Activity Modulator, Medicinal Agent Comprising Same, Use Of Cd300a Gene-deficient Mouse, And Anti-cd300a Antibody
Shibuya; Akira ; et al.
U.S. patent application number 16/656322 was filed with the patent office on 2020-02-06 for activity modulator, medicinal agent comprising same, use of cd300a gene-deficient mouse, and anti-cd300a antibody. This patent application is currently assigned to UNIVERSITY OF TSUKUBA. The applicant listed for this patent is UNIVERSITY OF TSUKUBA. Invention is credited to Syuichi Iino, Udayanga Sanath Kankanam Gamage, Haruka Miki, Tsukasa Nabekura, Chigusa Oda, Akira Shibuya, Satoko Tahara.
| Application Number | 20200040077 16/656322 |
| Document ID | / |
| Family ID | 48469634 |
| Filed Date | 2020-02-06 |






View All Diagrams
| United States Patent Application | 20200040077 |
| Kind Code | A1 |
| Shibuya; Akira ; et al. | February 6, 2020 |
ACTIVITY MODULATOR, MEDICINAL AGENT COMPRISING SAME, USE OF CD300A GENE-DEFICIENT MOUSE, AND ANTI-CD300A ANTIBODY
Abstract
The present invention aims to provide: an immunostimulant useful for maintaining, enhancing or suppressing an immune function associated with CD300a activation signaling, or an immunomodulator as an immunosuppressant useful for suppressing the immune function; use of a CD300a gene-deficient mouse for pathology analysis and the like; an anti-CD300a antibody; and the like.
| Inventors: | Shibuya; Akira; (Tsukuba-shi, JP) ; Oda; Chigusa; (Tsukuba-shi, JP) ; Tahara; Satoko; (Tsukuba-shi, JP) ; Nabekura; Tsukasa; (Tsukuba-shi, JP) ; Kankanam Gamage; Udayanga Sanath; (Tsukuba-shi, JP) ; Miki; Haruka; (Tsukuba-shi, JP) ; Iino; Syuichi; (Tsukuba-shi, JP) | ||||||||||
| Applicant: |
|
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Assignee: | UNIVERSITY OF TSUKUBA Tsukuba-shi JP |
||||||||||
| Family ID: | 48469634 | ||||||||||
| Appl. No.: | 16/656322 | ||||||||||
| Filed: | October 17, 2019 |
Related U.S. Patent Documents
| Application Number | Filing Date | Patent Number | ||
|---|---|---|---|---|
| 14359212 | Oct 8, 2014 | |||
| PCT/JP2012/078898 | Nov 7, 2012 | |||
| 16656322 | ||||
| Current U.S. Class: | 1/1 |
| Current CPC Class: | A61K 38/1774 20130101; A01K 2267/0368 20130101; A61K 38/1808 20130101; A01K 67/0276 20130101; C07K 2317/56 20130101; A61P 1/00 20180101; A61K 31/661 20130101; A61K 2039/54 20130101; A61K 2039/545 20130101; C07K 2317/76 20130101; A61K 31/04 20130101; A61K 2039/505 20130101; A61P 43/00 20180101; A61P 33/00 20180101; A61K 31/685 20130101; A61K 38/16 20130101; A61P 29/00 20180101; A61P 37/04 20180101; A01K 2227/105 20130101; A61P 31/04 20180101; C07K 16/2803 20130101; A61K 45/06 20130101; A61K 31/685 20130101; A61K 2300/00 20130101; A61K 38/16 20130101; A61K 2300/00 20130101; A61K 31/04 20130101; A61K 2300/00 20130101; A61K 31/661 20130101; A61K 2300/00 20130101 |
| International Class: | C07K 16/28 20060101 C07K016/28; A61K 38/16 20060101 A61K038/16; A61K 45/06 20060101 A61K045/06; A61K 31/661 20060101 A61K031/661; A61K 31/04 20060101 A61K031/04; A61K 31/685 20060101 A61K031/685; A01K 67/027 20060101 A01K067/027; A61K 38/17 20060101 A61K038/17; A61K 38/18 20060101 A61K038/18 |
Foreign Application Data
| Date | Code | Application Number |
|---|---|---|
| Nov 21, 2011 | JP | 2011-254151 |
Claims
1. A method for treatment of a disease or disease state in which inhibitory signal transduction of a cluster of differentiation 300a (CD300a)-expressing myeloid cell is involved, comprising: administering an effective amount of phosphatidyl serine, wherein said phosphatidyl serine promotes inhibitory signal transduction of a CD300a-expressing myeloid cell.
2. The method for treatment according to claim 1, wherein said disease is an inflammatory infection, allergic disease or autoimmune disease.
3. The method for treatment according to claim 1, wherein said disease is celiac disease.
4. A method for carrying out pathology analysis of atopic dermatitis, inflammatory bowel disease or asthma comprising use of a cluster of differentiation 300a (CD300a) gene-deficient mouse to which has been administered a substance that induces atopic dermatitis, inflammatory bowel disease or asthma as a model mouse that hardly develops atopic dermatitis, inflammatory bowel disease or asthma after administration of a substance that induces atopic dermatitis, inflammatory bowel disease or asthma.
5. The method for carrying out pathology analysis according to claim 4, wherein said substance that induces atopic dermatitis is mite antigen or ovalbumin, wherein said substance that induces inflammatory bowel disease is dextran sodium sulfate, and wherein said substance that induces asthma is mite antigen or ovalbumin.
6. A method for screening a candidate substance for efficacy in the treatment or prophylaxis of celiac disease, said method comprising: i) administering said candidate substance to a cluster of differentiation 300a (CD300a) gene-deficient mouse in which celiac disease has been induced following the administration to said mouse of a substance that induces celiac disease, and determining the presence or absence of a therapeutic effect; or ii) administering said candidate substance with a substance that induces celiac disease to a CD300a gene-deficient mouse, and determining the presence or absence of a prophylactic effect.
7. The method of claim 6, wherein said substance that induces celiac disease is a gluten-derived gliadin peptide.
8. An anti-human cluster of differentiation 300a (CD300a) antibody comprising: i) an H-chain variable region having the amino acid sequence of SEQ ID NO: 19 or an amino acid sequence that is the same as the amino acid sequence of SEQ ID NO: 19 except that 1, 2, 3, 4, or 5 amino acid(s) is/are each substituted, added, inserted or deleted in a region other than the complementarity determining regions of SEQ ID NO: 19; and ii) an L-chain variable region having the amino acid sequence of SEQ ID NO: 20 or an amino acid sequence that is the same as the amino acid sequence of SEQ ID NO: 20 except that 1, 2, 3, 4, or 5 amino acid(s) is/are each substituted, added, inserted or deleted in a region other than the complementarity determining regions of SEQ ID NO: 20.
9. An anti-mouse cluster of differentiation 300a (CD300a) antibody comprising: i) an H-chain variable region having the amino acid sequence of SEQ ID NO: 17 or an amino acid sequence that is the same as the amino acid sequence of SEQ ID NO: 17 except that 1, 2, 3, 4, or 5 amino acid(s) is/are each substituted, added, inserted or deleted in a region other than the complementarity determining regions of SEQ ID NO: 17; and ii) an L-chain variable region having the amino acid sequence of SEQ ID NO: 18 or an amino acid sequence that is the same as the amino acid sequence of SEQ ID NO: 18 except that 1, 2, 3, 4, or 5 amino acid(s) is/are each substituted, added, inserted or deleted in a region other than the complementarity determining regions of SEQ ID NO: 18.
10. A method of manufacturing a pharmaceutical composition, the method comprising: mixing the anti-human cluster of differentiation 300a (CD300a) antibody of claim 8 with a pharmaceutically acceptable carrier.
11. An anti-human cluster of differentiation 300a (CD300a) antibody comprising: i) an H-chain variable region having an amino acid sequence that is the same as the amino acid sequence of SEQ ID NO: 19 except that 1, 2, 3, 4, or 5 amino acid(s) is/are each substituted, added, inserted or deleted in a region other than the complementarity determining regions of SEQ ID NO: 19; and ii) an L-chain variable region having an amino acid sequence that is the same as the amino acid sequence of SEQ ID NO: 20 except that 1, 2, 3, 4, or 5 amino acid(s) is/are each substituted, added, inserted or deleted in a region other than the complementarity determining regions of SEQ ID NO: 20.
12. An anti-mouse cluster of differentiation 300a (CD300a) antibody comprising: i) an H-chain variable region having an amino acid sequence that is the same as the amino acid sequence of SEQ ID NO: 17 except that 1, 2, 3, 4, or 5 amino acid(s) is/are each substituted, added, inserted or deleted in a region other than the complementarity determining regions of SEQ ID NO: 17; and ii) an L-chain variable region having or an amino acid sequence that is the same as the amino acid sequence of SEQ ID NO: 18 except that 1, 2, 3, 4, or 5 amino acid(s) is/are each substituted, added, inserted or deleted in a region other than the complementarity determining regions of SEQ ID NO: 18.
13. A method for treatment of a disease or disease state in which inhibitory signal transduction of a cluster of differentiation 300a (CD300a)-expressing myeloid cell is involved via an immunoreceptor tyrosine-based inhibitory motif (ITIM) of CD300a, comprising: administering to a subject an effective amount of a neutralizing antibody against CD300a which inhibits binding between CD300a and phosphatidyl serine; wherein said neutralizing antibody against CD300a suppresses inhibitory signal transduction of the CD300a-expressing myeloid cell, and wherein said neutralizing antibody against CD300a is: an anti-human CD300a antibody comprising: i) an H-chain variable region having the amino acid sequence of SEQ ID NO: 19 or an amino acid sequence that is the same as the amino acid sequence of SEQ ID NO: 19 except that 1, 2, 3, 4, or 5 amino acid(s) is/are each substituted, added, inserted or deleted in the complementarity determining regions of SEQ ID NO: 19; and ii) an L-chain variable region having the amino acid sequence of SEQ ID NO: 20 or an amino acid sequence that is the same as the amino acid sequence of SEQ ID NO: 20 except that 1, 2, 3, 4, or 5 amino acid(s) is/are each substituted, added, inserted or deleted in the complementarity determining regions of SEQ ID NO: 20.
Description
CROSS-REFERENCE TO RELATED APPLICATION
[0001] This application is a Divisional of copending U.S. application Ser. No. 14/359,212, filed on Oct. 8, 2014, which is a 371 of International Application No. PCT/JP2012/078898, filed Nov. 7, 2012, which claims the benefit of priority from the prior Japanese Patent Application No. 2011-254151, filed on Nov. 21, 2011, the entire contents of all of which are incorporated herein by reference.
TECHNICAL FIELD
[0002] The present invention relates to an activity modulator for suppressing or promoting inhibitory signal transduction of a CD300a-expressing myeloid cell, a medicament comprising it, use of a CD300a gene-deficient mouse, and an anti-CD300a antibody.
BACKGROUND ART
[0003] Invasion of a pathogen (bacterium, virus, parasite or the like) into a host (human body or animal body) or generation of an endogenous inflammatory substance causes inflammatory reactions in which, for example, temporary contraction of arteriolae occurs at the site of invasion of the pathogen or the site of generation of the inflammatory substance, and expansion and hyperemia then occur, leading to local slowness of blood flow at the site of invasion of the pathogen or the site of generation of the inflammatory substance.
[0004] This causes adhesion of leukocytes to the vascular wall, and chemical mediators released from various immunocytes then act on the leukocytes to cause them to pass through the vascular wall by amoeboid movement and to allow their migration. Known examples of the chemical mediators include histamine, serotonin and lymphokines. Mast cells, which produce and release histamine and serotonin, are a type of lymphocytes that play a central role in the inflammatory reaction. Similarly to mast cells, macrophages also produce and release chemical mediators such as TNF.
[0005] The leukocytes whose migration was induced by the inflammatory reaction are attracted by the pathogen or the like, and this causes elimination (clearance) of the pathogen from the body by humoral immunity accompanied by antigen-antibody reaction and by cell-mediated immunity in which cytotoxic T cells and the like are involved, resulting in prevention of the spread of infection. Thus, the inflammatory reaction, and immune reactions that occur based on the inflammatory reaction, are extremely important for maintaining homeostasis of a living body.
[0006] On the other hand, the inflammatory reaction causes not only the biological defense described above, but also adverse signs/symptoms such as flare, fever, swelling, pain and dysfunction. Specific examples of such symptoms include allergic diseases, and various types of acute and chronic inflammations. Also in autoimmune diseases, in which the absence of immunological tolerance causes an autoimmune response, tissue injury occurs due to the inflammatory reaction.
[0007] That is, for prevention of a disease accompanied by the inflammatory reaction, it is important to kill the pathogen that causes the inflammatory reaction using antibiotics (antimicrobial agents), or to administer an agent that increases the immune function in the living body (immunostimulant) to eliminate the pathogen before an excessive inflammatory reaction occurs.
[0008] On the other hand, known examples of methods for amelioration or treatment of a disease accompanied by the inflammatory reaction include suppression of inflammation by administration of an agent (anti-inflammatory agent (immunosuppressant)) that decreases excessively activated immune function by, for example, suppression of release of chemical mediators.
[0009] For example, Patent Document 1 discloses, as an immunostimulant, an activating agent for the function of dendritic cells, which are antigen-presenting cells responsible for activation of various immunocytes. More specifically, the agent comprises as an effective component(s) at least one branched chain amino acid selected from isoleucine, leucine and valine.
[0010] Patent Document 2 discloses, as an anti-inflammatory agent (immunosuppressant), an agent comprising the SPARC (Secreted protein which is acidic and rich in cystein) peptide and a pharmaceutical carrier.
[0011] The following autoimmune diseases, allergic diseases and the like are known.
[0012] Celiac disease, or coeliac disease, is an autoimmune disease and is a progressive enteritis that is triggered by an immune reaction to gluten, which is a protein contained in wheat, barley, rye and the like. The incidence of this disease in the United Kingdom, Europe and the United States is one or more per 300 individuals.
[0013] Gluten, which is a mixture of two proteins gliadin and glutenin, is found in wheat, barley and rye. Gluten reacts with the small intestine and activates an immune system that attacks the fine small-intestinal epithelium, which is necessary for absorption of nutrients and vitamins, to cause injury.
[0014] When a patient with celiac disease takes a food or the like containing gluten, a gliadin-derived peptide, which cannot be digested by digestive enzymes of human and is contained in wheat as a fraction of a plant protein gluten, is deaminated by TG2 in the duodenal submucosal tissue to produce an antigen, which then causes production of autoantibodies.
[0015] Celiac disease occurs in genetically sensitive individuals having any of: HLA-DQ2 (which is retained in about 90% of individuals), which is encoded by HLA-DQA1 and HLA-DQB1; HLA-DQ2 mutants; and HLA-DQ8.
[0016] Such individuals show induction of an immune response to peptides induced from water-insoluble proteins in flour, gluten, and related proteins in rye and barley, which immune response is limited to inappropriate HLA-DQ2 and/or DQ8 and mediated by CD4.sup.+ T cells.
[0017] This immune reaction triggers an attack of the autoimmune system on the small-intestinal epithelial tissue to cause inflammation and then injury of villi and the like, leading to destruction of the epithelial cells themselves. As a result, nutrients cannot be absorbed from the small intestine, and the patient suffers from malnutrition irrespective of the dietary intake and the like.
[0018] Ulcerative colitis, which is a representative inflammatory bowel disease (IBD), is a collective term for chronic diseases that cause inflammation of unknown origin mainly in the digestive tract, and is a chronic disease in which inflammation occurs in the large intestine to form ulcers.
[0019] Inflammatory bowel disease (IBD) is a collective term used for describing two gastrointestinal disorders (Crohn's disease (CD) and ulcerative colitis (UC)) whose causes are unknown.
[0020] In Crohn's disease, a larger area is affected compared to ulcerative colitis, and inflammation and ulcers are found in almost the whole area of the digestive tract. Inflammatory bowel disease (IBD) occurs worldwide, and as many as two million people are reported to have suffered from Crohn's disease. The progression and prognosis of IBD widely vary.
[0021] In inflammatory bowel disease (IBD), diarrhea and bloody stool continue for a long period while the severity of symptoms changes with time. The cause of the disease has not been elucidated, and, in a major hypothesis, the continuous enteritis is thought to be caused by immune reactions to foods and enterobacteria due to disorder of immunological tolerance in the intestinal tract.
[0022] At present, there is no fundamental therapeutic method for the disease, and examples of therapies for inflammatory bowel disease include dietetic treatment; and use of an antidiarrheal (e.g., anticholinergic, loperamide or diphenoxylate), anti-inflammatory agent (e.g., steroid drug such as aminosalicylic acid, sulfasalazine, mesalamine, olsalazine, balsalazide or prednisolone; or aminosalicylic acid) or immunosuppressant (e.g., azathioprine, mercaptopurine or cyclosporin).
[0023] Surgical operations are required over a period of 10 years in 10% to 15% of the patients with IBD, and they have higher risk of occurrence of intestinal cancer.
[0024] In Crohn's disease, bacteria are involved in the onset and the progression of the disease, and intestinal inflammation in Crohn's disease is well-known for its frequent responsiveness to antibiotics and susceptibility to bacterial fecal flow. Common intestinal microorganisms and novel pathogens have been suggested to have associations with Crohn's disease, based on direct detection or anti-microbial immune responses associated with the disease.
[0025] Moreover, in a number of genetically susceptible models of chronic colitis, luminal microorganisms are indispensable cofactors for the disease, and animals kept in a microbe-free environment do not develop colitis.
[0026] The combination of genetic factors, exogenous causes and the endogenous microbiota may contribute to the immune-mediated injury of the intestinal mucosa found in inflammatory bowel disease.
[0027] It is also known that development of ulcerative colitis is associated with polymorphisms in 3 gene regions, that is, the FCGR2A gene, which encodes a receptor protein present on the surface of immunocytes; s8LC26A3, which encodes a transporter of chlorine ions and hydrogen carbonate ions; and a gene in the 13q12 region whose function is unknown (Non-patent Document 3).
[0028] However, in spite of abundant direct and indirect evidence on the role of intestinal microorganisms in Crohn's disease, no pathogenic organism or antigen has been identified to contribute to the impaired immunoregulation found in this disease. Tools useful for elucidation of the pathologies of the inflammatory bowel diseases (Crohn's disease and inflammatory enteritis), and medicaments for treatment and the like of these diseases have been demanded.
[0029] Atopic dermatitis is caused by entrance of an allergic substance (antigen) into the body followed by production of periostin due to stimulation by substances (interleukins 4 and 13) secreted from activated immunocytes, and then binding of the periostin to another protein "integrin" on the surface of keratinocytes in the skin, to cause inflammation.
[0030] The binding of periostin to integrin causes production of other inflammation-inducing substances, and the symptoms continue even in the absence of the antigen, resulting in chronicity. It has been shown, by an experiment using mice, that inhibition of binding of periostin to integrin using an inhibitor prevents occurrence of atopic dermatitis (Non-patent Document 4).
[0031] Although the major cause of atopic dermatitis has become evident, further elucidation of the pathology of atopic dermatitis, analysis of association of atopic dermatitis with other inflammatory diseases, and medicaments for atopic dermatitis that can be used in combination with the above inhibitor, are demanded.
[0032] Bronchial asthma is a respiratory disease in which bronchial inflammation triggered by an allergic reaction or infection with a bacterium or virus becomes chronic to thereby cause increased airway hyperresponsiveness and reversible airway narrowing, leading to symptoms such as attacks of wheezing, and cough. Further, bronchial asthma is said to be caused by the combination of airway hyperresponsiveness, allergic diathesis and environment. Recurrent symptoms such as wheezing, apnea, chest tightness and cough occur especially at night or in the early morning.
[0033] A number of cells and cellular components, especially mast cells, eosinophils, T-lymphocytes, macrophages, neutrophils and epithelial cells play roles in inflammation of the airway. Inflammation is associated with plasma exudation, edema, smooth muscle enlargement, mucus plugging, and epithelial changes. Further, inflammation causes associated increases in bronchial hyperresponsiveness to various stimuli.
[0034] Inflammation of the airway induces atrophy of airway smooth muscle, microvascular rupture and bronchial hyperresponsiveness. As the responsiveness of the airway increases, the symptoms become more severe and continuous, and daily variation of the pulmonary function increases. The mechanism of involvement of airway inflammation in the bronchial responsiveness is unknown, and tools useful for elucidation of the pathology of asthma, and medicaments and the like have been demanded.
[0035] It is known that a group of receptor molecules called MAIR (Myeloid Associated Ig like Receptors) are expressed on the cell membrane of myeloid (bone marrow) cells responsible for natural immunity (Non-patent Document 1). Among these, MAIR-I, which is also known as CD300a (also referred to as "LMIR1" or "CLM-8"), is expressed in macrophages, mast cells, granulocytes (neutrophils) and dendritic cells, and known to be an inhibitory receptor that associates with phosphatase via the ITIM (Immunoreceptor tyrosine-based inhibitory motif) sequence in the intracellular domain to transmit an inhibitory signal (Non-patent Document 2). However, the ligand for this receptor is unknown, and the receptor has been the so-called orphan receptor.
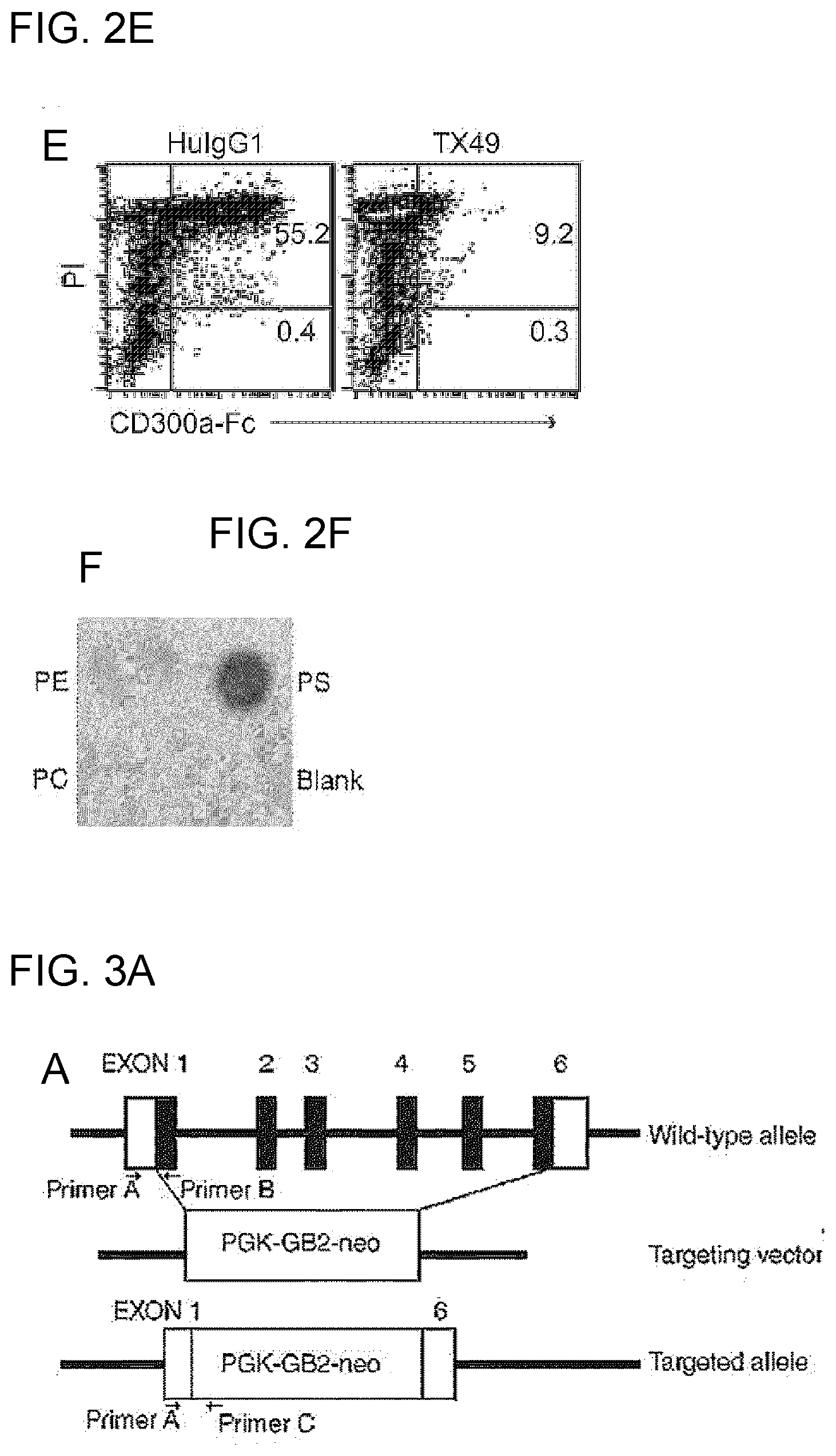
PRIOR ART DOCUMENTS
Patent Documents
[0036] [Patent Document 1] JP 2007-297379 A [0037] [Patent Document 2] JP 2011-516609 A
Non-Patent Documents
[0037] [0038] [Non-patent Document 1] Yotsumoto et al., J Exp Med 198 (2), 223-233, 2003 [0039] [Non-patent Document 2] Okoshi Y et al., Int Immunol., 17, 65-72, 2005. [0040] [Non-patent Document 3] Asano K et al., Nature Genetics 41. 1325-1329 (2009) [0041] [Non-patent Document 4] Miho Masuoka et al., J Clin Invest. 2012; doi: 10. 1172/JC158978
SUMMARY OF THE INVENTION
Problems to be Solved by the Invention
[0042] The present invention aims to elucidate the regulatory mechanism of activation of the innate immune response via the inhibitory signal transduction of CD300a, and to provide means for regulating the activity of CD300a-expressing cells involved in the inhibitory signal transduction, means for treating or preventing diseases or disease states in which the activity is involved, and techniques and the like associated with these.
Means for Solving the Problems
[0043] In order to elucidate the ligand of CD300a and the function of CD300a, the present inventors intensively studied to discover the following.
[0044] (i) The ligand of CD300a is phosphatidyl serine (PS).
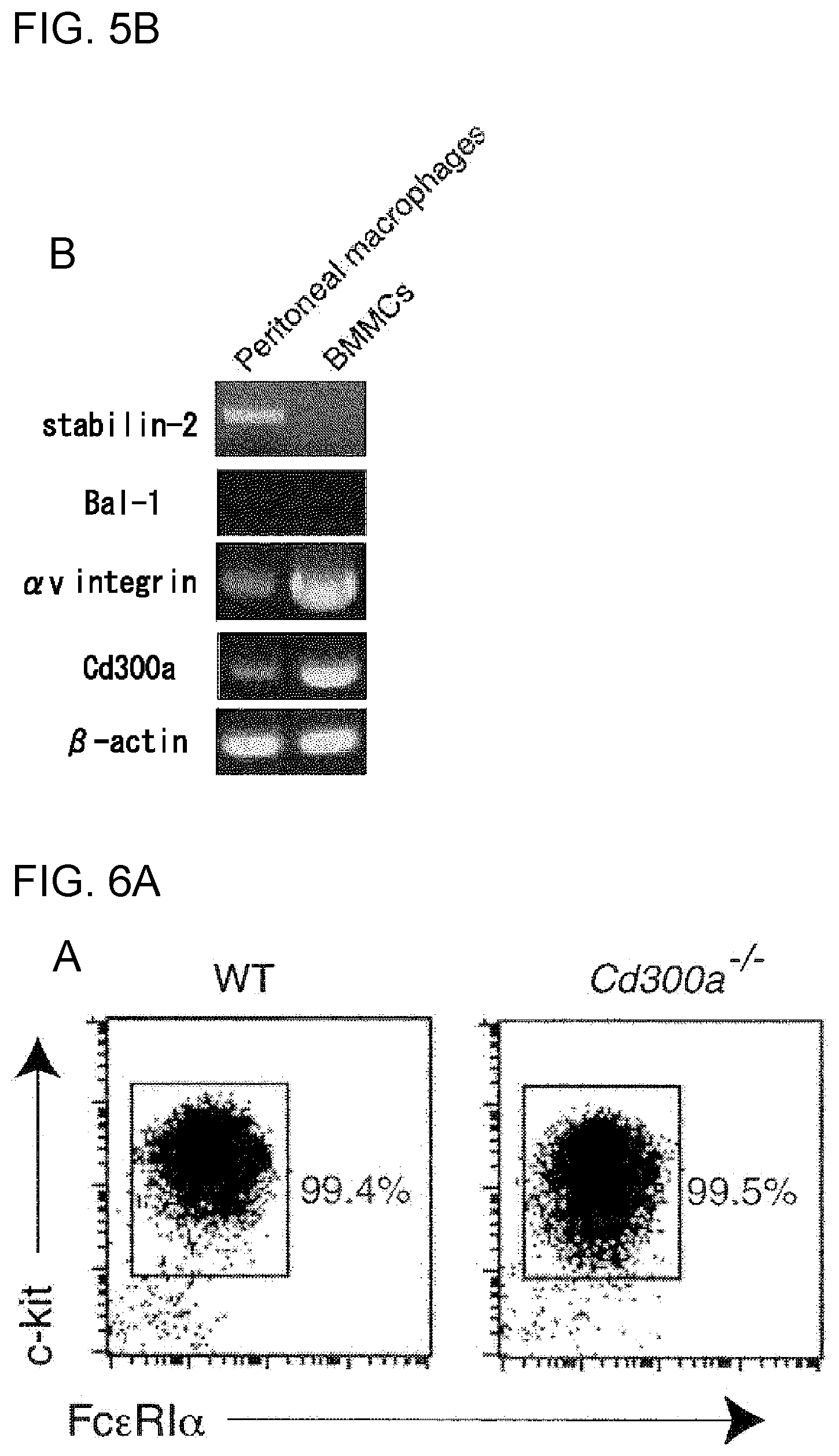
[0045] (ii) Binding of PS to CD300a on mast cells and the like promotes inhibitory signal transduction via CD300a, and activation of the mast cells and the like are also suppressed thereby.
[0046] (iii) Inhibition of binding of CD300a on mast cells and the like to PS by coexistence of a phosphatidyl serine-binding substance or CD300a-binding substance suppresses inhibitory signal transduction of CD300a, and the active state of mast cells and the like is maintained.
[0047] (iv) Through the suppression or maintenance of the active state, inflammatory infections, allergic diseases, autoimmune diseases and the like can be treated.
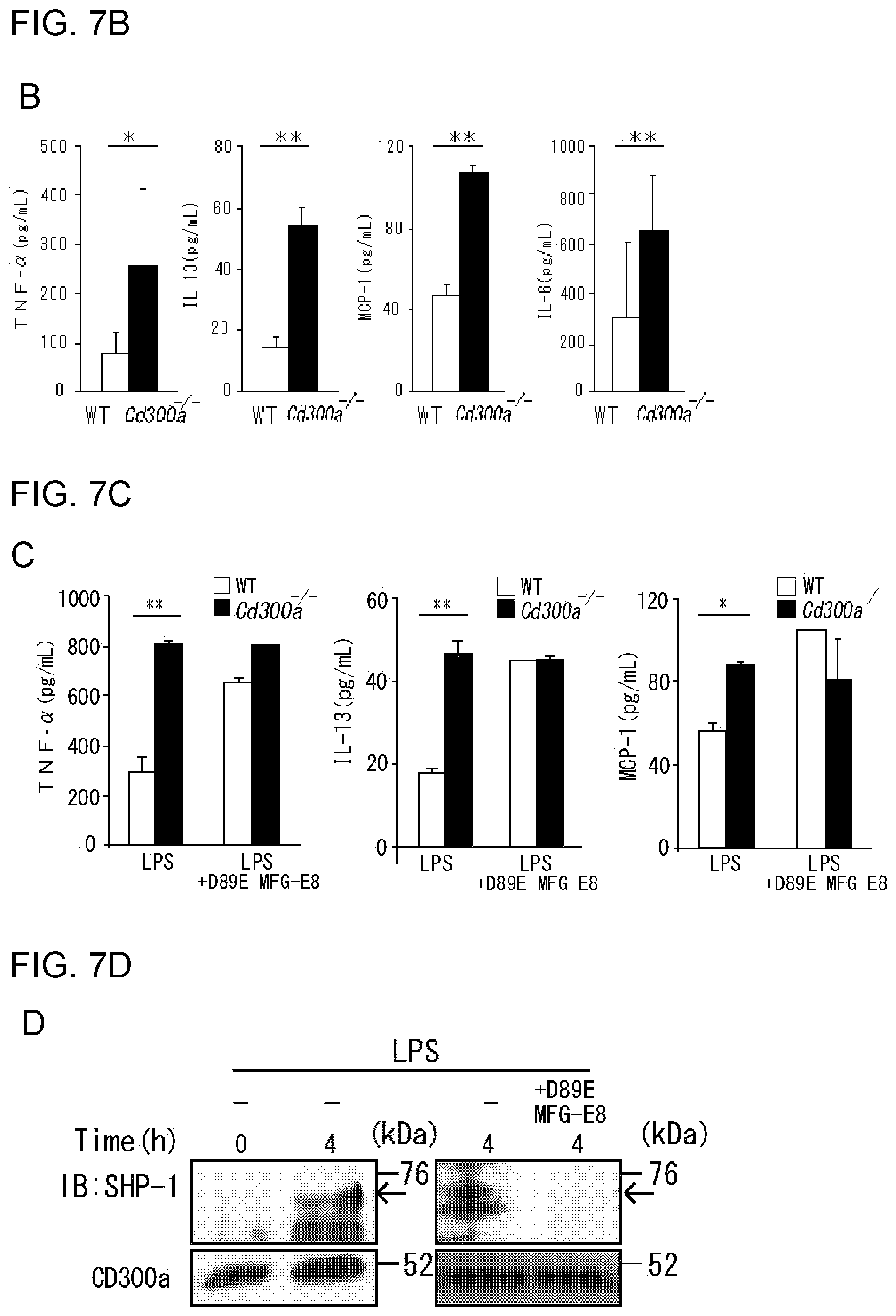
[0048] (v) CD300a gene-deficient mice can be a tool for performing pathology analysis of allergic diseases and autoimmune diseases, and screening of effective components of medicaments.
[0049] The present invention was carried out based on these discoveries, and provides, for example, the activity modulators, medicaments, use of a CD300a gene-deficient mouse, anti-CD300a antibody, and the like described in [1] to [23] below.
[0050] [1] An activity modulator for suppressing inhibitory signal transduction of a CD300a-expressing myeloid cell, the activity modulator comprising a substance that inhibits binding of CD300a to phosphatidyl serine.
[0051] [2] The activity modulator according to [1], wherein the substance that inhibits binding of CD300a to phosphatidyl serine is a phosphatidyl serine-binding substance.
[0052] [3] The activity modulator according to [2], wherein the phosphatidyl serine-binding substance is at least one selected from the group consisting of MFG-E8, MFG-E8 mutants, T cell immunoglobulin, soluble TIM-1, soluble TIM-4, soluble stabilin and soluble integrin .alpha.v.beta.3.
[0053] [4] The activity modulator according to [1], wherein the substance that inhibits binding of CD300a to phosphatidyl serine is a CD300a-binding substance.
[0054] [5] The activity modulator according to [4], wherein the CD300a-binding substance is an anti-human CD300a antibody comprising: an H-chain variable region having the amino acid sequence of SEQ ID NO:19 or an amino acid sequence that is the same as the amino acid sequence except that 1, 2, 3, 4, or 5 amino acid(s) is/are substituted, added, inserted and/or deleted; and an L-chain variable region having the amino acid sequence of SEQ ID NO:20 or an amino acid sequence that is the same as the amino acid sequence except that 1, 2, 3, 4, or 5 amino acid(s) is/are substituted, added, inserted and/or deleted.
[0055] [6] The activity modulator according to [4], wherein the CD300a-binding substance is an anti-mouse CD300a antibody comprising: an H-chain variable region having the amino acid sequence of SEQ ID NO:17 or an amino acid sequence that is the same as the amino acid sequence except that 1, 2, 3, 4, or 5 amino acid(s) is/are substituted, added, inserted and/or deleted; and an L-chain variable region having the amino acid sequence of SEQ ID NO:18 or an amino acid sequence that is the same as the amino acid sequence except that 1, 2, 3, 4, or 5 amino acid(s) is/are substituted, added, inserted and/or deleted.
[0056] [7] An activity modulator for promoting inhibitory signal transduction of a CD300a-expressing myeloid cell, the activity modulator comprising a substance that promotes binding of CD300a to phosphatidyl serine.
[0057] [8] The activity modulator according to [7], wherein the substance that promotes binding of CD300a to phosphatidyl serine is phosphatidyl serine.
[0058] [9] A medicament for treatment or prophylaxis of a disease or disease state in which inhibitory signal transduction of a CD300a-expressing myeloid cell is involved, the medicament comprising the activity modulator according to any of [1] to [8].
[0059] [10] The medicament according to [9], wherein the disease or disease state in which inhibitory signal transduction of a CD300a-expressing myeloid cell is involved is an inflammatory infection, allergic disease or autoimmune disease.
[0060] [11] The medicament according to [10] for treatment or prophylaxis of peritonitis or sepsis caused thereby, the medicament comprising the activity modulator according to any of [1] to [6].
[0061] [12] The medicament according to [10] for treatment of inflammatory bowel disease, the medicament comprising the activity modulator according to any of [1] to [6].
[0062] [13] The medicament according to [10] for treatment of celiac disease, the medicament comprising the activity modulator according to [7] or [8].
[0063] [14] The medicament according to [10] for treatment of atopic dermatitis, the medicament comprising the activity modulator according to any of [1] to [6].
[0064] [15] The medicament according to [10] for treatment of asthma, the medicament comprising the activity modulator according to any of [1] to [6].
[0065] [16] Use of a CD300a gene-deficient mouse for carrying out pathology analysis of an allergic disease or autoimmune disease, or for screening of a possible candidate substance for an effective component of a therapeutic agent or prophylactic agent for the disease.
[0066] [17] The use according to [16], comprising use of the CD300a gene-deficient mouse as a model mouse in which inflammatory bowel disease is hardly induced after administration of a substance that induces inflammatory bowel disease.
[0067] [18] The use according to [16], comprising use of the CD300a gene-deficient mouse as a model mouse that develops celiac disease after administration of a substance that induces celiac disease.
[0068] [19] The use according to [16], comprising the step of: administering a candidate substance for a therapeutic agent for celiac disease to the CD300a gene-deficient mouse that developed celiac disease, and determining the presence or absence of a therapeutic effect; or administering a candidate substance for a prophylactic agent for celiac disease together with the substance that induces celiac disease to the CD300a gene-deficient mouse before development of celiac disease, and determining the presence or absence of a prophylactic effect.
[0069] [20] The use according to [16], comprising use of the CD300a gene-deficient mouse as a model mouse that hardly develops atopic dermatitis after administration of a substance that induces atopic dermatitis.
[0070] [21] The use according to [16], comprising use of the CD300a gene-deficient mouse as a model mouse that hardly develops asthma after administration of a substance that induces asthma.
[0071] [22] An anti-human CD300a antibody comprising: an H-chain variable region having the amino acid sequence of SEQ ID NO:19 or an amino acid sequence that is the same as the amino acid sequence except that 1, 2, 3, 4, or 5 amino acid(s) is/are substituted, added, inserted and/or deleted; and an L-chain variable region having the amino acid sequence of SEQ ID NO:20 or an amino acid sequence that is the same as the amino acid sequence except that 1, 2, 3, 4, or 5 amino acid(s) is/are substituted, added, inserted and/or deleted.
[0072] [23] An anti-mouse CD300a antibody comprising: an H-chain variable region having the amino acid sequence of SEQ ID NO:17 or an amino acid sequence that is the same as the amino acid sequence except that 1, 2, 3, 4, or 5 amino acid(s) is/are substituted, added, inserted and/or deleted; and an L-chain variable region having the amino acid sequence of SEQ ID NO:18 or an amino acid sequence that is the same as the amino acid sequence except that 1, 2, 3, 4, or 5 amino acid(s) is/are substituted, added, inserted and/or deleted.
