Multiple Virus Resistance In Plants
Huang; Shihshieh ; et al.
U.S. patent application number 16/279798 was filed with the patent office on 2019-08-15 for multiple virus resistance in plants. The applicant listed for this patent is MONSANTO TECHNOLOGY LLC. Invention is credited to Stanislaw Flasinski, Alessandra Frizzi, Brad Gabor, Charles Hagen, Shihshieh Huang, John Kao, Raquel Salati.
| Application Number | 20190249189 16/279798 |
| Document ID | / |
| Family ID | 42272567 |
| Filed Date | 2019-08-15 |








View All Diagrams
| United States Patent Application | 20190249189 |
| Kind Code | A1 |
| Huang; Shihshieh ; et al. | August 15, 2019 |
MULTIPLE VIRUS RESISTANCE IN PLANTS
Abstract
The present invention provides gene targets, constructs and methods for the genetic control of plant disease caused by multiple plant viruses. The present invention relates to achieving a plant protective effect through the identification of target coding sequences and the use of recombinant DNA technologies for post-transcriptionally repressing or inhibiting expression of the target coding sequences of plant-parasitic viruses. Protein-expression based approaches may also be utilized to augment phenotype resistance. Thus, transcription of a single transgenic event comprising one or more plant expression cassettes can allow for broad spectrum resistance of a plant to multiple plant viral strains and species among the geminiviruses, tospoviruses, and potexviruses.
| Inventors: | Huang; Shihshieh; (Woodland, CA) ; Flasinski; Stanislaw; (Ballwin, MO) ; Frizzi; Alessandra; (Davis, CA) ; Gabor; Brad; (Woodland, CA) ; Hagen; Charles; (Davis, CA) ; Kao; John; (Davis, CA) ; Salati; Raquel; (Hollister, CA) | ||||||||||
| Applicant: |
|
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Family ID: | 42272567 | ||||||||||
| Appl. No.: | 16/279798 | ||||||||||
| Filed: | February 19, 2019 |
Related U.S. Patent Documents
| Application Number | Filing Date | Patent Number | ||
|---|---|---|---|---|
| 15598095 | May 17, 2017 | |||
| 16279798 | ||||
| 13750971 | Jan 25, 2013 | 9670502 | ||
| 15598095 | ||||
| 12763790 | Apr 20, 2010 | 8455716 | ||
| 13750971 | ||||
| 61171021 | Apr 20, 2009 | |||
| Current U.S. Class: | 1/1 |
| Current CPC Class: | C12N 15/8218 20130101; C12N 15/8283 20130101 |
| International Class: | C12N 15/82 20060101 C12N015/82 |
Claims
1. A tomato plant comprising resistance to a plurality of plant virus species, wherein the resistance is provided by at least two different modes of action selected from the group consisting of dsRNA, miRNA, and inhibition of virion assembly.
2. The plant of claim 1, wherein: (a) the resistance is provided by at least three different modes of action; (b) wherein the resistance comprises resistance against a begomovirus, tospovirus or potexvirus; (c) wherein the resistance provided to at least one of the plant virus species is provided by expression of a nucleic acid construct that produces dsRNA; (d) wherein resistance provided to at least one of the plant virus species is provided by expression of a dsRNA fusion construct or wherein resistance provided to at least one of the plant virus species is provided by expression of a nucleic acid construct that produces dsRNA; or (e) wherein resistance is provided against a begomovirus or tospovirus by a sequence encoded by a stacked miRNA expression cassette.
3-5. (canceled)
6. The plant of claim 2, wherein the dsRNA interferes with expression of a virus coat protein gene, a virus movement protein gene or a virus replication gene; or wherein the miRNA interferes with expression of a virus coat protein gene, a virus movement protein gene or a virus replication gene.
7. The plant of claim 2, wherein the nucleic acid construct which produces dsRNA comprises a sequence selected from the group consisting of SEQ ID NOs:379-455.
8-10. (canceled)
11. The plant of claim 2, wherein the miRNA comprises a sequence selected from the group consisting of SEQ ID NOs:1, 7, 13, 19, 25, 31, 37, 43, 47, 51, 55, 59, 63, 67, 71, 85, 99, 113, 127, and 141.
