Gly Thr Asp
Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro65 70 75 80Glu Asp Phe Ala
Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro 85 90 95Thr Phe Gly
Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala 100 105 110Pro
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly 115 120
125Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
Ser Gln145 150 155 160Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
Thr Tyr Ser Leu Ser 165 170 175Ser Thr Leu Thr Leu Ser Lys Ala Asp
Tyr Glu Lys His Lys Val Tyr 180 185 190Ala Cys Glu Val Thr His Gln
Gly Leu Ser Ser Pro Val Thr Lys Ser 195 200 205Phe Asn Arg Gly Glu
Cys 210236455PRTHomo Sapiens 236Glu Val Gln Leu Val Glu Ser Gly Gly
Gly Leu Val Lys Pro Gly Gly1 5 10 15Ser Leu Lys Leu Ser Cys Ala Ala
Ser Gly Phe Thr Phe Ser Asp Tyr 20 25 30Tyr Met Tyr Trp Val Arg Gln
Thr Pro Glu Lys Arg Leu Glu Trp Val 35 40 45Ala Thr Ile Ser Asp Asp
Gly Ser Tyr Thr Tyr Tyr Pro Asp Ser Val 50 55 60Lys Gly Arg Phe Thr
Ile Ser Arg Asp Asn Ala Lys Asn Asn Leu Tyr65 70 75 80Leu Gln Met
Ser Ser Leu Lys Ser Glu Asp Thr Ala Met Tyr Tyr Cys 85 90 95Ala Arg
Glu Gly Leu Tyr Tyr Tyr Gly Ser Gly Ser Tyr Tyr Asn Gln 100 105
110Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser
Thr Ser 130 135 140Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
Tyr Phe Pro Glu145 150 155 160Pro Val Thr Val Ser Trp Asn Ser Gly
Ala Leu Thr Ser Gly Val His 165 170 175Thr Phe Pro Ala Val Leu Gln