[0073] As other aspects of the inventions described above, the following inventions are provided.
[0074] Another aspect of the invention of [1] provides a method for suppressing inhibitory signal transduction of a CD300a-expressing myeloid cell, which method comprises inhibiting binding of CD300a to phosphatidyl serine. This method is applicable either in vivo or ex vivo/in vitro, and, in cases of in vivo application, the species of organism may be either human or non-human (e.g., mammal such as mouse).
[0075] Still another aspect of the invention of [1] provides use of a substance that inhibits binding of CD300a to phosphatidyl serine in production of an activity modulator for suppressing inhibitory signal transduction of a CD300a-expressing myeloid cell.
[0076] As the substance that inhibits binding of CD300a to phosphatidyl serine for carrying out the method or use, the phosphatidyl serine-binding substance recited in [2], preferably MFG-E8 or the like recited in [3] may be used. Similarly, as the substance that inhibits binding of CD300a to phosphatidyl serine for carrying out the method, the CD300a-binding substance recited in [4], preferably the anti-human CD300a antibody comprising an H-chain variable region and an L-chain variable region having the prescribed amino acid sequences recited in [5], or the anti-mouse CD300a antibody comprising an H-chain variable region and an L-chain variable region having the prescribed amino acid sequences recited in [6], may be used.
[0077] Another aspect of the invention of [7] provides a method for promoting inhibitory signal transduction of a CD300a-expressing myeloid cell, which method comprises promotion of binding of CD300a to phosphatidyl serine. This method is applicable either in vivo or ex vivo/in vitro, and, in cases of in vivo application, the species of organism may be either human or non-human (e.g., mammal such as mouse).
[0078] Still another aspect of the invention of [7] provides use of a substance that promotes binding of CD300a to phosphatidyl serine in production of an activity modulator for promoting inhibitory signal transduction of a CD300a-expressing myeloid cell.
[0079] As the substance that promotes binding of CD300a to phosphatidyl serine for carrying out the method or use, the phosphatidyl serine recited in [8] may be used.
[0080] Another aspect of the invention of [9] provides a method for treatment or prophylaxis of a disease or disease state in which inhibitory signal transduction of a CD300a-expressing myeloid cell is involved, which method comprises inhibiting or promoting binding of CD300a to phosphatidyl serine, thereby suppressing or promoting inhibitory signal transduction of a CD300a-expressing myeloid cell. This method is applicable either in vivo or ex vivo/in vitro, and, in cases of in vivo application, the species of organism may be either human or non-human (e.g., mammal such as mouse).
[0081] Still another aspect of the invention of [9] provides use of an activity modulator that inhibits or promotes binding of CD300a to phosphatidyl serine to thereby suppress or promote inhibitory signal transduction of a CD300a-expressing myeloid cell, in production of a medicament for treatment or prophylaxis of a disease or disease state in which inhibitory signal transduction of a CD300a-expressing myeloid cell is involved.
[0082] Examples of the disease or disease state in the method or use include inflammatory infection, allergic disease, and autoimmune disease as recited in [10], and specific examples of the disease or disease state include the peritonitis or sepsis caused thereby, inflammatory bowel disease, celiac disease, atopic dermatitis, and asthma as recited in [11] to [15].
[0083] Another aspect of the invention of [16] provides a method for carrying out pathology analysis of an allergic disease or autoimmune disease, or for screening of a possible candidate substance for an effective component of a therapeutic agent or prophylactic agent for the disease, by using a CD300a gene-deficient mouse. The CD300a gene-deficient mouse used in the method is a model mouse in which inflammatory bowel disease, atopic dermatitis or asthma is hardly induced, or a model mouse that develops celiac disease as recited in [17], [18], [20] or [21] after administration of a substance that induces each allergic disease or autoimmune disease. The method is especially preferably a method for celiac disease, comprising the prescribed steps recited in [19].
Effect of the Invention
[0084] The present invention enables preparation of an activity modulator that can suppress or promote inhibitory signal transduction of a CD300a-expressing myeloid cell, and production of a medicament for treatment or prophylaxis of a disease or disease state in which inhibitory signal transduction of a CD300a-expressing myeloid cell is involved, which medicament comprises as an effective component the activity modulator. Further, the present invention enables use of a CD300a gene-deficient mouse as a model mouse or the like, and production of an anti-CD300a antibody having excellent neutralizing action.
BRIEF DESCRIPTION OF THE DRAWINGS
[0085] FIG. 1 shows the results of flow cytometry analysis obtained in Example 1A.
[0086] FIG. 2A shows the results of flow cytometry analysis obtained in Example 1B.
[0087] FIG. 2B shows the results of flow cytometry analysis obtained in Example 1B.
[0088] FIG. 2C shows the results of flow cytometry analysis obtained in Example 1C.
[0089] FIG. 2D shows the results of flow cytometry analysis obtained in Example 1D.
[0090] FIG. 2E shows the results of flow cytometry analysis obtained in Example 1D.
[0091] FIG. 2F shows the results of immunoblotting analysis obtained in Example 1E.
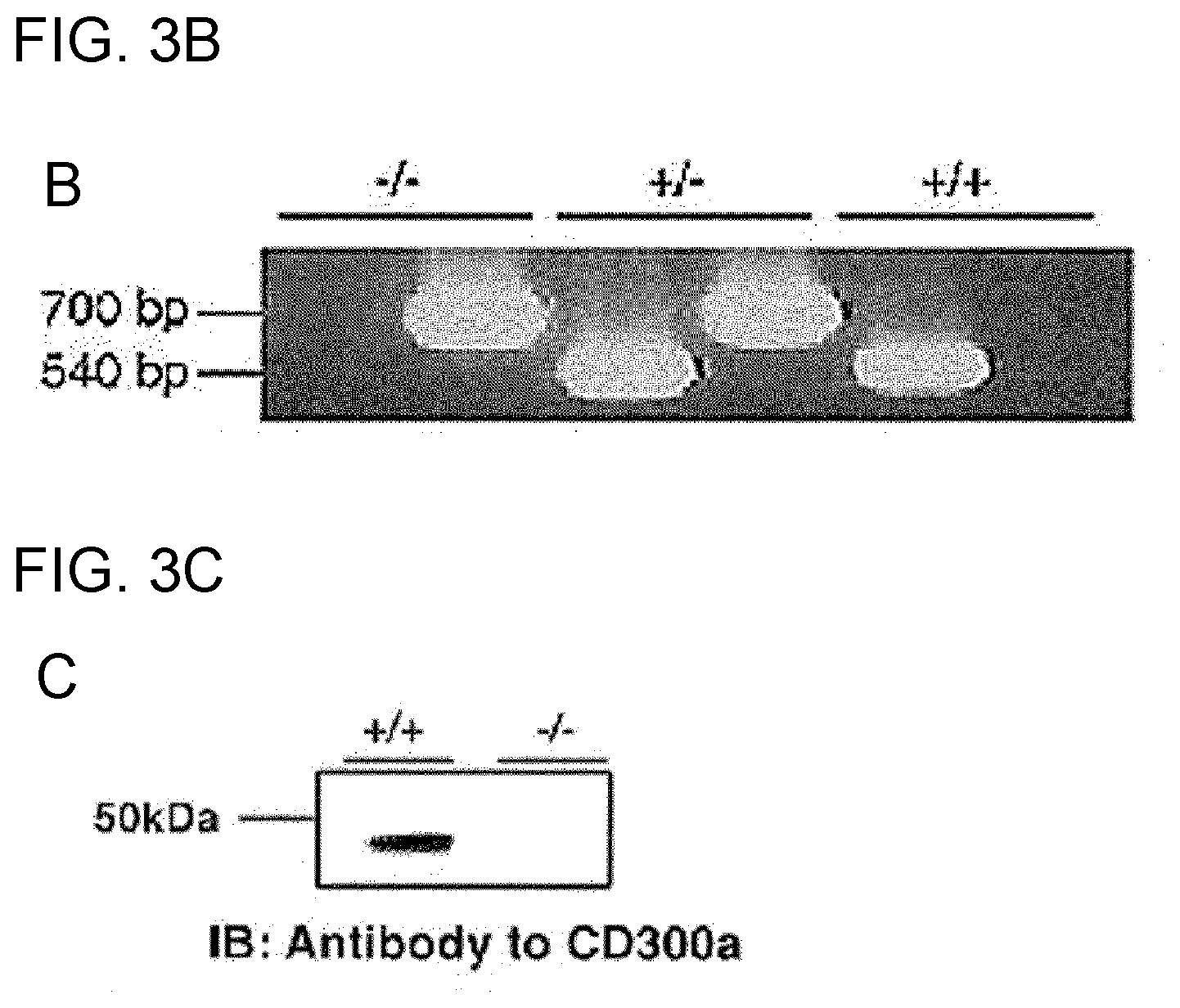
[0092] FIG. 3A is a schematic diagram for illustrating the structure of the CD300a gene in the wild-type allele, the targeting vector used for preparing a CD300a gene-deficient mouse, and the targeted allele.
[0093] FIG. 3B is a photograph taken after electrophoresis of PCR products from the wild-type allele and the mutant allele.
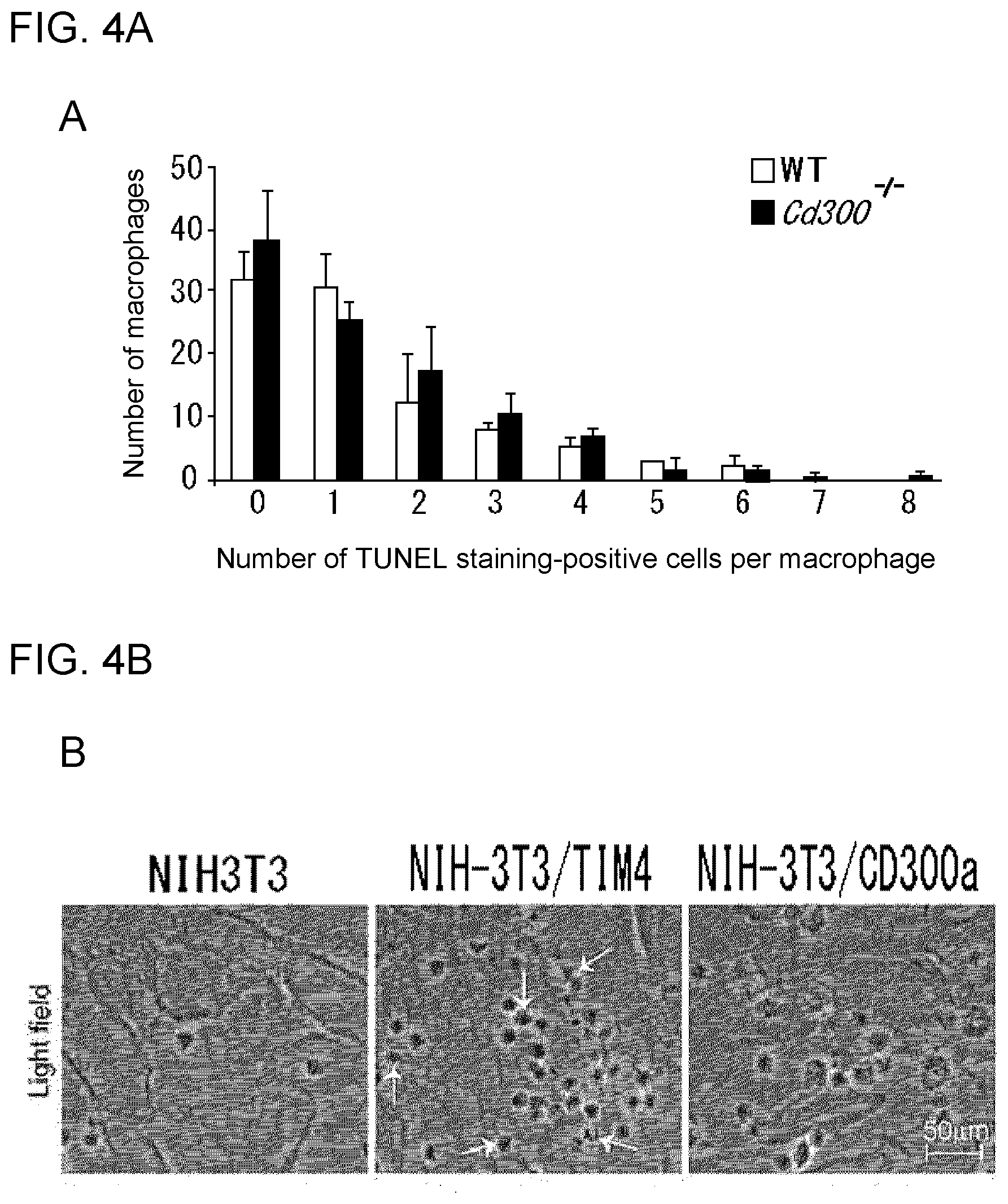
[0094] FIG. 3C shows the results of Western blotting using a wild-type mouse and a CD300a-deficient mouse.
[0095] FIG. 3D shows the results of flow cytometry analysis of a WT mouse and a CD300a gene-deficient mouse.
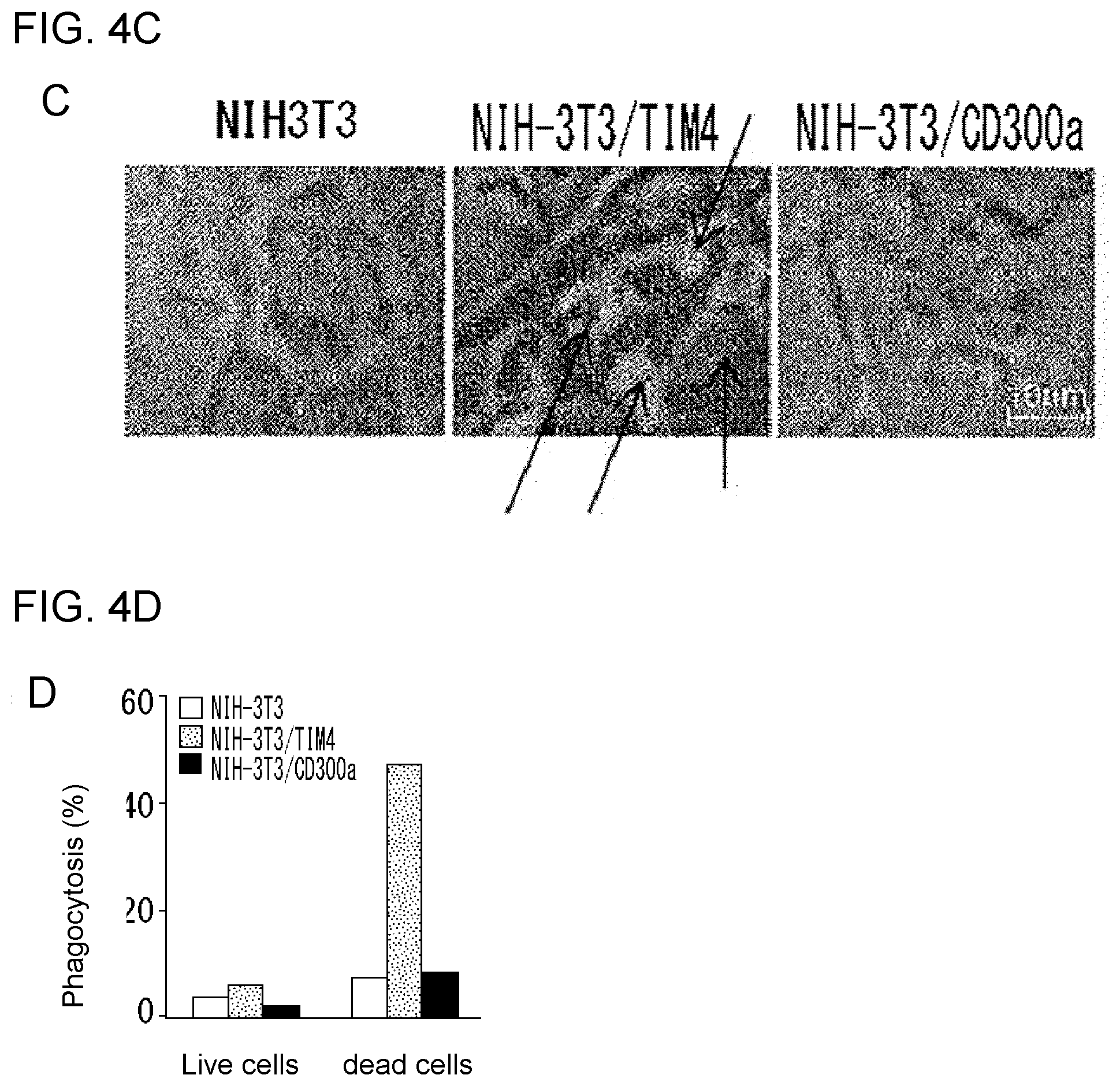
[0096] FIG. 4A shows the results of flow cytometry analysis obtained in Example 2A.
[0097] FIG. 4B shows the results of analysis under a light microscope obtained in Example 2C.
[0098] FIG. 4C shows the results of laser scanning confocal microscopy obtained in Example 2C.
[0099] FIG. 4D shows the results obtained in Example 2C illustrating the ratio of the number of cells of NIH3T3 or each transfectant showing incorporation of a thymocyte in the cytoplasm.
[0100] FIG. 5A shows the results of flow cytometry analysis obtained in Example 2B.
[0101] FIG. 5B shows the results of RT-PCR analysis obtained in Example 2B.
[0102] FIG. 6A shows the results of flow cytometry analysis obtained in Example 3A.
[0103] FIG. 6B shows the results of analysis by the .beta.-hexaminidase assay obtained in Example 3 A.
[0104] FIG. 7A shows the results of flow cytometry analysis obtained in Example 3B.
[0105] FIG. 7B shows the results obtained in Example 3C on the rates of increase in the amounts of various cytokines and chemokines released.
[0106] FIG. 7C shows a diagram showing the results obtained in Example 3E on the rates of increase in the amounts of various cytokines and chemokines released.
[0107] FIG. 7D shows the results of immunoblotting analysis obtained in Example 3F.
[0108] FIG. 7E shows a diagram showing the results of immunoblotting analysis obtained in Example 3G.
[0109] FIG. 7F shows a diagram showing the rate of increase in TNF-.alpha., obtained in Example 3G.
[0110] FIG. 8 shows the results of flow cytometry analysis obtained in Example 3D.
[0111] FIG. 9A shows the results of densitometric analysis obtained in Example 4B.
[0112] FIG. 9B shows the results of densitometric analysis obtained in Example 4B.
[0113] FIG. 10A shows a diagram showing the results of calculation of the CFU of aerobic bacteria, obtained in Example 4C.
[0114] FIG. 10B shows a diagram showing the numbers of neutrophils and macrophages obtained in Example 4C after induction of CD300a neutrophils.
[0115] FIG. 11 shows a diagram showing the rate of the number of each type of macrophages that showed phagocytosis of E. coli.
[0116] FIG. 12A shows a diagram showing the results of flow cytometry analysis obtained in Example 4D.
[0117] FIG. 12B shows a diagram showing the rate of survival of each type of mice, obtained in Example 4D.
[0118] FIG. 12C shows a diagram showing the bacterial clearance in the intestine in each type of mice in Example 4D, in terms of the bacterial CFU.
[0119] FIG. 12D shows the results of flow cytometry analysis obtained in Example 4E.
[0120] FIG. 12E shows a diagram showing the rate of survival of each group of mice after administration of TX41 in Example 4F.
[0121] FIG. 12F shows a diagram showing the clearance in the intestine after administration of TX41 in Example 4G.
[0122] FIG. 12G shows a diagram showing the result of analysis of the change in the number of neutrophils after administration of TX41 in Example 4G.
[0123] FIG. 13A shows a diagram illustrating the protocol for induction of asthma with ovalbumin.
[0124] FIG. 13B shows the total cell number in the alveolar lavage fluid from each mouse on Day 25 after the beginning of induction of asthma in Example 5A.
[0125] FIG. 13C shows the ratio of eosinophils in the alveolar lavage fluid from each mouse on Day 25 after the beginning of induction of asthma in Example 5B.
[0126] FIG. 14 shows the serum IgE value in each mouse on Day 14 after the beginning of induction of asthma in Example 5B.
[0127] FIG. 15 shows a diagram showing changes in the body weight with time due to DSS-induced enteritis in each group of mice (Example 6A).
[0128] FIG. 16A shows a graph showing the length of the large intestine in each type of mice on Day 6 after the beginning of administration of DSS (Example 6B).
[0129] FIG. 16B shows a diagram showing sections of large intestines of a WT mouse and CD300a.sup.-/- (Example 6C).
[0130] FIG. 17 shows a diagram for comparison of the degree of cell damage in each type of mice on Day 6 of administration of DSS, based on the clinical score (Example 6D).
[0131] FIG. 18A shows a diagram showing the amount of each cytokine in the large intestine of each mouse on Day 9 of administration of DSS (Example 6E).
[0132] FIG. 18B shows a diagram showing the number of each type of immunocytes on Day 0, Day 2, Day 4 and Day 6 after the beginning of administration of DSS in each type of mice (Example 6F).
[0133] FIG. 19 shows a diagram showing the results of flow cytometry that was performed for identifying dendritic cells expressing CD300a (Example 6G).
[0134] FIG. 20 shows a diagram showing the relative gene expression levels of cytokines in the large intestine in each type of mice (Example 6H).
[0135] FIG. 21 shows a diagram showing the relative gene expression levels of cytokines in CD4.sup.+ T cells (Example 61).
[0136] FIG. 22 shows a diagram showing changes with time (daily changes) in the number of times of scratching behavior per 30 minutes in each type of OVA-sensitized mouse (Example 7A).
[0137] FIG. 23 shows sections of the skin of each mouse at the end of the 3rd week after sensitization with OVA (Example 7B).
[0138] FIG. 24 shows a diagram showing the result of toluidine blue staining of a skin sample from each mouse at the end of the 3rd week after sensitization with OVA (Example 7C).
[0139] FIG. 25A shows a graph showing the number of cell layers in the epidermis in each group of mice at the end of the 3rd week after sensitization with OVA (Example 7D).
[0140] FIG. 25B shows the numbers of eosinophils and mast cells that showed infiltration into the dermis in skin samples of each group of mice at the end of the 3rd week after sensitization with OVA (Example 7E).
[0141] FIG. 26 shows the result of Langerin immunostaining of a skin sample from each group of mice at the end of the 3rd week after sensitization with OVA (Example 7F).
[0142] FIG. 27 shows counterstaining of each sample in FIG. 26.
[0143] FIG. 28 shows a diagram showing a state where mast cells are interacting with Langerin-positive skin cells (Example 7H).
[0144] FIG. 29 is a diagram showing a schedule of the test for confirming the therapeutic effect of TX41.
[0145] FIG. 30 shows the total serum IgE level in WT mice after administration of TX74 or TX41, as measured by the ELISA method.
[0146] FIG. 31 shows a diagram showing the number of times of scratching behavior in WT mice after administration of TX74 or TX41.
[0147] FIG. 32 shows a diagram showing the result of H&E staining of a skin section of a WT mouse after administration of TX74 or TX41.
[0148] FIG. 33 shows a diagram showing a state in which a skin section of a WT mouse after administration of TX74 or TX41 was counterstained by toluidine blue staining.
[0149] FIG. 34 shows a diagram showing changes in the body weight (BW) with time monitored from the beginning of feeding with a normal diet or high-gluten diet.
[0150] FIG. 35 shows photographs each showing the result of histopathological analysis of the intestine of a mouse at Week 20 after the beginning of feeding with a normal diet or high-gluten diet.
[0151] FIG. 36 shows a diagram showing the clinical score at Week 20 after the beginning of feeding with a normal diet or high-gluten diet in each group of mice.
[0152] FIG. 37 shows a diagram showing the number of intraepithelial lymphocytes per 100 intestinal epithelium cells at week 20 after the beginning of feeding with a normal diet or high-gluten diet in each group of mice.
[0153] FIG. 38 shows a diagram showing the amount of transglutaminase 2 (TG2) in a suspension of the jejunum at week 20 after the beginning of feeding with a normal diet or high-gluten diet in each type of mice.
[0154] FIG. 39 shows a diagram showing the results of flow cytometry analysis of expression of CD11b and CD11c in cells of the lamina propria gated for the CD45.sup.+PI.sup.- cell population.
[0155] FIG. 40 shows a diagram showing the frequency of each type of immunocytes in the jejunal lamina propria in WT mice and CD300a gene-deficient mice kept with a normal diet or high-gluten diet.
[0156] FIG. 41 shows a diagram showing the gene expression levels of various cytokines and chemokines in lamina propria (LP) macrophages in each group of mice.
[0157] FIG. 42 shows a diagram showing the result of analysis of expression of CD300a (MAIR-I) in each type of immunocytes by flow cytometry.
[0158] FIG. 43 shows a diagram showing the relative gene expression levels of cytokines and chemokines in CD11b.sup.+ dendritic cells in the lamina propria (LP) in each type of mice.
[0159] FIG. 44 shows a diagram showing changes in the expression levels of gliadin-induced cytokines caused by addition of a recombinant mouse MFG-E8 protein.
[0160] FIG. 45 shows a diagram showing the result of histopathological analysis of the large intestine of a Balb/c WT mouse or CD300a gene-deficient mouse fed with a normal diet or high-gluten diet.
[0161] FIG. 46 shows a diagram showing the anti-gliadin IgG and IgA antibody titers in sera derived from Balb/c WT or CD300a gene-deficient mice.
[0162] FIG. 47 shows a diagram showing changes in the rate of change in the body weight with time (weekly changes) in WT mice and CD300a gene-deficient mice fed with a gluten-free diet.
[0163] FIG. 48 shows a diagram showing the expression levels of cytokines in CD11b.sup.+ dendritic cells and macrophages in the normal state in the lamina propria (LP) in WT mice and CD300a gene-deficient mice.
[0164] FIG. 49 shows a diagram showing the expression levels of cytokines in macrophages in thioglycolate-induced peritoneal exudate cells after stimulation with the gliadin peptide P31-43.
[0165] FIG. 50 is a diagram showing the expression levels of IL-6, IL-15, TNF-.alpha., IFN-.beta., MCP1 and MCP5 in CD11b.sup.+ dendritic cells in the lamina propria.
[0166] FIG. 51 shows a diagram showing the expression levels of IL-6, TNF-.alpha. and IFN-.beta. in lamina propria (LP) macrophages in B6 WT mice or CD300a gene-deficient mice after stimulation with gliadin.
[0167] FIG. 52 shows a diagram showing expression of cytokines and chemokines in lamina propria (LP) macrophages (stimulated with gliadin) derived from microbiota-depleted WT mice or CD300a gene-deficient mice.
[0168] FIG. 53 shows a diagram showing the frequency of phosphatidyl serine-expressing cells in isolated lamina propria (LP) macrophages.
[0169] FIG. 54 shows a diagram showing the gene expression levels of .alpha.v and .beta.3 integrin subunits in lamina propria (LP) macrophages in WT mice and CD300a gene-deficient mice.
[0170] FIG. 55 shows a diagram showing the gene expression levels of phosphatidyl serine receptors (TIM-1, TIM-4, Stabiln-2, SR-PSOX, BAI1 and Mer).
[0171] FIG. 56 shows the results of homology analysis of each of the H-chain and L-chain of TX41 and TX49.
DESCRIPTION OF EMBODIMENTS
[0172] The activity modulator of the present invention, a medicament comprising it, use of a CD300a gene-deficient mouse, and an anti-CD300a antibody, are described below in detail. Literatures used for mentioning conventional knowledge or a known test method on the immune mechanism, CD300a or the like are listed in the end of Examples.
[0173] [Activity Modulator]
[0174] The activity modulator of the present invention includes those for suppressing inhibitory signal transduction of a CD300a-expressing myeloid cell, as well as those for promoting such inhibitory signal transduction.
[0175] The "CD300a-expressing myeloid cell" herein includes a mast cell, macrophage, neutrophil, dendritic cell and the like. CD300a is a collective term for those expressed in mammals such as human and mouse, and the species of organism is not limited.
[0176] The "inhibitory signal transduction" is signal transduction that occurs by association of the inhibitory receptor CD300a with phosphatase via the ITIM (Immunoreceptor tyrosine-based inhibitory motif) sequence in the intracellular domain.
[0177] In the following description, since the activity modulator for suppressing inhibitory signal transduction of a CD300a-expressing myeloid cell may have an action to activate an immune function as a result, it is also referred to as "immunostimulant" in the present invention (for example, in cases where it is used as an effective component of a medicament for an inflammatory infection). On the other hand, since the activity modulator for promoting inhibitory signal transduction of a CD300a-expressing myeloid cell may have an action to suppress an immune function as a result, it is also referred to as "immunosuppressant" in the present invention (for example, in cases where it is used as an effective component of a medicament for celiac disease).
[0178] <First Activity Modulator>
[0179] The first activity modulator by the present invention comprises a component having an action to suppress inhibitory signal transduction via CD300a. In the present invention, as such a component, a substance that inhibits binding of phosphatidyl serine to CD300a, that is, a phosphatidyl serine-binding substance or CD300a-binding substance may be used. The first activity modulator may contain either one of these, or may contain both of these.
[0180] (Phosphatidyl Serine-Binding Substance)
[0181] The phosphatidyl serine-binding substance as a first activity modulator is not limited as long as it binds to phosphatidyl serine (PS), which is a ligand of CD300a, to inhibit interaction (binding) between the phosphatidyl serine and CD300a expressed in a myeloid cell.
[0182] Specific examples of the phosphatidyl serine-binding substance include MFG-E8 (Milk Fat Globular Protein EGF-8); T cell immunoglobulin; and soluble proteins such as soluble TIM-1, soluble TIM-4, soluble stabilin and soluble integrin .alpha.v.beta.3. Among these, MFG-E8 is preferred.
[0183] The phosphatidyl serine-binding substance is not limited to native proteins such as MFG-E8, and may be one having an amino acid sequence in which one or several amino acids are deleted, substituted and/or added (mutant) (for example, "D89E MFG-E8" in Examples) as long as the binding capacity to phosphatidyl serine is not lost.
[0184] Such a mutant can be prepared by a known method such as site-directed mutagenesis or random mutagenesis.
[0185] The "soluble protein" described above means a protein prepared by modifying a native protein, such as a membrane protein, insoluble to the later-described diluent or body fluid by, for example, deleting a hydrophobic domain or adding a hydrophilic peptide by a known genetic recombination technique such that the protein becomes soluble to the diluent or body fluid.
[0186] (CD300a-Binding Substance)
[0187] The CD300a-binding substance as a first activity modulator is not limited as long as it binds to CD300a to inhibit interaction (binding) between the CD300a expressed in a myeloid cell and phosphatidyl serine.
[0188] Specific examples of the CD300a-binding substance include neutralizing antibodies against CD300a. The neutralizing antibody may be a single particular type of monoclonal antibody, or may be a combination of 2 or more types of monoclonal antibodies (or polyclonal antibodies). Further, the neutralizing antibody may be either a full-length antibody or an antibody fragment (Fab fragment, F(ab').sub.2 fragment or the like).
[0189] The neutralizing antibody can be prepared by a known method. In cases of a monoclonal antibody, anti-CD300a monoclonal antibodies can be generally prepared by, for example, a procedure comprising immunization with CD300a, preparation of hybridomas, screening, culturing, and recovery. From the thus prepared antibodies, an appropriate monoclonal antibody that has a desired capacity (neutralizing action) to inhibit binding of CD300a to phosphatidyl serine and can exert the action and effect of the present invention may be selected.
[0190] (TX41, TX49, and Antibodies Similar to these)
[0191] TX41 is an anti-mouse CD300a monoclonal antibody (rat IgG2a), and TX49 is an anti-human CD300a monoclonal antibody (mouse IgG1). Both of these are monoclonal antibodies prepared and used in the later-described Examples, and excellent in the function to suppress signal transduction by inhibition of binding of CD300a to phosphatidyl serine. Therefore, these are preferred as the CD300a-binding substance in the present invention. However, anti-CD300a antibodies that can be used in the present invention are not limited to TX41, TX49, and antibodies similar to these (having a variable region with an equivalent amino acid sequence).
[0192] The variable region in the H-chain of TX41 has the amino acid sequence of SEQ ID NO:17; the variable region in the L-chain of TX41 has the amino acid sequence of SEQ ID NO:18; the variable region in the H-chain of TX49 has the amino acid sequence of SEQ ID NO:19; and the variable region in the L-chain of TX49 has the amino acid sequence of SEQ ID NO:20. Each of these variable regions contains 3 complementarity determining regions (CDRs) and 4 framework regions. FIG. 56 shows the results of analysis of homology between the amino acid sequences of the variable regions of TX41 and TX49 (for each of the H-chain and L-chain).
[0193] The binding substance for mouse CD300a is preferably an antibody in which the variable region in the H-chain has the amino acid sequence of SEQ ID NO:17, and the variable region in the L-chain has the amino acid sequence of SEQ ID NO:18, according to TX41.
[0194] The binding substance for human CD300a is preferably an antibody in which the variable region in the H-chain has the amino acid sequence of SEQ ID NO:19, and the variable region in the L-chain has the amino acid sequence of SEQ ID NO:20, according to TX49.
[0195] Further, the binding substance for mouse CD300a may also be an antibody in which the H-chain variable region has an amino acid sequence that is the same as the amino acid sequence of SEQ ID NO:17 except that 1, 2, 3, 4, or 5 amino acid(s) is/are substituted, added, inserted and/or deleted, or an antibody in which the L-chain variable region has an amino acid sequence that is the same as the amino acid sequence of SEQ ID NO:18 except that 1, 2, 3, 4, or 5 amino acid(s) is/are substituted, added, inserted and/or deleted (one of the H-chain and the L-chain may have the above-described mutation(s), or both of these may have the above-described mutation(s)).
[0196] Further, the binding substance for human CD300a may also be an antibody in which the H-chain variable region has an amino acid sequence that is the same as the amino acid sequence of SEQ ID NO:19 except that 1, 2, 3, 4, or 5 amino acid(s) is/are substituted, added, inserted and/or deleted, or an antibody in which the L-chain variable region has an amino acid sequence that is the same as the amino acid sequence of SEQ ID NO:20 except that 1, 2, 3, 4, or 5 amino acid(s) is/are substituted, added, inserted and/or deleted (one of the H-chain and the L-chain may have the above-described mutation(s), or both of these may have the above-described mutation(s)).
[0197] The sites of such mutations are preferably not in the CDRs or vicinities thereof in the variable regions. Further, in cases where an amino acid is substituted, the substitution is preferably conservative amino acid substitution, in which substitution occurs between amino acids having similar side-chain structures and/or chemical properties.
[0198] The form (amino acid sequence, amino acid length) of the constant region, that is, the Fab region excluding the above-described variable region, and the Fc region, of the anti-CD300a antibody may be designed as appropriate as long as the action and effect of the present invention are not inhibited, since the form of the constant region hardly affects the binding capacity to CD300a, that is, the neutralizing action.
[0199] That is, the anti-CD300a antibody can be prepared as a fusion protein composed of the above prescribed amino acid sequence of the variable region and a known amino acid sequence of the constant region.
[0200] For example, use of a human constant region for preparation of an anti-human CD300a antibody as a human chimeric antibody is one of preferred embodiments. Such an anti-CD300a antibody can be prepared by a known method.
[0201] For example, by synthesizing a DNA encoding the above prescribed amino acid sequence of the variable region and linking the synthesized DNA to a DNA encoding an amino acid sequence of the constant region and another/other necessary DNA(s) (transcription factor(s) and/or the like), an expression vector for an anti-CD300a antibody gene can be constructed. By introducing this vector to a host cell and allowing expression of the gene, the anti-CD300a antibody of interest can be produced.
[0202] The above-mentioned TX41 and TX49, and antibodies similar to these can be potentially used also for an object other than the action and effect of the present invention, i.e., the inhibition of inhibitory signal transduction that occurs due to binding of phosphatidyl serine to CD300a.
[0203] Moreover, by suppressing expression of CD300a in myeloid cells in the affected area using an siRNA designed based on a gene sequence of CD300a (available from DNA databases such as DDBREMBL/GenBank=INSD), therapeutic effects for the various diseases described above can be obtained as in the cases where the CD300a gene is deleted or binding of CD300a to phosphatidyl serine is inhibited. In other words, an siRNA against the CD300a gene can also be said to be the substance that inhibits binding of CD300a to phosphatidyl serine in the present invention.
[0204] (Use of First Activity Modulator)
[0205] The first activity modulator of the present invention can be used for suppressing inhibitory signal transduction of a CD300a-expressing myeloid cell. In this case, the myeloid cell may be either a myeloid cell present in the body, or a myeloid cell separated from the body or cultured in vitro.
[0206] By maintaining or increasing activation signaling via CD300a of the myeloid cell by the above action, intercellular signal transduction via chemical mediators released from the myeloid cell is also maintained or increased, and inflammation, allergic reaction and the like that are caused by further intercellular signal transduction that occurs thereafter can then be influenced. Therefore, the first activity modulator can be used as an effective component (for example, as an immunostimulant) of the specific medicaments described later. Further, for example, the first activity modulator can also be used as an effective component of an agent to be used as a comparative analysis tool for comparative analysis performed after amelioration of the disease state of asthma, atopic dermatitis, inflammatory bowel disease or sepsis in a laboratory animal.
[0207] Those skilled in the art can sufficiently presume that CD300a-binding substances (neutralizing antibodies such as TX41 and TX49) and phosphatidyl serine-binding substances (MFG-E8 and the like) can be therapeutic agents for diseases that have been found to show amelioration of symptoms by deletion of the CD300a gene (that is, by complete prevention of binding of CD300a to phosphatidyl serine), such as atopic dermatitis and the like described in Examples.
[0208] On the other hand, those skilled in the art can also sufficiently presume that phosphatidyl serine can be a therapeutic agent for diseases that have been found to show development or exacerbation of symptoms by deletion of the CD300a gene (that is, by complete prevention of binding of CD300a to phosphatidyl serine), such as celiac disease described in Examples.
[0209] <Second Activity Modulator>
[0210] The second activity modulator contains a component having an action to promote inhibitory signal transduction via CD300a (that is, to suppress activation signaling of CD300a). In the present invention, such a component may be a substance that promotes binding of phosphatidyl serine to CD300a. The substance is especially phosphatidyl serine, which is a ligand of CD300a.
[0211] Further, by performing screening using the CD300a gene-deficient mice provided by another aspect of the present invention, agonists (low molecular compounds, antibodies and the like) for CD300a having the same action as phosphatidyl serine may be discovered, and such agonists can also be used as substances that promote binding of phosphatidyl serine to CD300a.
[0212] (Phosphatidyl Serine)
[0213] Phosphatidyl serine (PS) is a ligand for CD300a expressed in myeloid cells, and interaction (binding) between PS and CD300a promotes inhibitory signaling of CD300a-expressing cells. For example, in mast cells, inflammatory reaction-associated activities that cause release of chemical mediators such as histamine, cytokines and chemokines are suppressed via this inhibitory signaling. PS is industrially produced, and can be easily obtained.
[0214] For CD300a-expressing myeloid cells placed in vitro (in a test environment), apoptotic cells presenting PS (it is known that PS is present inside the cell (in the cytoplasm-side layer of the lipid bilayer) in a normal cell, but presented outside the cell upon occurrence of apoptosis) can also be a second activity modulator. Further, liposomes and the like having a PS-containing lipid membrane formed in the outside can be potentially used as second activity modulators.
[0215] (Calcium Salt)
[0216] Since the interaction between PS and CD300a in mast cells requires calcium ions, the second activity modulator preferably contains a calcium salt that generates a calcium ion by ionization (e.g., calcium chloride).
[0217] The content of the calcium salt in the second activity modulator may be determined appropriately in consideration of the calcium ion concentration in the site of administration, the amount of PS contained in the second activity modulator, and the like.