12. The plant of claim 1, wherein: (a) resistance against a begomovirus is provided by expression of dsRNA which interferes with expression of a begomovirus replication gene; (b) resistance against a tospovirus or potexvirus is provided by expression of a dsRNA which interferes with expression of a virus coat protein gene or virus movement protein gene; (c) resistance against a potexvirus is provided by expression of a nucleic acid construct which produces miRNA; or (d) resistance against a begomovirus or tospovirus is provided by a sequence encoded by a stacked miRNA expression cassette.
13. The plant of claim 1, wherein resistance provided to at least one of the plant virus species is provided by expression of a tospovirus genome segment terminal sequence that inhibits virion assembly.
14. The plant of claim 1, wherein resistance provided to at least one of the plant virus species is provided by inhibiting virion assembly, wherein virion assembly is inhibited by a sequence comprised within a nucleic acid construct comprising a first nucleic acid segment and a second nucleic acid segment, wherein the first and second segments are substantially inverted repeats of each other and are linked together by a third nucleic acid segment, and wherein the third segment comprises at least one terminal sequence of a tospovirus genome segment that inhibits virion assembly.
15. The plant of claim 14, wherein the third nucleic acid comprises a tospovirus genome terminal sequence selected from the group consisting of: a terminal sequence of a CaCV or GBNV L genome segment, a terminal sequence of a CaCV or GBNV M genome segment, a terminal sequence of a CaCV or GBNV S genome segment, a tospovirus genome terminal repeat sequence, a nucleic acid sequence comprising SEQ ID NO: 167, a nucleic acid sequence comprising SEQ ID NO:168, a nucleic acid sequence comprising SEQ ID NO:376, a nucleic acid sequence comprising SEQ ID NO:377, a nucleic acid sequence comprising SEQ ID NO:378, and a nucleic acid sequence comprising SEQ ID NO: 455.
16. The plant of claim 15, wherein the tospovirus genome segment terminal repeat sequence comprises SEQ ID NO:167 or SEQ ID NO:168.
17. The plant of claim 1, wherein the plant comprises resistance to viruses of at least two of the Geminiviridae, Bunyaviridae and Flexiviridae families; or wherein the viruses are selected from the genera Potexvirus, Tospovirus, and Begomovirus.
18. (canceled)
19. The plant of claim 18, wherein the viruses are selected from the group consisting of: a) at least one of TYLCV, ToSLCV, ToLCNDV, PHYVV, PepGMV; b) one or more of TSWV, GBNV, CaCV; and c) PepMV.
20. The plant of claim 17, wherein the Potexvirus is Pepino mosaic virus; the begomovirus is TYLCV, ToLCNDV, PHYVV, ToSLCV, or PepGMV; or the tospovirus is CaCV, GBNV, or TSWV.
21. The plant of claim 18, wherein the begomovirus is TYLCV, ToLCNDV, PHYVV, ToSLCV, or PepGMV.
22. The plant of claim 18, wherein the topovirus is CaCV, GBNV, or TSWV.
23. The plant of claim 18, wherein the begomovirus is TYLCV and the Potexvirus is Pepino mosaic virus; or the tospovirus is TSWV and the potexvirus is Pepino mosaic virus; or the wherein the begomovirus is TYLCV, the potexvirus is Pepino mosaic virus, and the tospovirus is TSWV.
24. The plant of claim 1, further comprising a sequence selected from the group consisting of SEQ ID NOs:156, 158, 160, 162, 164, 166, and 363-375; or further comprising a sequence selected from the group consisting of: SEQ ID NOs:1, 7, 13, 19, 25, 31, 37, 43, 47, 51, 55, 59, 63, 67, 71, 85, 99, 113, 127, 141, and 379-454.
25. (canceled)
26. The plant of claim 1, comprising (a) at least one sequence selected from the group consisting of SEQ ID NOs:379-454 and at least one sequence selected from the group consisting of SEQ ID NOs:167, 168, 376, 377, 378, and 455; (b) at least one sequence selected from the group consisting of SEQ ID NOs:379-454 and at least one sequence selected from the group consisting of SEQ ID NOs:1, 7, 13, 19, 25, 31, 37, 43, 47, 51, 55, 59, 63, 67, 71, 85, 99, 113, 127, and 141; or (c) at least one sequence selected from the group consisting of SEQ ID NOs:1, 7, 13, 19, 25, 31, 37, 43, 47, 51, 55, 59, 63, 67, 71, 85, 99, 113, 127, and 141 and at least one sequence selected from the group consisting of SEQ ID NOs:167, 168, 376, 377, 378, and 455.