Ser Ser Gly Leu Tyr Ser Leu Ser Ser 180 185 190Val Val Thr Val Pro
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys 195 200 205Asn Val Asn
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu 210 215 220Pro
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro225 230
235 240Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
Lys 245 250 255Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
Val Val Val 260 265 270Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
Asn Trp Tyr Val Asp 275 280 285Gly Val Glu Val His Asn Ala Lys Thr
Lys Pro Arg Glu Glu Gln Tyr 290 295 300Asn Ser Thr Tyr Arg Val Val
Ser Val Leu Thr Val Leu His Gln Asp305 310 315 320Trp Leu Asn Gly
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu 325 330 335Pro Ala
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg 340 345
350Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
355 360 365Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
Ser Asp 370 375 380Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
Asn Asn Tyr Lys385 390 395 400Thr Thr Pro Pro Val Leu Asp Ser Asp
Gly Ser Phe Phe Leu Tyr Ser 405 410 415Arg Leu Thr Val Asp Lys Ser
Arg Trp Gln Gln Gly Asn Val Phe Ser 420 425 430Cys Ser Val Met His
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser 435 440 445Leu Ser Leu
Ser Pro Gly Lys 450 455237213PRTHomo Sapiens 237Gln Leu Thr Gln Ser
Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg1 5 10 15Val Thr Ile Thr