[0218] (Use of Second Activity Modulator)
[0219] The second activity modulator of the present invention can be used for promoting inhibitory signal transduction of a CD300a-expressing myeloid cell. In this case, the myeloid cell may be either a myeloid cell present in the body, or a myeloid cell separated from the body or cultured in vitro.
[0220] By suppressing activation signaling via CD300a of the myeloid cell by the above action, intercellular signal transduction via chemical mediators released from the myeloid cell is also suppressed, and inflammation, allergic reaction and the like that are caused by further intercellular signal transduction that occurs thereafter can then be influenced. Therefore, the second activity modulator can be used as an effective component (for example, as an immunosuppressant) of the specific medicaments described later. Further, for example, the second activity modulator can also be used as an effective component of an agent to be used as a comparative analysis tool for comparative analysis performed after amelioration of celiac disease in a laboratory animal.
[0221] [Medicament]
[0222] The medicament (pharmaceutical composition) of the present invention contains the activity modulator as described above as an effective component (e.g., immunostimulant or immunosuppressant), and may further contain various pharmaceutically acceptable additives (e.g., a carrier), if necessary.
[0223] Such a medicament can be formulated for treatment or prophylaxis of a disease or symptom (especially inflammation reaction) in which inhibitory signal transduction of a CD300a-expressing myeloid cell is involved, such as an inflammatory infection, allergic disease or autoimmune disease.
[0224] The "treatment" includes not only curing of a disease or symptom, but also amelioration (alleviation) of a disease or symptom. The "prophylaxis" includes not only prevention of a disease or symptom in advance, but also prevention of recurrence of a disease or symptom after curing of the disease or symptom.
[0225] More specifically, by blending the first activity modulator as an effective component, a medicament or the like for treatment or prophylaxis of bacterial peritonitis or sepsis caused thereby, inflammatory bowel disease, atopic dermatitis or asthma can be prepared.
[0226] Further by blending the second activity modulator as an effective component, a medicament or the like for treatment or prophylaxis of celiac disease can be prepared.
[0227] The site of administration of the medicament is not limited, and may be a site where excessive immune function (inflammation reaction) is occurring, depending on the disease or disease state to which the medicament is applied. Examples of the site include intraperitoneal, intratracheal, subcutaneous, intradermal, and intraurogenital sites.
[0228] Since myeloid cells that express CD300a are usually present in submucosal tissues and connective tissues in mammals, the medicament is preferably directly administered to the submucosal tissue or connective tissue at the above-described site, or in the vicinity thereof The administration may be carried out by injection such as intravenous injection, intraarterial injection, subcutaneous injection, intradermal injection, intramuscular injection or intraperitoneal injection. For example, in cases of treatment or prophylaxis of an inflammatory infection (e.g., bacterial peritonitis), intraperitoneal injection is preferred.
[0229] The dose per administration and the number of doses of the medicament (or the effective component contained therein) vary depending on the age, sex and body weight of the patient; symptoms; degree of the therapeutic effect required; administration method; treatment period; type of the effective component; and the like; and may be appropriately controlled. The number of doses is, for example, 1 to several doses per day.
[0230] In cases where the medicament contains as an effective component a phosphatidyl serine-binding substance as the first activity modulator, the medicament may be formulated such that the dose per administration of the phosphatidyl serine-binding substance is usually 3 to 15 mg, preferably 5 to 10 mg per 1 kg of the human or animal subjected to the administration.
[0231] In cases where the medicament contains as an effective component a CD300a-binding substance as the first activity modulator, the medicament may be formulated such that the dose per administration of the CD300a-binding substance is usually 50 to 150 mg, preferably 50 to 100 mg per 1 kg of the human or animal subjected to the administration.
[0232] In cases where the medicament contains as an effective component a second activity modulator, the medicament may be formulated such that the dose per administration of phosphatidyl serine is usually 3 to 150 mg, preferably 5 to 100 mg per 1 kg of the human or animal subjected to the administration in view of further increasing the immunosuppressive effect.
[0233] (Pharmaceutically Acceptable Carrier)
[0234] The medicament of the present invention may contain a pharmaceutically acceptable carrier, if necessary.
[0235] The pharmaceutically acceptable carrier is not limited as long as it does not deteriorate the purpose of the medicament, and examples of the carrier include diluents such as aqueous diluents and nonaqueous diluents; stabilizers/preservatives such as antioxidants (e.g., sulfite); buffers such as phosphates; emulsifiers such as surfactants; coloring agents; thickeners; local anesthetics such as lidocaine; solubilizers such as glycols; isotonic agents such as sodium chloride and glycerin; and other additives.
[0236] For example, in cases where the dosage form of the medicament of the present invention is an injection solution, the effective component is preferably dissolved or dispersed in a diluent by blending the diluent such that a desired viscosity and desired concentrations of components are achieved.
[0237] Examples of such a diluent include aqueous diluents such as physiological saline and commercially available distilled water for injection; and nonaqueous diluents such as polyethylene glycol, and alcohols including ethanol.
[0238] The medicament whose dosage form is an injection solution may be sterilized by filtration through a filter, or may be sterilized by blending a microbicide or the like, according to a conventional method.
[0239] In cases where the activity modulator is administered as an injection solution, it may be in the form of an injection solution to be prepared at the time of use. For example, a solid dosage form containing the activity modulator may be prepared by freeze-drying or the like, and may then be dissolved or dispersed in a diluent to prepare an injection solution at the time of administration.
[0240] <Medicament for Inflammatory Infection>
[0241] Inhibition of binding of phosphatidyl serine to CD300a-expressing cells (mast cells) enables maintenance or improvement of the activity of the mast cells. This increases the number of neutrophils, and activates attack of neutrophils to pathogens (bacteria, parasites and the like), thereby improving the function to suppress the growth of pathogens, and the pathogen clearance function. Thus, by using the first activity modulator (phosphatidyl serine-binding substance or CD300a-binding substance) as an effective component (immunostimulant), a medicament for an inflammatory infection can be obtained.
[0242] <Medicament for Inflammatory Bowel Disease>
[0243] Inhibition of binding of phosphatidyl serine to CD300a-expressing cells (CD11b-positive dendritic cells in the large intestinal lamina propria) increases production, by CD4.sup.+ cells and the like, of IL-10, which is known to suppress growth induction of inflammatory T cells. This suppresses activation of inflammatory T cells, and allows maintenance of homeostasis of the gut immune system. Thus, by using the first activity modulator as an effective component, a medicament for inflammatory bowel disease can be obtained.
[0244] <Medicament for Atopic Dermatitis>
[0245] Inhibition of binding of phosphatidyl serine to CD300a-expressing cells can suppress cellular infiltration of eosinophils, mast cells (which are known to interact with skin Langerin-positive dendritic cells in the dermis to activate CD4-positive T cells) and monocytes; suppress hyperplasia of epidermis (fibroblasts); and decrease the total serum IgE level. Thus, by using the first activity modulator as an effective component, a medicament for atopic dermatitis can be obtained.
[0246] <Medicament for Asthma>
[0247] Inhibition of binding of phosphatidyl serine to CD300a-expressing cells can suppress eosinophilic airway inflammation, and decrease the total serum IgE level. Thus, by using the first activity modulator as an effective component, a medicament for asthma can be obtained.
[0248] <Medicament for Celiac Disease>
[0249] In cases where binding of phosphatidyl serine to CD300a-expressing cells (macrophages in the large intestinal lamina propria) is maintained, MyD88- and TRIF-mediated inhibitory gliadin signaling pathways play protective roles in the progression of celiac disease. Thus, by using the second activity modulator as an effective component (immunosuppressant), a medicament for celiac disease can be obtained.
[0250] [Use of CD300a Gene-Deficient Mouse]
[0251] <Uses Related to Celiac Disease>
[0252] According to a discovery obtained in the present invention, induction of celiac disease by administration of a gluten-derived gliadin peptide, which is known to be a substance that induces celiac disease, to CD300a gene-deficient mice causes more severe symptoms of celiac disease than administration to wild-type mice.
[0253] That is, a CD300a gene-deficient mouse can be used as a model mouse that develops celiac disease by administration of a substance that induces celiac disease.
[0254] The model mouse of celiac disease in the present invention is a mouse in which the CD300a gene is inactivated and release of anti-inflammatory cytokines from mast cells and macrophages is derepressed.
[0255] The mouse shows symptoms of celiac disease (inflammation of the small intestine) in cases where the mouse is fed from an early stage (from the late middle age) with a substance that induces celiac disease. Thus, the mouse is useful for elucidation of the cause of celiac disease, and plays an important role in development of a therapeutic agent for celiac disease.
[0256] Symptoms of celiac disease can be confirmed by, for example, a known method in which an inflammation-inducing substance is administered to the mouse, and a section of the small-intestinal epithelium of the mouse is then observed under the microscope.
[0257] The celiac disease model mouse of the present invention can be used also for screening of therapeutic agents for celiac disease. In particular, since the mouse securely shows symptoms of celiac disease, the mouse is useful for development of not only therapeutic agents to be applied after the onset of symptoms of celiac disease, but also prophylactic agents for celiac disease.
[0258] Further, the model mouse is also useful for evaluation of therapeutic agents for celiac disease that have already been demonstrated to be therapeutically effective.
[0259] For example, the model mouse is useful for studying the optimal concentration of a therapeutic agent that has a therapeutic effect within a certain range of concentration but does not have the effect at a concentration lower than the range and shows toxicity at a concentration higher than the range.
[0260] More specifically, the celiac disease model mice are divided into a test mouse group and a control mouse group, and a therapeutic agent to be tested is administered to the test mouse group. By comparing sections of the small-intestinal epithelium between the groups, the effect of the therapeutic agent can be evaluated.
[0261] Further, by giving the therapeutic agent to the test mouse group at different concentrations, a therapeutically effective and optimal concentration of the therapeutic agent can be studied.
[0262] That is, the CD300a gene-deficient mouse can be employed for uses in which a candidate substance for a therapeutic agent for celiac disease is administered to the CD300a gene-deficient mouse with celiac disease to see whether the agent is therapeutically effective or not; uses in which a candidate substance for a prophylactic agent for celiac disease is administered together with the substance that induces celiac disease to the CD300a gene-deficient mouse before development of celiac disease to see whether the agent is prophylactically effective or not; and uses for analyzing the pathology of celiac disease.
[0263] The CD300a (MAIR-I) receptor is positioned upstream of the signal transduction to suppress production of anti-inflammatory cytokines. Therefore, by administration of a CD300a (MAIR-I) agonist or signal transducer that maintains or promotes the signal transduction to suppress production of anti-inflammatory cytokines, symptoms of celiac disease are expected to be ameliorated.
[0264] Thus, the CD300a gene-deficient mouse can also be preferably used in screening for finding agonists; for finding signal transducers that complement signal transduction downstream of the MAIR-I receptor, and substances that induce such signal transducers; and for finding genes involved in production of such signal transducers and the like.
[0265] This screening can be carried out by differential analysis between the CD300a gene-deficient mice and WT mice by DNA microarray analysis, two-dimensional protein electrophoresis or the like.
[0266] <Uses Related to Atopic Dermatitis>
[0267] Atopic dermatitis is hypersensitivity associated with allergic reaction, and accompanied by skin inflammation (eczema or the like).
[0268] Atopic dermatitis is caused by entrance of an allergic substance (antigen) into the body followed by production of periostin due to stimulation by substances (interleukins 4 and 13) secreted from activated immunocytes and then binding of the periostin to another protein "integrin" on the surface of keratinocytes in the skin, to cause inflammation.
[0269] The binding of periostin to integrin causes production of other inflammation-inducing substances, and the symptoms continue even in the absence of the antigen, resulting in chronicity. It has been shown, by an experiment using mice, that inhibition of binding of periostin to integrin prevents occurrence of atopic dermatitis (Document 31 listed below).
[0270] According to a discovery obtained in the present invention, administration of a substance that induces atopic dermatitis (mite antigen, ovalbumin or the like) to CD300a gene-deficient mice causes milder symptoms of atopic dermatitis than administration to wild mice.
[0271] That is, the CD300a gene-deficient mouse can be used as a model mouse that hardly develops atopic dermatitis. Further, the CD300a gene-deficient mouse can be used for analysis of signaling pathways involved in atopic dermatitis, pathology analysis of atopic dermatitis, and the like in relation to IL-4 and IL-13 production.
[0272] <Uses Related to Asthma>
[0273] Bronchial Asthma is a respiratory disease in which bronchial inflammation triggered by an allergic reaction or infection with a bacterium or virus becomes chronic to cause increased airway hyperresponsiveness and reversible airway narrowing, leading to attacks of wheezing, cough, and the like. Further, bronchial asthma is said to be caused by the combination of airway hyperresponsiveness, allergic diathesis and environment.
[0274] According to a discovery obtained in the present invention, administration of a substance that induces asthma (mite antigen, ovalbumin or the like) to CD300a gene-deficient mice to induce asthma causes milder symptoms of asthma than administration to wild mice.
[0275] That is, the CD300a gene-deficient mouse can be used as a model mouse that hardly develops asthma after administration of a substance that induces asthma, and can also be used for elucidation of signaling pathways involved in asthma, pathology analysis of asthma, and the like.
[0276] <Uses Related to Inflammatory Bowel Disease>
[0277] According to a discovery obtained in the present invention, induction of inflammatory bowel disease in CD300a gene-deficient mice by a known method such as administration of an inflammation-inducing substance (e.g., DSS) causes milder symptoms of inflammatory bowel disease than in wild-type mice.
[0278] That is, the CD300a gene-deficient mouse can be used as a model mouse that hardly develops an inflammatory disease after administration of a substance that induces inflammatory bowel disease, and can also be used for pathology analysis and the like of inflammatory bowel disease in relation to the causative gene of the disease.
[0279] (Method for Preparing CD300a Gene-Deficient Mouse)
[0280] The CD300a gene-deficient mouse of the present invention is a mouse in which the CD300a gene on the chromosome is replaced by an inactive CD300a gene and hence the function of CD300a protein is deficient.
[0281] The "inactive CD300a gene" means a gene that is incapable of expressing normal CD300a protein due to, for example, partial deletion of the CD300a gene, insertion of another base sequence to the coding region of the CD300a gene, a point mutation(s) in the CD300a gene, or mutation in a regulatory region for expression of the CD300a gene. Examples of the deficient CD300a gene include, but are not limited to, genes in which at least one of the exons 1 to 6 contained in the CD300a gene is deleted.
[0282] The term "function of CD300a protein is deficient" means that at least a part of the function of CD300a protein involved in the inhibitory signal transduction related to the present invention is lost (for example, the function of CD300a protein is partially lost by replacement of one of the alleles by an inactive form, as in a heterozygous knockout mouse), preferably means that the function is completely lost.
[0283] A common method for obtaining a CD300a gene-deficient mouse using a gene cassette is as follows. However, the CD300a gene-deficient mouse in the present invention is not limited to those obtained by this method.
[0284] A targeting vector having a targeted allele (mutant allele) in which the exons of the wild-type allele of the CD300a gene are replaced by an antibiotic resistance gene (marker gene) is prepared. According to a conventional method, a chimeric mouse is obtained using the targeting vector and ES cells. The chimeric mouse is crossed with a wild-type mouse to obtain F1 mice (heterozygote (+/-)), and the F1 mice are crossed with each other to obtain F2 mice.
[0285] Genomic DNA is extracted from the F2 mice, and genomic DNA of each mouse is subjected to PCR to investigate the presence/absence of the wild-type allele and the mutant allele in the genomic DNA, to obtain an F2 mouse having only the mutant allele (homozygote (-/-)). Further, in order to confirm the absence of expression of CD300a, cells derived from the mouse are subjected to confirmation by Western blotting using an anti-CD300a antibody, to provide a CD300a gene-deficient mouse.
EXAMPLES
[0286] The present invention is described below more concretely by way of Examples.
[0287] However, the present invention is not limited by the Examples below.
1. Preparation Example
[0288] (1) Preparation of CD300a-Fc Fusion Protein, MFG-E8 and D89E MFG-E8
[0289] (1-1) A CD300a fusion protein having the Fc region of human IgG (CD300a-Fc) was prepared as described in the Document 25 listed below from a chimeric cDNA containing a cDNA of a gene encoding the whole extracellular domain of mouse or human CD300a and a cDNA of a gene encoding human IgG1Fc. The fusion protein in which the extracellular domain of CD300a is derived from mouse is referred to as "mouse CD300a-Fc", and the fusion protein in which the extracellular domain of CD300a is derived from human is referred to as "human CD300a-Fc".
[0290] (1-2) MFG-E8 was provided by Mr. Masato Tanaka (RCAI, Yokohama, Japan).
[0291] (1-3) D89E MFG-E8 is a mutant of MFG-E8 obtained by site-directed mutagenesis in the RGD motif of MFG-E8 as described in the Document 4 below.
[0292] In the obtained D89E MFG-E8, the 89th amino acid as counted from the N-terminus is substituted from aspartic acid to glutamic acid. MFG-E8 (native) binds to both phosphatidyl serine (PS) and .alpha.v.beta.3 integrin to thereby cross-link apoptotic cells to phagocytes expressing .alpha.v.beta.3 integrin (Document 4 listed below). On the other hand, D89E MFG-E8 does not bind to .alpha.v.beta.3 integrin while it binds to phosphatidyl serine (PS).
[0293] (2) Mice, and Cecal Ligation and Puncture (CLP)
[0294] The knockout mice used in the Examples were prepared or provided as follows.
[0295] (i) CD300a Gene-Deficient (Cd300a.sup.-/-) Mouse
[0296] Using a bacterial artificial chromosome (BAC) system, the exons 1 to 6 of the wild-type allele of the Cd300a gene were replaced by a neomycin resistance gene cassette (PGK-GB2-neo) to prepare a targeted allele (mutant allele) (FIG. 3A). Subsequently, a chimeric mouse was obtained according to a conventional method, and the chimeric mouse was crossed with a wild-type mouse to obtain F1 mice (heterozygote (+/-)). The F1 mice were crossed with each other to obtain F2 mice.
[0297] In order to select CD300a gene-deficient (Cd300a.sup.-/-) mice from the F2 mice, genomic DNA was extracted from the tail of each F2 mouse, and the genomic DNA was subjected to PCR to investigate the presence/absence of the WT allele and the mutant allele in the genomic DNA.
[0298] As shown in FIG. 3B, the PCR product corresponding to the WT allele and the PCR product corresponding to the mutant allele are detected as a band of about 540 bp and a band of about 700 bp, respectively.
[0299] Further, in order to confirm the absence of expression of CD300a in the CD300a gene-deficient mice, cells derived from the CD300a gene-deficient mice were subjected to Western blot analysis using an anti-CD300a antibody. As shown in FIG. 3C, a wild-type mouse showed a band of about 50 kDa derived from CD300a, but this band was not detected in a CD300a gene-deficient mouse.
[0300] (ii) C57BL/6J-Kit.sup.W-sh/W-sh Mouse
[0301] C57BL/6J-kit.sup.W-sh/W-sh mice (hereinafter referred to as "kit.sup.W-sh/W-sh mice" or "mast cell-deficient mice") were provided from RIKEN BioResource Center (Tsukuba, Japan). These mice are known to show deficiency of mast cells (Document 21 listed below), and to show incorporation of DNA by phagocytes without apoptotic DNA fragmentation, followed by degradation of DNA in the phagocytes (Document 12 listed below).
[0302] (iii) CAD-Deficient Mouse
[0303] The CAD (Caspase-activated DNase)-deficient mouse described in the Document 12 listed below was used.
[0304] (v) In Vivo Removal of Macrophages and Neutrophils
[0305] According to description in the Document 30 listed below, clodronate liposomes and control PBS liposomes (Encapsula NanoSciences) were prepared. Subsequently, at Hour 24 after CLP, 0.5 mL of the liposomes were injected to the abdominal cavity of the mouse to remove macrophages.
[0306] Further, at Hour 24 after CLP, an anti-Gr-1 antibody was injected into the abdominal cavity of the mouse to remove neutrophils.
[0307] All the operations of preparation using mice and the evaluation tests in the Examples were carried out in accordance with the guideline prepared by Animal Care and Use Committee of Laboratory Animal Resource Center, University of Tsukuba.
[0308] (3) Antibodies
[0309] The manufacturer, or the method for preparation, of each antibody is shown in the table below.
TABLE-US-00001 TABLE 1 Antibody name Manufacturer or preparation method Control rat antibody BD Pharmingen Anti-CD11b (M1/70) antibody BD Pharmingen Anti-F4/80 antibody BD Pharmingen Anti-c-Kit (2B8) antibody BD Pharmingen Anti-Gr-1 (RB6) antibody BD Pharmingen Anti-Fc.epsilon.RI (MAR-1) antibody BD Pharmingen Anti-TNF-.alpha. antibody BD Pharmingen Human CD300a monoclonal Prepared according to a Document antibody TX49 (mouse IgG1) (*) listed below Mouse CD300a monoclonal Prepared according to a Document antibody TX41 (rat IgG2a) (*) listed below Anti-MCP-1 antibody BioLegend Anti-IL-13 antibody eBiosciences Anti-SHP-1 (C-19) antibody Upstate Biotechnology Anti-SHP-2 (C-18) antibody Upstate Biotechnology (*): Preparation was carried out by the method described in the Document 1 below.
[0310] (4) Preparation of Cells
[0311] Cells were prepared as follows.
[0312] (i) Bone Marrow-Derived Mast Cells (BMMCs)
[0313] In complete RPMI 1649 medium supplemented with a cell growth factor (SCF) (10 ng/mL), IL-3 (4 ng/mL) and fetal bovine serum (FBS) (10%) placed in a 10-cm dish, 2.times.10.sup.8 mouse bone marrow cells were cultured for not less than 5 weeks to prepare bone marrow-derived mast cells (BMMCs). The BMMCs were subcultured every week with fresh medium. Flow cytometry analysis showed that more than 95% of the prepared BMMCs were c-Kit.sup.+Fc.epsilon.RI.sup.+ cells.
[0314] (ii) Bone Marrow-derived Macrophages (BMM.phi.)
[0315] In complete RPMI 1649 medium supplemented with M-CSF (10 ng/mL) and fetal bovine serum (FBS) (10%) placed in a 10-cm dish, 2.times.10.sup.6 mouse bone marrow cells were cultured for 1 week to prepare bone marrow-derived macrophages (BMM.phi.).
[0316] (iii) NIH3T3 Cell Transfectant
[0317] According to a conventional method, a pMX-neo retrovirus vector plasmid containing a cDNA of Flag-tagged CD300a or cDNA of Flag-tagged CD300d was prepared.
[0318] NIH3T3 cells were transfected with the thus prepared plasmid, to obtain a transfectant that stably expresses CD300a or CD300d. The transfectant that stably expresses CD300a and the transfectant that stably expresses CD300d obtained are referred to as "NIH3T3 transfectant (CD300a)" and "NIH3T3 transfectant (CD300d)", respectively.
[0319] The NIH3T3 cell transfectant that stably expresses TIM-4 was provided by Mr. T. Kitamura (University of Tokyo). This transfectant is referred to as "NIH3T3 transfectant (TIM-4)".
[0320] [Method of Evaluation]
[0321] Conditions for the method of evaluation are described below. In the survival test, the Kaplan-Meier log-rank test was used, and, in other evaluation tests, the unpaired Student's t test was used to perform statistical analysis. Statistical significance was assumed at P<0.05.
[0322] (5) Binding Assay Etc.
[0323] (i) Binding Assay
[0324] Cells were stained for 30 minutes in phosphate buffer supplemented with 2% FBS in the presence or absence of 1 mM CaCl.sub.2 using CD300a or a control human IgG, and then washed twice with the same buffer, followed by incubation with the F(ab').sub.2 fragment of an FITC-conjugated goat anti-human IgG. Subsequently, for staining with annexin V, the cells were incubated in 10 mM HEPES-NaOH buffer supplemented with 140 mM NaCl and 2.5 mM CaCl.sub.2 together with annexin V for 15 minutes at room temperature.
[0325] (ii) Binding Inhibition Assay
[0326] Cells were preincubated with a monoclonal antibody against mouse CD300a (TX41), control isotype antibody or MFG-E8 for 30 minutes, and then stained with CD300a-Fc as in the above binding assay. Further, in order to analyze whether CD300a-Fc was bound to phospholipid or not, an assay was carried out using a PIP Strip (manufactured by Echelon Biosciences) according to the manufacturer's instructions.
[0327] (6) Measurement of CFU (Colony Forming Unit) of Aerobic Bacteria
[0328] Serial dilutions of mouse peritoneal perfusate were plated, and the dilutions were cultured on plates containing brain-heart infusion (BHI) agar at 37.degree. C. for 48 hours. Subsequently, the CFU of aerobic bacteria was calculated by measuring the number of colonies in 1 mL of the peritoneal perfusate as described in the Document 27 listed below.
Example 1; Identification of CD300a Ligand
Example 1A
[0329] In order to confirm expression of the mouse CD300a ligand in hematopoietic stem cell lines and tumor cell lines, the following test was carried out.
[0330] The bone marrow-derived macrophages (BMM.phi.) obtained in "1. Preparation Example", bone marrow-derived dendritic cells (BMDCs) or IL-3-dependent hematopoietic cell line cells (BaF/3 cells) (2.times.10.sup.5 cells per each type of cells) were incubated in PBS (phosphate buffered saline) containing CD300a-Fc (1 .mu.g) and calcium chloride (1 mM) at 20.degree. C. for 30 minutes, and then stained using a buffer containing an FITC-conjugated anti-human IgG antibody (0.1 .mu.g) and propidium iodide (PI) (1 .mu.g).
[0331] The stained cells of each type were subjected to analysis using a flow cytometer (FACSCalibur, manufactured by Becton Dickinson; model number, "E6133").
[0332] Further, a control test was carried out in the same manner as in the above test method except that a control human IgG (1 .mu.g) was used instead of mouse CD300a-Fc.
[0333] The results of flow cytometry analysis for BMM.phi., BMDC and BaF/3 cells are shown in FIG. 1A, FIG. 1B and FIG. 1C, respectively (the results of the control test are shown as "Ctrl Ig").
[0334] As shown in FIG. 1A to FIG. 1C, it was found that, in cases where calcium chloride is contained, mouse CD300a-Fc binds to PI.sup.- cells (live cells) but does not bind to PI.sup.+ cells (dead cells). That is, the mouse CD300a ligand is suggested to be expressed in dead cells.
Example 1B
[0335] In order to test whether mouse CD300a-Fc binds to apoptotic cells, which are a type of dead cells, the following test was carried out.
[0336] Thymocytes derived from a C57BL/6 mouse (wild-type mouse) were incubated with dexamethasone (manufactured by Sigma) (10 .mu.M) in RPMI medium to prepare apoptotic thymocytes.
[0337] The obtained apoptotic cells (cell number, 2.times.10.sup.5) were incubated in a medium (PBS) containing CD300a-Fc (1 .mu.g), APC-conjugated annexin V (manufactured by BD Pharmingen) (1 .mu.l) and calcium chloride (1 mM) at 20.degree. C. for 30 minutes, and stained using a buffer containing an FITC-conjugated anti-human IgG antibody (0.1 .mu.g) and propidium iodide (PI) (1 .mu.g).
[0338] The stained cells of each type were subjected to analysis using a flow cytometer (FACSCalibur, manufactured by Becton Dickinson; model number, "E6133") (results: FIG. 2A).
[0339] Further, the cells were subjected to flow cytometry analysis under the same conditions as in the above test except that a medium supplemented with no calcium chloride was used instead of the medium supplemented with calcium chloride (results: FIG. 2B).
[0340] According to FIG. 2A, it can be seen that, in the presence of calcium chloride, mouse CD300a-Fc bound to thymocytes that are stained with annexin V but did not bind to thymocytes that are not stained with annexin V (annexin V). That is, it is suggested that mouse CD300a-Fc binds to apoptotic thymocytes.
[0341] On the other hand, it can be seen as shown in FIG. 2B that mouse CD300a-Fc did not bind to apoptotic thymocytes in the absence of calcium chloride.
[0342] From these test results, it can be understood that mouse CD300a binds to apoptotic cells dependently on calcium ions.
Example 1C
[0343] The apoptotic cells obtained in Example 1B (cell number, 2.times.10.sup.5) were incubated in a medium (PBS) supplemented with CD300a-Fc (1 .mu.g), APC-conjugated annexin V (manufactured by BD Pharmingen) (1 .mu.l), control human IgG1 (1 .mu.g) and calcium chloride (1 mM) at 20.degree. C. for 30 minutes, and stained using a buffer containing an FITC-conjugated anti-human IgG antibody (0.1 .mu.g) and propidium iodide (PI) (1 .mu.g).
[0344] The stained cells of each type were subjected to analysis using a flow cytometer (FACSCalibur, manufactured by Becton Dickinson; model number, "E6133") (results: FIG. 2C "HuIgGl").
[0345] Further, the cells were subjected to flow cytometry analysis under the same conditions as in the above test method except that "TX41", which is a monoclonal antibody, or "MFG-E8", which is a protein expressed in the macrophage, was used instead of the control human IgG1 (results: FIG. 2C "TX41" and "MFG-E8").
[0346] As described above, the "TX41" used herein is an anti-mouse CD300a monoclonal antibody, and blocks binding of mouse CD300a to the ligand.
[0347] "MFG-E8" is known to bind to both phosphatidyl serine (PS) and .alpha.v.beta.3 integrin to thereby cross-link apoptotic cells to phagocytes expressing .alpha.v.beta.3 integrin (Document 4 listed below).
[0348] As can be seen by comparison among the results of flow cytometry shown in FIG. 2C, mouse CD300a did not bind to apoptotic cells in the presence of TX41 or MFG-E8.
[0349] From this viewpoint, it is suggested that mouse CD300a-Fc has binding capacity to PS (that is, the ligand of CD300a is PS).
Example 1D
[0350] In order to test whether or not human CD300a binds to apoptotic cells similarly to mouse CD300a, the following test was carried out.
[0351] First, Jurkat cells (human T-cell line) were suspended in RPMI 1640 medium, and the resulting medium was irradiated with UV for 60 minutes to prepare apoptotic Jurkat cells.
[0352] Flow cytometry analysis was carried out under the same test conditions as in Example 1A except that the "Jurkat cell-derived apoptotic cells" were used as apoptotic cells instead of the apoptotic cells derived from wild-type mouse thymocytes, and "human CD300a-Fc" was used instead of the "mouse CD300a-Fc" (results: FIG. 2D).
[0353] Further, cytometry analysis was carried out under the same test conditions as in Example 1B except that the "Jurkat cell-derived apoptotic cells" were used as apoptotic cells instead of the apoptotic cells derived from wild-type mouse thymocytes; "human CD300a-Fc" was used instead of the "mouse CD300a-Fc"; and "TX49" or "control human IgG1" was used instead of "TX41" (results: FIG. 2E "TX49" or "HuIgGl"). "TX49" is an anti-human CD300a antibody (monoclonal antibody), and blocks binding of CD300a to the ligand.
[0354] As can be seen from FIG. 2D to FIG. 2E, human CD300a-Fc bound to annexin V.sup.+ cells but did not bind thereto in the presence of the anti-human CD300a antibody.
[0355] That is, it is suggested that, similarly to mouse CD300a-Fc, human CD300a also binds to apoptotic cells.
Example 1E
[0356] Liquids (test liquids) containing various phospholipids (PS, PC, PE) (100 pmol) were spotted on a membrane (PIP-strip (manufactured by Echelon Bioscience)) to allow adsorption of the phospholipids on the membrane.
[0357] Subsequently, the membrane was immersed in TBST buffer (pH 8.0) containing mouse CD300a-Fc (1.5 .mu.g/mL), supplemented with calcium chloride (1 mM) and BSA, at 20.degree. C. for 2 hours.
[0358] Thereafter, the membrane was washed 3 times with TBST buffer that does not contain mouse CD300a-Fc (pH 8.0) to remove CD300a-Fc unbound to the phospholipids on the membrane. Detection was then carried out using TBST buffer (pH 8.0) containing HRP-conjugated anti-human IgG (manufactured by Jackson Immun), supplemented with BSA (results: FIG. 2F).
[0359] In FIG. 2F, "PE", "PC" and "PS" indicate the positions where phosphatidyl ethanolamine, phosphatidyl choline and phosphatidyl serine were spotted on the membrane, respectively, and "Blank" indicates a position where no phospholipid was spotted on the membrane.
[0360] According to FIG. 2F, CD300a bound to neither PE nor PC, and bound specifically to PS.
[0361] From the results of Examples 1A to 1E, it can be understood that CD300a binds to phosphatidyl serine (PS) dependently on calcium ions (that is, the ligand of CD300a is PS).
Example 2: Functional Analysis of CD300a (1)
[0362] Some PS-binding receptors are known to be expressed in phagocytes and to be involved in removal of apoptotic cells under physiological and pathological conditions (Documents 4 to 9 listed below).
[0363] PS is known to mediate the so-called "eat me" signal in phagocytes (macrophages and the like), which are cells expressing CD300a (Documents 10 and 11 listed below). In view of this, the tests described in the Examples 2A to 2C below were carried out to test whether CD300a is involved in phagocytosis of apoptotic cells or not.
Example 2A
[0364] Thymocytes derived from a CAD-deficient mouse were treated in the same manner as in Example 1B to prepare apoptotic thymocytes.
[0365] Subsequently, macrophages (thioglycolate-induced peritoneal macrophages) derived from a CD300a gene-deficient mouse (2.times.10.sup.5 cells) were co-cultured with the apoptotic thymocytes derived from a CAD-deficient mouse at a ratio of 1:5 (macrophages:apoptotic thymocytes (cell numbers)) in a 8-well Lab-TeK II chamber slide (manufactured by Nalge Nunc) at 37.degree. C. for 1 hour.
[0366] Subsequently, as described in the Documents 5 and 26 listed below, the co-cultured macrophages were washed with cold PBS and fixed with a fixative containing paraformaldehyde (1%). The fixed macrophages where then subjected to TUNEL staining using a buffer containing FITC-labeled dUTP (manufactured by Roche).
[0367] Not less than 50 cells randomly selected from the stained macrophages were analyzed using a laser scanning confocal microscope ("FV10i", manufactured by Olympus Corporation; product number, 1B22358), and the number of TUNEL-positive cells (apoptotic cells) contained per one macrophage cell was counted. The ratios of macrophages containing apoptotic cells in the numbers of 0 to 8 (phagocytosis rates) were calculated as percentages with respect to the total number of macrophages (results: FIG. 4A "Cd300a.sup.-/-").
[0368] Further, the phagocytosis rates were measured under the same conditions as in the above test except that macrophages derived from a wild-type mouse were used instead of the macrophages derived from a CD300a gene-deficient mouse (results: FIG. 4A "WT").
[0369] As shown in FIG. 4A, no evident difference in the phagocytosis rate was found between the case where the macrophages were derived from a CD300a gene-deficient mouse and the case where the macrophages were derived from a wild-type mouse.
Example 2B
[0370] In order to test whether mast cells express known PS receptors (TIM-1, TIM-4, stabilin 2 and integrin .alpha.v.beta.3), the following test was carried out.
[0371] Bone marrow-derived mast cells (BMMCs) (cell number, 2.times.10.sup.5) were incubated at 20.degree. C. for 30 minutes in a medium (PBS) containing a PE (Phycoerythrin)-conjugated TIM-1 monoclonal antibody (0.1 .mu.g), APC-conjugated TIM-4 monoclonal antibody (0.1 .mu.g) and Alexa-conjugated anti-mouse CD300a monoclonal antibody (TX41) (0.5 .mu.g).
[0372] Subsequently, the stained cells were washed twice using PBS, and subjected to analysis using a flow cytometer (FACSCalibur, manufactured by Becton Dickinson; model number, "E6133") (results: FIG. 5A "BMMCs").
[0373] Further, flow cytometry analysis was carried out under the same conditions as in the above test method except that peritoneal macrophages or BM-derived macrophages were used instead of the BMMCs (results: FIG. 5A "Peritoneal macrophages" and "BM derived macrophages").
[0374] Further, using High Capacity cDNA Reverse Transcription Kit (manufactured by Applied Biosystems), cDNAs were prepared from peritoneal macrophages and BMMCs. Using each prepared cDNA, the expression levels of stabilin 2, BA1-1, .alpha.v integrin, Cd300a and .beta.-actin (loading control) were analyzed by RT-PCR (results: FIG. 5B).
[0375] As can be seen from FIG. 5A and FIG. 5B, unlike the cases of macrophages, the mast cells expressed CD300a and .alpha.v.beta.3 integrin, but showed only low levels of expression of TIM-1, TIM-4 and stabilin 2, which are PS receptors involved in phagocytosis.
Example 2C
[0376] The NIH3T3 transfectant (CD300a) (cell number, 6.times.10.sup.4) was co-cultured with FITC-labeled cells (apoptotic thymocytes or thymocytes (live cells)) for 2 hours, and washed with PBS, followed by analysis under a light microscope (BZ-9000, manufactured by Keyence) (results: FIG. 4B).
[0377] Further, the cells obtained after the co-culture and washing were fixed with a fixative containing paraformaldehyde, Vectashield (manufactured by Vector Laboratories), and analyzed using a laser scanning confocal microscope (results: FIG. 4C). In FIG. 4C, green areas (indicated by arrows) indicate phagocytosed cells (apoptotic thymocytes or thymocytes (live cells)).
[0378] Further, analysis using a light microscope and a laser scanning confocal microscope was carried out under the same conditions as in the above test except that NIH3T3 untransfected cells (negative control) or "NIH3T3 transfectant (TIM-4)" cells (positive control) were used instead of the NIH3T3 transfectant (CD300a).
[0379] In FIG. 4B and FIG. 4C, "NIH-3T3" and "NIH-3T3/Tim4" show images of "NIH3T3" (untransfected cells) and "NIH3T3 transfectant (TIM-4)", respectively, which images were obtained using a light microscope and a laser scanning confocal microscope.
[0380] The images obtained with the laser scanning confocal microscope were used to measure the ratios of the number of untransfected cells and the number of cells of each transfectant that incorporated apoptotic thymocytes into the cytoplasm (percentages with respect to the number of co-cultured untransfected cells or to the number of co-cultured cells of each transfectant) (results: FIG. 4D "apoptotic").
[0381] Further, similarly, the ratios of the number of NIH3T3 cells and the number of cells of each transfectant that incorporated thymocytes (live cells) into the cytoplasm (percentages with respect to the number of co-cultured untransfected cells or to the number of co-cultured cells of each transfectant) were measured (results: FIG. 4D "Live").
[0382] As shown in FIG. 4B, unlike NIH3T3, both of the above transfectants adhered to apoptotic thymocytes. However, based on FIG. 4C and FIG. 4D, it can be seen that only the NIH3T3 transfectant (TIM-4) incorporated apoptotic thymocytes to show phagocytosis.
[0383] Although data are not shown, the NIH3T3 transfectant (CD300a) did not show phagocytosis of live cells (thymocytes), similarly to NIH3T3.
[0384] From the results of Examples 2A to 2C, it can be understood that CD300a is not involved in phagocytosis of apoptotic cells by macrophages.
Example 3: Functional Analysis of CD300a (2)
[0385] As shown in FIG. 5, mast cells express CD300a, but, unlike macrophages, the cells do not express TIM-1, TIM-4 and stabilin, which are PS receptors.
[0386] PS is known to bind to these PS receptors directly or indirectly, and contribution of these receptors to incorporation of apoptotic cells is known (Document 13 listed below). In view of this, the tests described below in the Examples 3A to 3C were carried out in order to test whether CD300a also has such a function or not (whether or not there is functional overlap in incorporation of apoptotic cells).