27. The plant of claim 1, wherein the plant comprises at least one heterologous nucleic acid sequence that confers viral resistance selected from the group consisting of a) a nucleic acid sequence that encodes an RNA sequence that is complementary to all or a part of a first target gene; b) a nucleic acid sequence that comprises multiple copies of at least one anti-sense DNA segment that is anti-sense to at least one segment of said at least one first target gene; c) a nucleic acid sequence that comprises a sense DNA segment from at least one target gene; d) a nucleic acid sequence that comprises multiple copies of at least one sense DNA segment of a target gene; e) a nucleic acid sequence that transcribes to RNA for suppressing a target gene by forming double-stranded RNA and that comprises at least one segment that is anti-sense to all or a portion of the target gene and at least one sense DNA segment that comprises a segment of said target gene; f) a nucleic acid sequence that transcribes to RNA for suppressing a target gene by forming a single double-stranded RNA that comprises multiple serial anti-sense DNA segments that are anti-sense to at least one segment of the target gene and multiple serial sense DNA segments that comprise at least one segment of said target gene; g) a nucleic acid sequence that transcribes to RNA for suppressing a target gene by forming multiple double strands of RNA and comprises multiple segments that are anti-sense to at least one segment of said target gene and multiple sense DNA segments of the target gene, and wherein said multiple anti-sense DNA segments and said multiple sense DNA segments are arranged in a series of inverted repeats; h) a nucleic acid sequence that comprises nucleotides derived from a plant miRNA; and i) a nucleic acid sequence encoding at least one tospovirus terminal sequence that interferes with virion assembly.
28. The plant of claim 27, further comprising a non-transgenic plant virus resistance trait.
29. The plant of claim 27, wherein expression of the at least one heterologous nucleic acid sequence results in resistance to two or more viruses selected from the group consisting of: tospoviruses, begomoviruses, and potexviruses.
30. A transgenic seed or cell of the plant of claim 1.
31. A method for conferring resistance in a tomato plant to a plurality of plant virus species, the method comprising expressing in the plant at least two nucleic acid sequences that collectively provide resistance to said plurality of plant virus species, wherein at least 2 different modes of action are utilized to provide such resistance, comprising expression of at least two sequences selected from the group consisting of: dsRNA, miRNA, and a sequence which interferes with virion assembly.
32. The method of claim 31, wherein: (a) the resistance comprises resistance against a begomovirus, tospovirus or potexvirus; (b) the resistance provided to at least one of the plant virus species is provided by expression of a nucleic acid construct that produces dsRNA; (c) the resistance provided to at least one of the plant virus species is provided by expression of a dsRNA fusion construct; (d) the resistance provided to at least one of the plant virus species is provided by expression of a nucleic acid construct that produces miRNA; (e) the resistance against a begomovirus or tospovirus is provided by a sequence encoded by a stacked miRNA expression cassette; or (f) the resistance provided to at least one of the plant virus species is provided by expression of a tospovirus genome segment terminal sequence that inhibits virion assembly.
33-34. (canceled)
35. The method of claim 32, wherein the dsRNA interferes with expression of a virus coat protein gene, a virus movement protein gene or a virus replication gene; or wherein the nucleic acid construct comprises a sequence selected from the group consisting of SEQ ID NOs:379-455.
36.-38. (canceled)
39. The method of claim 32, wherein the miRNA interferes with expression of a virus coat protein gene, a virus movement protein gene or a virus replication gene; or wherein resistance provided to at least one of the plant virus species is provided by expression of a nucleic acid construct that produces miRNA.
40. (canceled)
41. The method of claim 31, wherein: (a) resistance against a begomovirus is provided by expression of dsRNA which interferes with expression of a begomovirus replication gene; (b) resistance against a tospovirus or potexvirus is provided by expression of a dsRNA which interferes with expression of a virus coat protein gene or virus movement protein gene; (c) resistance against a potexvirus is provided by expression of a nucleic acid construct which produces miRNA; or (d) resistance against a begomovirus or tospovirus is provided by a sequence encoded by a stacked miRNA expression cassette.