Cys Arg Ala Ser Gln Gly Leu Ser Ser Ala Leu Ala 20 25 30Trp Tyr Arg
Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Asp 35 40 45Ala Ser
Ser Leu Glu Ser Gly Val Pro
Ser Arg Phe Ser Gly Ser Gly 50 55 60Ser Gly Thr Asp Phe Thr Leu Thr
Ile Ser Ser Leu Gln Pro Glu Asp65 70 75 80Phe Ala Thr Tyr Tyr Cys
Gln Gln Phe Thr Ser Tyr Pro Gln Ile Thr 85 90 95Phe Gly Gln Gly Thr
Arg Leu Glu Ile Lys Arg Thr Val Ala Ala Pro 100 105 110Ser Val Phe
Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr 115 120 125Ala
Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys 130 135
140Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
Glu145 150 155 160Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
Ser Leu Ser Ser 165 170 175Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
Lys His Lys Val Tyr Ala 180 185 190Cys Glu Val Thr His Gln Gly Leu
Ser Ser Pro Val Thr Lys Ser Phe 195 200 205Asn Arg Gly Glu Cys
21023815PRTHomo Sapiens 238Pro Glu Phe Arg Asp Ser Tyr Pro Ile Lys
Tyr Val His Ala Phe1 5 10 1523916PRTHomo Sapiens 239Phe Ala Gln Ser
Lys Pro Asp Ser Ala Glu Pro Met Asp Arg Ser Ala1 5 10
1524016PRTHomo Sapiens 240Pro Gly Ala Gln Leu Ala Arg Gln Ile Gly
Ala Ser Leu Asn Asp Asp1 5 10 15
References
D00001
D00002
D00003
D00004
D00005
D00006
D00007
D00008
D00009
D00010
D00011
D00012
D00013
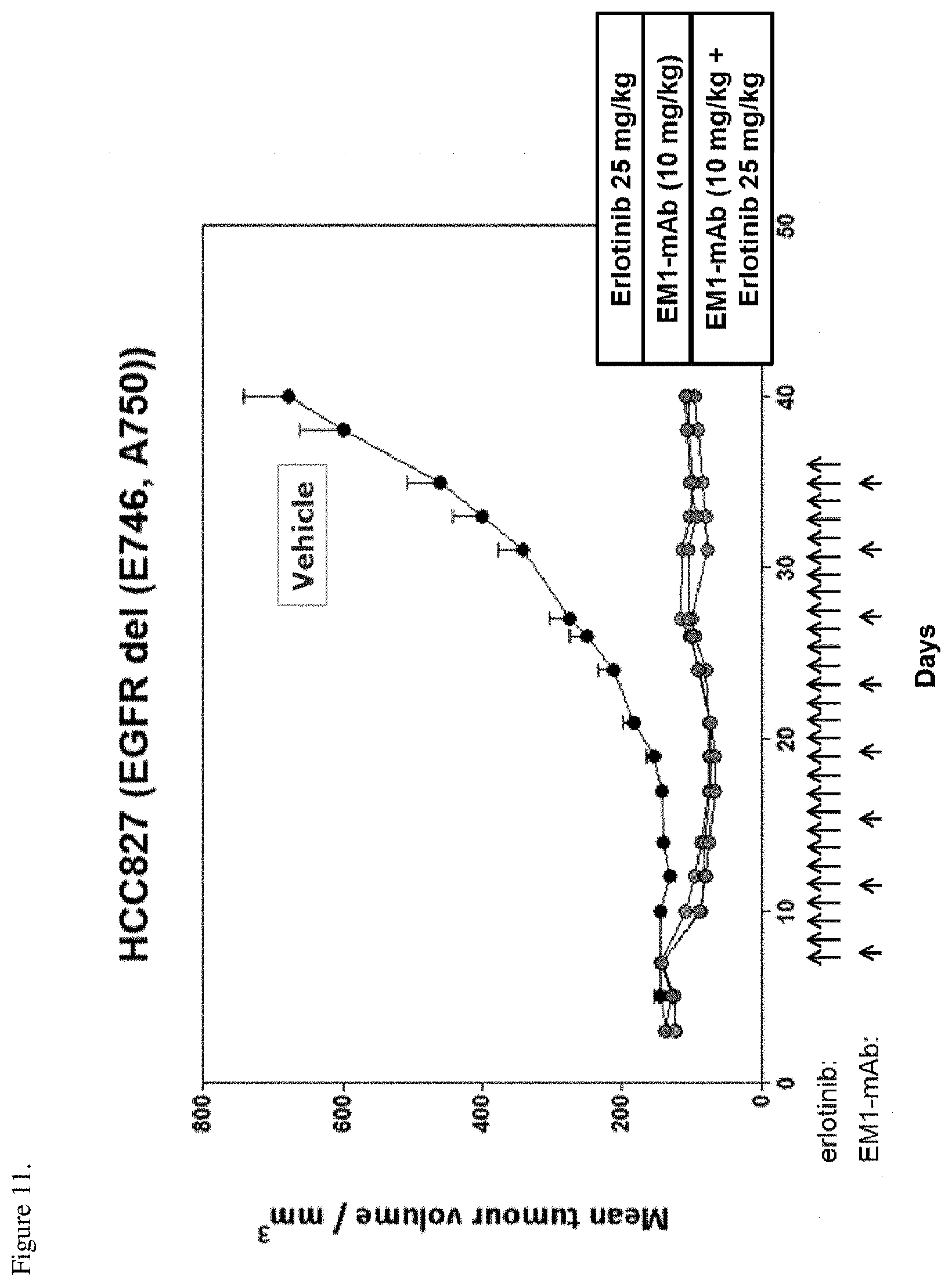
D00014
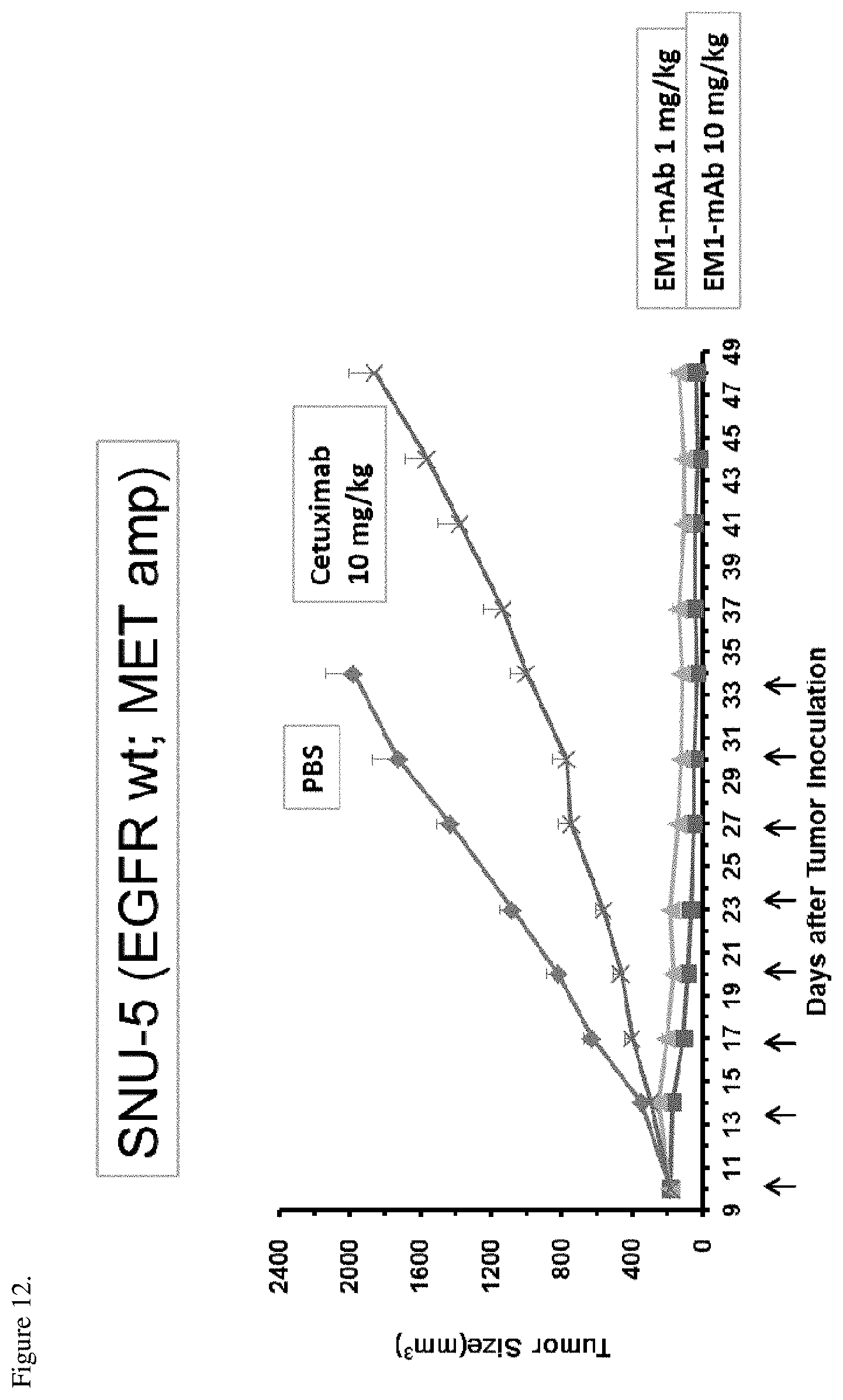
D00015