Example 3A
[0387] In complete RPMI 1649 medium supplemented with a cell growth factor (SCF) (10 ng/mL), IL-3 (4 ng/mL) and fetal bovine serum (FBS) (10%) placed in a 10-cm dish, 2.times.10.sup.8 bone marrow cells (BM cells) derived from a CD300a gene-deficient mouse were cultured for 4 weeks to prepare bone marrow-derived mast cells (BMMCs) of the CD300a gene-deficient mouse. The BMMCs were subcultured every week with fresh medium.
[0388] Subsequently, the obtained BMMCs were incubated in RPMI1649 medium containing an FITC-conjugated anti-Fc.epsilon.RI.alpha. antibody (0.1 .mu.g) and a PE-conjugated anti-c-Kit antibody (0.1 .mu.g/mL) at 4.degree. C. for 30 minutes, and analyzed by flow cytometry (results: FIG. 6A "CD300.sup.-/-").
[0389] Further, flow cytometry analysis was carried out under the same test conditions as in the above test except that BMMCs were prepared using bone marrow cells derived from a wild-type mouse instead of the bone marrow cells derived from a CD300a gene-deficient mouse (results: FIG. 6A "WT"). Each number in FIG. 6A indicates the ratio of the cells shown in each box. Each test was repeated 3 times independently.
[0390] Using each type of BMMCs, a .beta.-hexosaminidase release assay (degranulation assay) was carried out as follows (for detailed conditions, see the Document 29 below).
[0391] First, 1.times.10.sup.5 to 2.times.10.sup.5 BMMCs of each type in the logarithmic growth phase were cultured at 37.degree. C. for one day and night in a 24-well plate coated with gelatin (manufactured by Sigma), and then incubated at 37.degree. C. for 1 hour in a medium that contains a biotin-conjugated mouse anti-trinitrophenol IgE (0.5 mg/mL) but does not contain a supplement.
[0392] Subsequently, streptavidin was added to the medium to cause cross-linking between the biotin-conjugated mouse anti-trinitrophenol IgE molecules, and culture was performed at 37.degree. C. for 45 minutes, followed by collecting the supernatant.
[0393] To the collected supernatant, a buffer (pH 4.5) containing p-nitrophenyl-N-acetyl-.beta.-D-glucosamide (manufactured by Sigma), citric acid (0.4 M) and sodium phosphate (0.2 M) was added, and the resulting mixture was incubated at 37.degree. C. for 3 hours to allow hydrolysis reaction of p-nitrophenyl-N-acetyl-.beta.-D-glucosamide by released .beta.-hexosaminidase. This reaction was stopped by adding 0.2 M glycine-NaOH (pH 10.7), and the absorbance at a wavelength of 415 nm, which increases as hydrolysis of p-nitrophenyl-N-acetyl-.beta.-D-glucosamide proceeds, was measured to quantify the amount of .beta.-hexosaminidase released. The rate (%) of increase in the amount of .beta.-hexosaminidase released with respect to the amount observed with untreated BMMCs of each type is shown in FIG. 6B.
[0394] FIG. 6B shows the ratio of BMMCs that released .beta.-hexosaminidase. As shown in FIG. 6A and FIG. 6B, no significant difference was found in expression of Fc.epsilon.RI.alpha. and c-Kit (marker proteins for mast cells) and the rate of increase in the amount of .beta.-hexosaminidase released between the case where the BMMCs were derived from a CD300a gene-deficient mouse and the case where the BMMCs were derived from a wild-type mouse.
[0395] That is, it can be seen that differentiation from bone marrow cells and degranulation mediated by FceRI are not influenced by CD300a.
Example 3B
[0396] BMMCs derived from a CD300a gene-deficient mouse and the apoptotic cells obtained in Example 1B (BMMCs:apoptotic cells=10:1 (ratio in terms of the cell number)) were incubated in PBS containing calcium chloride (1 mM), APC-conjugated annexin V (1 .mu.l), CD300a-Fc (1 .mu.g/mL) and MFG-E8 (5 .mu.g) at 20.degree. C. for 30 minutes, and then stained using a buffer containing an FITC-conjugated anti-human IgG antibody (0.1 .mu.g/mL) and propidium iodide (PI) (1 .mu.g).
[0397] The stained cells were subjected to analysis using a flow cytometer (FACSCalibur, manufactured by Becton Dickinson; model number, "E6133") (results: FIG. 7A "MFG-E8").
[0398] Further, flow cytometry analysis was carried out under the same conditions as in the above test except that a control IgG was used instead of MFG-E8 (results: FIG. 7A "Ctrl Ig").
[0399] From the results shown in FIG. 7A, it can be seen that CD300a-Fc bound to apoptotic cells (annexin V.sup.+) in the presence of the control IgG, but that the binding was specifically inhibited in the presence of MFG-E8 (PS-binding substance).
[0400] The concentrations of cytokines and chemokines in the supernatant of this sample mixture were quantified using ELISA kits manufactured by BD Pharmingen (TNF-.alpha. and IL-6) and R&D Systems (MIP-2, MCP-1, IL-13 and MIP-1a). As a result, none of the cytokines and chemokines could be detected.
Example 3C
[0401] In order to test whether or not stimulation by LPS (lipopolysaccharide) changes the amounts of cytokines released in the coexistence of BMMCs and apoptotic cells, the following test was carried out.
[0402] BMMCs derived from a CD300a gene-deficient mouse and apoptotic cells (BMMCs:apoptotic cells=10:1 (ratio in terms of the cell number)) were incubated in RPMI containing LPS (1 .mu.g/mL) for 4 hours, and the supernatant of the medium was then collected.
[0403] Subsequently, the levels of cytokines and chemokines were measured 3 times using ELISA kits manufactured by BD Pharmingen (TNF-.alpha. and IL-6) and R&D Systems (MIP-2, MCP-1, IL-13 and MIP-1.alpha.), and the rate of increase in the amount of each cytokine or chemokine released with respect to the amount observed with the BMMCs that had not been subjected to the above LPS treatment was calculated (results: FIG. 7B "Cd300a.sup.-/-").
[0404] Further, the rate of increase in the amount of each cytokine or chemokine was calculated under the same conditions as in the above test except that BMMCs derived from a wild-type mouse were used instead of the BMMCs derived from a CD300a gene-deficient mouse (results: FIG. 7B "WT").
[0405] As shown in FIG. 7B, LPS increased the amounts of cytokines released in both types of BMMCs. However, the BMMCs derived from a CD300a gene-deficient mouse showed significantly larger increases in the amounts of TNF-.alpha., IL-13 and MCP-1 than the BMMCs derived from a wild-type mouse.
Example 3D
[0406] Further, the following test was carried out in order to test the rates of increase in intracellular cytokines and chemokines in BMMCs.
[0407] BMMCs derived from a CD300a gene-deficient mouse and the apoptotic cells obtained in Example 1B (BMMCs:apoptotic cells=10:1 (ratio in terms of the cell number)) were incubated in a medium (RPMI) containing LPS (lipopolysaccharide) (1 .mu.g/mL) for 4 hours, and the BMMCs and apoptotic cells after the incubation were then incubated in a medium (FIX & PERM, manufactured by Invitrogen) supplemented with fluorescently labeled antibodies against various cytokines and chemokines and formaldehyde at 4.degree. C. for 20 minutes. The stained cells were subjected to analysis using a flow cytometer (FACSCalibur, manufactured by Becton Dickinson; model number, "E6133") (results: FIG. 8, arrows (1)). Further, flow cytometry analysis was carried out under the same conditions as in the above test except that a control antibody was used instead of the fluorescently labeled antibodies against various cytokines and chemokines (control test (results: FIG. 8, arrows (2))).
[0408] Further, flow cytometry analysis was carried out under the same conditions as in the above test except that BMMCs derived from a wild-type mouse were used instead of the BMMCs derived from a CD300a gene-deficient mouse (results: FIG. 8, arrows (3)). Further, flow cytometry analysis was carried out under the same conditions as in the above test except that a control antibody was used instead of the fluorescently labeled antibodies against various cytokines and chemokines (control test (results: FIG. 8, arrows (4))).
[0409] Each graph in FIG. 8 shows the amount of increase in MFI (mean fluorescence intensity) observed for each cytokine or chemokine in each type of BMMCs, relative to that of LPS-untreated BMMCs.
[0410] According to FIG. 8, in both types of BMMCs, the amounts of cytokines and chemokines in the cytoplasm increased compared to the case where the LPS treatment was not carried out. In particular, it can be seen that the BMMCs derived from a CD300a gene-deficient mouse showed significantly larger increases in the amounts of TNF-.alpha. and the like in the cytoplasm than the BMMCs derived from a wild-type mouse.
Example 3E
[0411] D89E MFG-E8 is a variant (mutant) of MFG-E8 and has a point mutation (D89E) in the RGD motif. D89E MFG-E8 binds to PS, but does not bind to .alpha.v.beta.3 integrin.
[0412] In view of this, the following test was carried out in order to test whether or not the amounts of cytokines and chemokines released from BMMCs change in the presence of D89E MFG-E8.
[0413] TNF-.alpha., IL-13, MCP-1 and IL-6 were quantified in the same manner as in Example 3C except that LPS as well as D89E MFG-E8 (5 .mu.g/mL) were added to the medium containing BMMCs derived from a CD300a gene-deficient mouse and apoptotic cells (results: FIG. 7C "CD300a.sup.-/-").
[0414] Further, TNF-.alpha., IL-13, MCP-1 and IL-6 were quantified under the same conditions as in the above test except that BMMCs derived from a wild-type mouse were used instead of the BMMCs derived from a CD300a gene-deficient mouse (results: FIG. 7C "WT").
[0415] As shown in FIG. 7C, in the presence of D89E MFG-E8, no significant difference was found in the concentrations of cytokines between the case where the BMMCs were derived from a CD300a gene-deficient mouse and the case where the BMMCs were derived from a wild-type mouse.
Example 3F
[0416] CD300a is known to have an immunoreceptor tyrosine-based inhibitory motif (ITIM) in the intracellular domain, and to induce SHP-1 upon cross-linking by an anti-CD300a antibody (Document 14 listed below).
[0417] In view of this, the following test was carried out in order to test whether CD300a interacts with SHP-1 or not. As in Example 3C, BMMCs derived from a CD300a gene-deficient mouse or wild-type mouse were co-cultured with apoptotic cells in the presence of LPS for 4 hours, and a homogenate of the cells was subjected to immunoprecipitation with an anti-CD300a antibody (TX41).
[0418] Using the thus obtained immunoprecipitates, immunoblotting with an anti-SHP-1 antibody or an anti-CD300a antibody was carried out as described in Document 14 listed below (results: FIG. 7D).
[0419] As can be seen from these results, the BMMCs responded to the stimulation with LPS to induce (recruit) SHP-1 when they were co-cultured with apoptotic cells. However, CD300a did not recruit SHP-1 in the presence of D89E MFG-E8.
[0420] That is, it is thought that induction (recruitment) of SHP-1 by CD300a in response to LPS stimulation requires binding of PS to CD300a.
Example 3G
[0421] In order to investigate whether SHP-1 is involved in CD300a-mediated signaling or not, first, Ptpn6 (SHP-1 gene) of BMMCs derived from a wild-type mouse was knocked out with an siRNA to prepare SHP-1-deficient (Ptpn6-KD) wild-type mouse-derived BMMCs under the following conditions. Further, similarly, SHP-1-deficient (Ptpn6-KD) CD300a gene-deficient mouse-derived BMMCs were prepared from BMMCs derived from a CD300a gene-deficient mouse.
[0422] With 1 mL of X-treme Gene siRNA transfection reagent (manufactured by Roche), 0.5 mM siRNA (SHP-1 siRNA) (siGENOME SMARTpool; ThermoScientific Dharmacom) targeting the SHP-1 gene (Ptpn6 gene) in BMMCs was mixed, and 5.times.10.sup.5 BMMCs derived from a CD300a gene-deficient mouse were transfected therewith as described in the Document 28 listed below, to prepare SHP-1 knockdown BMMCs derived from a CD300a gene-deficient mouse (BMMCs (CD300a.sup.-/- Ptpn6-KD)).
[0423] Further, SHP-1 knockdown BMMCs derived from a wild-type mouse (BMMCs (WT Ptpn6-KD)) were prepared under the same conditions as in the above test except that BMMCs derived from a wild-type mouse were used instead of the BMMCs derived from a CD300a gene-deficient mouse.
[0424] Here, in order to confirm that the BMMCs of each type were transfected with the SHP-1 siRNA and that the expression level of SHP-1 was decreased, a lysate of BMMCs (CD300a.sup.-/- Ptpn6-KD) or BMMCs (WT Ptpn6-KD) was subjected to immunoblotting analysis using an anti-SHP-1 antibody, anti-SHP-2 antibody or anti-.beta.-actin antibody (results: FIG. 7E).
[0425] As can be seen from FIG. 7E, the BMMCs transfected with the SHP-1 siRNA showed a decreased expression level of SHP-1. "Ctrl" shows results of immunoblotting analysis using BMMCs transfected with a control siRNA instead of the SHP-1 siRNA.
[0426] Subsequently, BMMCs (CD300a.sup.-/- Ptpn6-KD) or BMMCs (WT Ptpn6-KD), and the apoptotic cells obtained in Example 1B (BMMCs:apoptotic cells=10:1 (ratio in terms of the cell number)) were incubated in RPMI supplemented with calcium chloride (1 mM) and LPS (lipopolysaccharide) (1 .mu.g/mL) for 4 hours, and the amount of TNF-.alpha. released was measured in the same manner as in Example 3B (results: FIG. 7F).
[0427] As shown in FIG. 7F (left graph), the BMMCs derived from a CD300a gene-deficient mouse produced a significantly larger amount of TNF-.alpha. than the BMMCs derived from a wild-type mouse. On the other hand, as shown in FIG. 7F (right graph), in the cases where the BMMCs derived from a wild-type mouse or a CD300a gene-deficient mouse were transfected with the SHP-1 siRNA, the amount of TNF-.alpha. released was almost the same between the BMMCs derived from a wild-type mouse and the BMMCs derived from a CD300a gene-deficient mouse, and no significant difference was found between these.
[0428] These results suggest that binding of PS to CD300a causes CD300a to induce SHP-1 and to thereby mediate signaling that causes suppression of the activity of BMMCs, resulting in suppression of secretion of TNF-.alpha..
[0429] From the results of Example 3, it can be understood that the interaction between PS and CD300a inhibits production of inflammation-inducing (LPS-inducing) cytokines and chemokines from BMMCs, and that the interaction recruits SHP-1, resulting in suppression of secretion of TNF-.alpha..
Example 4: Functional Analysis of CD300a (3)
[0430] TNF-.alpha., IL-3 and MCP-1 produced by mast cells are chemoattractants for neutrophils, and known to play important roles in bacterial clearance in CLP peritonitis mice (Patent Documents 15 to 19 listed below).
[0431] In view of this, in order to study whether CD300a has a bacterial clearance function or not, the Examples 4A to 4H below were carried out.
Example 4A
[0432] A wild-type mouse was subjected to midline incision of 1 to 2 cm on the cecum (ventral region), and the end portion was ligated. After performing two times of puncture using a 27-gauge needle in the ligated area, the cecum was returned to the abdomen. Thereafter, 1 mL of sterile physiological saline was subcutaneously injected for rehydration, and the incision site was closed by suturing. Details of the procedure and conditions for the CLP are described in the Document 16 listed below.
[0433] Before performing the CLP and 4 hours after performing the CLP, peritoneal perfusate was collected. Subsequently, APC-conjugated annexin V (1 .mu.g) and CD300-Fc (1 .mu.g) were added to the peritoneal perfusate, and staining was performed with an FITC-conjugated anti-human IgG and PI (propidium iodide), followed by performing analysis by flow cytometry (results: FIG. 12A).
[0434] As can be seen from the results shown in FIG. 12A, the site of peritonitis was a site where a number of cells were undergoing apoptosis, as described in the Document 20 listed below.
[0435] That is, the immune regulation by mast cells in the site of peritonitis is suggested to be influenced by CD300a.
Example 4B
[0436] In order to test the relationship between CD300a and the immune regulation by mast cells, proteome analysis was carried out as follows.
[0437] First, CLP was carried out in the same manner as in Example 4A using a wild-type mouse and a mast cell-deficient mouse (kit.sup.W-sh/W-sh).
[0438] Four hours after performing the CLP, peritoneal perfusate was collected from each mouse, and the collected peritoneal perfusate was subjected to proteome analysis of cytokines and chemokines using Proteome Profiler Array (manufactured by R&D Systems) according to the manufacturer's instructions.
[0439] FIG. 9A shows the results of densitometry analysis (proteome analysis) using the peritoneal perfusate from each of the wild-type mouse and the mast cell-deficient mouse (kit.sup.W-sh/W-sh mouse) (in FIG. 9A, "PC" indicates a positive control).
[0440] FIG. 9B shows the pixel densities of the signals for the chemokines and cytokines, which were obtained from the densitometry images shown in FIG. 9A.
[0441] As can be seen from the results shown in FIG. 9B, at Hour 4 after the CLP, the concentrations of chemokines were higher in the kit.sup.W-sh/W-sh mouse than in the wild-type mouse. Similar results were obtained in 2 replicates of the test.
Example 4C
[0442] In the same manner as in Example 4B, peritoneal perfusate was collected from wild-type mice and mast cell-deficient mice (kit.sup.W-sh/W-sh mice) (n=3 per each type of mice).
[0443] Subsequently, a dilution series of each peritoneal perfusate was prepared, and the prepared serial dilutions of peritoneal perfusate were plated to perform culture on plates containing brain-heart infusion (BHI) agar at 37.degree. C. for 48 hours. Thereafter, the CFU of aerobic bacteria was calculated by measuring the number of colonies in 1 mL of the peritoneal perfusate as described in the Document 27 listed below (results: FIG. 10A).
[0444] Further, the numbers of neutrophils and macrophages in each peritoneal perfusate were also counted. The results are shown in FIG. 10B as "neutrophil" and "macrophage", respectively.
[0445] From a wild-type mouse and mast cell-deficient mouse (kit.sup.W-sh/W-sh), BM-derived macrophages were prepared. These macrophages (cell number: 1.times.10.sup.6) were co-cultured for 1 hour in a medium containing fluorescein-labeled E. coli placed in a 24-well plate, and the number of each type of macrophages that phagocytosed E. coli was counted by flow cytometry to calculate the ratio of phagocytosing macrophages (results: FIG. 11 "BM macrophage").
[0446] A test was carried out under the same conditions as in the above test except that PEC macrophages derived from a wild-type mouse or mast cell-deficient mouse (kit.sup.W-sh/W-sh mouse) were used instead of the BM-derived macrophages derived from a wild-type mouse or mast cell-deficient mouse (kit.sup.W-sh/W-sh mouse), to calculate the ratio of phagocytosing macrophages (results: FIG. 11 "PEC macrophage").
[0447] As shown in FIG. 10A, it can be seen that, at Hour 4 after the CLP, the mast cell-deficient mouse (kit.sup.W-sh/W-sh mouse) showed a lower intraperitoneal bacterial CFU and a larger number of neutrophils than the wild-type mouse. On the other hand, as shown in FIG. 10B and FIG. 11, the number of macrophages and their phagocytosis were not significantly different between the genotypes.
Example 4D
[0448] In order to test whether CD300a is involved in induction (recruitment) of neutrophils or not, the following Example was carried out.
[0449] To mast cell-deficient mice (Kit.sup.W-sh/W-sh mice), PBS buffer containing BMMCs derived from a wild-type mouse (cell number, 1.times.10.sup.6) was administered by intraperitoneal injection (n=20). On Day 28 after the administration, CLP was performed in the same manner as in Example 4A, and the survival rate of the mice was measured (results: FIG. 12B "WT BMMCs.fwdarw.Kit.sup.W-sh/W-sh").
[0450] Further, a test was carried out under the same conditions as in the above test except that BMMCs derived from a CD300a gene-deficient mouse were used instead of the BMMCs derived from a wild-type mouse, and the survival rate of mice was measured (results: FIG. 12B "CD300a.sup.-/- BMMCs.fwdarw.Kit.sup.W-sh/W-sh/Kit.sup.W-sh/W-sh"). "Kit.sup.W-sh/W-sh/Kit.sup.W-sh/W-sh" in FIG. 12B indicates the survival rate of mast cell-deficient mice (Kit.sup.W-sh/W-sh mice) subjected to CLP without administration of BMMCs.
[0451] According to FIG. 12B, the mast cell-deficient mice (Kit.sup.W-sh/W-sh mice) subjected to administration of BMMCs derived from a wild-type mouse showed a higher survival rate even after the CLP, compared to the case where administration of BMMCs was not carried out.
[0452] However, it can be seen that the mast cell-deficient mice (Kit.sup.W-sh/W-sh mice) subjected to administration of BMMCs derived from a CD300a gene-deficient mouse showed a significantly higher survival rate even after CLP, compared to the mice subjected to administration of BMMCs derived from a wild-type mouse and the mice that had not been subjected to administration of BMMCs (FIG. 12B).
[0453] Further, as a result of measuring the bacterial CFU in the same manner as in Example 4C using peritoneal perfusate of each type of mice at Hour 4 after the CLP, the mast cell-deficient mice (Kit.sup.W-sh/W-sh mice) subjected to administration of BMMCs derived from a CD300a gene-deficient mouse showed a significantly higher bacterial clearance than other mice (results: FIG. 12C).
Example 4E
[0454] In order to test whether or not the amount of TNF-.alpha. released increases by intraperitoneal administration of BMMCs to a mast cell-deficient mouse (Kit.sup.W-sh/W-sh mouse), the following Example was carried out.
[0455] Twenty four hours before CLP, a CFSE-labeled BMMC mixture (BMMCs derived from a CD300a gene-deficient mouse:BMMCs derived from a wild-type mouse=1:1 (ratio in terms of the cell number)) was administered to mast cell-deficient mice (Kit.sup.W-sh/W-sh mice) by intraperitoneal injection.
[0456] The mice after injection of the BMMC mixture was subjected to CLP in the same manner as in Example 4A, and, at Hour 4 after the CLP, peritoneal perfusate was collected. Each type of BMMCs contained in the peritoneal perfusate were subjected to analysis using a flow cytometer (FACSCalibur, manufactured by Becton Dickinson; model number, "E6133") (results: FIG. 12D).
[0457] As shown in FIG. 12D, it can be seen that, at Hour 4 after the CLP, the BMMCs derived from a CD300a gene-deficient mouse (CD300a.sup.-CFSE.sup.+ cells) showed production of a significantly larger amount of TNF-.alpha. than the BMMCs derived from a wild-type mouse (CD300a.sup.+CFSE.sup.+ cells).
Example 4F
[0458] In order to test the influence of administration of an anti-CD300a monoclonal antibody (TX41) on CD300a, the following Example was carried out.
[0459] First, CLP was carried out in the same manner as in Example 4A except that 500 .mu.g of an anti-CD300a monoclonal antibody (TX41) (n=13) was intraperitoneally injected to wild-type mice 1 hour or 18 hours before the CLP, and the survival rate of the mice was measured in the same manner as in Example 4D (results: FIG. 12E "Antibody to CD300a").
[0460] Further, a test was carried out under the same conditions as in the above test except that an isotype control antibody (n=11) was used as a control instead of TX41 (n=13), and the survival rate of the mice was measured (results: FIG. 12E "Control antibody").
[0461] As shown in FIG. 12E, it was found that the survival time of wild type mice was longer in the cases where CLP was carried out 1 hour or 18 hours after administration of TX41 by intraperitoneal injection, compared to the cases of administration of the control antibody.
Example 4G
[0462] From each mouse at Hour 4 after the CLP in Example 4F, peritoneal perfusate was collected. The obtained peritoneal perfusate was treated in the same manner as in Example 4C to measure the bacterial CFU and the number of neutrophils (control antibody: n=5 and anti-CD300a monoclonal antibody: n=5) (FIG. 12F and FIG. 12G, respectively).
[0463] The administration of TX41 by intraperitoneal injection did not cause damage to myeloid cells including mast cells in the mice.
[0464] As shown in FIG. 12F and FIG. 12G, the administration of TX41 to wild-type mice by intraperitoneal injection 1 hour or 18 hours before CLP resulted in a significant increase in neutrophils and an increased bacterial clearance in the abdominal cavity.
[0465] From the results of Example 4, it can be understood that inhibition of the interaction between PS and CD300a by TX41 or the like allows prevention of sepsis induced by peritonitis.
[0466] Under physiological conditions, a number of cells undergo apoptosis. In this process, PS receptors play a central role in incorporation of the apoptotic cells, and are indispensable for preventing the progression of autoimmune diseases (Document 22 listed below).
[0467] On the other hand, it is known that, under pathological conditions such as microbial infection, cell death due to apoptosis remarkably increases, and this causes inflammation reaction by mast cells via receptors (e.g., Toll-like receptors) against pathogen-associated molecular patterns (PAMPs) (Documents 15, 23 and 24 listed below). Further, mast cells are known to play important roles in immune reaction against pathogens.
[0468] Thus, from the results in the above Examples, it can be understood that PS not only provides an incorporation signal for phagocytes via several kinds of PS receptors, but also has an effect to effectively suppress inflammation reaction caused by mast cells via CD300a, as newly discovered in the present invention.
[0469] It can therefore be understood that phosphatidyl serine-binding substances (e.g., MFG-E8) and CD300a-binding substances (e.g., neutralizing antibodies against CD300a) inhibit the interaction between PS and CD300a in mast cells to thereby activate the mast cells or maintain such an activity.
[0470] That is, it can be understood that these substance are useful as effective components of immunostimulants used for prophylaxis of various LPS-induced inflammatory infections (and sepsis caused thereby).
[0471] Further, since PS suppresses activation signaling of CD300a and hence suppresses activation of mast cells, PS can be understood to be useful as, for example, an effective component of immunosuppressants to be used for suppressing inflammation reaction in allergic diseases and autoimmune diseases (for example, for suppressing release of chemical mediators such as histamine) to alleviate or treat symptoms of the allergic diseases and autoimmune diseases (i.e., to suppress excessive immune function).
[0472] <Asthma>
[0473] (Materials and Methods)
[0474] (Mice)
[0475] C57BL/6J mice were purchased from Clea Japan, Inc. CD300a gene-deficient mice (CD300a.sup.-/- mice) were obtained by crossing Balb/c CD300a gene-deficient mice prepared in the inventors' laboratory with the purchased WT C57BL/6J mice for 12 generations, and then performing back-crossing. Male and female mice that were 8 to 10 weeks old at the beginning of induction of asthma were used.
[0476] (OVA-Induced Asthma)
[0477] FIG. 13A shows the protocol for inducing asthma with ovalbumin. On Days 0, 7 and 14 after the beginning of induction of asthma, a mixture of 100 .mu.s of ovalbumin (OVA, chicken egg albumin, manufactured by Sigma) and 100 .mu.L of aluminum hydroxide gel (ALUM, ALhydrogel 2%, manufactured by Invitrogen) was intraperitoneally injected to each mouse.
[0478] Further, on Days 21, 22 and 23 after the beginning of induction of asthma, each mouse was subjected to inhalation of 10% ovalbumin prepared by dilution with PBS, for 30 minutes using an ultrasonic nebulizer (NE-U17, Omuron). On Day 25 after the beginning of induction of asthma, each mouse was subjected to bronchoalveolar lavage (BAL), and serum was collected from each mouse.
[Example 5A] (FIG. 13A, FIG. 13B)
[0479] (Bronchoalveolar Lavage BAL)
[0480] After subjecting each mouse to tracheotomy, washing was performed 3 times with 1 mL of 2% FBS/PBS, followed by collecting the washing liquid and measuring the cell number. As shown in FIG. 13A and FIG. 13B, the cell number in the bronchoalveolar lavage fluid (BAL fluid) of each mouse on Day 25 after the beginning of induction of asthma was significantly smaller in the CD300a gene-deficient mice than in the WT mice in terms of both the total cell number and the number of eosinophils. This result indicates that CD300a exacerbated eosinophilic airway inflammation caused by ovalbumin (OVA), and that the CD300a gene-deficient mice showed amelioration of symptoms of eosinophilic airway inflammation.
[Example 5B] (Analysis of Cells in Bronchoalveolar Lavage Fluid) (FIG. 13C)
[0481] With CD45.2-FITC, Siglec-F-PE, CD11b-APC Cy7, CD11c-PEC Cy7 and F4/80-Alexa (all of these were purchased; BD), 1.times.10.sup.6 cells in the collected bronchoalveolar lavage fluid were stained, and CD45.sup.+SiglecF.sup.-CD11b.sup.+CD11c.sup.-F4/80.sup.- was analyzed as the eosinophil fraction by flow cytometry (FACS).
[0482] FACS analysis of cells in the bronchoalveolar lavage fluid of the mice on Day 25 after the beginning of induction of asthma showed the ratio of eosinophils among CD45-positive cells (CD45.sup.+SiglecF.sup.-CD11b.sup.+CD11c.sup.-F4/80.sup.-). The CD300a gene-deficient mice showed a significantly smaller ratio of infiltrating eosinophils in the BAL fluid than the WT mice.
[Example 5C] (Measurement of Serum IgE Value) (FIG. 14)
[0483] Serum IgE was measured by ELISA using a rat anti-mouse IgE (BD) and a biotinylated anti-mouse IgE (BD).
[0484] As shown in FIG. 14, the CD300a gene-deficient mice showed significantly lower serum IgE values during the period of sensitization to cause ovalbumin-induced asthma.
[0485] Based on comparison of the serum IgE value (index of the degree of allergy) among the OVA-induced asthma model mice on Disease Day 14, the CD300a gene-deficient mice showed significantly lower serum IgE values than the WT mice.
[0486] It is thought that administration of an anti-CD300a antibody, which suppresses signal transduction of CD300a, causes significant suppression of serum IgE in ovalbumin-induced asthma.
[0487] <Enteritis>
[0488] (Materials and Methods)
[0489] (Mice)
[0490] C57BL/6 mice (WT mice) were purchased from Clea. CD300a gene-deficient mice established from Balb/cA-derived ES cells in the inventors' laboratory were backcrossed with C57BL/6, and mice of the 12th or later generation were used.
[0491] The mice used for the experiment were kept under a Specific Pathogen-Free (SPF) environment in Laboratory Animal Resource Center, University of Tsukuba.
[0492] (Induction of Enteritis)
[0493] Female C57BL/6 mice of 10 to 12 weeks old were made to drink 2.5% (w/v) dextran sulfate sodium salt, reagent grade (DSS, MP Biomedicals, molecular weight (MW)=36000 to 50000) continuously for 8 days for induction of enteritis.
[0494] (Antibodies)
[0495] BD biotinylated anti-CD300a (TX41) was prepared in the inventors' laboratory. Other antibodies, which are described below, were purchased from Bioscience. [0496] Purified anti-mouse TNF-.alpha. [0497] Biotin-labeled anti-mouse TNF-.alpha. [0498] Purified mouse IL-6 [0499] Biotin-labeled anti-mouse IL-6 [0500] Purified anti-mouse IL-10 [0501] Biotin-labeled anti-mouse IL-10 [0502] Allophycocyanin-Cy7 (APC-Cy7)-labeled anti-mouse CD11b (M1/70) [0503] Phycoerythrin-Cy7 (PE-Cy7)-labeled anti-mouse CD11c (HL3) [0504] Fluorescein isothiocyanate (FITC)-labeled anti-mouse CD4 (L3T4)
[0505] (Large Intestine Tissue Culture Liquid)
[0506] After removal of the large intestine from each mouse on Day 9 after administration of DSS or water, the large intestine was washed with phosphate buffer to remove feces. The large intestine was longitudinally incised, and 5 3-mm tissue pieces were cut out from the portion 3 mm distant from the anus, and the tissue pieces were subjected to 12 hours of culture in 10% FBS RPMI 1640 (manufactured by GIBCO). The supernatant was then collected to provide a large intestine tissue culture liquid. TNF-.alpha., IL-6 and IL-10 contained in the culture liquid were measured by sandwich ELISA.
[0507] (Isolation of Cells of Large Intestinal Lamina Propria)
[0508] Mice were sacrificed, and the large intestine of each mouse was collected. The large intestine was cut into 4 pieces with scissors, and then washed with phosphate buffer to remove feces. The remaining tissue was placed in 1 mM DTT/5 mM MEDTA/Hank's balanced salt solution (manufactured by Sigma), and incubated in an incubator (shaker type) at 37.degree. C. for 30 minutes.
[0509] Subsequently, the tissue was washed again with phosphate buffer to remove detached epithelium and contaminants. The remaining tissue was cut into small pieces with scissors, and placed in 1 mg/mL collagenase type 3 (manufactured by Worthington)/0.1 mg/mL DNase (manufactured by Worthington)/5% Fetal bovine serum/Hanks's balanced salt solution, and incubated in an incubator (shaker type) at 37.degree. C. for 2 hours to allow complete lysis.
[0510] The cell suspension was passed through 70-.mu.m nylon, and then centrifuged. The obtained cells were suspended in 40% Percoll, and overlaid on 70% Percoll. After centrifugation, cells in the intermediate layer were collected to be used as cells of the large intestinal lamina propria in flow cytometry.
[0511] (Isolation of CD11b.sup.+CD11c.sup.+ Cells and CD4.sup.+ T Cells)
[0512] The isolated cells of the large intestinal lamina propria were labeled with APC-Cy7-labeled anti-CD 11b (M1/70), PE-Cy7-labeled anti-CD11c (HL3) and FITC-labeled anti-mouse CD4 (L3T4). The labeled cells were sorted by FACSAria (BD) to provide CD11b.sup.+CD11c.sup.+ cells and CD4.sup.+ T cells, respectively.
[0513] (Measurement of mRNA Level)
[0514] The cells sorted by FACSAria were lysed with ISOGEN-LS (Nippon Gene). From the lysed cells, mRNA was extracted using an mRNA extraction kit, and cDNA was prepared using a reverse transcription kit. The prepared cDNA was mixed with primers (see SEQ ID NOs:1 to 16 in SEQUENCE LISTING) and Power Cyber Green (Applied Biosystems), and measurement of the mRNA levels was carried out using 7500 Fast Real Time PCR System (Applied Biosysytems).
[Example 6A] Body Weight Change Rate (FIG. 15)
[0515] WT mice and CD300a gene-deficient mice were made to drink 2.5% (w/v) dextran sodium sulfate (DSS) continuously for 8 days to induce enteritis, and the body weight of each mouse was measured every day.
[0516] The mice used were female mice of 10 to 12 weeks old, and the rate of change in the body weight was observed for 15 WT mice and 15 CD300a gene-deficient mice. Statistical significance was determined by Student's t-test (**: p<0.01, ***: p<0.001).
[0517] As shown in FIG. 15, the weight loss due to enteritis induced by drinking of 2.5% DSS was significantly milder in the CD300a gene-deficient mice having no MAIR-I (.quadrature.) than in the WT mice (.circle-solid.).
[Example 6B] Measurement of Length of Large Intestine (FIG. 16A)
[0518] The large intestine was collected from 7 WT mice and 7 CD300a gene-deficient mice on Day 6 of the administration of DSS, and the length from the anal region to the cecum region was measured. Statistical significance was determined by Student's t-test (**: p<0.01).
[0519] As shown in FIG. 16A, the CD300a gene-deficient mice had longer large-intestines, and large-intestinal atrophy due to the DSS-induced enteritis was milder in the CD300a gene-deficient mice than in the WT mice.
[Example 6C] Measurement of Length of Large Intestine (FIG. 16B)
[0520] Histological evaluation of the large intestine was simultaneously carried out. As a result, it was observed that the CD300a gene-deficient mice had milder histological changes in the large intestine than the WT mice (FIG. 16B).
[Example 6D] Measurement of Cytotoxic Activity (FIG. 17)
[0521] Large-intestine tissues of the WT mice and the CD300a gene-deficient mice on Day 6 of the DSS administration were stained by HE (hematoxylin-eosin) staining, and scored from 0 to 5 (0: no change; 1: hypertrophy and structural changes in crypts; 2: remarkable decrease in goblet cells; 3: crypts are found, but their structures are hardly maintained; 4: disappearance of crypts is found, but no detachment of the epithelium is found; 5: disappearance of crypts, detachment of the epithelium, and remarkable cellular infiltration are found).
[0522] Each of the large intestines of 7 WT mice and 7 CD300a gene-deficient mice was photographed with visual field enhancement at 10 areas from the anal side, and the obtained images were scored. Statistical significance was determined by Student's t-test (***: p<0.001). The results are shown in FIG. 17.
[0523] As shown in FIG. 17, tissue injury due to the DSS enteritis was mild in the CD300a gene-deficient mice.
[Example 6E] Measurement of IL-10 (FIG. 18A)
[0524] DSS or water (control group) was administered, and the large intestine of each mouse was removed on Day 9 after the administration of DSS. The large intestine was washed with phosphate buffer, and cut into 3-mm.sup.3 pieces from the anal side.
[0525] The minced tissue of the large intestine was cultured in 10% FBS RPMI 1640 (GIBCO) for 12 hours, and the supernatant was then collected. The collected culture supernatant was subjected to measurement of the protein contents of cytokines (TNF-.alpha., IL-6 and IL-10) by sandwich ELISA. Statistical significance was determined by Student's t-test (*: p<0.05).
[0526] As shown in FIG. 18, in the large-intestine tissue with DSS-induced enteritis, the contents of IL-6 and TNF-.alpha. were not different between the WT mice and the CD300a gene-deficient mice. However, higher-level production of IL-10 was found in the CD300a gene-deficient mice (FIG. 18A).
[0527] It is known that, in Mreg cells, IL-10 suppresses expression of CD80 and CD86, which are proteins required for activation of inflammatory T cells (Document 33 listed below).
[0528] As shown in FIG. 18A, the CD300a gene-deficient mice showed an increased production of IL-10 at the 5% significance level. Thus, it is thought that IL-10 suppresses growth induction of inflammatory T cells and activation of inflammatory T cells, thereby contributing maintenance of homeostasis of the gut immune system.
[Example 6F] Measurement of Number of Cells of Each Type (FIG. 18B)
[0529] In mice on Day 0, Day 2, Day 4 and Day 6 after administration of DSS, changes in the cell populations of CD4.sup.+ T cells, CD8.sup.+ T cells, B cells, macrophages and dendritic cells were investigated.
[0530] As a result, as shown in FIG. 18B, no difference was found in the number of cells of each cell population that have moved to the large intestine between the WT mice and the CD300a gene-deficient mice.
[Example 6G] Expression Analysis of CD300a (FIG. 19)
[0531] Cells isolated from the large intestine were stained with APC-Cy7-labeled anti-CD11b (M1/70), PE-Cy7-labeled anti-CD11c (HL3), and Biotin-anti-CD300a (TX41). The cells in the CD11b.sup.-CD11c.sup.- fraction, CD11b.sup.+CD11c.sup.- fraction, CD11b.sup.-CD11c.sup.+ fraction, and CD11b.sup.+CD11c.sup.+ fraction were subjected to measurement of expression of CD300a by flow cytometry. The results are shown in FIG. 19.
[0532] As shown in FIG. 19, in the large intestinal lamina propria, expression of CD300a was found in CD11b.sup.+CD11c.sup.+ cells, which can be said to be special cells.
[Example 6H] Quantification of Expression in CD11b.sup.+CD11c.sup.+ Cells (FIG. 20)
[0533] The CD11b.sup.+CD11c.sup.+ cells isolated from the large intestine of each mouse were sorted by FACSAria, and mRNA was then extracted from the cells. cDNA was prepared from the extracted mRNA, and the mRNA levels of IL-10, TNF-.alpha. and IL-6 were measured by quantitative PCR.
[0534] As a result, as shown in FIG. 20, no difference in the mRNA levels of IL-10, TNF-.alpha. and IL-6 could be found in the CD11b.sup.+CD11c.sup.+ cells between the WT mice and the CD300a gene-deficient mice.