42. (canceled)
43. The method of claim 31, wherein resistance provided to at least one of said plant virus species is provided by inhibiting virion assembly, wherein virion assembly is inhibited by a sequence comprised within a nucleic acid construct comprising a first nucleic acid segment and a second nucleic acid segment, wherein the first and second segments are substantially inverted repeats of each other and are linked together by a third nucleic acid segment, and wherein the third segment comprises at least one terminal sequence of a tospovirus genome segment, expression of which inhibits virion assembly.
44. The method of claim 43, wherein the third nucleic acid comprises a tospovirus genome terminal sequence selected from the group consisting of: a terminal sequence of a CaCV or GBNV L genome segment, a terminal sequence of a CaCV or GBNV M genome segment, a terminal sequence of a CaCV or GBNV S genome segment, and a tospovirus genome terminal repeat sequence.
45. The method of claim 44, wherein the terminal sequence or terminal repeat sequence comprises SEQ ID NO:167, SEQ ID NO:168, SEQ ID NO:376, SEQ ID NO:377, or SEQ ID NO: 378.
46. The method of claim 31, wherein the plurality of plant virus species are selected from at least two of the Geminiviridae, Bunyaviridae and Flexiviridae families; or wherein the viruses are selected from the genera Potexvirus, Tospovirus, and Begomovirus.
47. (canceled)
48. The method of claim 47, wherein the viruses are selected from the group consisting of: a) one or more of TYLCV, ToSLCV, ToLCNDV, PHYVV, PepGMV; b) one or more of TSWV, GBNV, CaCV; and c) PepMV.
49. The method of claim 31, wherein the nucleic acid sequence comprises at least one gene suppression element for suppressing at least one first target gene.
50. The method of claim 31, comprising expressing in the plant: a) at least one sequence selected from the group consisting of SEQ ID NOs:379-454 and at least one sequence selected from the group consisting of SEQ ID NOs:167, 168, 376, 377, 378, and 455; (b) at least one sequence selected from the group consisting of SEQ ID NOs:379-454 and at least one sequence selected from the group consisting of SEQ ID NOs:1, 7, 13, 19, 25, 31, 37, 43, 47, 51, 55, 59, 63, 67, 71, 85, 99, 113, 127, and 141; or (c) at least one sequence selected from the group consisting of SEQ ID NOs:1, 7, 13, 19, 25, 31, 37, 43, 47, 51, 55, 59, 63, 67, 71, 85, 99, 113, 127, and 141 and at least one sequence selected from the group consisting of SEQ ID NOs:167, 168, 376, 377, 378, and 455.
51. (canceled)
Description
[0001] This application claims the priority of U.S. Provisional Appl. Ser. No. 61/171,021, filed Apr. 20, 2009, the entire disclosure of which is incorporated herein by reference.
BACKGROUND OF THE INVENTION
1. Field of the Invention
[0002] The present invention relates generally to methods and compositions for enhancing resistance to multiple plant viruses.
2. Description of Related Art
[0003] Solanaceous plants are subject to multiple potential disease causing agents, including virus-induced diseases that are responsible for major crop losses worldwide. For many RNA viruses, expression of transgenic coat protein (CP) or replicase blocks the progression of the virus infectious process. RNA-based resistance makes use of the plant post-transcriptional gene silencing (PTGS) mechanism to degrade viral RNAs. However, such approaches may yield resistance that is narrowly based and/or not durable, especially with rapidly spreading/evolving new viral species or isolates. In some instances, classically-defined (non-transgenic) resistance traits are available to aid in development of virus resistant plants. Additionally, control of plant pests, such as insects that serve to transmit plant viruses, may help to limit losses due to viral infection of plants.