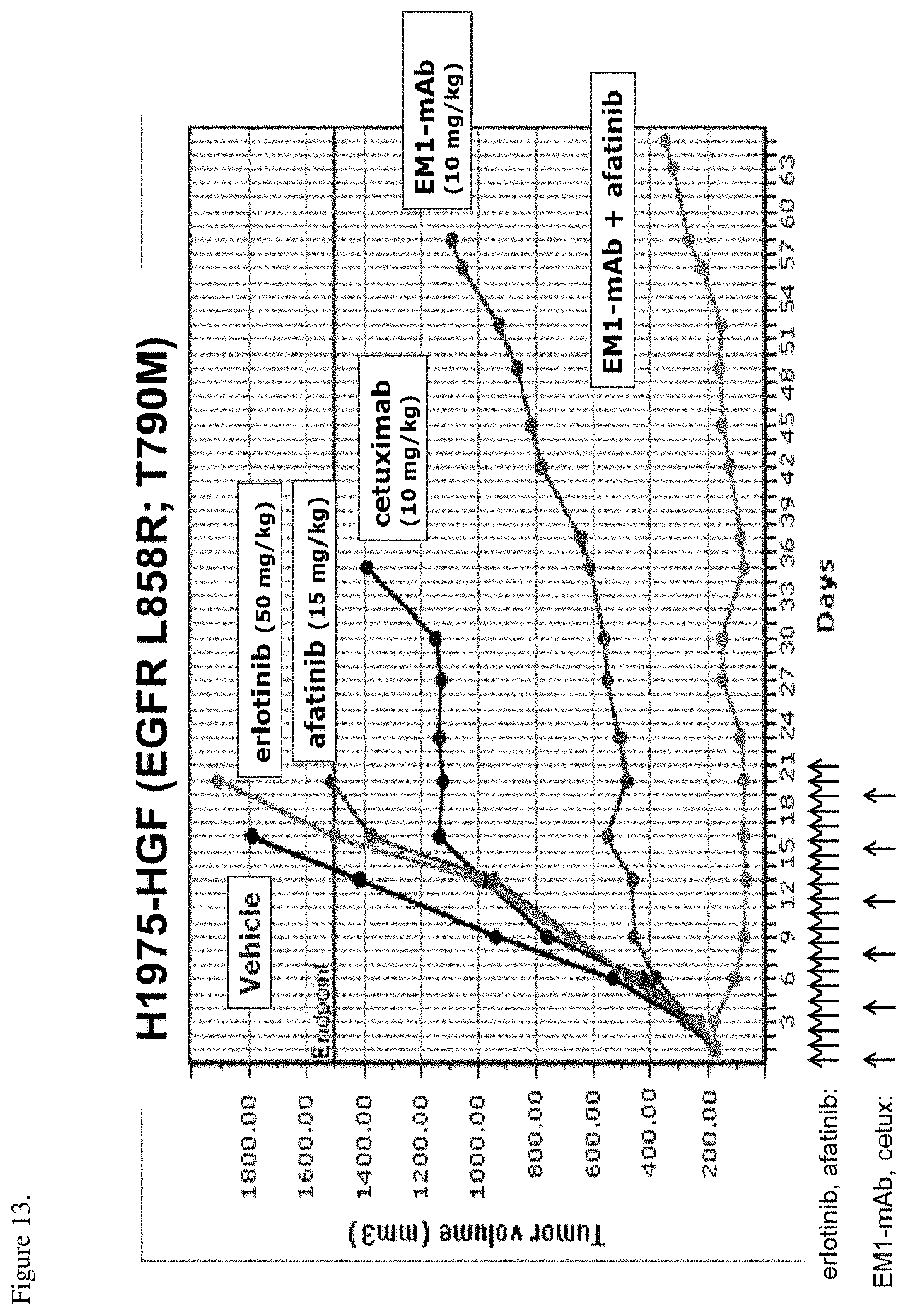
D00016
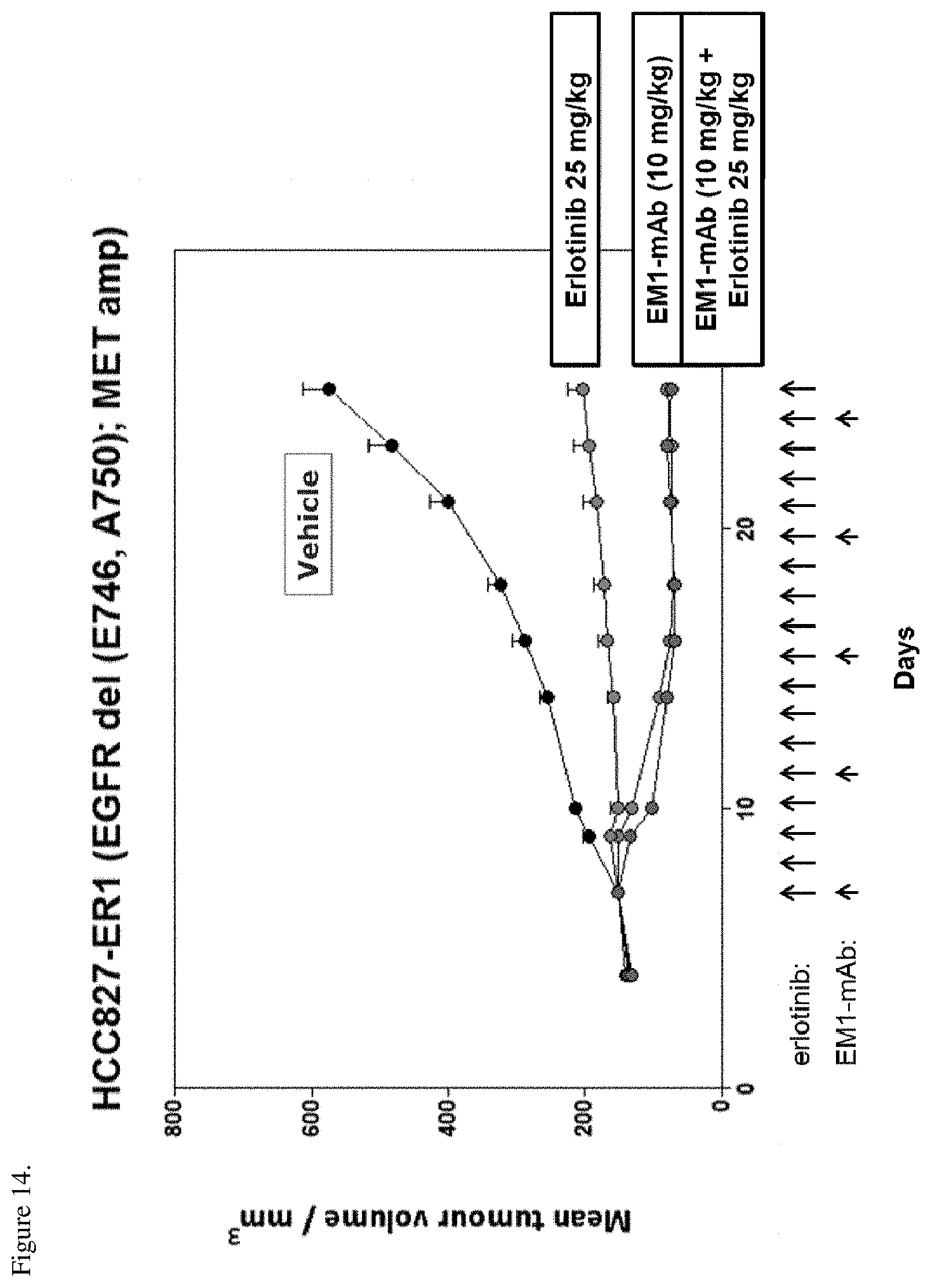
D00017
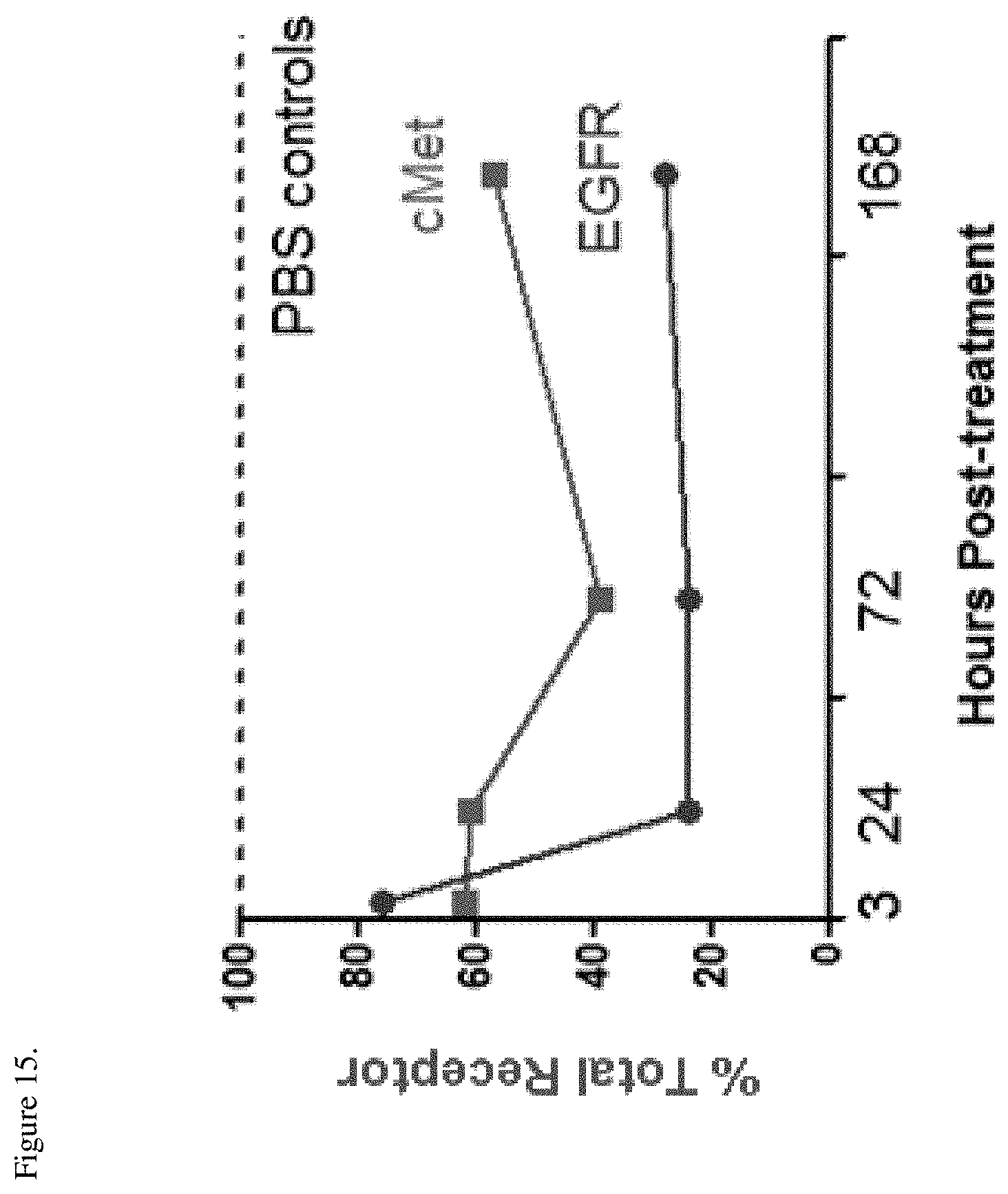
D00018

D00019
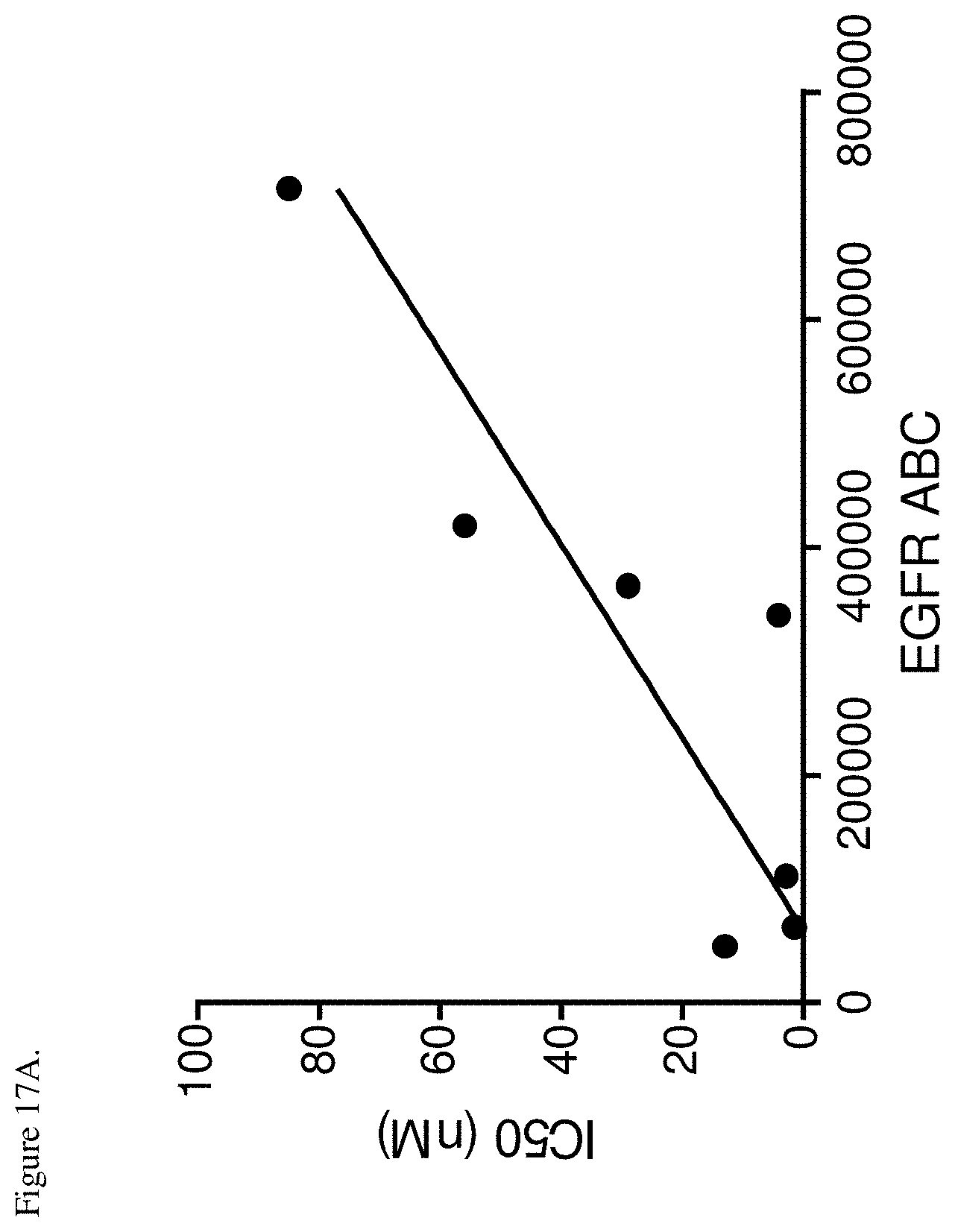
D00020
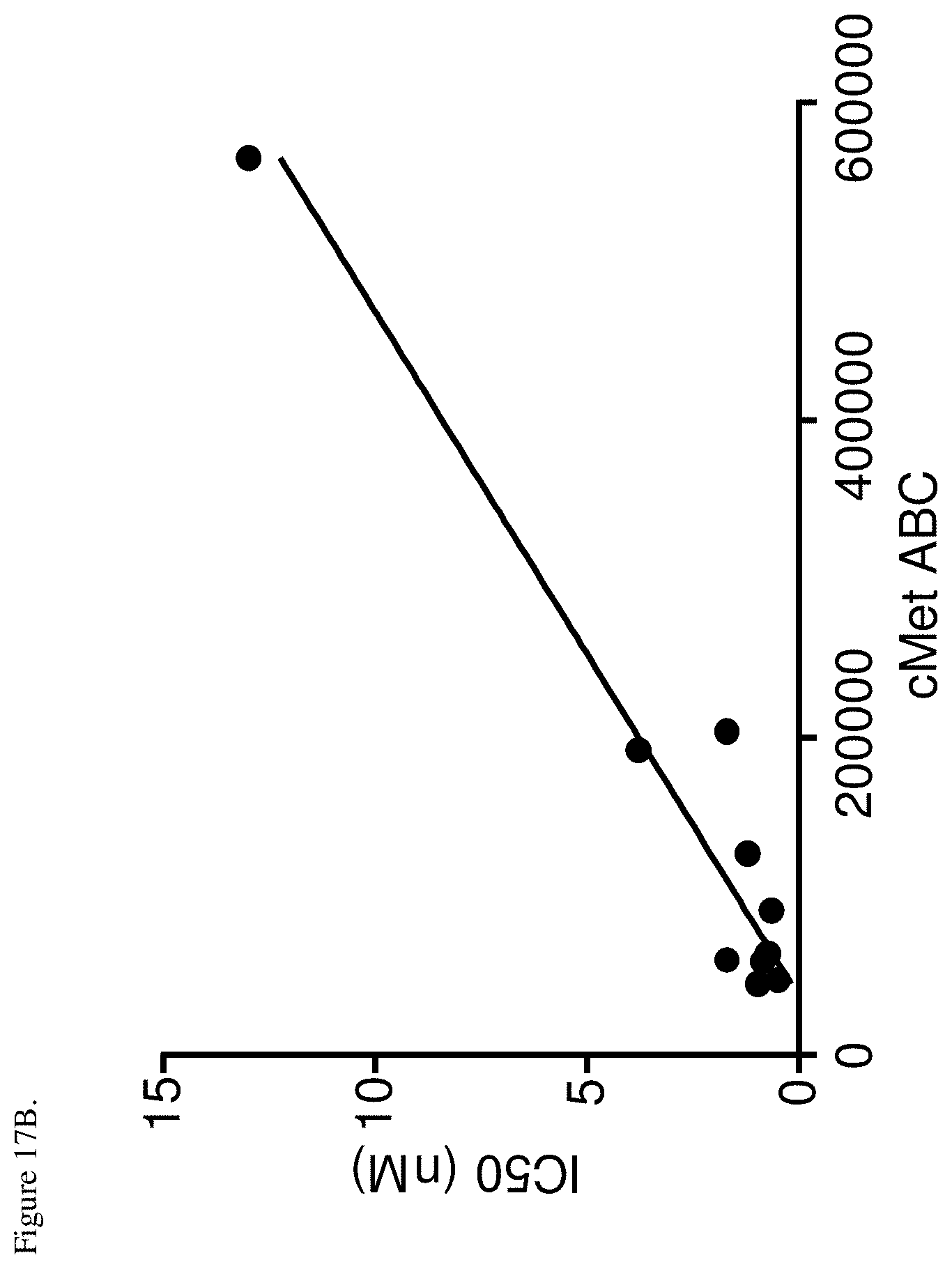
D00021

D00022
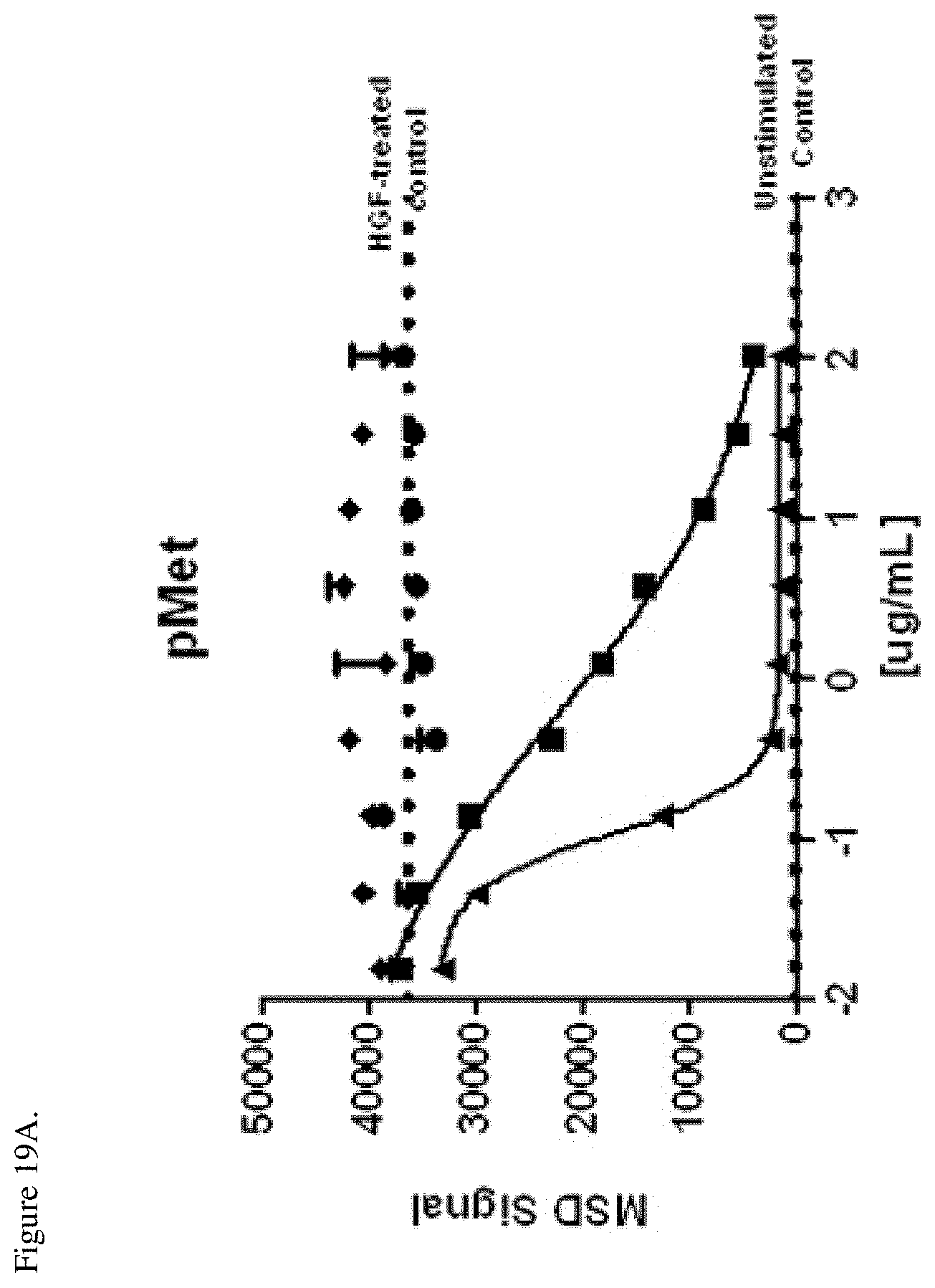
D00023
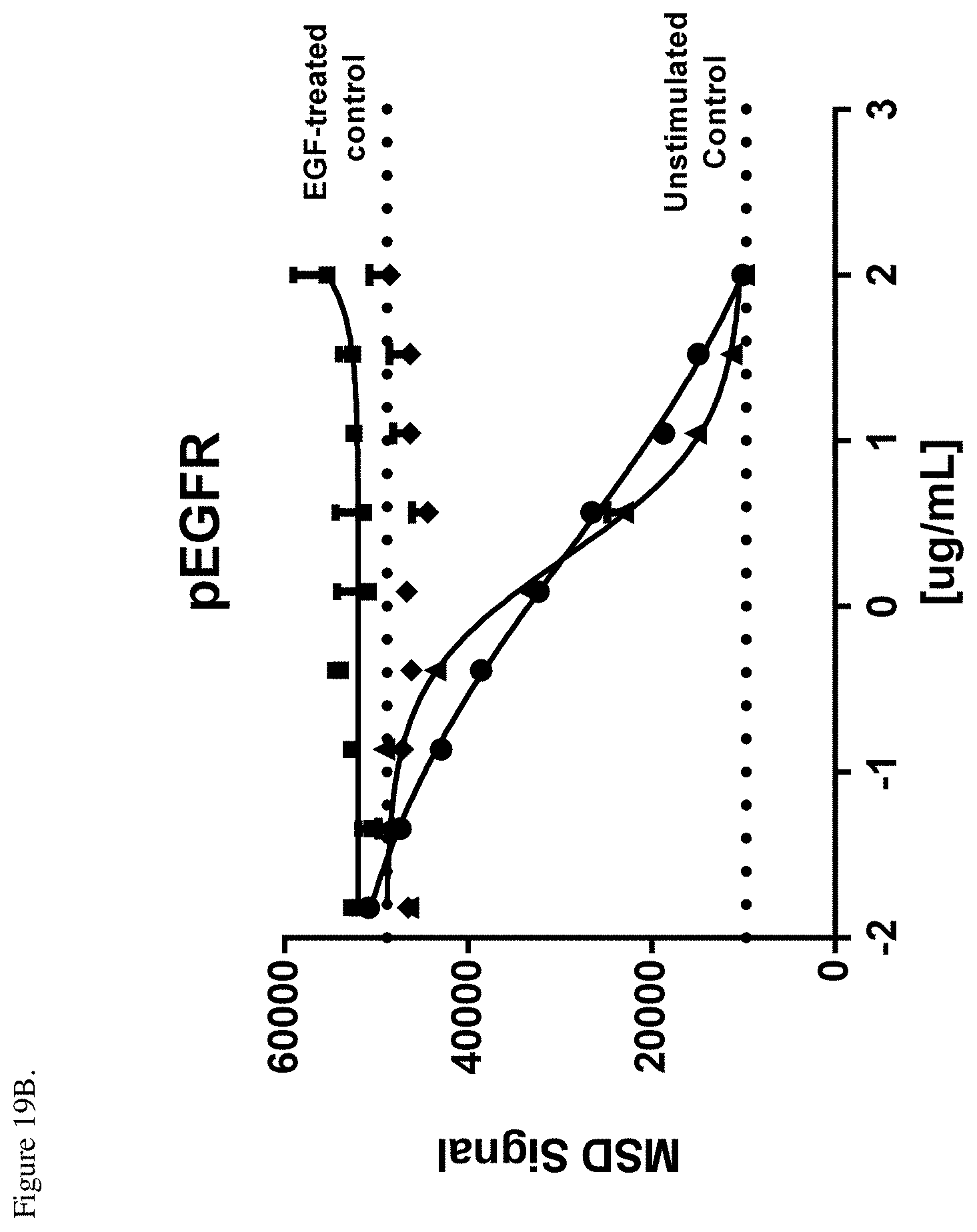
D00024
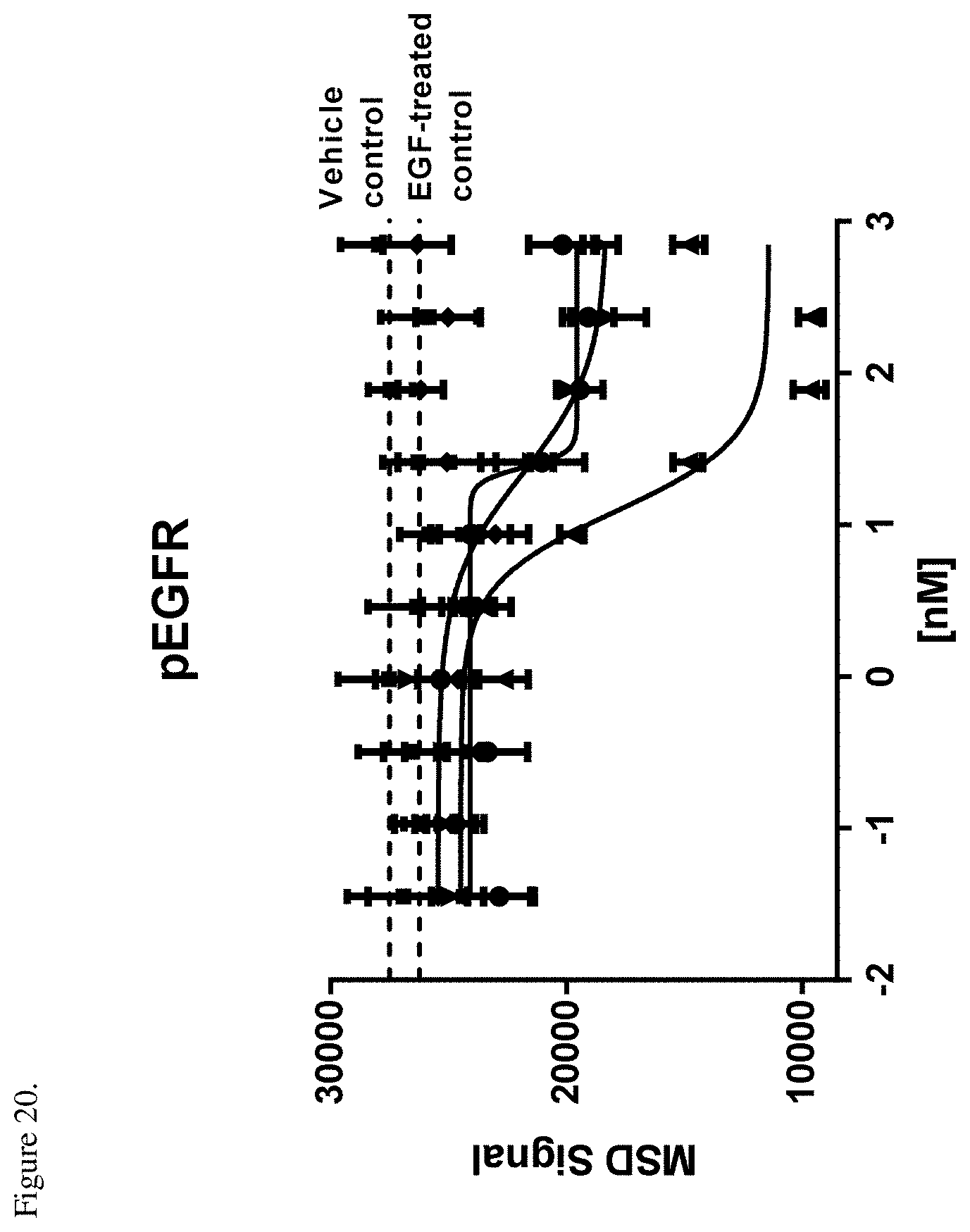
D00025
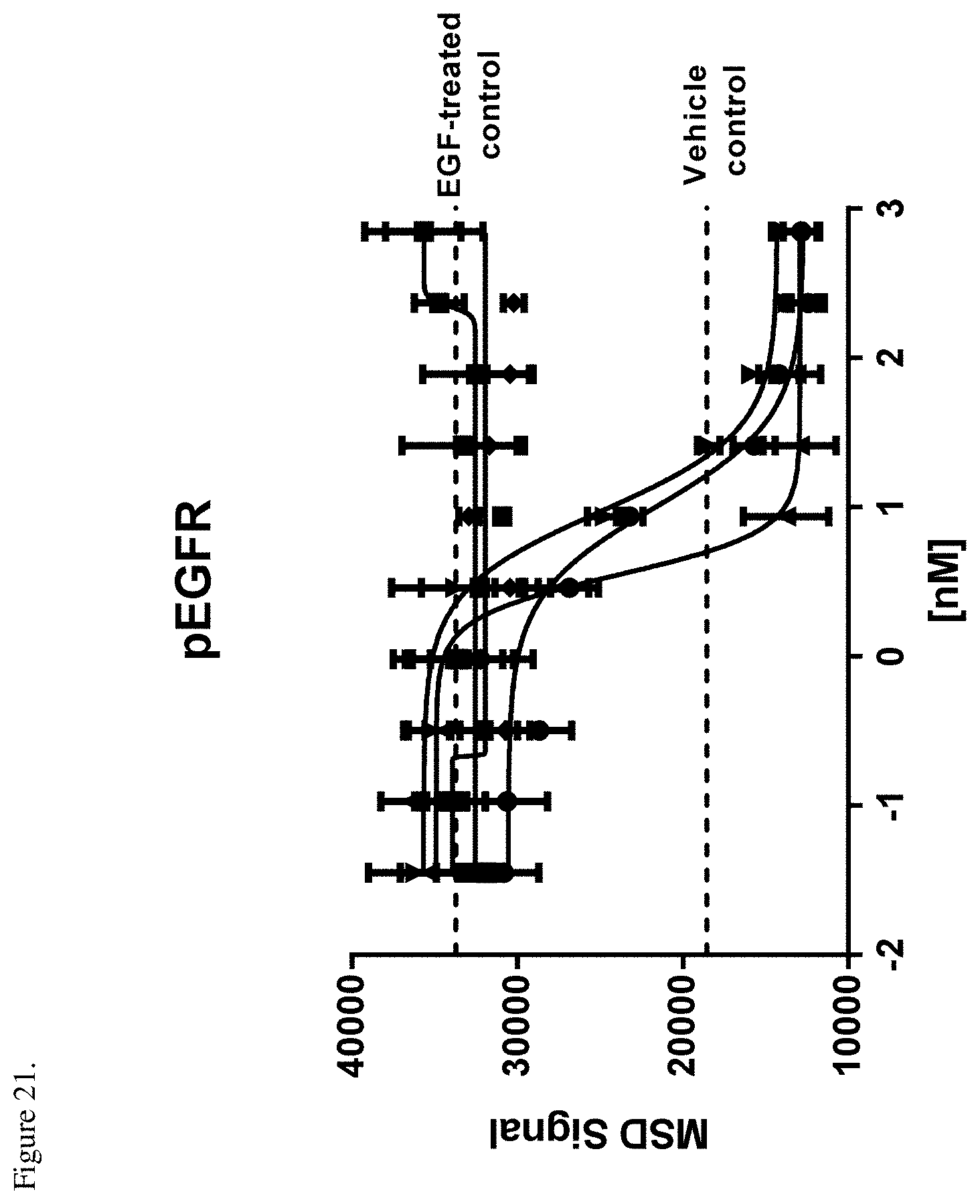
S00001
XML
uspto.report is an independent third-party trademark research tool that is not affiliated, endorsed, or sponsored by the United States Patent and Trademark Office (USPTO) or any other governmental organization. The information provided by uspto.report is based on publicly available data at the time of writing and is intended for informational purposes only.
While we strive to provide accurate and up-to-date information, we do not guarantee the accuracy, completeness, reliability, or suitability of the information displayed on this site. The use of this site is at your own risk. Any reliance you place on such information is therefore strictly at your own risk.
All official trademark data, including owner information, should be verified by visiting the official USPTO website at www.uspto.gov. This site is not intended to replace professional legal advice and should not be used as a substitute for consulting with a legal professional who is knowledgeable about trademark law.