[Example 61] Quantification of Expression in CD4.sup.+ T Cells (FIG. 21)
[0535] CD4.sup.+ cells isolated from the large intestine of each mouse after administration of DSS were sorted by FACSAria, and mRNA was then extracted from the cells. cDNA was prepared from the extracted mRNA, and the mRNA levels of IL-10, Foxp3, TGF-.beta., T-bet, GATA-3 and ROR.gamma.t were measured by quantitative PCR using ABI 7500 fast. Statistical significance was determined by Student's t-test (*: p<0.05, **: p<0.01).
[0536] As shown in FIG. 21, the CD300a gene-deficient mice showed higher mRNA levels of IL-10, Foxp3 and Tgf.beta. in the CD4.sup.+ T cells.
[0537] (Discussion)
[0538] As shown by the results of Examples 6A to 61, in mice with DSS-induced enteritis, deficiency of MAIR-I causes production of anti-inflammatory cytokines such as IL-10 (FIG. 21), leading to amelioration of the disease state of enteritis.
[0539] It is thought that the above phenomenon is not due to an increase in a certain cell population (FIG. 18B), but due to regulation of IL-10 production via CD300a (MAIR-I) by the CD11b-positive dendritic cells per se or certain cells influenced by those dendritic cells.
[0540] That is, it can be predicted that suppression or inhibition of signal transduction of CD300a (MAIR-I) via CD11b-positive dendritic cells may lead to amelioration of the disease state of enteritis.
[0541] <Atopic Dermatitis>
[0542] Involvement of CD300a in atopic dermatitis was investigated. The materials and methods, and Examples are shown below.
[0543] (Experimental Animals)
[0544] Balb/c mice were purchased from Clea Japan, Inc., and kept in the inventors' laboratory under approved breeding room conditions. The mice used in this study were 8 CD300a (MAIR-I) gene-deficient mice and 8 Balb/c wild-type (WT: Wild Type) mice. During the experimental period, each mouse was provided with food and water ad libitum, and kept under normal laboratory conditions.
[0545] (Percutaneous Sensitization)
[0546] Each mouse was mildly anesthetized with isoflurane (Mylan, Osaka, Japan), and the hair on the back was shaved with an electric shaver. An area (1 cm.sup.2) on the back skin of each mouse was subjected to at least 10 times of tape stripping using adhesive cellophane tape.
[0547] On the gauze of Band Aid (registered trademark) tape, 100 .mu.g of ovalbumin (OVA) in 100 .mu.L of phosphate buffered saline was placed, and the resulting tape was applied to the bodies of 5 mice in each group subjected to the tape stripping. To the remaining 3 mice, PBS was applied with tape. The tape was once replaced on Day 2, and OVA sensitization with the tape was carried out every day for 1 week.
[0548] Each mouse was kept without OVA sensitization during Week 2. During Week 3, the mouse was subjected again to OVA sensitization in the same manner as described above. At the end of Week 3, each mouse was sacrificed, and samples for histology and ELISA were collected.
[0549] (Number of Times of Scratching Behavior)
[0550] The number of times of scratching behavior was counted by careful observation of each mouse for 30 minutes at the end of each of Week 1 to Week 3.
[0551] (Histology)
[0552] For histological observation, the skin of the OVA-sensitized area in each mouse was collected. Each collected skin sample was cut into small tissue blocks, and immersed in 4% paraformaldehyde at 4.degree. C. for 24 hours.
[0553] After this fixation, a dehydration step was carried out. All skin samples were then quickly frozen in acetone in a container containing dry ice. The samples were stored at -30.degree. C. until use.
[0554] The skin samples were then cut into sections with a thickness of 4 .mu.m using a frozen section preparation apparatus, Coldtome HM560E (manufactured by Carl Zeiss, Jena, Germany), and placed on slide glasses precoated with New Silane III (Muto Pure Chemicals, Co., Ltd., Tokyo, Japan).
[0555] The tissue sections were stained with hematoxylin-eosin (manufactured by Sakura Finetek Japan), and then stained with toluidine blue (manufactured by Santa Cruz Biotechnology, Inc.).
[0556] Evaluation of histological finding of the tissue was performed for the skin depth (cell layer); infiltration of eosinophils, monocytes and mast cells; and the level of hyperplasia of fibroblasts.
[Example 7A] Number of Times of Scratching Behavior (FIG. 22)
[0557] FIG. 22 shows the number of times of scratching behavior per 30 minutes in each mouse. Pruritus (a disease that causes itch without causing eruption) is a common condition for atopic dermatitis. Therefore, the evaluation was carried out by counting the number of times of scratching behavior by careful observation of each animal for 30 minutes during the OVA sensitization.
[0558] As shown in FIG. 22, the OVA-sensitized WT mice (.diamond-solid.) showed a severe condition of scratching behavior after the OVA sensitization, and a larger number of times of scratching behavior than the OVA-sensitized CD300a gene-deficient mice (.tangle-solidup.). The largest number of times of scratching behavior was observed at the end of the 4th OVA sensitization.
[0559] During the OVA sensitization, the most severe symptom of scratching behavior was observed in the WT mice (FIG. 22). Appearance of a severe symptom of scratching behavior is one of the most common pathological features of atopic dermatitis. Thus, involvement of CD300a (MAIR-I)-positive cells in an important role in atopic dermatitis might become clear.
[Example 7B] Observation (see FIG. 23)
[0560] The skin of each OVA-sensitized mouse was observed.
[0561] FIG. 23(A) shows the skin of a WT mouse that had not undergone OVA sensitization. In untreated WT mice, which had not been subjected to ovalbumin sensitization, cellular infiltration was not found in the dermis, and a thin epidermis could be observed. The lower panels in FIG. 23 show magnified views of the rectangular areas in the upper panels. The length of each thick bar corresponds to 10 .mu.m.
[0562] On the other hand, as shown in FIG. 23(B), the skin of the OVA-sensitized WT mice showed hyperplasia of the epidermis and hyperplasia of fibroblasts. Obvious infiltration of eosinophils (see white arrowheads) and monocytes was found in the dermis of the skin in the WT mice subjected to sensitization with ovalbumin.
[0563] As shown in FIG. 23(C), in the epidermis of the CD300a gene-deficient mice that had not been subjected to OVA sensitization, cellular infiltration was not found, and a thin epidermis could be observed similarly to FIG. 23(A).
[0564] Surprisingly, as shown in FIG. 23(D), the skin of the OVA-sensitized CD300a gene-deficient mice showed an increased thickness of the epidermis, but did not show hyperplasia of the epidermis. Further, the dermis showed neither cellular infiltration nor hyperplasia of fibroblasts.
[Example 7C] Staining of Mast Cells (See FIG. 24)
[0565] As shown in FIG. 24, all mouse skin samples were subjected to toluidine blue staining. Toluidine blue staining is a staining method that preferentially stains intracellular granules in mast cells.
[0566] FIG. 24(A) shows the skin of a WT mouse that had not been subjected to OVA sensitization. FIG. 24(B) shows the skin of an OVA-sensitized WT mouse. FIG. 24(C) shows the skin of a CD300a gene-deficient mouse that had not been subjected to OVA sensitization. FIG. 24(D) shows the skin of an OVA-sensitized WT mouse. Stained mast cells are indicated by open arrowheads.
[0567] As shown in FIG. 24, after the OVA sensitization, an increased number of mast cells were found in the skin of the WT mouse. Moreover, the epidermis in the mouse was thicker than that in the CD300a gene-deficient mouse.
[Example 7D] Measurement of Number of Cell Layers (FIG. 25A)
[0568] All mouse skin samples were subjected to counting of the number of cell layers in the epidermis (see FIG. 25A). After the OVA sensitization, the WT mice showed the largest number of cell layers. On the other hand, in the OVA-sensitized CD300a gene-deficient mice, the number of cell layers was less than half of this number.
[Example 7E] Counting of Number of Eosinophils and Number of Mast Cells (FIG. 25B)
[0569] As shown in FIG. 25B, all mouse epidermal samples were subjected to counting of the number of eosinophils and the number of mast cells showing infiltration into the dermis, which are indices for atopic dermatitis. The largest cell numbers were observed in the epidermis of the WT mice after the sensitization with ovalbumin.
[0570] On the other hand, in the OVA-sensitized CD300a gene-deficient mice, the increases in the number of eosinophils and the number of mast cells were milder than those in the WT mice. After the OVA sensitization, hyperplasia of epidermis and hyperplasia of fibroblasts appeared most severely in the WT mice (FIG. 23 and FIG. 25A). Hyperplasia of epidermis and fibroblasts is another major pathological feature of atopic dermatitis.
[0571] Further, a high level of infiltration of eosinophils, mast cells and monocytes was found in the skin of the OVA-sensitized WT mice (FIG. 23 to FIG. 25B). Interaction between infiltrating eosinophils and hyperplasia of fibroblasts causes secretion of IL-31. IL-31 is an itch-inducing cytokine (Document 34 listed below: Wong C K et al., 2012).
[0572] Therefore, after OVA sensitization, WT mice show more severe features of atopic dermatitis than CD300a gene-deficient mice.
[0573] (Immunohistology)
[0574] In order to detect CD300a (MAIR-I) and the Langerin antigen on serial sections of the skin, the single-step or two-step method of enzyme immunohistochemistry was carried out. First, all sections were rinsed 3 times with phosphate-buffered saline supplemented with 0.05% Tween (TPBS, pH 7.4). The sections were then immersed for 30 minutes in each of cold absolute methanol and 0.5% H.sub.2O.sub.2.
[0575] After the washing in TPBS, the sections were subjected to blocking treatment using the Blocking One Histo reagent (Nacalai Tesque, Inc., Kyoto, Japan) for 10 minutes, and then washed with TPBS. The skin sections were cultured at 4.degree. C. for 18 hours in the presence of anti-Langerin goat IgG (Santa Cruz Biotechnology, Inc., Santa Cruz, Calif., US) and biotinylated anti-CD300a (MAIR-I) rat IgG 2a.lamda. (prepared in the inventors' laboratory), and then at room temperature for 1 hour in the presence of biotinylated donkey anti-goat IgG as the secondary antibody for Langerin.
[0576] Finally, these were cultured in the presence of DAB/Metal Concentrate (Thermo Scientific, Waltham, Mass., US), and counterstained with hematoxylin. Negative control sections were cultured in the presence of TPBS or an isotype control antibody instead of the primary antiserum.
[Example 7F] Langerin Immunostaining (FIG. 26)
[0577] The epidermis of each mouse was investigated to see whether or not it is positive for Langerin, which is a dendritic cell marker in immunostaining. Langerin is a C-type lectin expressed in Langerhans cells, and also expressed in part of dermal dendritic cells. Langerin is involved in recognition and incorporation of antigens, and formation of Birbeck granules, which are responsible for intracellular antigen delivery.
[0578] (Results)
[0579] FIG. 26(A) shows the epidermis of an untreated WT mouse (mouse without OVA sensitization). FIG. 26(B) shows the epidermis of an OVA-sensitized WT mouse. FIG. 26(C) shows the epidermis of an untreated CD300a gene-deficient mouse. FIG. 26(D) shows the epidermis of an OVA-sensitized CD300a gene-deficient mouse. The panels a, b, c and d are magnified views of the rectangular areas in the upper panels.
[0580] After the OVA sensitization, a significantly larger number of Langerin-positive cells were found in the epidermis of WT mice than in the epidermis of CD300a gene-deficient mice. The arrowheads indicate Langerin-positive cells. Each scale bar represents 10 .mu.m.
[Example 7G] Counterstaining after Langerin Immunostaining (FIG. 27)
[0581] Immunopositivity of the Langerin antibody was evaluated by counterstaining with toluidine blue. The skin of the WT mouse showed an increased number of Langerin-positive cells in the dermis. In the dermis, interaction of several Langerin-positive cells with mast cells was found. Mast cells were stained in purple. Each scale bar represents 10 .mu.m.
[0582] Langerhans cells and skin dendritic cells are major antigen-presenting cells in the skin. Langerhans cells are positive for the Langerin antigen in their cell membranes, and Langerin-positive cells are also present among skin dendritic cells (Document 35 listed below: Nakajima S. et al, 2012).
[0583] Skin Langerin-positive dendritic cells interact with mast cells to activate CD4-positive T cells (Document 36 below: Otsuka A. et al., 2011). In the OVA-sensitized model, the numbers of mast cells (FIG. 24 and FIG. 25B) and Langerin-positive cells (see FIG. 26) largely increased in the skin of the OVA-sensitized WT mice (see white arrowheads in FIG. 26 for comparison).
[0584] In the skin of the OVA-sensitized WT mice, interaction between mast cells and Langerin-positive cells was found (FIG. 27).
[0585] (Discussion)
[0586] Thus, it can be deduced that atopic dermatitis more severely appears in the skin of WT mice than the skin of CD300a gene-deficient mice. These results suggest that CD300a (MAIR-I) plays an important role in atopic dermatitis.
[0587] In the dermis of the WT skin, CD300a (MAIR-I)-positive cells largely increased after the OVA sensitization (FIG. 27). This result further confirms that CD300a (MAIR-I)-positive cells play an important role in atopic dermatitis.
[0588] (Treatment of Atopic Dermatitis)
[0589] (Treatment with Anti-CD300a (MAIR-I) Antibody)
[0590] In the present experiment, 6 Balb/c mice of 7 weeks old were used. According to the protocol shown in FIG. 29, 3 animals out of the 6 animals were subjected to intravenous injection of anti-CD300a (MAIR-I) rat IgG 2a.lamda. (TX41), and the remaining 3 animals were subjected to intravenous injection of a rat IgG 2a.lamda. control antibody (TX74). Both of these antibodies were prepared and checked in the inventors' laboratory. "TX74" is an isotype control antibody of TX41, and does not have a neutralizing action as described below. Each antibody was diluted with sterile PBS to an antibody concentration of 1600 .mu.g/mL, and 150 .mu.L of the dilution was injected at once.
[0591] <Blocking of CD300a (MAIR-I) Antigen by Injection of CD300a (MAIR-I) Antibody>
[0592] FIG. 29 shows the procedure to block the CD300a (MAIR-I) antigen in a Balb/c WT mouse. Each thick line indicates the period of OVA sensitization. The arrowheads indicate the schedule of injection of the antibodies.
[0593] Total serum IgE was evaluated at the end of each week of sensitization. Histological samples were collected after the continuous sensitization. The anti-rat CD300a (MAIR-I) antibody IgG 2a.lamda. (TX41), and the control antibody (TX74), which is an isotype thereof, were used for intravenous injection.
[0594] (ELISA)
[0595] Peripheral blood was collected from the retro-orbital cavity using a plain glass hematocrit tube (Drummond Scientific Company, Broomall, Pa., US), and centrifuged at 12000 rpm for 5 minutes.
[0596] Serum was collected by cutting the tube, and the whole serum was diluted with a blocking serum before use in ELISA. The ELISA experiment was carried out according to the standard protocol for total IgE, recommended by BD Biosciences, California, US.
[Example 7H] Immunoreaction with Anti-CD300a Antibody (Confirmation of Presence of Receptors) (FIG. 28)
[0597] Immunopositivity of an anti-CD300a antibody was evaluated with a skin sample of each mouse. FIG. 28(A) shows untreated epidermis of a WT mouse, and (B) shows OVA-sensitized epidermis of a WT mouse.
[0598] The epidermis of WT mice after OVA sensitization showed cells that are significantly immunopositive. The epidermis of CD300a gene-deficient mice did not show immunopositive reaction. The arrowheads in (B) indicate CD300a (MAIR-I)-positive cells. Each scale bar represents 10
[Example 71] Treatment by Administration of CD300a Antibody (FIG. 30)
[0599] (ELISA)
[0600] As shown in FIG. 29, each of TX41 and TX74 was administered to WT mice according to the above-described procedure, and the IgE level, which is an index of atopic dermatitis, was measured.
[0601] FIG. 30 shows the total serum IgE level measured by ELISA. The IgE level after OVA sensitization was higher in the mice to which TX74 was injected than in the mice to which TX41 was injected.
[Example 7J] Treatment by Administration of Anti-CD300a Antibody (FIG. 31)
[0602] As shown in FIG. 31, after the OVA sensitization, the WT mice to which TX74 was injected showed a more severe scratching behavior than the WT mice to which TX41 was injected.
[Example 7K] H&E Staining (FIG. 32)
[0603] As shown in FIG. 32, skin sections of the WT mice in Example 71 were subjected to H&E staining. After the OVA sensitization, the skin of the WT mice to which TX74 was injected showed higher levels of hyperplasia of the epidermis and infiltration of monocytes than the skin of the WT mice to which TX41 was injected. In FIG. 32, the scale bar in the lower right corner of each photograph represents 10
[Example 7L] Toluidine Blue Staining (FIG. 33)
[0604] As shown in FIG. 33, the skin of each WT mouse was subjected to toluidine blue staining. After the OVA sensitization, the skin of the WT mice to which TX71 was injected showed more mast cells than the skin of the WT mice to which TX41 was injected. Each scale bar represents 10 .mu.m, similarly to FIG. 32.
[0605] (Discussion)
[0606] The total IgE and the number of scratching behavior were higher in the mice to which TX74 was injected than the mice to which TX41 was injected (see FIG. 30 and FIG. 31). The epidermal thickness, number of infiltrating cells, number of fibroblasts and number of mast cells were also higher in the mice to which TX74 was injected than the mice to which TX41 was injected (see FIG. 32 and FIG. 33).
[0607] From these results, it could be further confirmed that CD300a (MAIR-I) plays an important role in atopic dermatitis, and the effect of TX41, which is an anti-CD300a antibody, as a therapeutic agent for atopic dermatitis could be confirmed.
[0608] <Celiac Disease>
[0609] Celiac disease (CD) is a progressive enteritis caused by immune response to dietary gluten protein. The adaptive immune response specific to the gliadin peptide derived from gluten is involved in the progression of celiac disease. The innate immune response as the fundamental cause of the disease has not been completely elucidated.
[0610] Here, we demonstrate that CD300a (MAIR-I), which is expressed in lamina propria macrophages (macrophages in the lamina propria) and is a member of the bone marrow-associated immunoglobulin-like receptor family, plays a regulatory role in the progression of dietary gluten-induced intestinal diseases.
[0611] CD300a gene-deficient mice, which lack CD300a (MAIR-I), fed with a high-gluten diet showed celiac disease-like symptoms. Compared to wild-type (WT) mice fed with a high-gluten diet, each CD300a gene-deficient mouse showed a mild increase in the body weight, high clinical score, large amount of transglutaminase 2, and accumulation of lamina propria macrophages in the jejunum.
[0612] Compared to the WT mice fed with a high-gluten diet, lamina propria macrophages of each CD300a gene-deficient mouse fed with a high-gluten diet showed higher expression of IL-6, IL-15, TNF-.alpha., IFN-.beta., MCP1 and MCP5.
[0613] After in vitro stimulation with the toxic gliadin peptide (P31-43), lamina propria macrophages of each CD300a gene-deficient mouse showed higher-level expression of the above-described cytokines.
[0614] Enhanced expression of IL-6, TNF-.alpha. and IFN-.beta. occurred in lamina propria macrophages of CD300a gene-deficient mice, which lack CD300a (MAIR-I) and have a MyD88-deficient and TRIF-deficient genetic background.
[0615] Further, blocking of binding of phosphatidyl serine, which is the ligand of CD300a (MAIR-I) on apoptotic cells, to CD300a (MAIR-I) promoted production of cytokines in lamina propria macrophages of WT mice.
[0616] In summary, these results indicate that the interaction between macrophages in the intestinal lamina propria and CD300a (MAIR-I) on apoptotic cells plays a protective role in progression of celiac disease, by MyD88- and TRIF-mediated inhibitory gliadin signaling pathways.
[0617] (Materials and Methods)
[0618] (Mice)
[0619] Male Balb/c wild-type (WT) mice of 9 to 14 weeks old, and littermates of Balb/c CD300a gene-deficient mice, which do not have CD300a (MAIR-I), were provided. These mice were used in an experiment in which they were fed with a normal diet (ND), high-gluten diet (HGD) or gluten-free diet (GFD).
[0620] For in vitro experiments, Balb/c WT mice; and CD300a gene-deficient Balb/c or C57BL/6 B6 mice, which have no CD300a (MAIR-I); were used. These mice had the same sex and age. MyD88-gene deficient B6 mice and TRIF gene-deficient B6 mice were purchased from OrientalBio Service, Inc. (Kyoto, Japan). All mice were kept under specific pathogen-free conditions.
[0621] (Feeding Test)
[0622] WT mice and CD300a (MAIR-I) mice were kept with normal powder diet containing less than 2% gluten (MF, Oriental Yeast Co., Ltd., Tokyo, Japan) (ND). WT mice and CD300a gene-deficient mice were kept with a high-gluten diet (HGD), which is the same as MF except that gluten is contained at 30% (Sigma-Aldrich, St. Louis, Mo.). In several experiments, mice were fed with pellets of ND (MF) or a gluten-free diet (GFD; AIN-76A, Research Diets, Inc., New Brunswick, N.J.).
[0623] (Depletion of Microbiota)
[0624] Depletion of the microbiota was carried out as in the [Document 37 listed below]. Depletion of the microbiota was confirmed by observing colony forming units in feces from each mouse using an agarose plate containing brain-heart infusion medium.
[0625] (Histopathological Analysis)
[0626] Histopathological changes in mice were observed in the later-described specific weeks after feeding with ND, HGD or GFD. Samples of the jejunum and colon were isolated and fixed with formalin, followed by staining with hematoxylin-eosin.
[0627] The standard for the clinical score for the jejunum was defined as in the [Document 38 listed below]. Briefly, the score of the ratio between crypts and villi was calculated according to the average depths of crypts and villi (score 0 to 3). Further, the score (score 0 to 3) of infiltration of monocytes was calculated according to the average diameters of the lamina propria of villi, and crypts. The final clinical score was represented as the total of 0 to 6.
[0628] The number of intraepithelial lymphocytes in intestinal epithelial cells (IECs) of the jejunum was counted, and represented as the IEL number per 100 intestinal epithelial cells ([Document 39 listed below]). Quantification of transglutaminase 2 (TG2) in the jejunum was carried out using a tissue suspension of the jejunum. The test was carried out using TG2-CovTest (Zedira, Darmstadt, Germany) according to the manufacturer's instructions.
[0629] (Titration of IgG and IgA Antibodies Against Gliadin)
[0630] The serum antibody titers of IgG and IgA against gliadin in mice were determined by an enzyme immunoassay. This was carried out by the method described in the [Document 40 listed below] with some modifications.
[0631] An anti-mouse IgG antibody labeled with horseradish peroxidase (HRP) (GE Healthcare, Little Chalfont, UK), and an anti-mouse IgA antibody labeled with HRP (Southern Biotech, Birmingham, Ala.) were used as anti-gliadin IgG and IgA detection antibodies, respectively. OPD Reagent (Sigma-Aldrich) was used as the substrate of HRP in colorimetric analysis.
[0632] (Isolation of Lamina Propria Macrophages and Dendritic Cells)
[0633] Lamina propria macrophages (LP M.phi.) and dendritic cells (DCs) were isolated according to the [Document 41 listed below] with some modifications.
[0634] After removal of the mesentery and Peyer patches, the jejunum was cut into small pieces. The pieces were washed a total of 3 times with PBS buffer supplemented with 2 mM EDTA (Sigma Aldrich) and 20% FCS, with shaking at 37.degree. C. for 15 minutes.
[0635] The remaining tissue was homogenized, and then digested in PBS buffer supplemented with 1.5 mg/mL type VIII collagenase (Sigma-Aldrich) and 20% FCS, with shaking at 37.degree. C. for 20 minutes.
[0636] Lamina propria macrophages and lamina propria dendritic cells expressing CD45 in the tissue suspension were concentrated using biotinylated anti-mouse CD45 (30-F11, BD Biosciences, Franklin Lakes, N.J.) and streptavidin particles plus IMAG (BD Biosciences).
[0637] The CD45-expressing cells in the lamina propria were stained with a fluorescein isothiocyanate-conjugated anti-mouse CD11b (M1/70), phycoerythrin-conjugated CD11c (HL3), propidium iodide, Alexa 647-labeled TX41 (anti-mouse CD300a (anti-mouse MAIR-I), rat IgG2a) or Alexa 647-labeled TX74 (anti-FLAG, isotype control for TX41), biotinylated anti-mouseCD45, and streptavidin allophycoerythrin Cy7 (allophycoerythrin-Cy7).
[0638] All fluorescently labeled antibodies and streptavidin allophycoerythrin-Cy7 were purchased from BD Biosciences.
[0639] CD11b.sup.+CD11c.sup.low cells and CD11c.sup.+ cells gated based on the number of CD45.sup.+PI.sup.- cells were isolated as lamina propria macrophages and lamina propria dendritic cells using FACSAria (BD Biosciences).
[0640] An important principle of flow cytometry data analysis is to selectively visualize the particles of interest while removing unnecessary particles (dead cells, residues and the like). This operation is called gating (gate).
[0641] (Flow Cytometry Analysis)
[0642] Macrophages, CD11b.sup.+ dendritic cells and CD11b.sup.- dendritic cells among the cells of lamina propria, which are CD45.sup.+PI.sup.-, were gated based on the numbers of CD11b.sup.+CD11c.sup.low, CD11b.sup.+CD11c.sup.+ and CD11b.sup.-CD11c.sup.+, respectively.
[0643] Intraepithelial lymphocytes and intestinal epithelial cells were obtained by washing small pieces of jejunum with PBS buffer supplemented with 2 mM EDTA and 20% FCS. The intraepithelial lymphocytes and intestinal epithelial cells contained in the suspension were gated based on the number of CD45.sup.+PI.sup.- cells and the number of CD45.sup.-PI.sup.- cells.
[0644] Expression of CD300a (MAIR-I) in macrophages, CD11b.sup.+ dendritic cells and CD11b.sup.- dendritic cells in the lamina propria, intraepithelial lymphocytes, and intestinal epithelial cells were analyzed with Alexa 647-labeled TX41 (anti-mouse MAIR-I, rat IgG2a) or Alexa 647-labeled TX74 (anti-FLAG of TX41, isotype control).
[0645] Apoptotic cells presenting phosphatidyl serine (PS) were analyzed using allophycocyanin-conjugated annexin V. Flow cytometry analysis was carried out using FACSAria (BD Biosciences).
[0646] (Quantitative RT-PCR Method)
[0647] In Isogen LS (Nippon Gene, Tokyo, Japan), 10000 to 20000 lamina propria macrophages and lamina propria dendritic cells that were stimulated with or not stimulated with the toxic gliadin peptide P31-43 were resuspended, and total RNA was isolated from the resulting suspension according to the manufacturer's instructions.
[0648] Single-stranded DNA was synthesized from total RNA using a cDNA reverse transcription kit (Applied Biosystems, Foster City, Calif.). The primer sets used for quantitative reverse transcriptase-mediated polymerase chain reaction (Q-RT-PCR) were designed by PrimerBank (http://pga.mgh.harvard.edu/primerbank/) [Document 42 listed below].
[0649] Q-RT-PCR was carried out using Platinum SYBR Green Super Mix UDG (Invitrogen, Carlsbad, Calif.) and ABI 7500 Fast (Applied Biosystems). The data were analyzed by the DDCT (delta delta CT) method.
[0650] Control samples expressing all genes tested in this study were prepared from spleen cells stimulated with lipopolysaccharide (1000 ng/mL, 6 hours), and were single-stranded DNA. The results were shown based on comparative quantification with samples showing expression of the respective genes.
[0651] (Gliadin Stimulation)
[0652] The toxic .alpha.-gliadin peptide P31-43 derived from gluten [Document 43 listed below] and the ovalbumin peptide P323-339 as a control were synthesized by Operon Biotechnologies (Huntsville, Ala.).
[0653] The purities of these peptides were not less than 95%, and the endotoxin unit/mL was less than 0.001. This was confirmed with Limulus Color KY Test Wako (Wako Pure Chemical Industries, Ltd., Osaka).
[0654] Macrophages in thioglycolate-induced peritoneal exudate cells (PECs) prepared from a B6 mouse were stimulated using 100 .mu.g/mL gliadin peptide P31-43 or ovalbumin P323-339 as described in the [Document 44 listed below].
[0655] In wells of a 96-well flat-bottom plate, 10000 to 20000 lamina propria macrophages and CD11b.sup.+ dendritic cells derived from WT mice or CD300a gene-deficient mice were cultured in RPMI 1640 medium supplemented with 10% FCS, glutamine/streptomycin/penicillin, HEPES and non-essential amino acids.
[0656] Lamina propria macrophages of Balb/c mice or B6 mice were stimulated for 10 hours or 3 hours with 100 .mu.g/mL gliadin peptide P31-43. In several experiments, the lamina propria macrophages were treated or not treated with MFG-E8 (final concentration, 5 .mu.g/mL) of a recombinant mouse at 4.degree. C. for 30 minutes, and further, stimulated with the gliadin peptide P31-43 at 37.degree. C. for 3 hours.
[0657] (Statistical Analyses)
[0658] All statistical analyses were carried out using the Mann-Whitney U test. Statistical significance was judged at P<0.05.
[Example 8A] Body Weight (BW) Change (FIG. 34)
[0659] In order to investigate whether or not CD300a (MAIR-I) is involved in the progression of celiac disease (CD), CD300a gene-deficient Balb/c mice or wild-type (WT) mice were kept with a normal diet containing less than 2% gluten (ND) or a high-gluten diet containing 30% gluten (HGD). After starting feeding with the normal diet or high-gluten diet, changes in the body weight (BW) were monitored.
[0660] ".smallcircle." represents "WT mice, n=7"; ".circle-solid." represents "CD300a gene-deficient mice, normal diet, n=9"; ".quadrature." represents "WT mice, high-gluten diet, n=10"; and ".box-solid." represents "CD300a gene-deficient mice, high-gluten diet, n=14".
[0661] The CD300a gene-deficient mice kept with the high-gluten diet (HGD) showed a significantly smaller degree of increase in the body weight (BW) than the WT mice kept with the high-gluten diet (HGD) (FIG. 34). The CD300a gene-deficient mice were confirmed to show progression of celiac disease-like enteropathy after feeding with the high-gluten diet. These results suggest that CD300a (MAIR-I) plays a protective role in the progression of the intestinal disease in the jejunum after feeding with a high-gluten diet.
[Example 8B] H&E Staining (FIG. 35)
[0662] Histopathological analysis of the intestines of mice at Week 20 after feeding with a normal diet or a high-gluten diet (HGD) was carried out. The jejunum of each mouse was fixed with formalin, followed by staining with hematoxylin-eosin. FIG. 35 shows a representative result obtained for a mouse in each group.
[0663] The CD300a gene-deficient mice kept with a high-gluten diet (HGD) showed a more advanced intestinal disease in the jejunum compared to the WT mice kept with a high-gluten diet (HGD) (see FIG. 35)
[Example 8C] Number of Intraepithelial Lymphocytes (FIG. 36, FIG. 37)
[0664] According to the above-described method, the clinical score and the number of intraepithelial lymphocytes (IELs) per 100 intestinal epithelial cells (IECs) in the jejunum of each mouse were counted at Week 20 after feeding with the normal diet or the high-gluten diet. "* and **" represent p<0.05 and p<0.01, respectively.
[0665] As described above, the CD300a gene-deficient mice showed atrophy of villi in the jejunum (FIG. 35), and significantly higher clinical scores (see FIG. 36). Further, an increase in intraepithelial lymphocytes (IELs) in small-intestinal epithelial cells (IECs) was found as compared to the CD300a gene-deficient mice kept with FD as well as the WT mice kept with a high-gluten diet (HGD) (see FIG. 37).
[Example 8D] TG2-CovTest (Colorimetric Analysis for Quantitation of Specific TG2 Cross-Linking Activity) (FIG. 38)
[0666] The level of transglutaminase 2 (TG2) is known to be influenced by intestinal inflammation. TG2 is a key enzyme for the progression of celiac disease. WT mice and CD300a gene-deficient mice were fed with a normal diet or high-gluten diet for at least 20 weeks, and the level of transglutaminase 2 (TG2) in a jejunum suspension of each mouse was quantified with TG2-CovTest.
[0667] As a result, the TG2 level in the intestine was highest in the CD300a gene-deficient mice kept with a high-gluten diet (HGD) among the groups of mice (see FIG. 38).
[Example 8E] Flow Cytometry (FIG. 39)
[0668] Expression of CD11b and CD11c in lamina propria cells gated for the CD45.sup.+PI.sup.- cell population was analyzed by flow cytometry. Lamina propria (LP) cells of WT mice or CD300a gene-deficient mice of the Balb/c strain fed with the normal diet or high-gluten diet were analyzed by flow cytometry. CD45-expressing LP cells were concentrated using the I-Mag technology (cell separation system using magnetic beads).
[0669] CD11b.sup.+CD11c.sup.low cells are lamina propria macrophages, and CD11b.sup.+CD11c.sup.+ and CD11b.sup.-CD11c.sup.+ are lamina propria (LP) dendritic cells (DCs).
[Example 8F] Quantitative and Qualitative Changes in Immunocytes (FIG. 40)
[0670] In each of WT mice and CD300a gene-deficient mice, the frequency of lamina propria (LP) macrophages (open bar), the frequency of lamina propria (LP) CD11b.sup.+ dendritic cells (closed bar), and the frequency of lamina propria (LP) CD11b.sup.- dendritic cells (shaded bar) were measured.
[0671] That is, quantitative and qualitative changes in the immunocytes in the lamina propria of the jejunum of WT mice and CD300a gene-deficient mice kept with a normal diet (ND) or high-gluten diet (HGD) were investigated.
[0672] The frequency of lamina propria macrophages expressing CD11b.sup.+CD11c.sup.low (lp Mf) was significantly higher in the CD300a gene-deficient mice kept with a high-gluten diet (HGD) than in the WT mice kept with a high-gluten diet (HGD) or in the CD300a gene-deficient mice kept with a normal diet (ND) (see and compare the cell populations 5.8 in the lower right panel, 2.3 in the lower left panel, and 2.2 in the upper right panel of FIG. 39; and see FIG. 40).
[0673] Lamina propria macrophages in the intestine are cells differentiated from inflammatory monocytes expressing CCR2 in peripheral blood [Document 45 listed below].
[0674] In view of the fact that CCR2 is a receptor of MCP1 and MCP5, the increased expression of these chemokines in lamina propria macrophages after stimulation with gliadin is consistent with the high frequency of lamina propria macrophages in the CD300a gene-deficient mice kept with a high-gluten diet (HGD) (see FIG. 40).
[Example 8H] Gene Expression Levels of Cytokines and Chemokines (FIG. 41)
[0675] The gene expression levels of cytokines and chemokines in lamina propria (LP) macrophages isolated from each mouse were analyzed by quantitative RT-PCR. In FIG. 41, the expression level of each gene is expressed as the relative level with respect to the level in WT mice fed with a normal diet, which is taken as "1" to be used as a standard.
[0676] Lamina propria macrophages in the CD300a gene-deficient mice kept with a high-gluten diet (HGD) showed significantly higher levels of inflammatory cytokines (IL-6, IL-15, TNF-.alpha., IFN-b and MCP1) and some chemokines such as MCP5 compared to the WT mice kept with a high-gluten diet (HGD) (see FIG. 41).
[0677] The enhanced expression of inflammatory cytokines and chemokines from lamina propria macrophages in the CD300a gene-deficient mice kept with a high-gluten diet (HGD) is thought to be due to the intestinal disease observed in the mice.
[0678] Further, interestingly, lamina propria macrophages in the CD300a gene-deficient mice kept with a normal diet (ND) showed higher expression of IL-15 and p19 compared to the WT mice kept with ND (FIG. 48).
[Example 81] (FIG. 42)
[0679] Expression of CD300a (MAIR-I) in lamina propria macrophages, CD11b.sup.+ dendritic cells, CD11b.sup.- dendritic cells, intestinal epithelial cells and intraepithelial lymphocytes was analyzed by flow cytometry.
[0680] CD300a (MAIR-I) was detected using TX41 (anti-MAIR-I mouse monoclonal antibody). Lamina propria macrophages (M.phi.), CD11b.sup.+ dendritic cells and CD11b dendritic cells were gated based on the number of CD45.sup.+PI.sup.- lamina propria cells.
[0681] Intraepithelial lymphocytes (IELs) and intestinal epithelial cells (IECs) were gated into the CD3.sup.+CD45.sup.+PI.sup.- and CD45.sup.-PI.sup.- fractions, respectively.
[0682] CD300a (MAIR-I) is expressed in most of bone marrow cells including macrophages, granulocytes, mast cells and dendritic cells ([Document 46 listed below]).
[0683] As described above, the present inventors analyzed expression of CD300a (MAIR-I) in macrophages on the lamina propria, CD11b.sup.+ inflammatory dendritic cells in the lamina propria, CD11b.sup.- tolerogenic dendritic cells, intestinal epithelial cells (IELs) and intraepithelial lymphocytes (IECs), and lamina propria macrophages, and further investigated the CD300a (MAIR-I)-expressing subpopulations of lamina propria CD11b.sup.+ dendritic cells and lamina propria CD11b.sup.- dendritic cells. As a result, no expression of CD300a (MAIR-I) was found in the intraepithelial lymphocytes (IELs) and intraepithelial lymphocytes (IECs) (FIG. 42).
[Example 8J] (FIG. 43)
[0684] Although the receptor of the gliadin peptide has not been identified, earlier studies reported that the gliadin P31-43 peptide accumulates in early endosomes and inhibits their maturation (Documents 47 and 48 listed below).
[0685] Lamina propria (LP) CD11b.sup.+ dendritic cells were isolated from Balb/c WT mice or CD300a gene-deficient mice, and stimulated in vitro with 100 mg/mL toxic gliadin peptide P31-43 for 10 hours.
[0686] The gene expression levels of cytokines and chemokines (IL-6, IL-15, TNF-.alpha., IFN-.beta., MCP1 and MCP5) in lamina propria (LP) CD11b.sup.+ dendritic cells in the WT mice and the CD300a gene-deficient mice after the gliadin stimulation were analyzed by quantitative RT-PCR. The expression level of each gene is expressed as the relative level with respect to the level in untreated WT mice, which is taken as "1" to be used as a standard.
[0687] As a result of the investigation of the gene expression levels, the expression levels of IL-6, IL-15, TNF-.alpha., IFN-.beta., MCP1 and MCP5 after stimulation with the gliadin peptide P31-43 were significantly higher in lamina propria macrophages of CD300a gene-deficient mice than in lamina propria macrophages of WT mice (FIG. 43).
[0688] Inflammatory lamina propria CD11b.sup.+ dendritic cells of CD300a gene-deficient mice kept with a normal diet (ND) showed higher expression of IL-15 as compared to WT mice kept with a normal diet (ND).
[Example 8K] Suppression of Gliadin-Induced Expression of Cytokines (FIG. 44)
[0689] The present inventors tested whethe
References
D00001
D00002
D00003
D00004
D00005
D00006
D00007
D00008
D00009
D00010
D00011
D00012
D00013
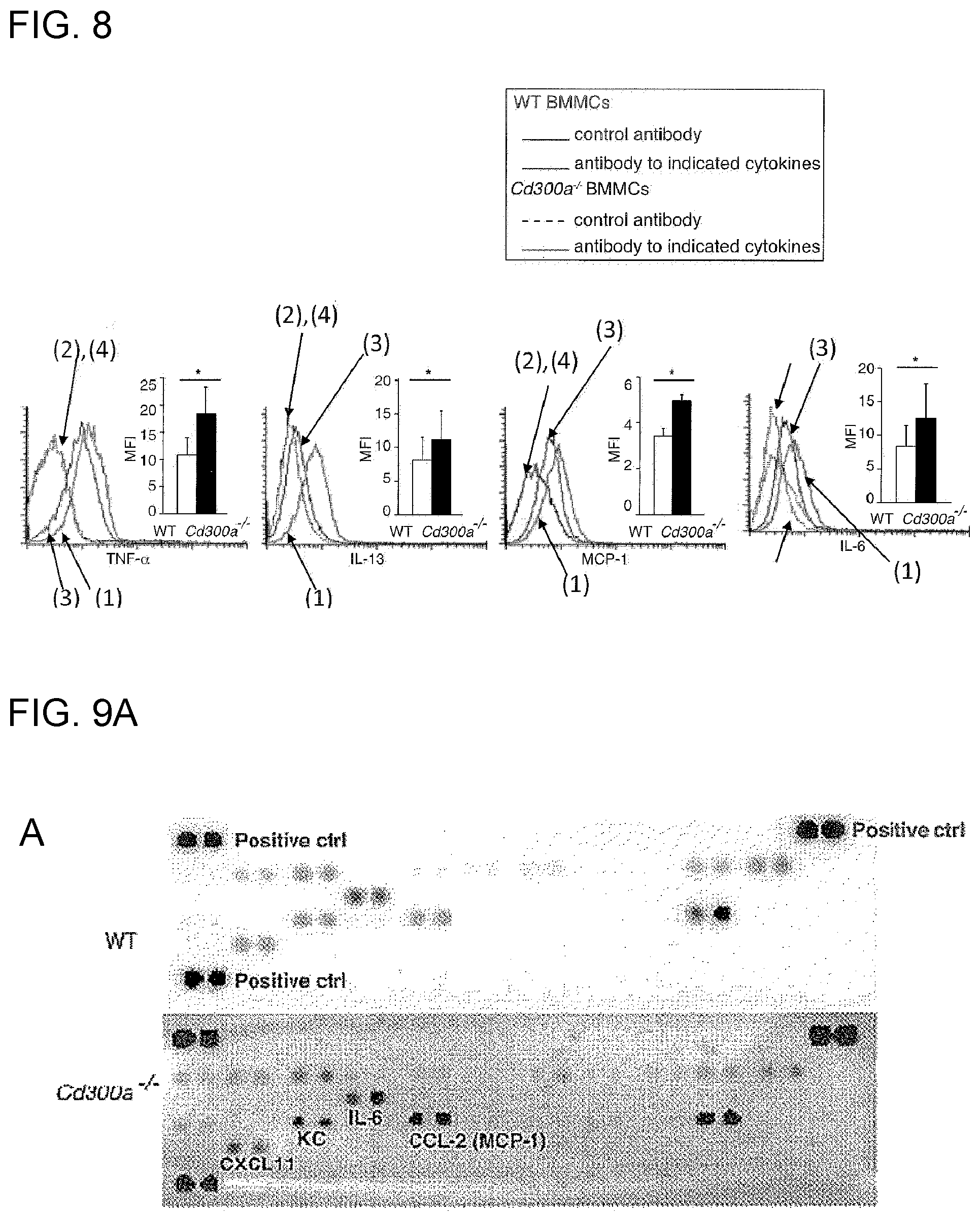
D00014
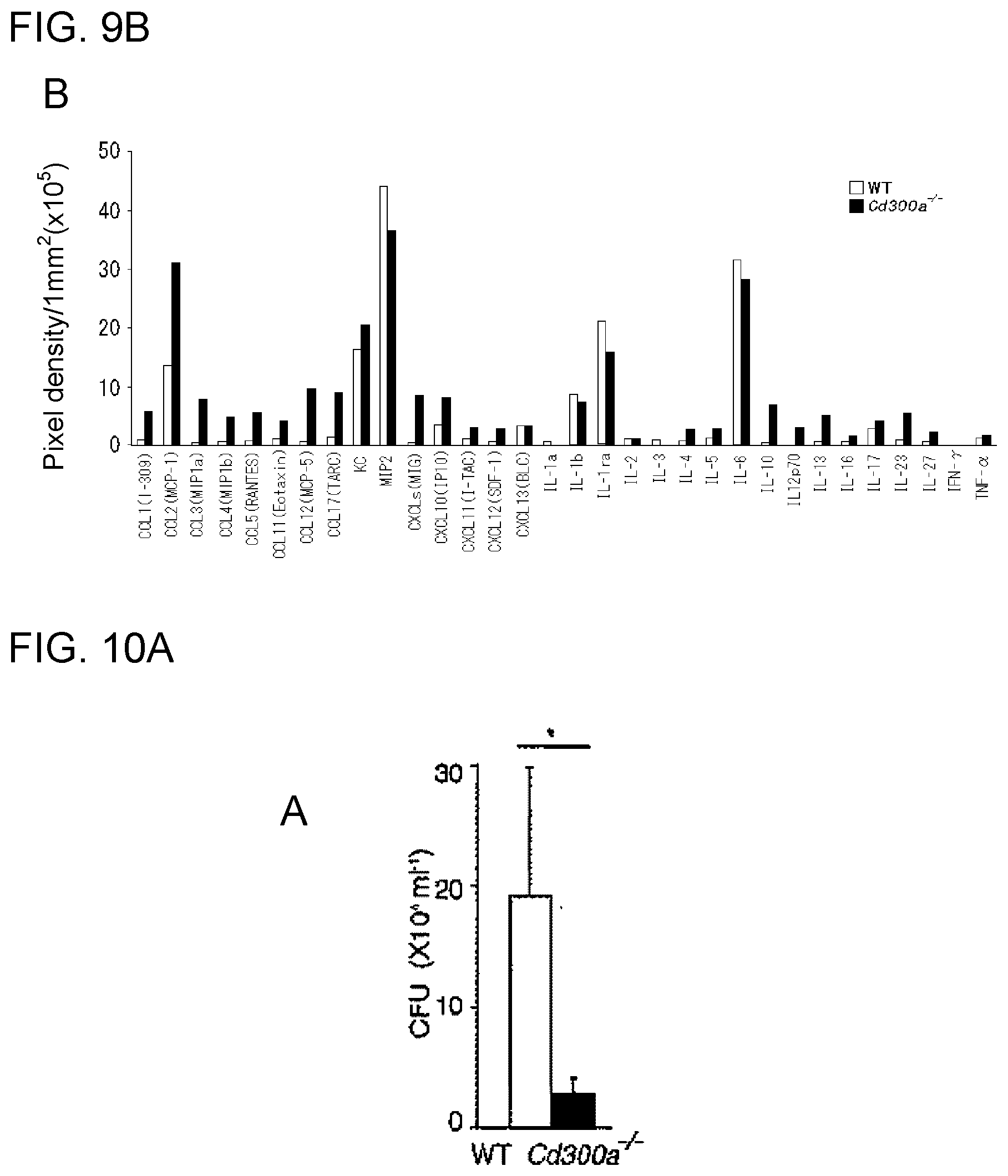
D00015
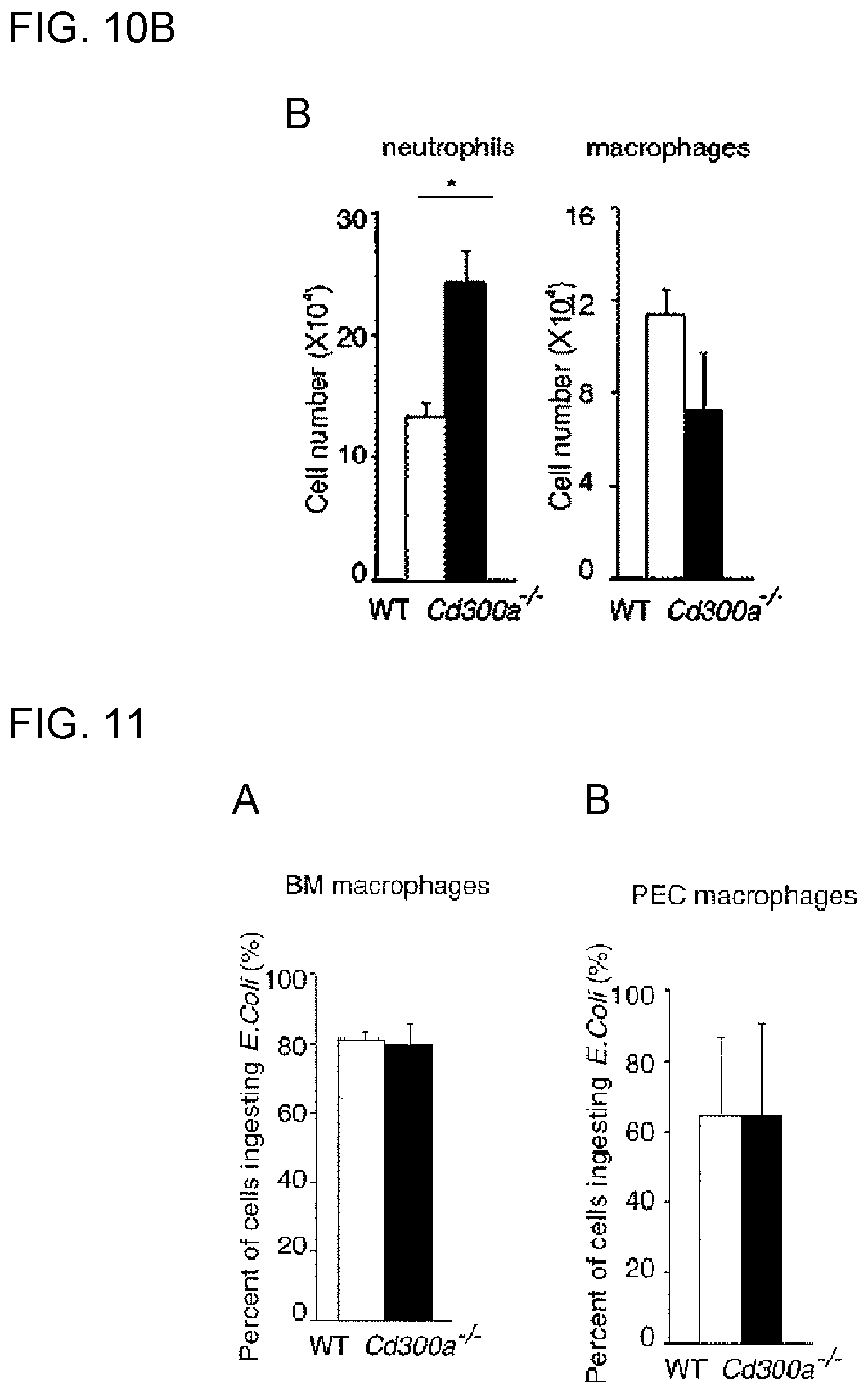
D00016
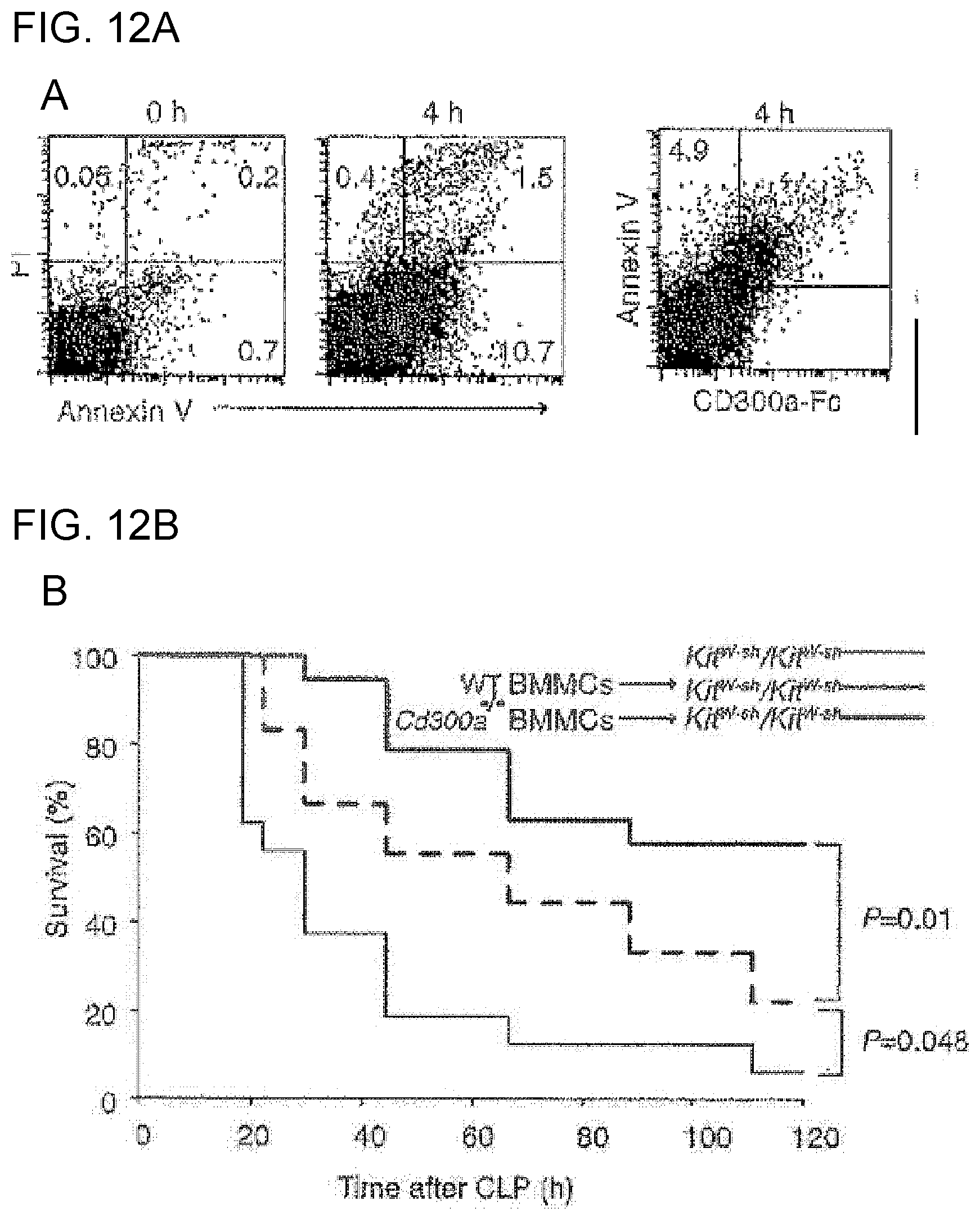
D00017
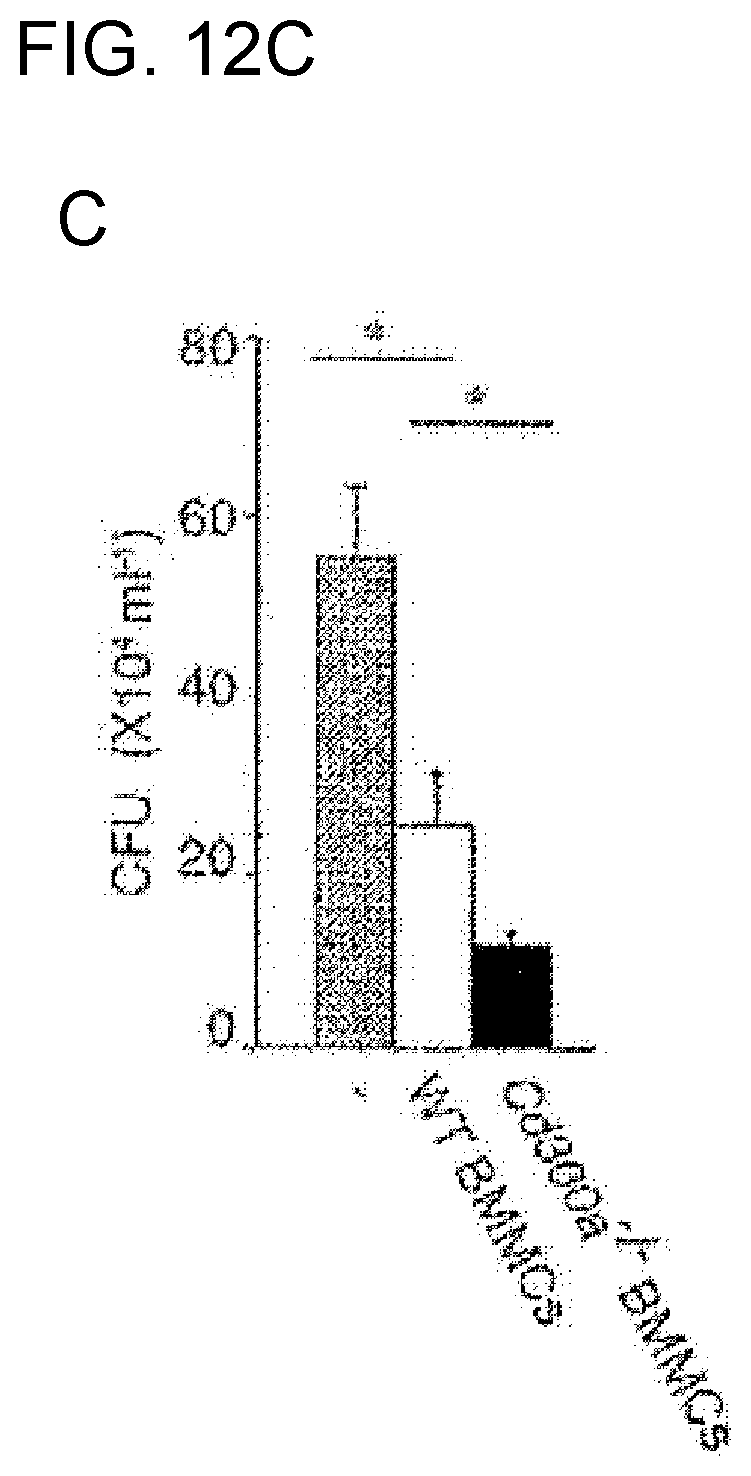
D00018
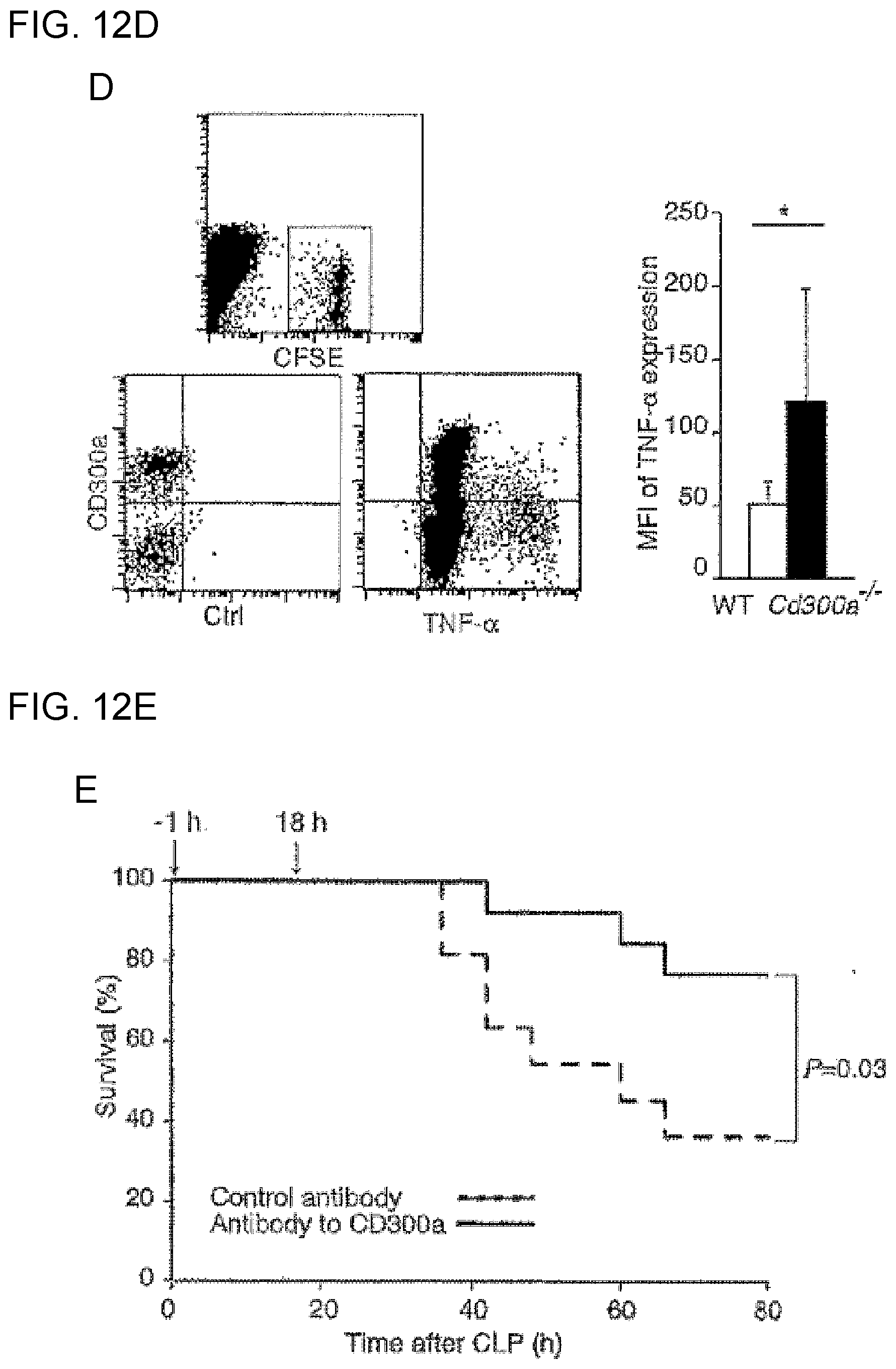
D00019
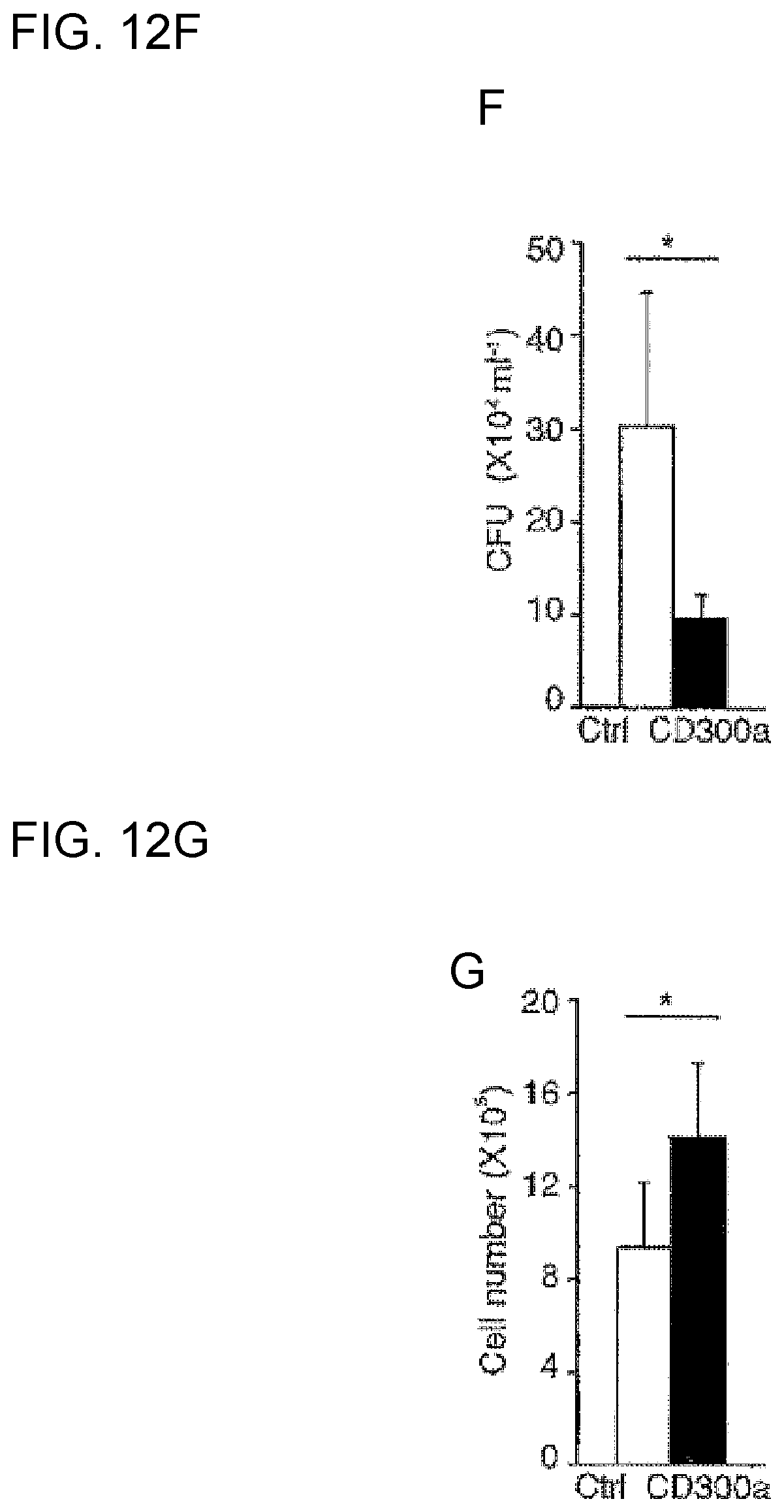
D00020
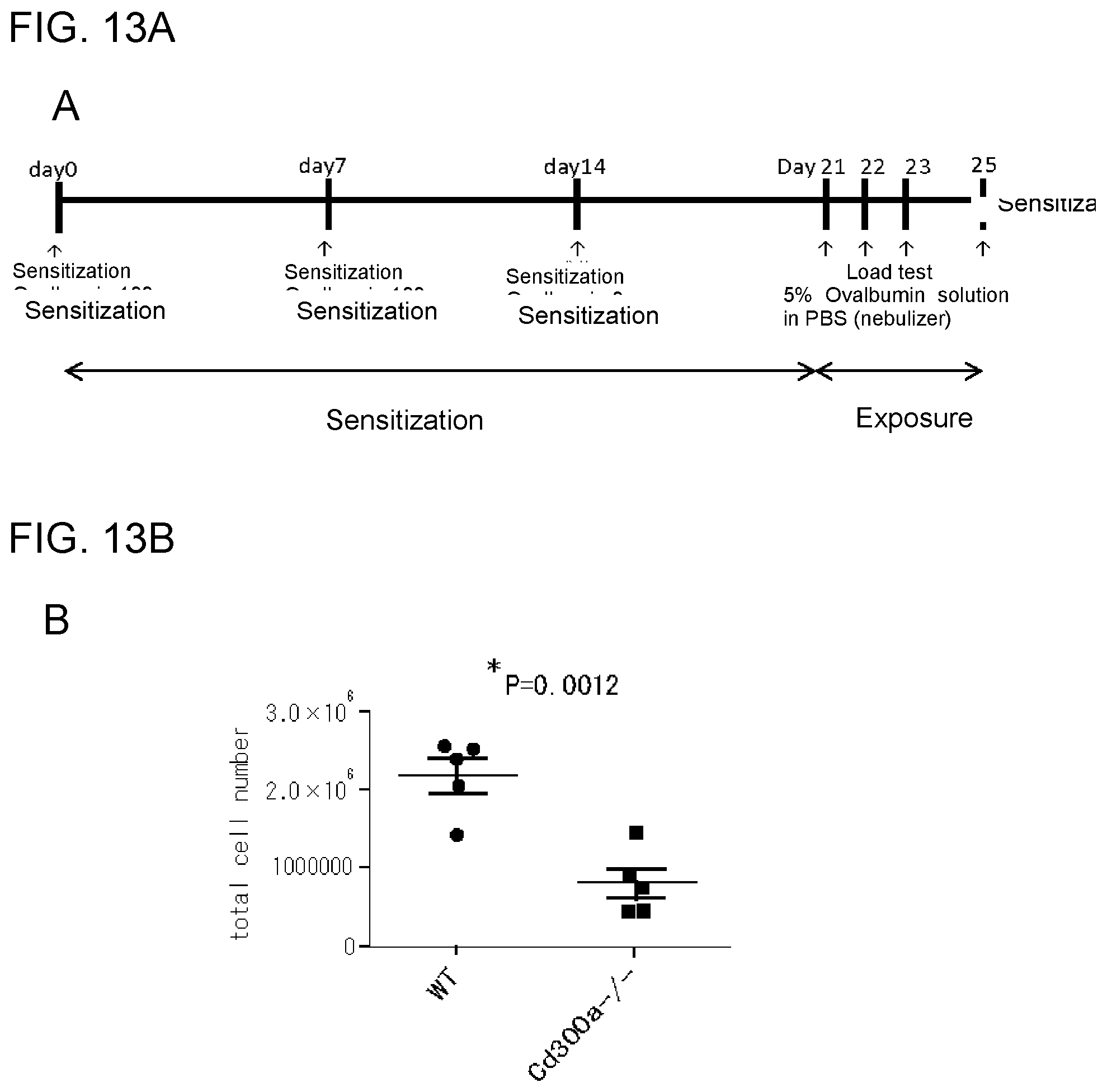
D00021

D00022

D00023
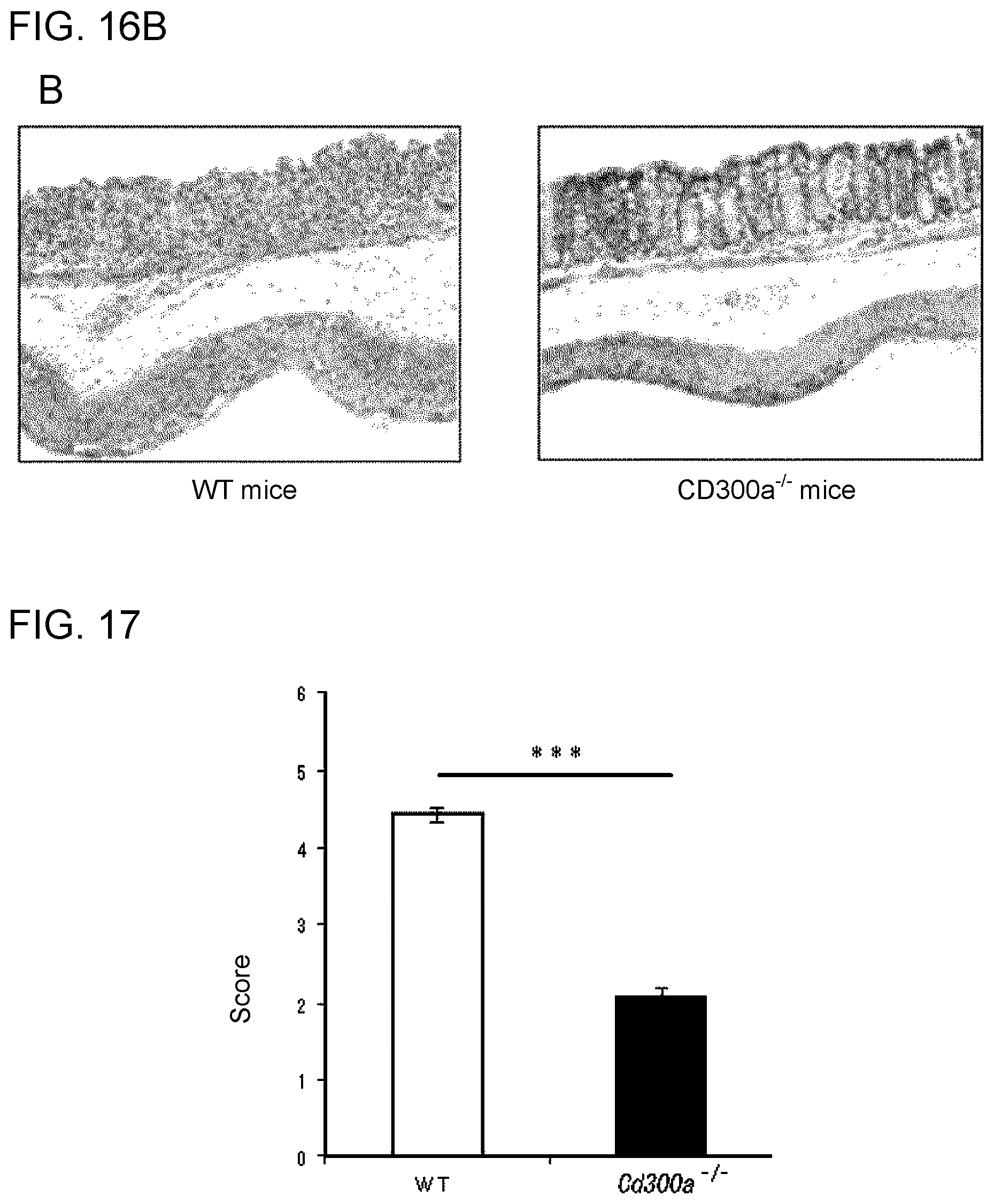
D00024

D00025

D00026
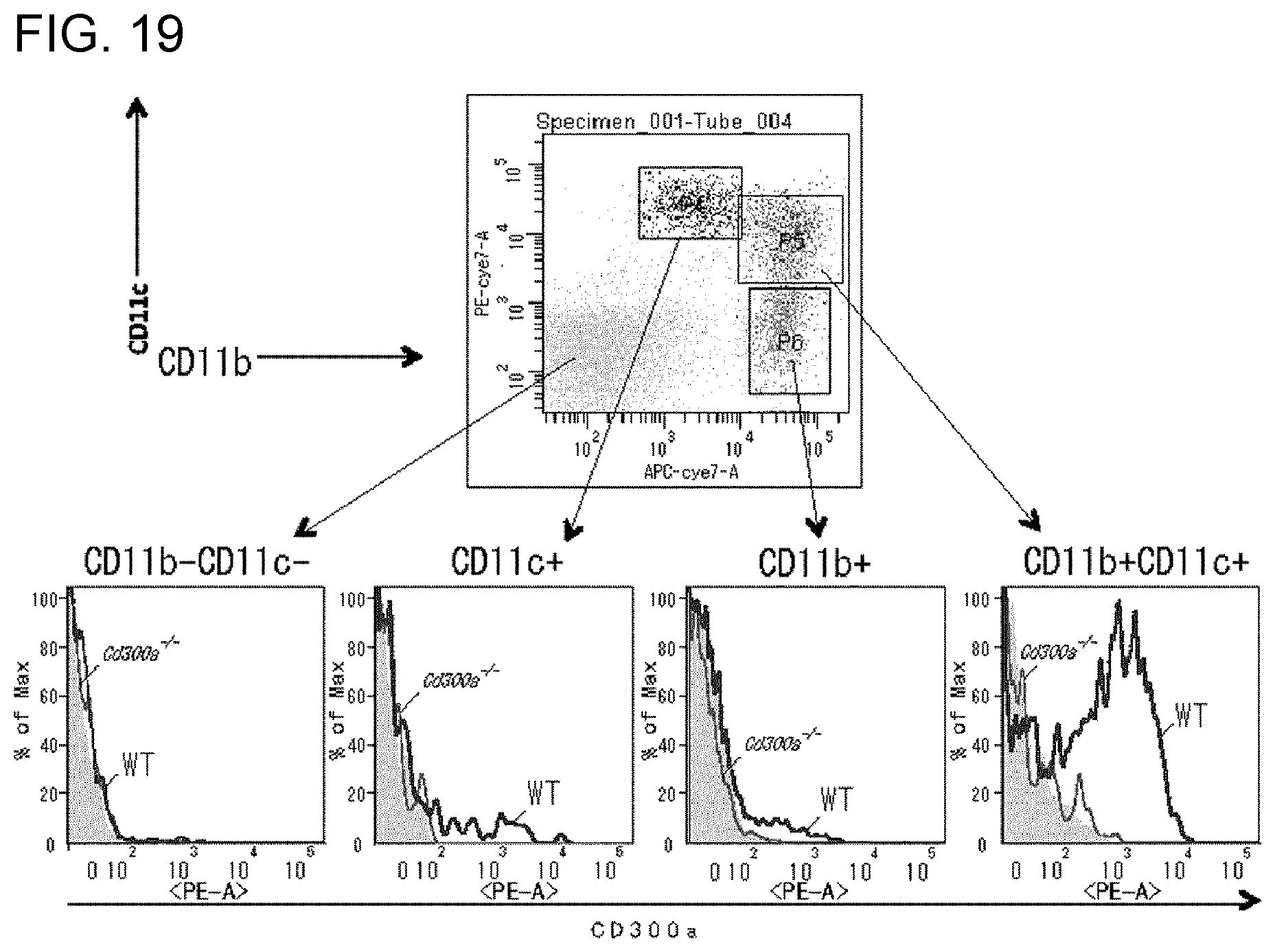
D00027
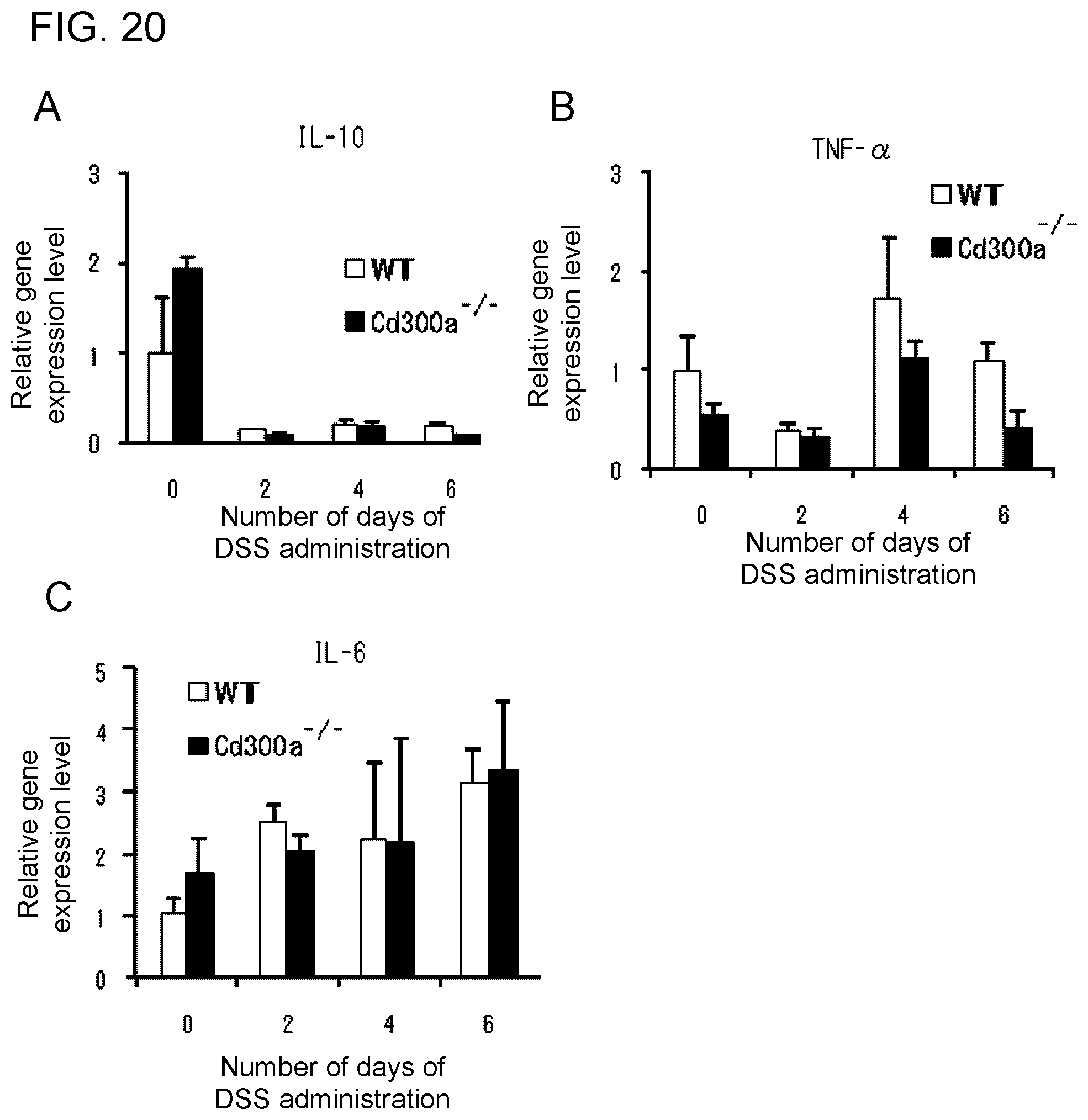
D00028
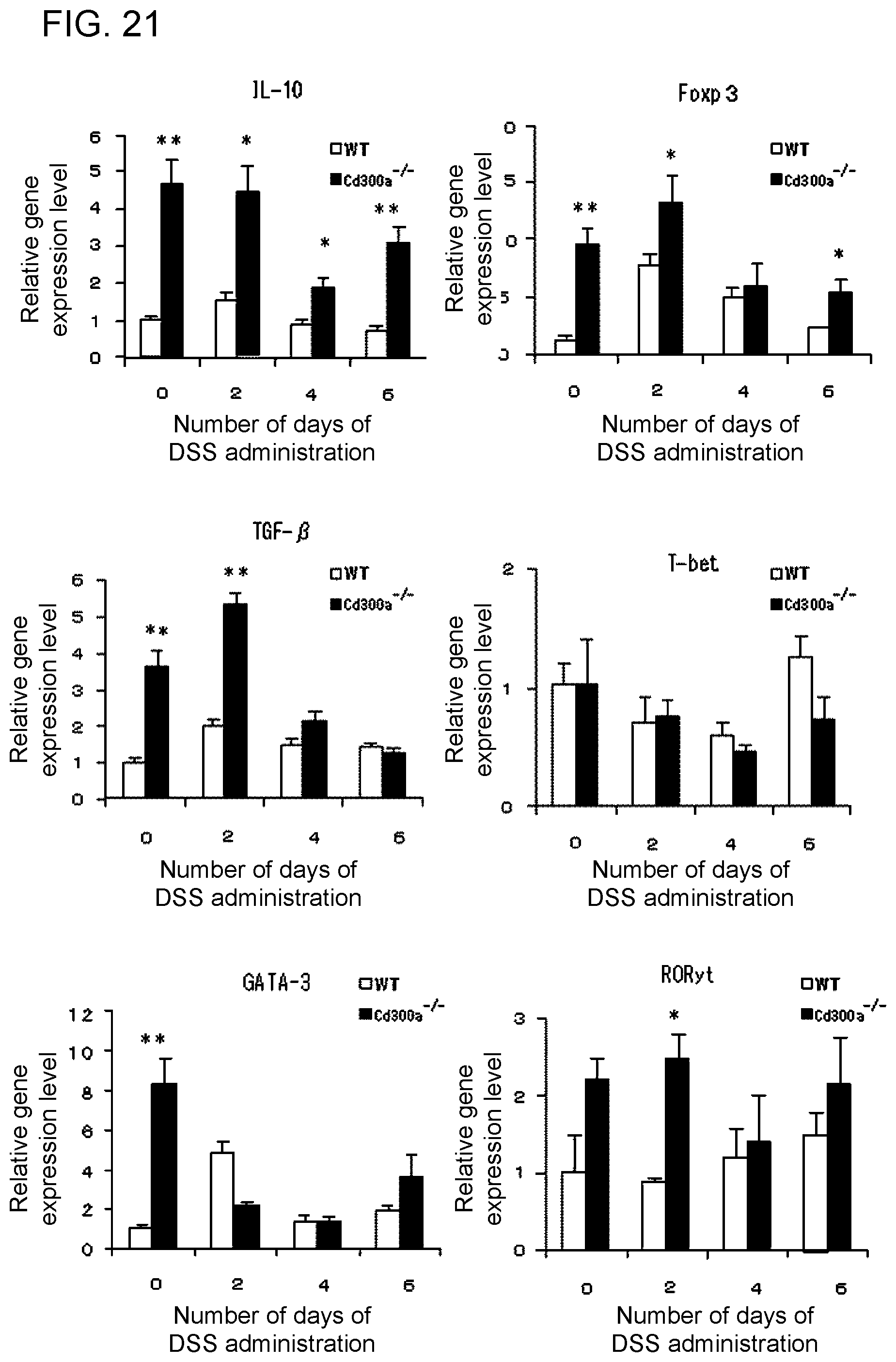
D00029
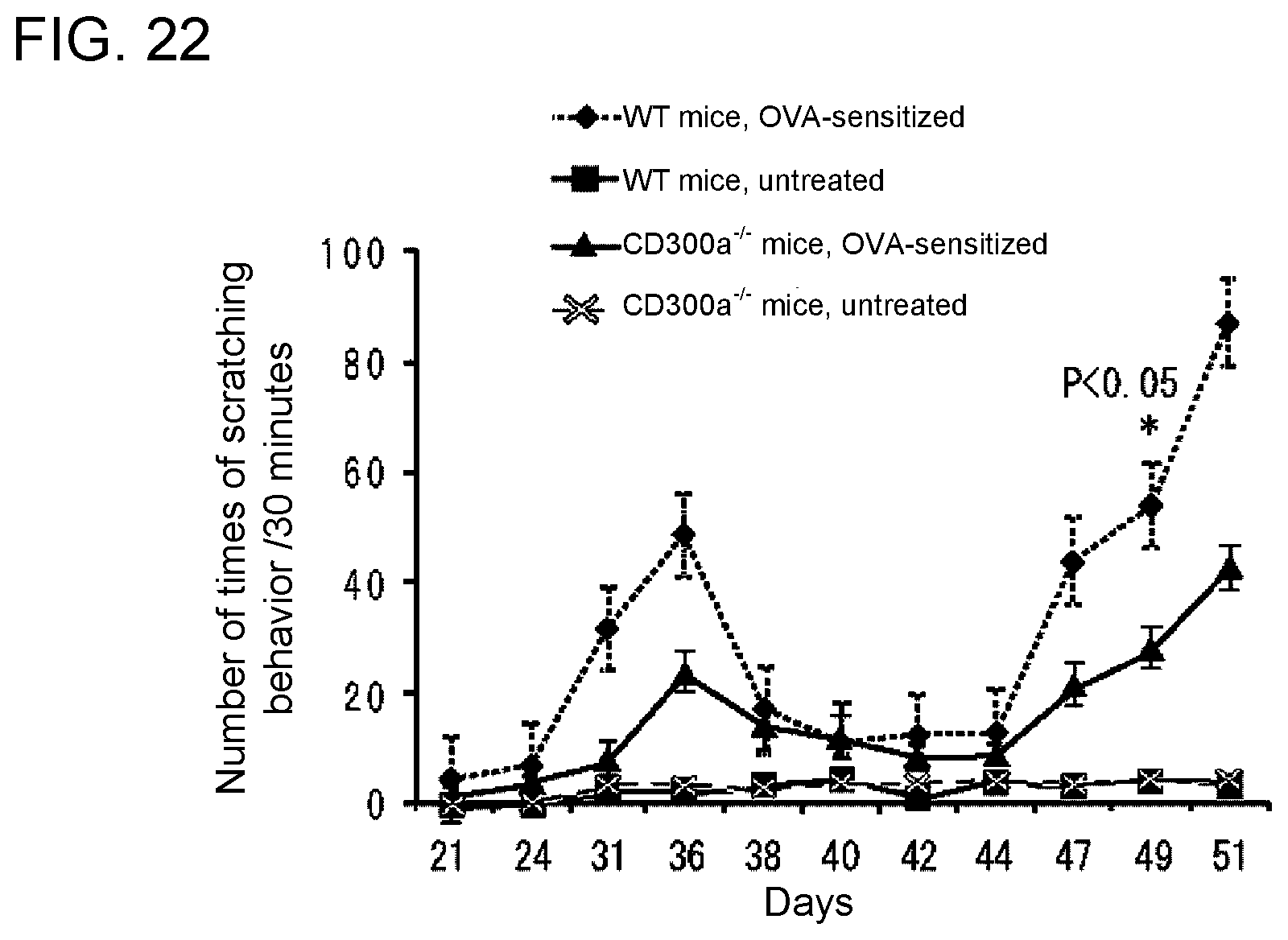
D00030

D00031

D00032
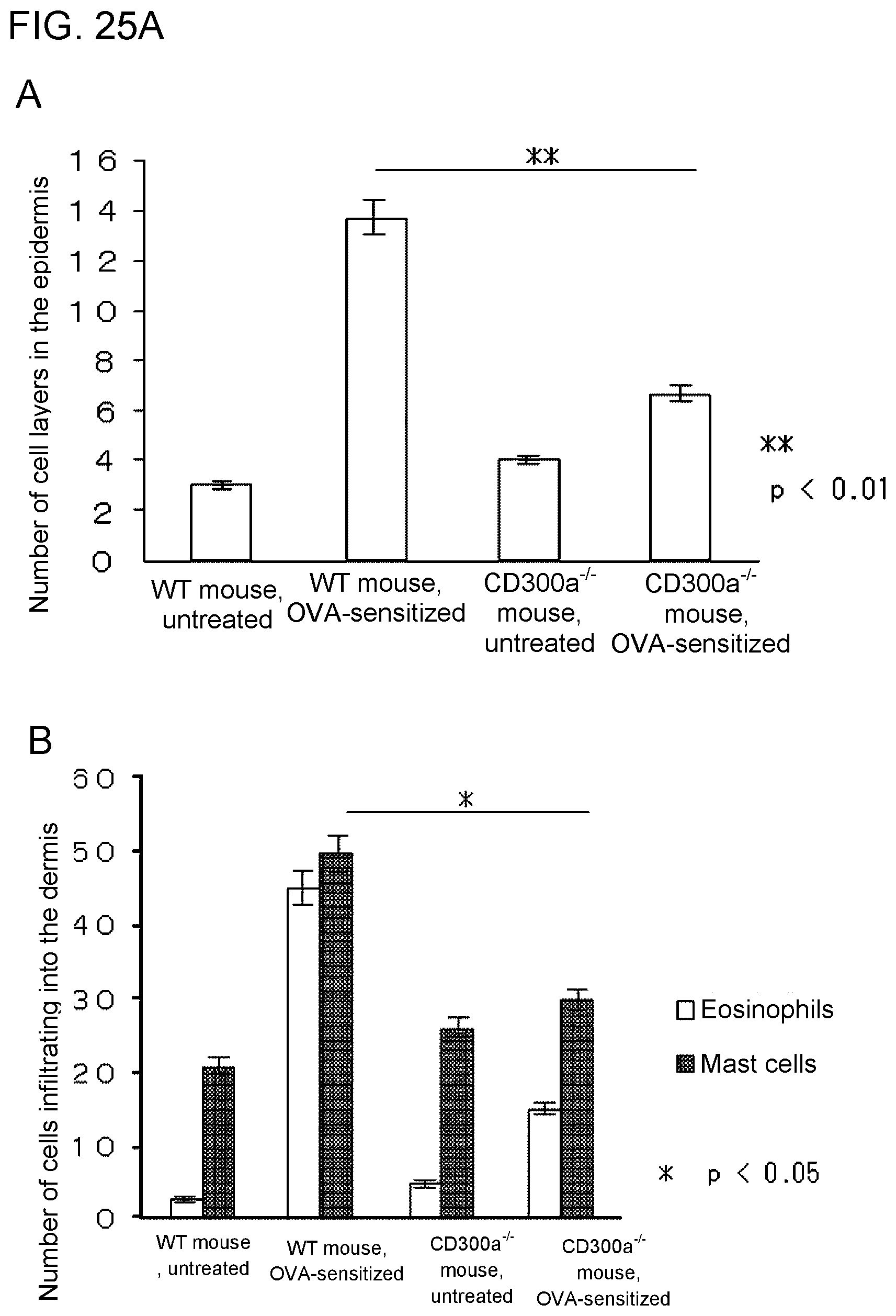
D00033

D00034
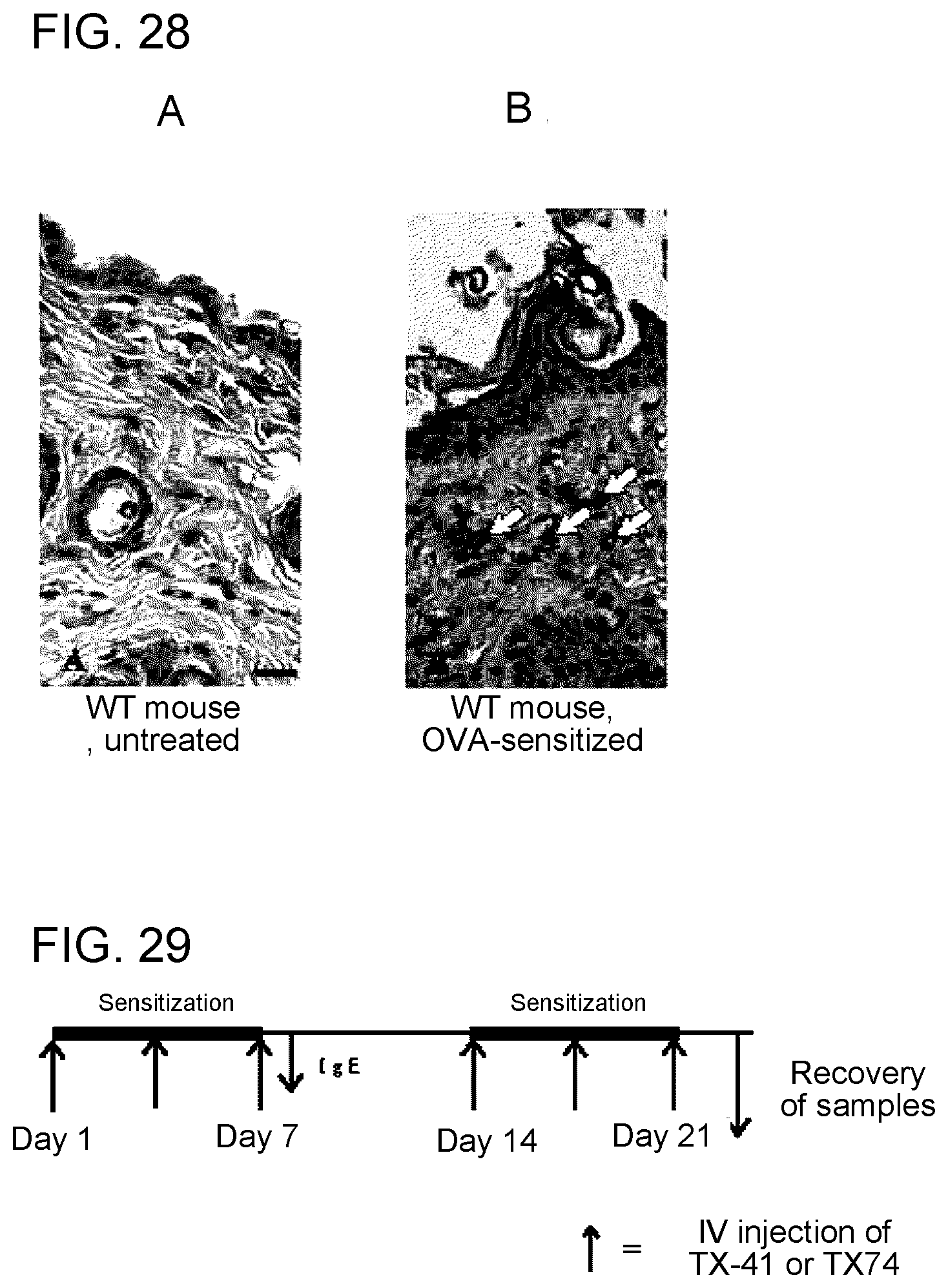
D00035
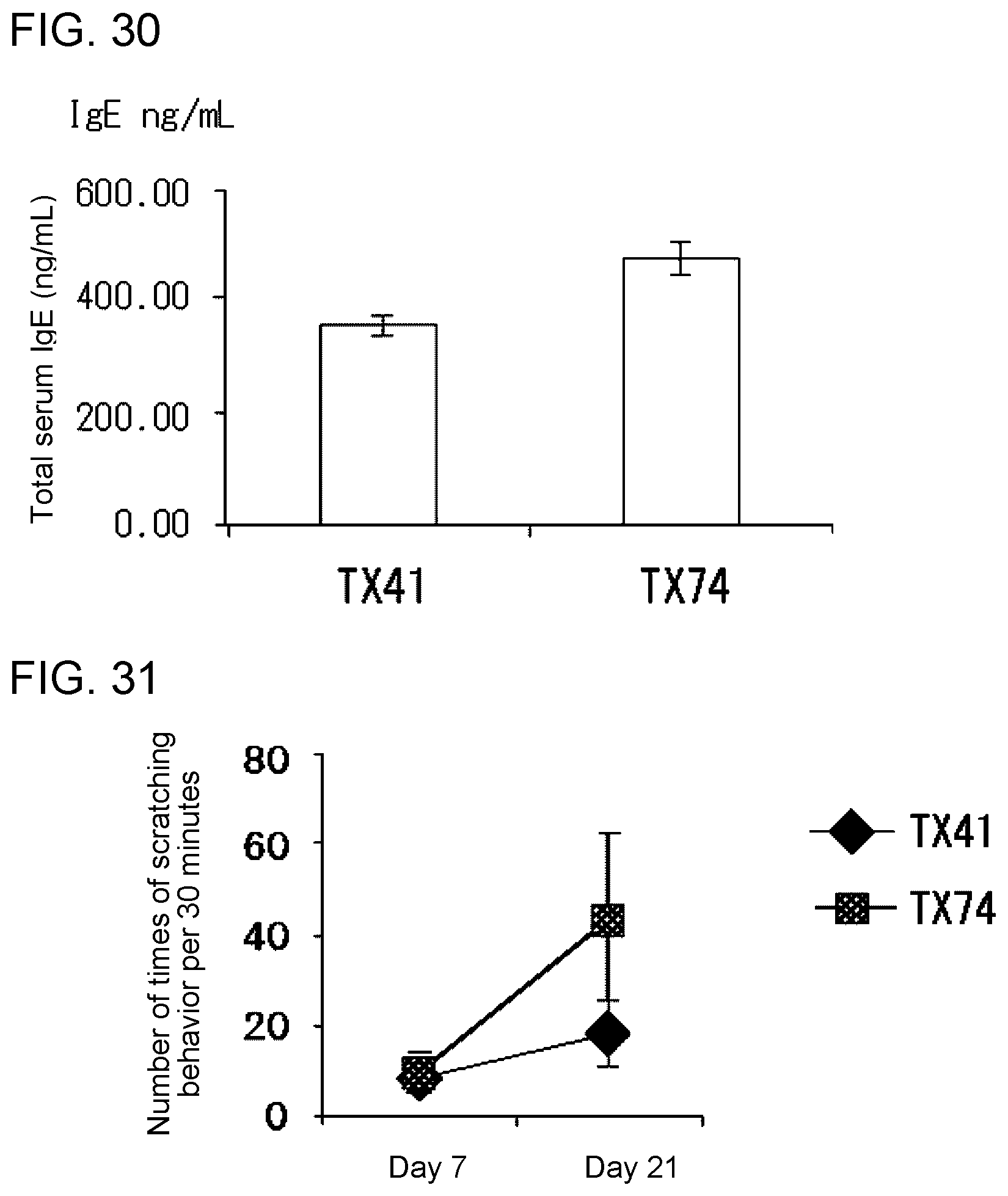
D00036
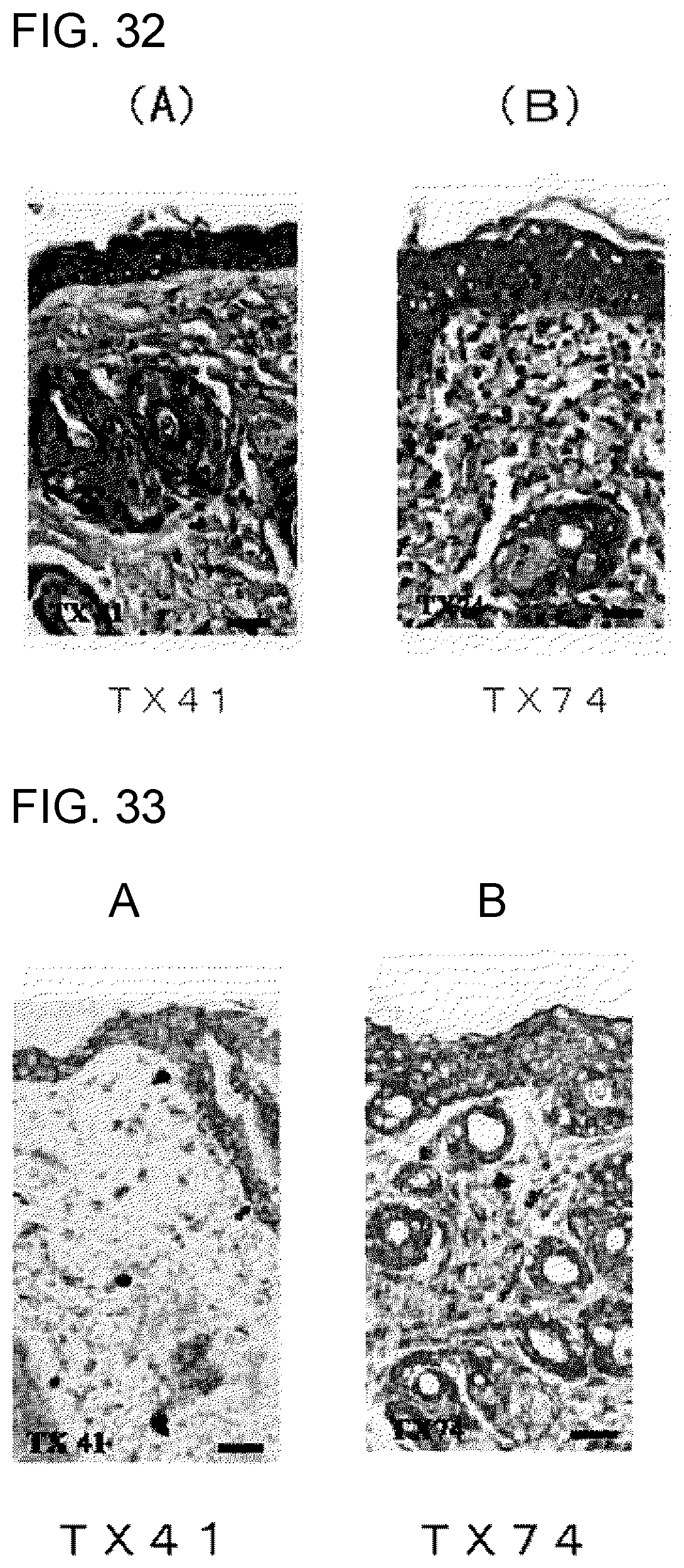
D00037
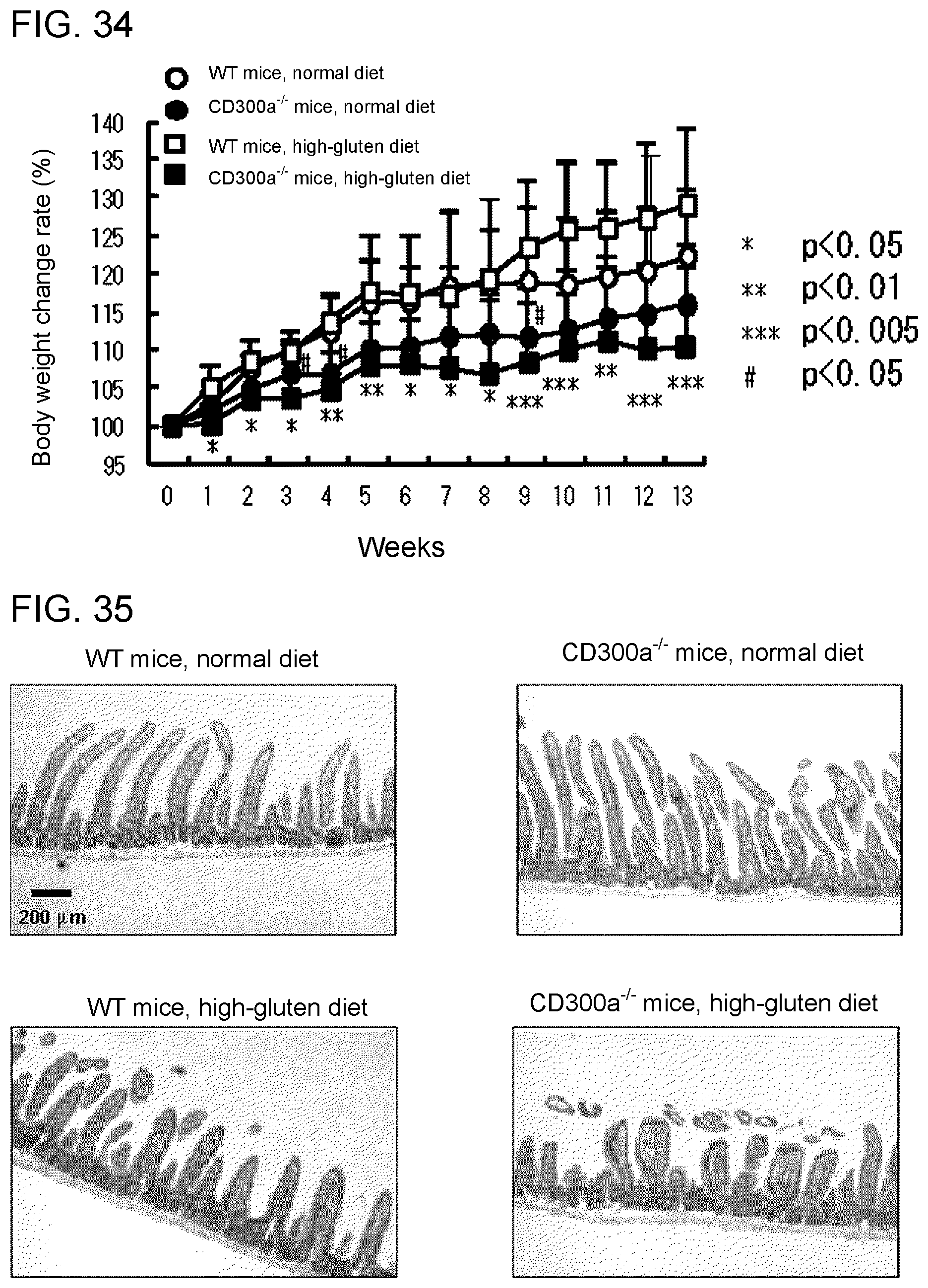
D00038
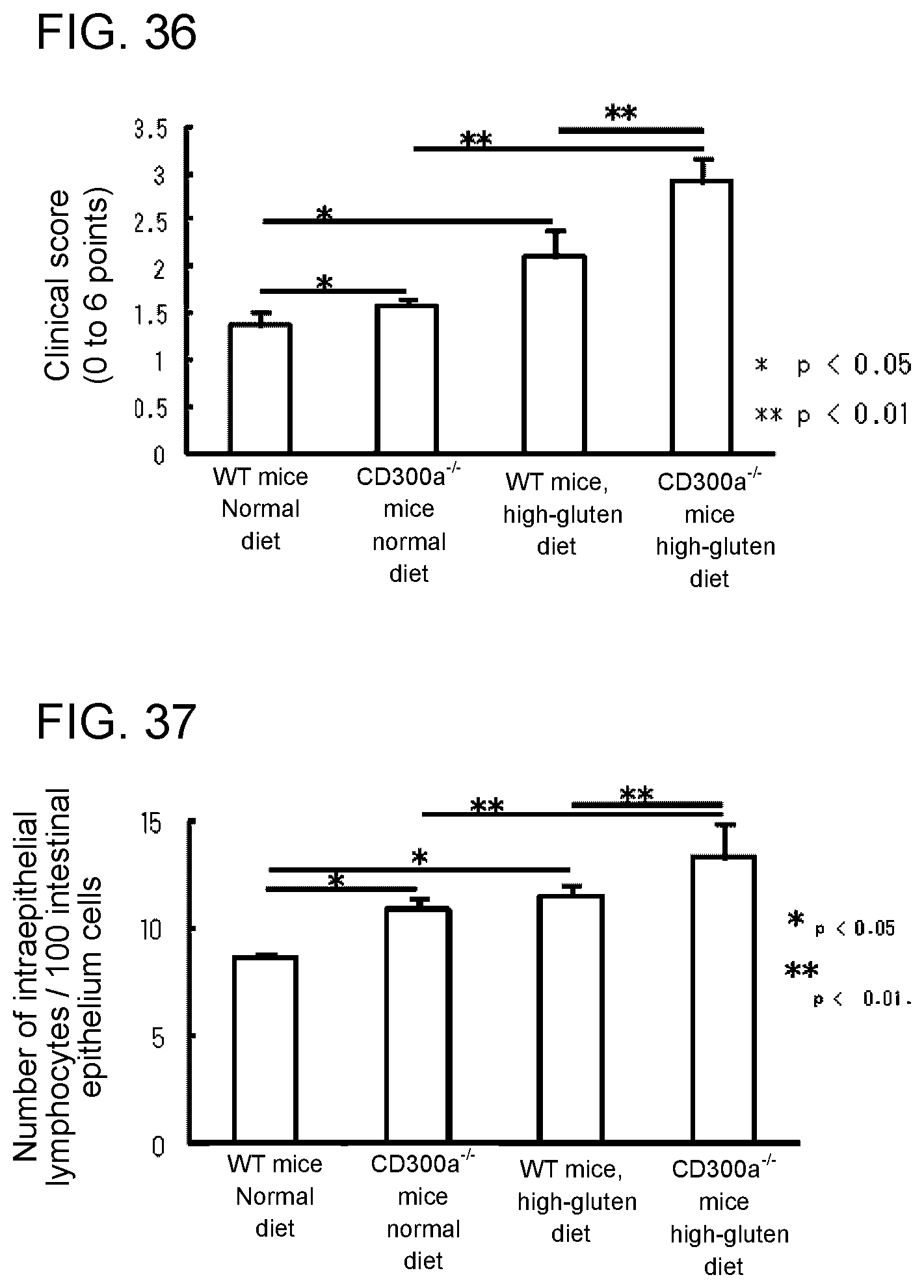
D00039

D00040
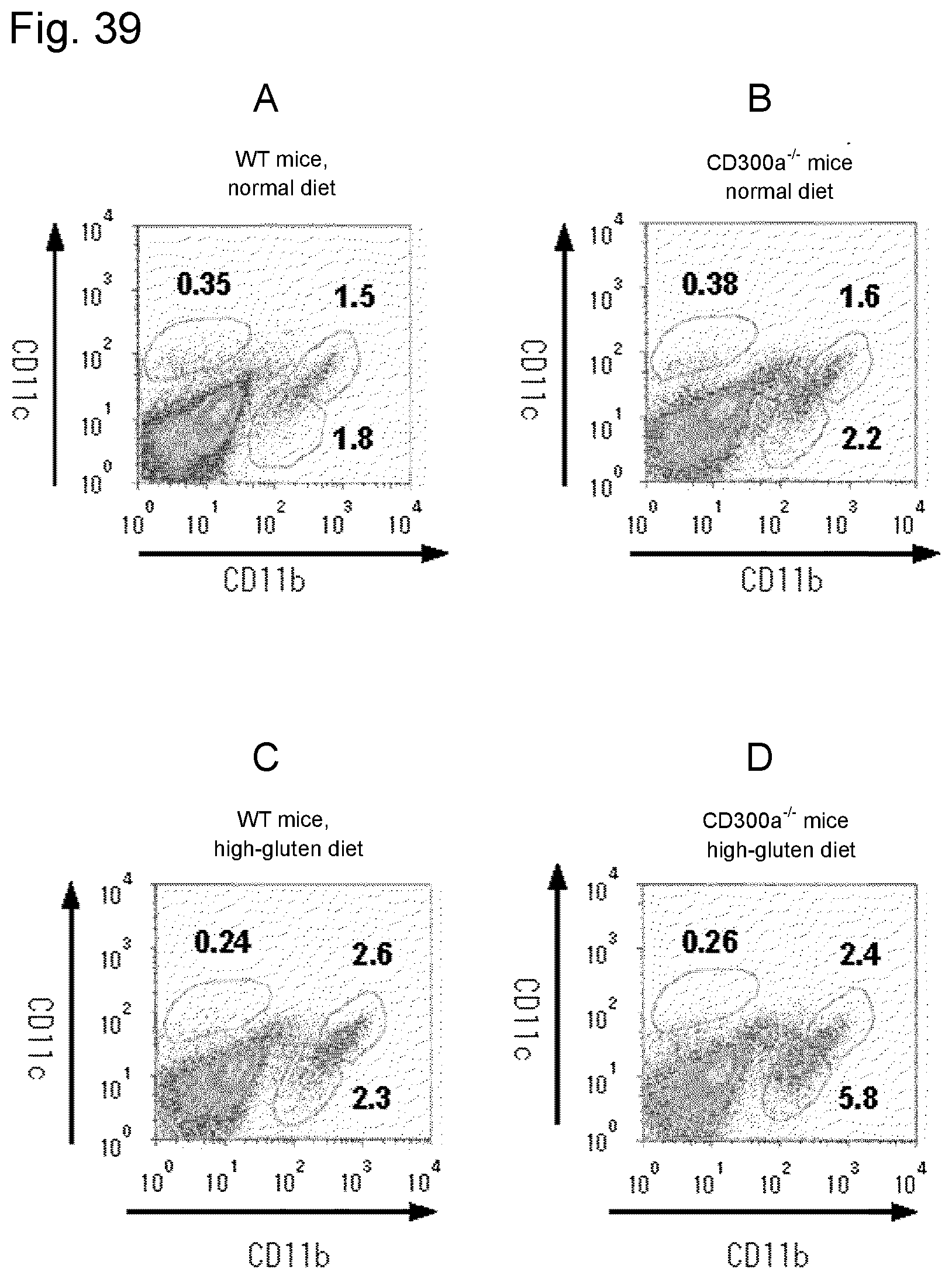
D00041

D00042
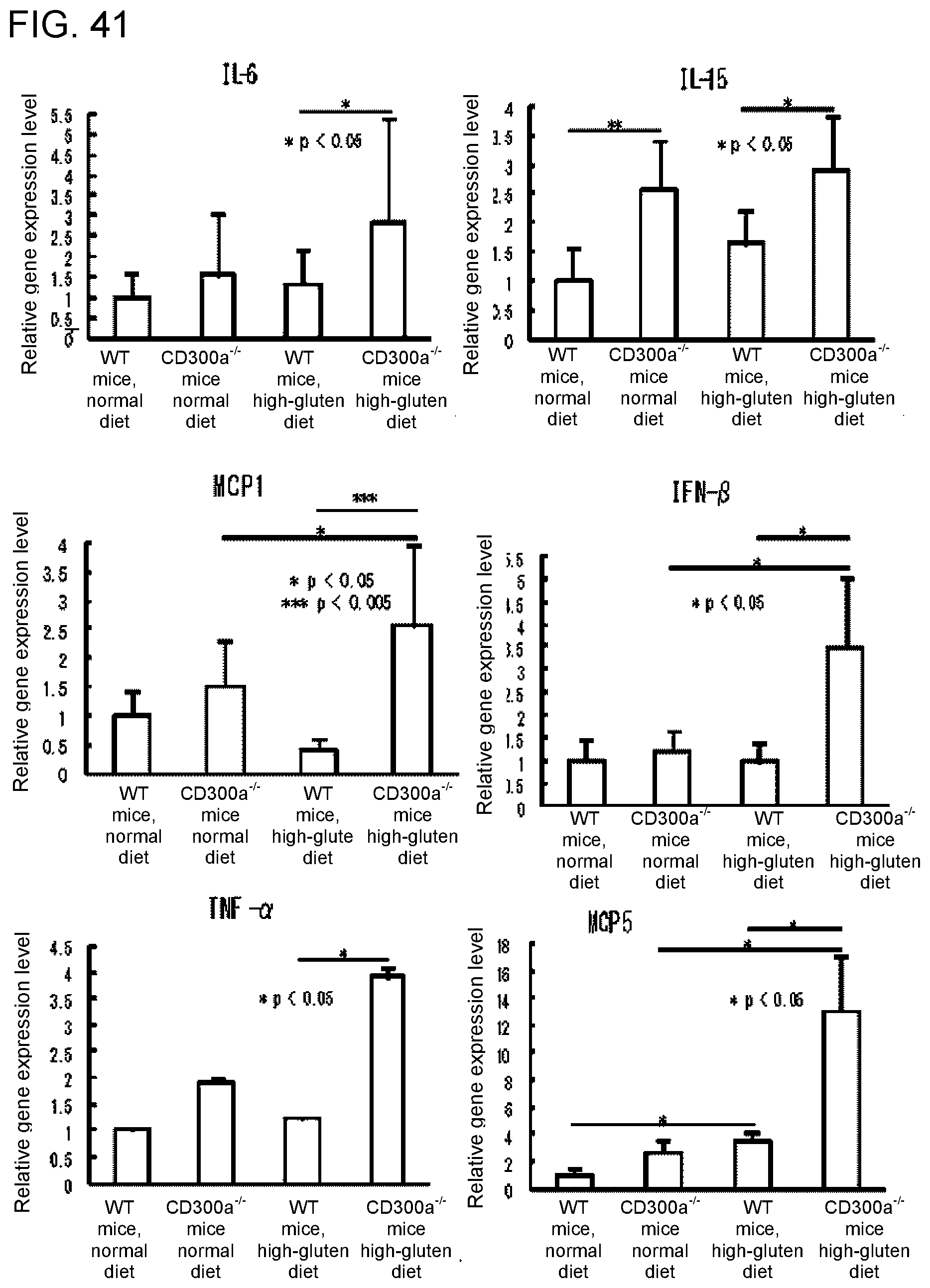
D00043
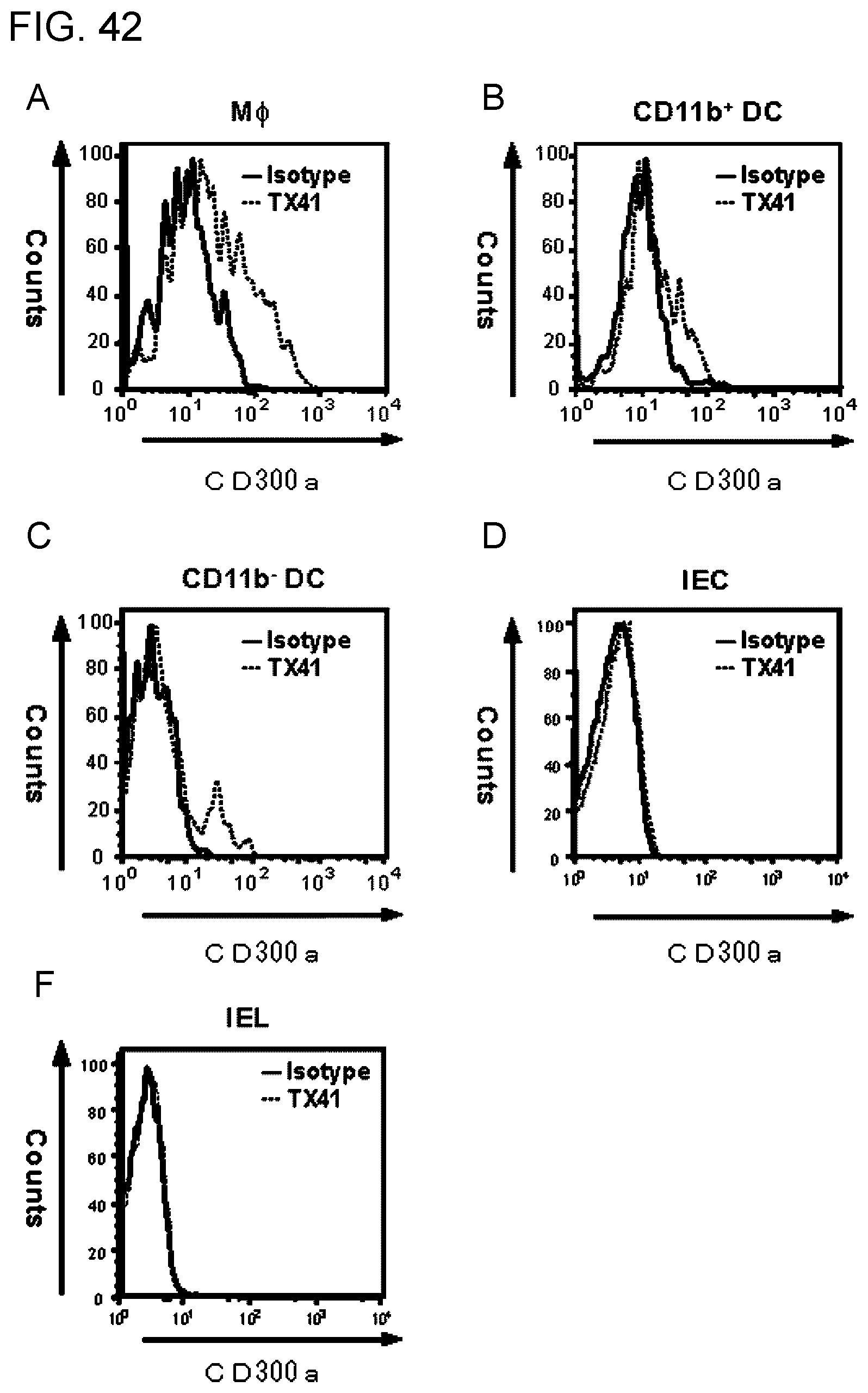
D00044
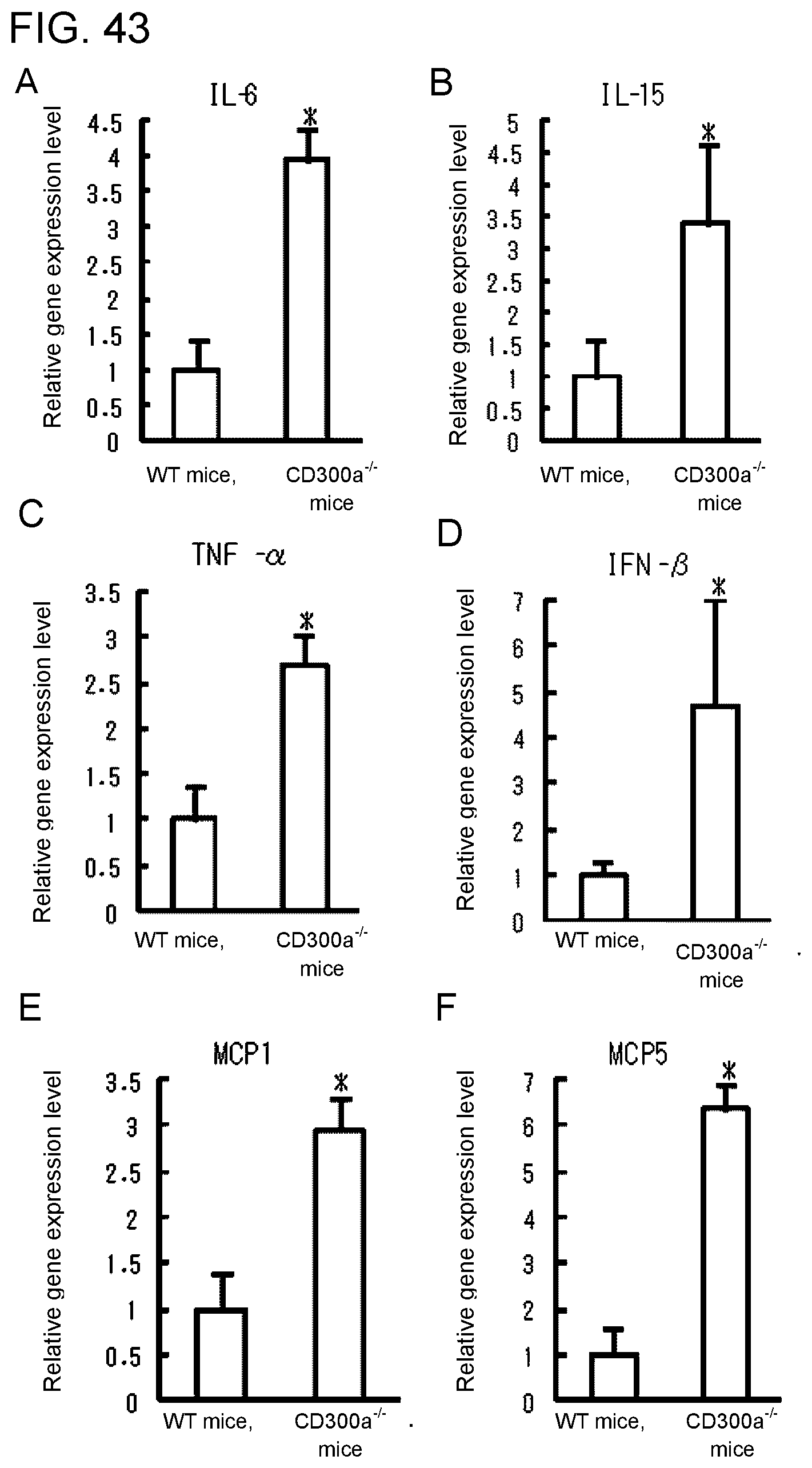
D00045
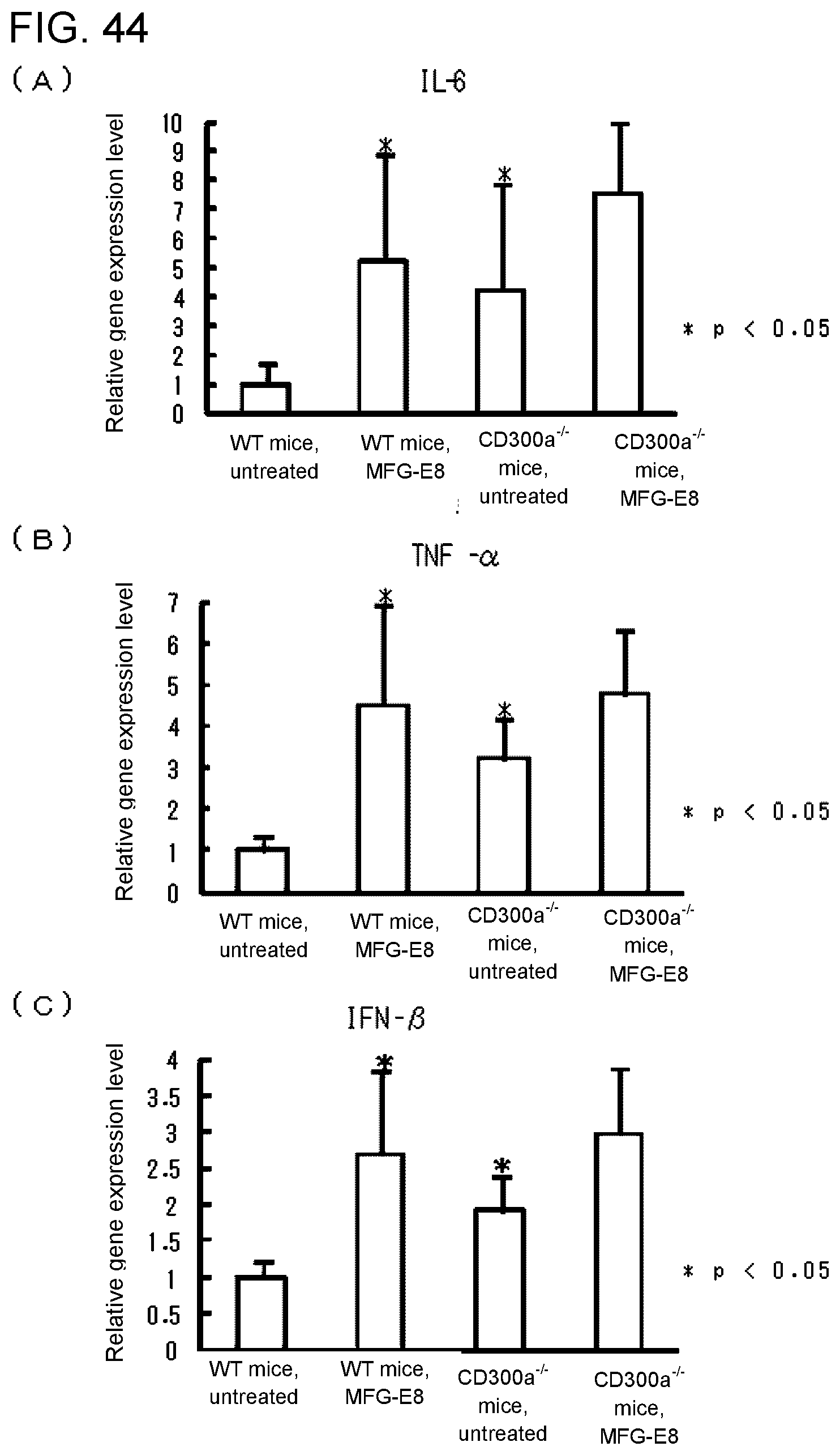
D00046

D00047
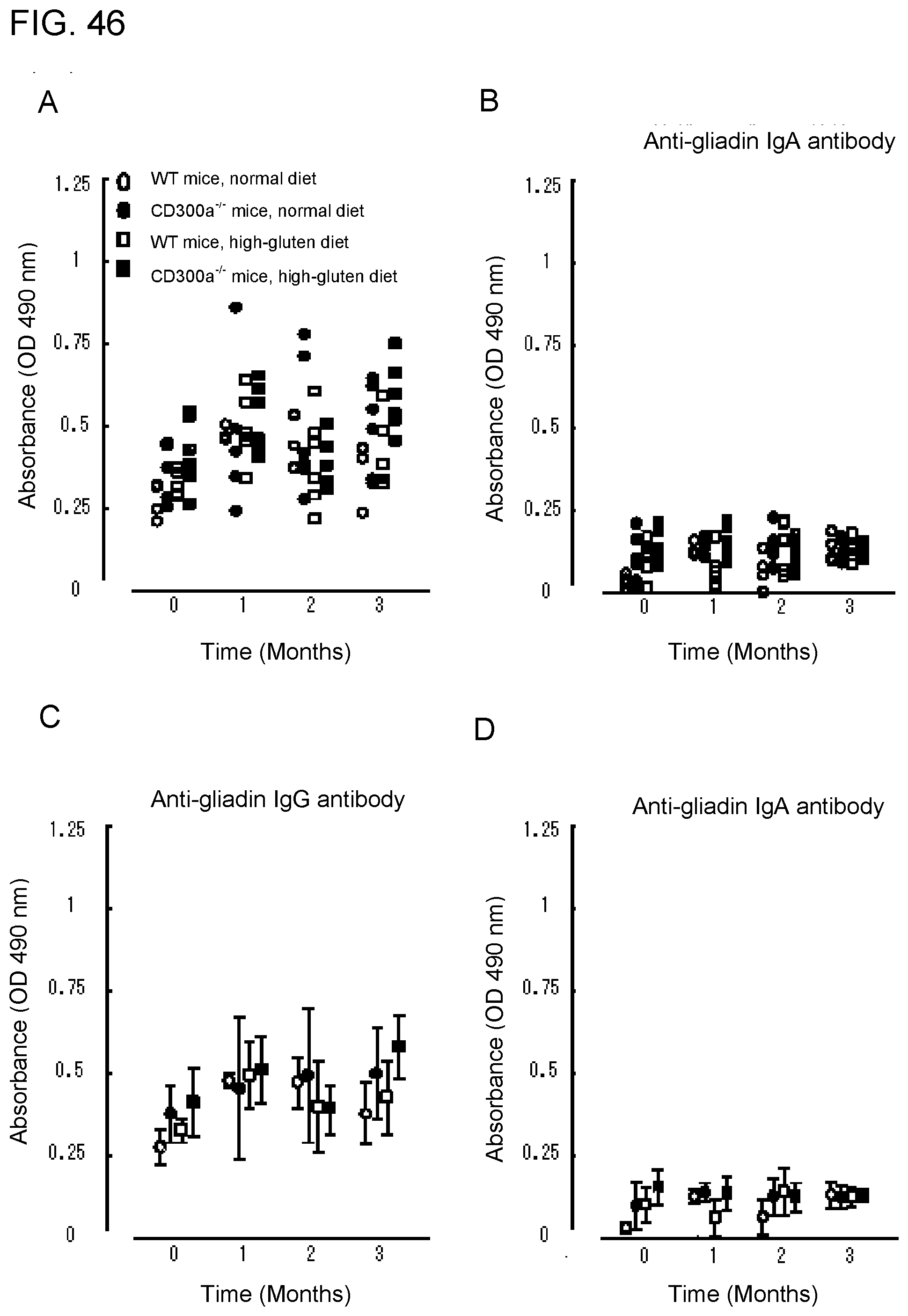
D00048

D00049
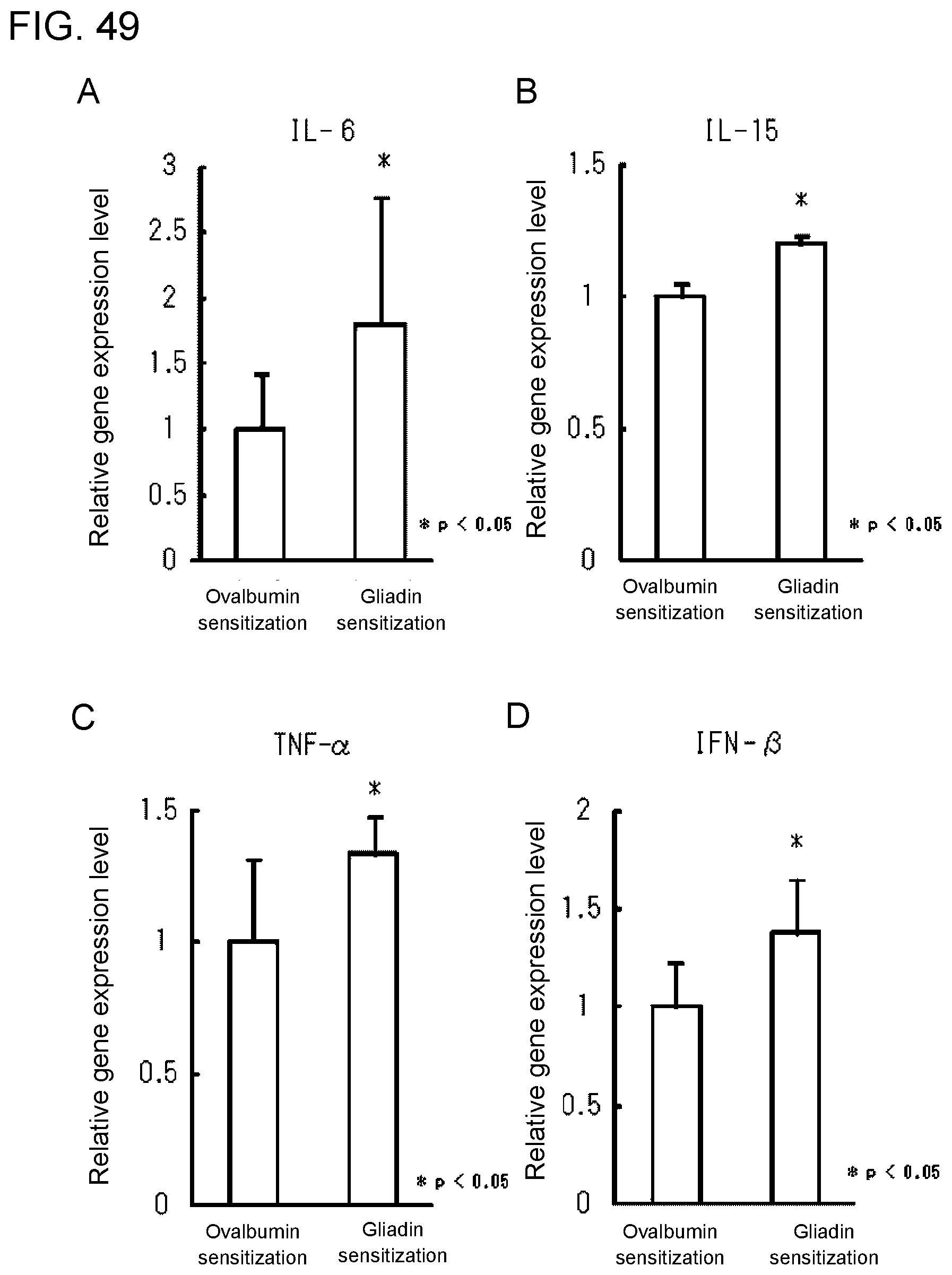
D00050
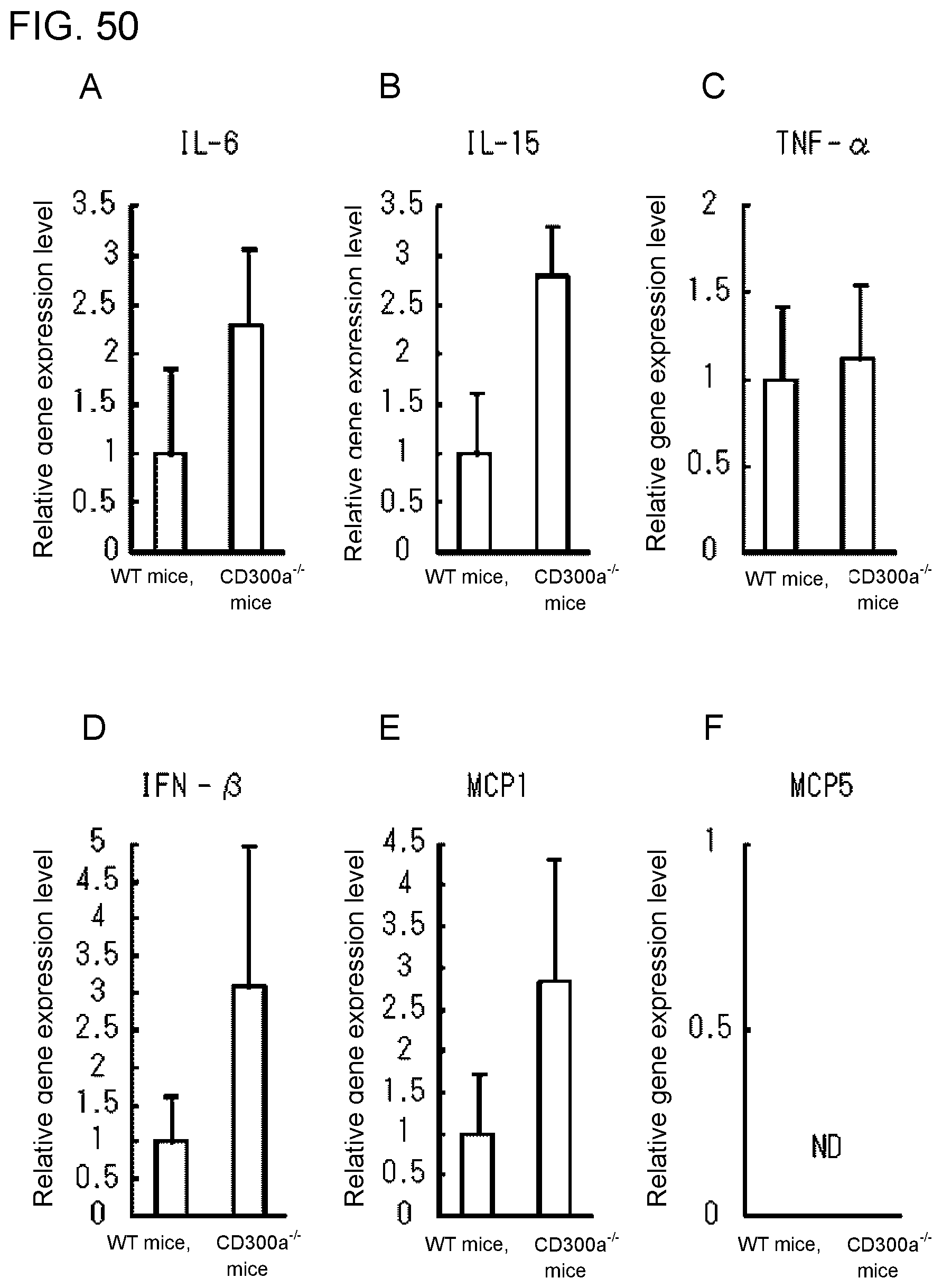
D00051
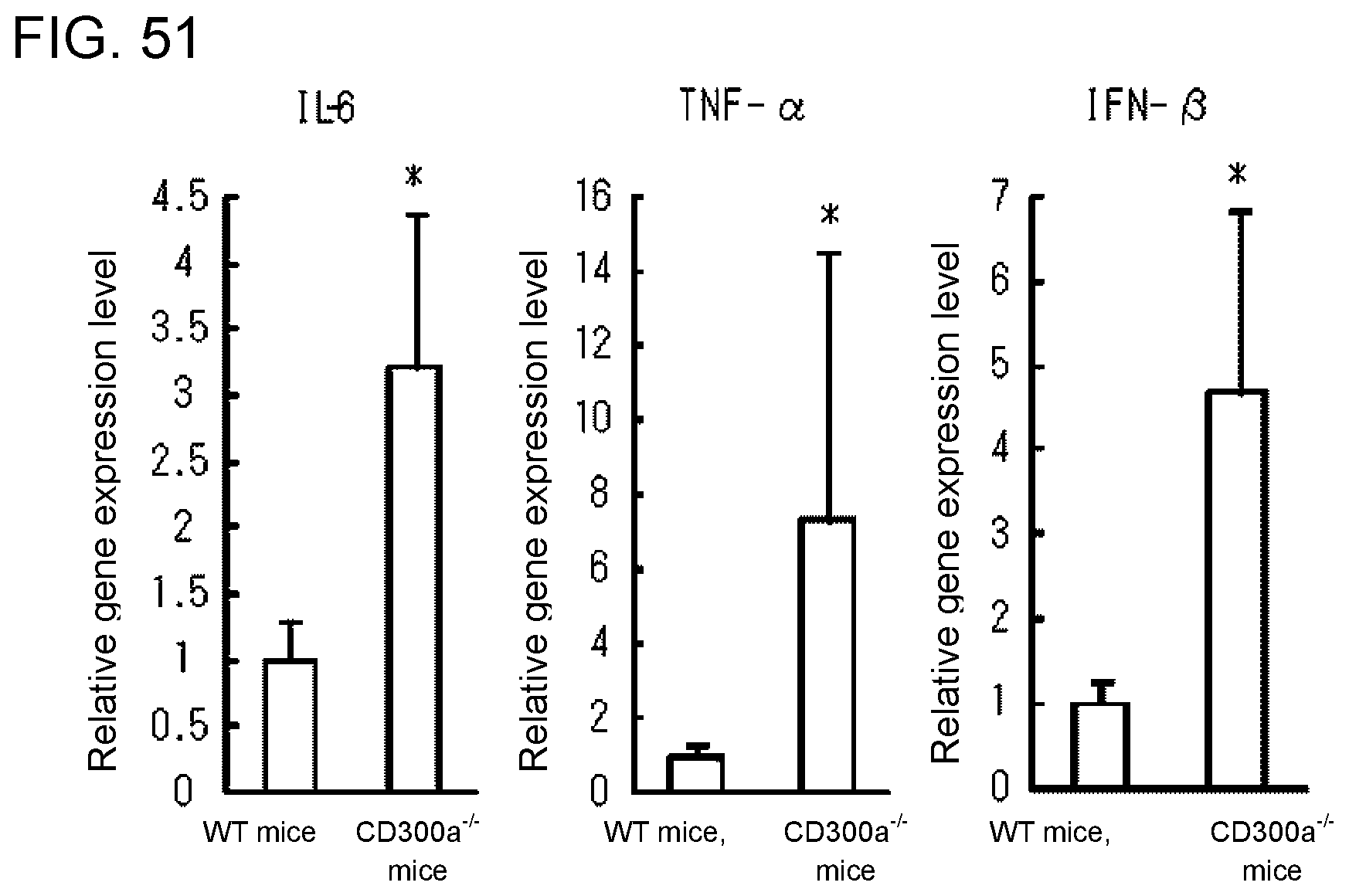
D00052
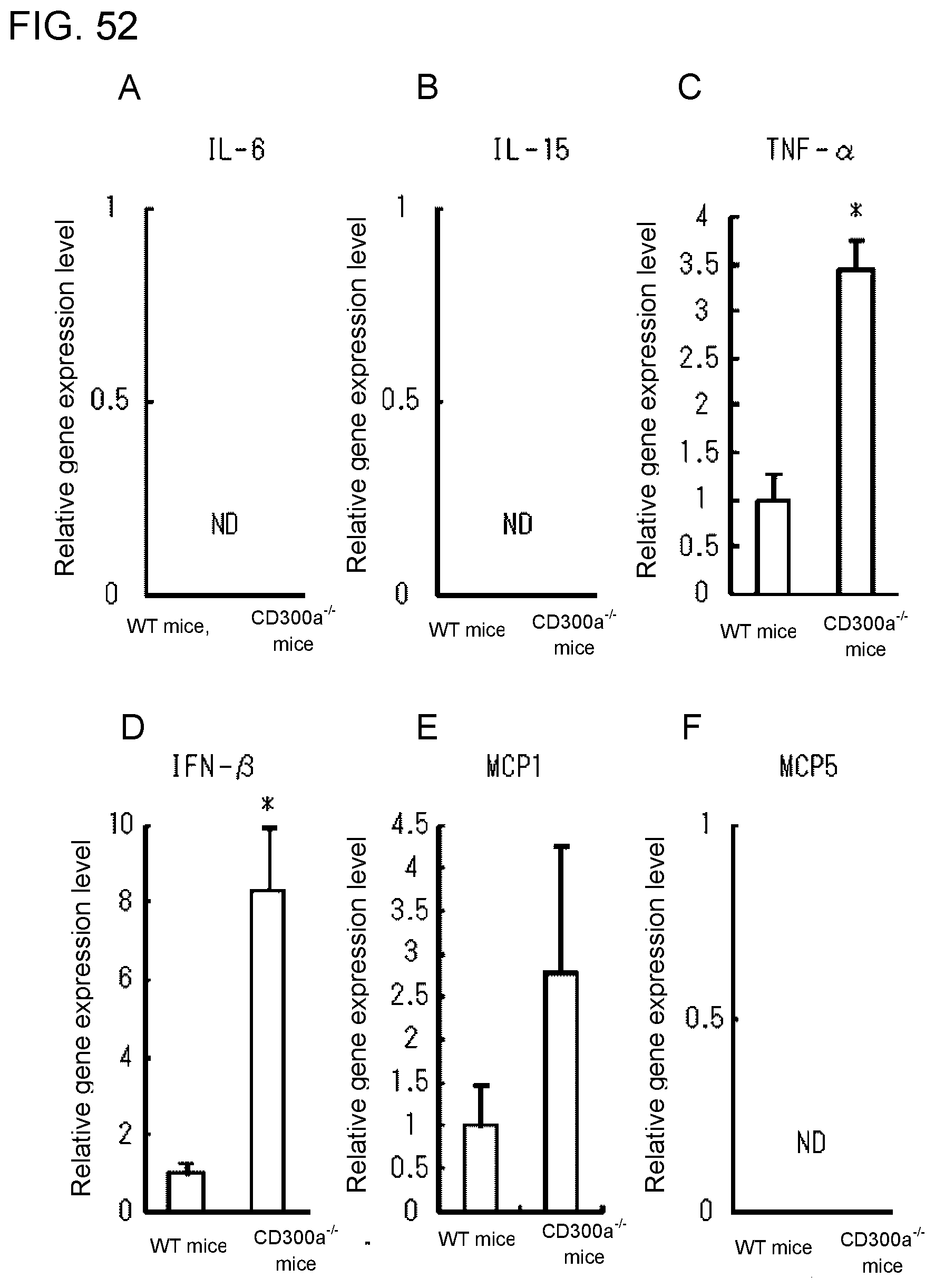
D00053
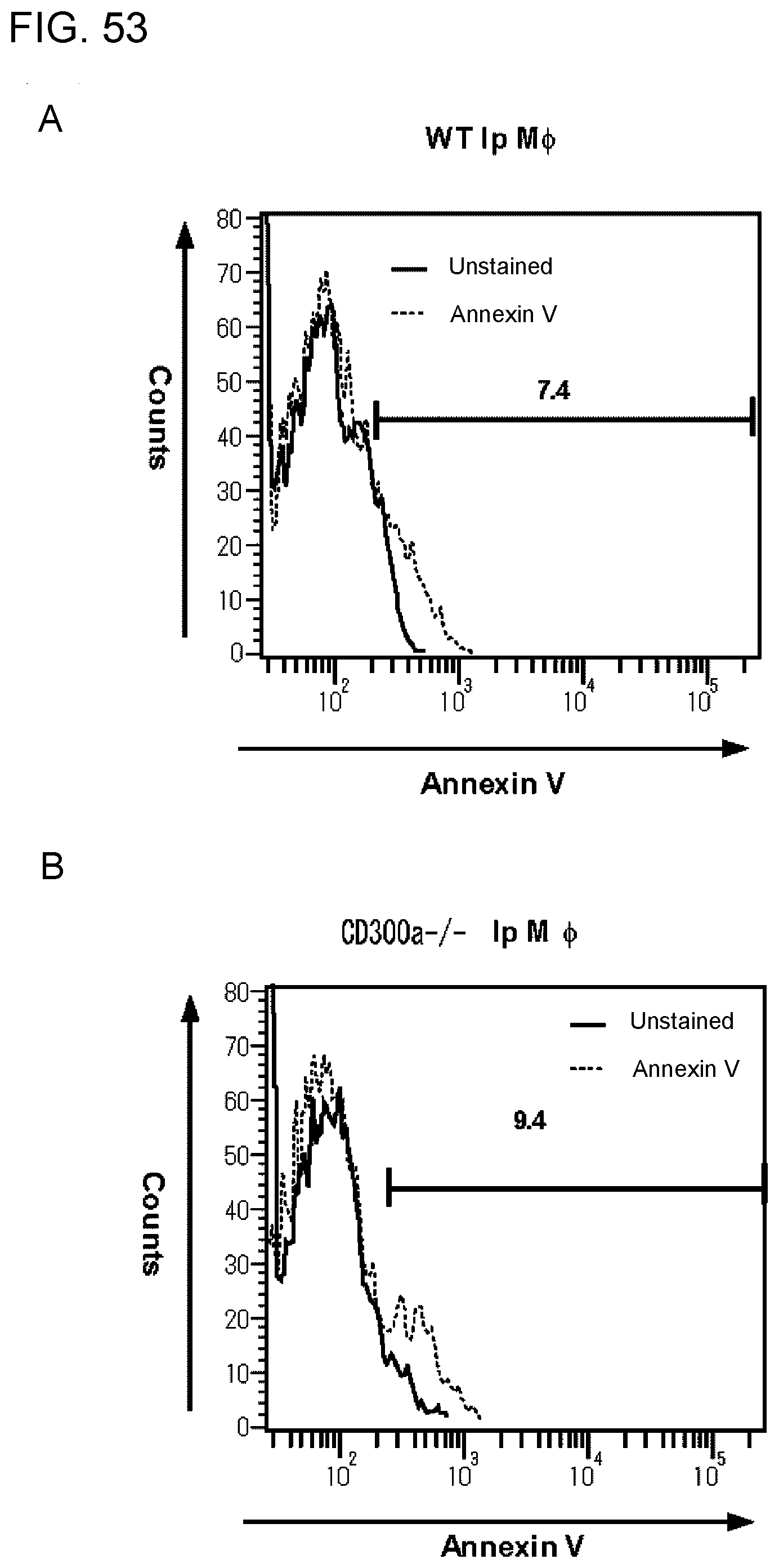
D00054
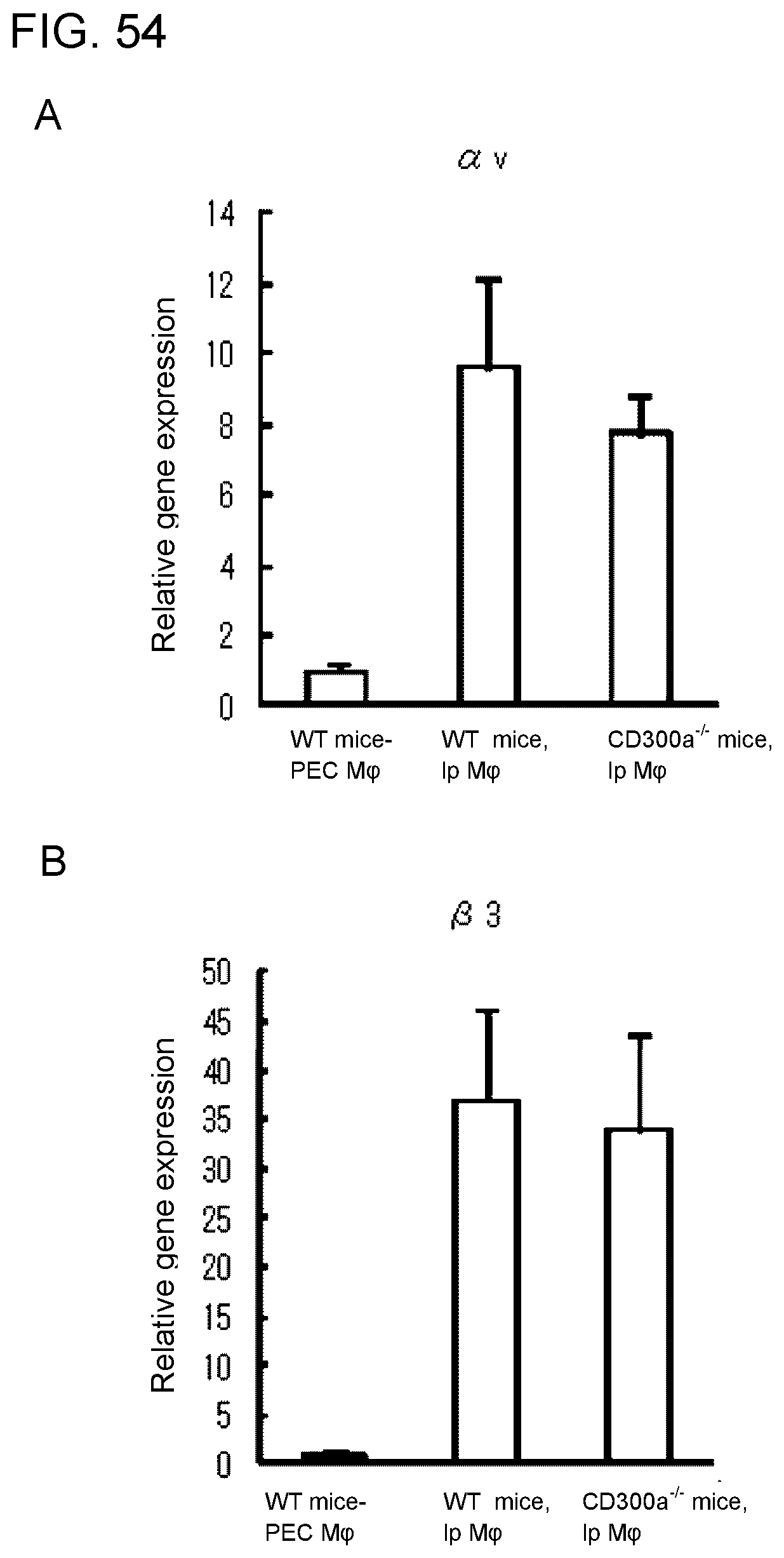
D00055
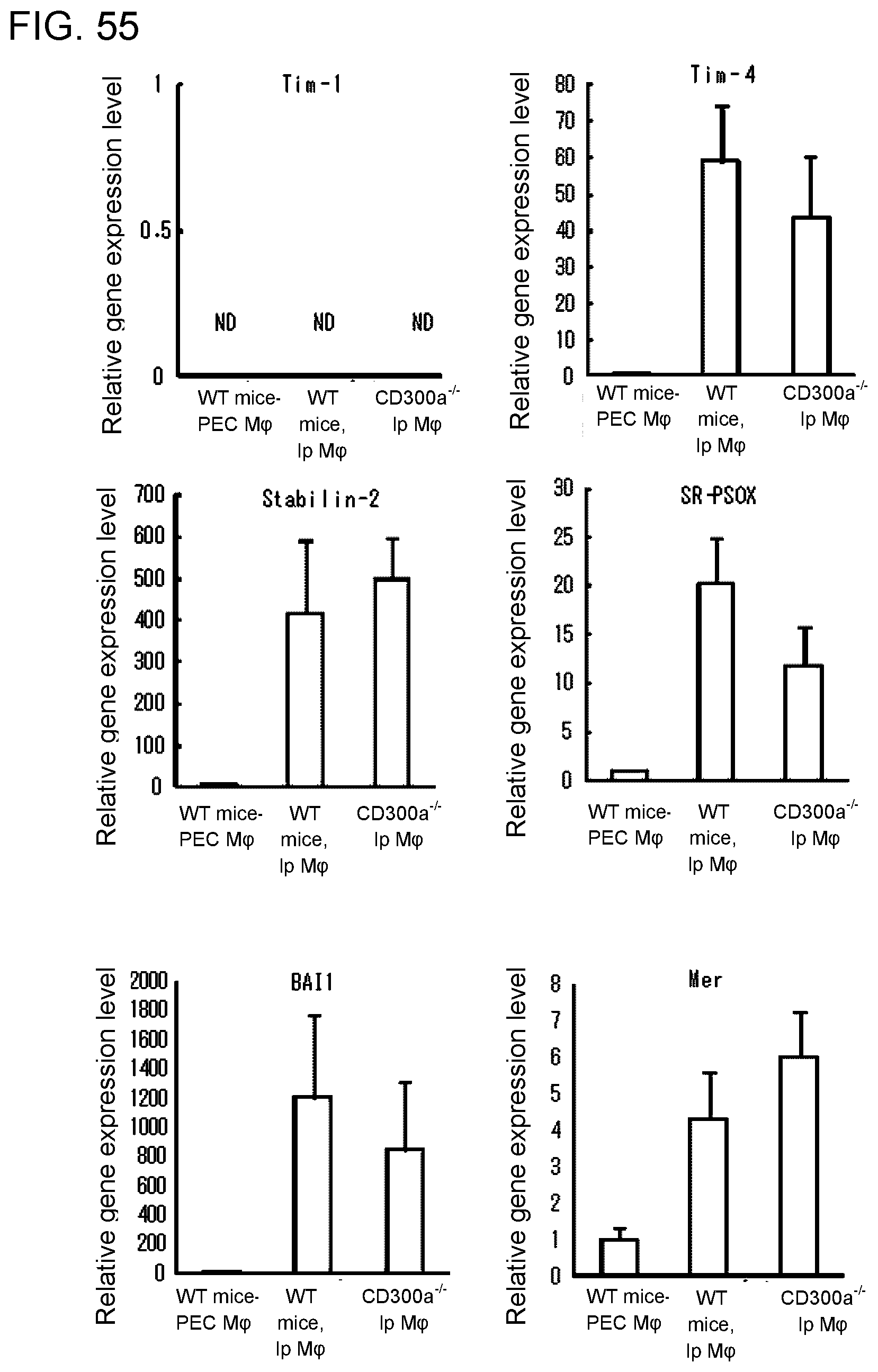
D00056

S00001
XML
uspto.report is an independent third-party trademark research tool that is not affiliated, endorsed, or sponsored by the United States Patent and Trademark Office (USPTO) or any other governmental organization. The information provided by uspto.report is based on publicly available data at the time of writing and is intended for informational purposes only.
While we strive to provide accurate and up-to-date information, we do not guarantee the accuracy, completeness, reliability, or suitability of the information displayed on this site. The use of this site is at your own risk. Any reliance you place on such information is therefore strictly at your own risk.
All official trademark data, including owner information, should be verified by visiting the official USPTO website at www.uspto.gov. This site is not intended to replace professional legal advice and should not be used as a substitute for consulting with a legal professional who is knowledgeable about trademark law.