BRIEF DESCRIPTION OF THE FIGURES
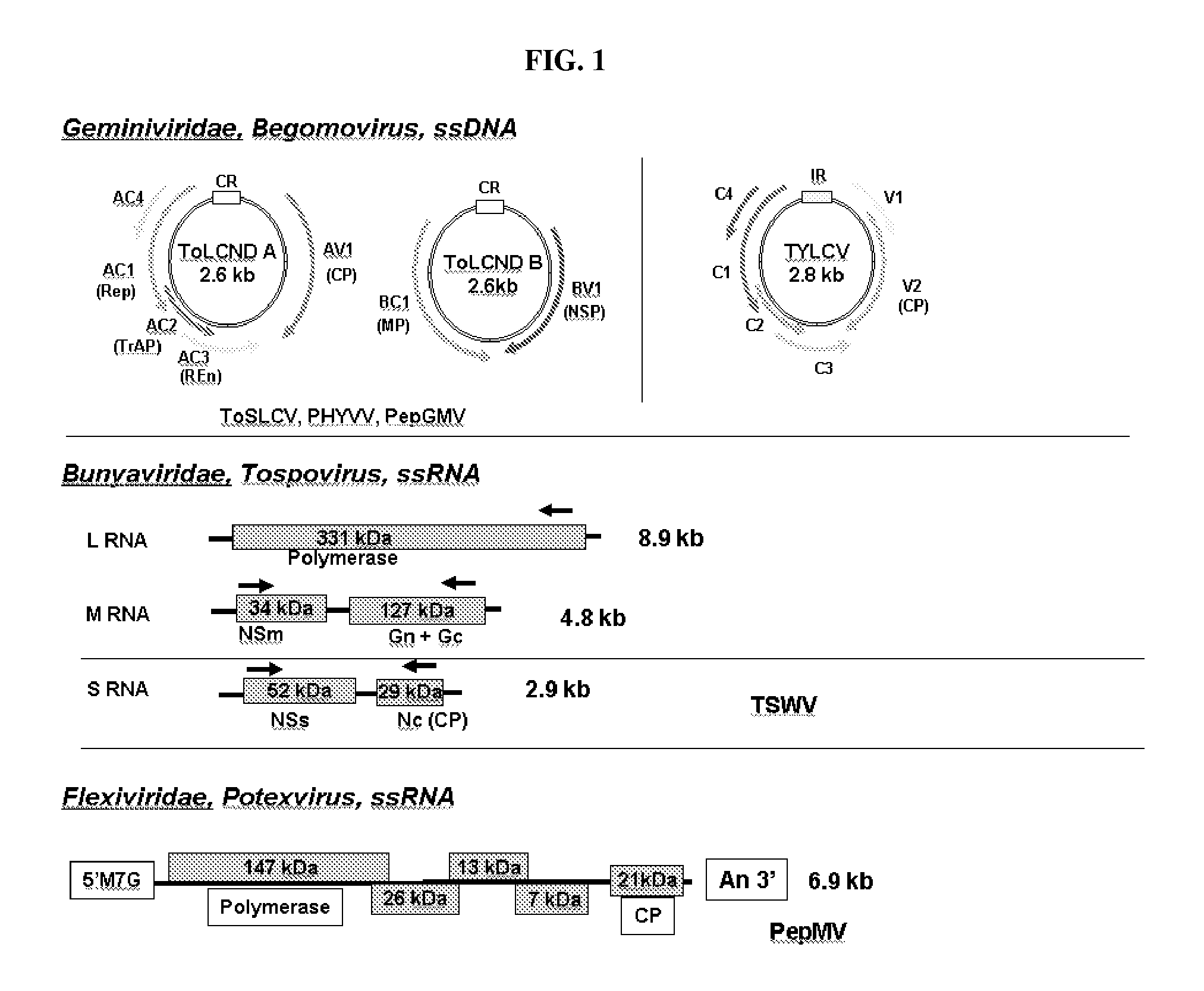
[0004] FIG. 1: Schematic diagram of genome organization of viruses of interest.
[0005] FIG. 2A: Schematic diagram illustrating approach for identifying sequence efficacious for plant virus control.
[0006] FIG. 2B: Schematic diagram of a typical begomovirus DNA-A genome showing location of regions screened for effectiveness for viral control when expressed as inverted repeats. Numbered gray arrows represent portions of genome that were tested.
[0007] FIG. 2C: Schematic diagram of a typical potexvirus (Pepino mosaic virus; PepMV) genome showing location of regions screened for effectiveness for viral control when expressed as inverted repeats. Numbered gray arrows represent portions of genome that were tested.
[0008] FIG. 2D: Schematic diagram of a tospovirus (e.g. Tomato spotted wilt virus (TSWV) genome showing location of regions screened for effectiveness for viral control when expressed as inverted repeats. Numbered gray arrows represent portions of genome that were tested.
[0009] FIG. 3: Virus resistance correlating with siRNA production in transformed tomato plants.
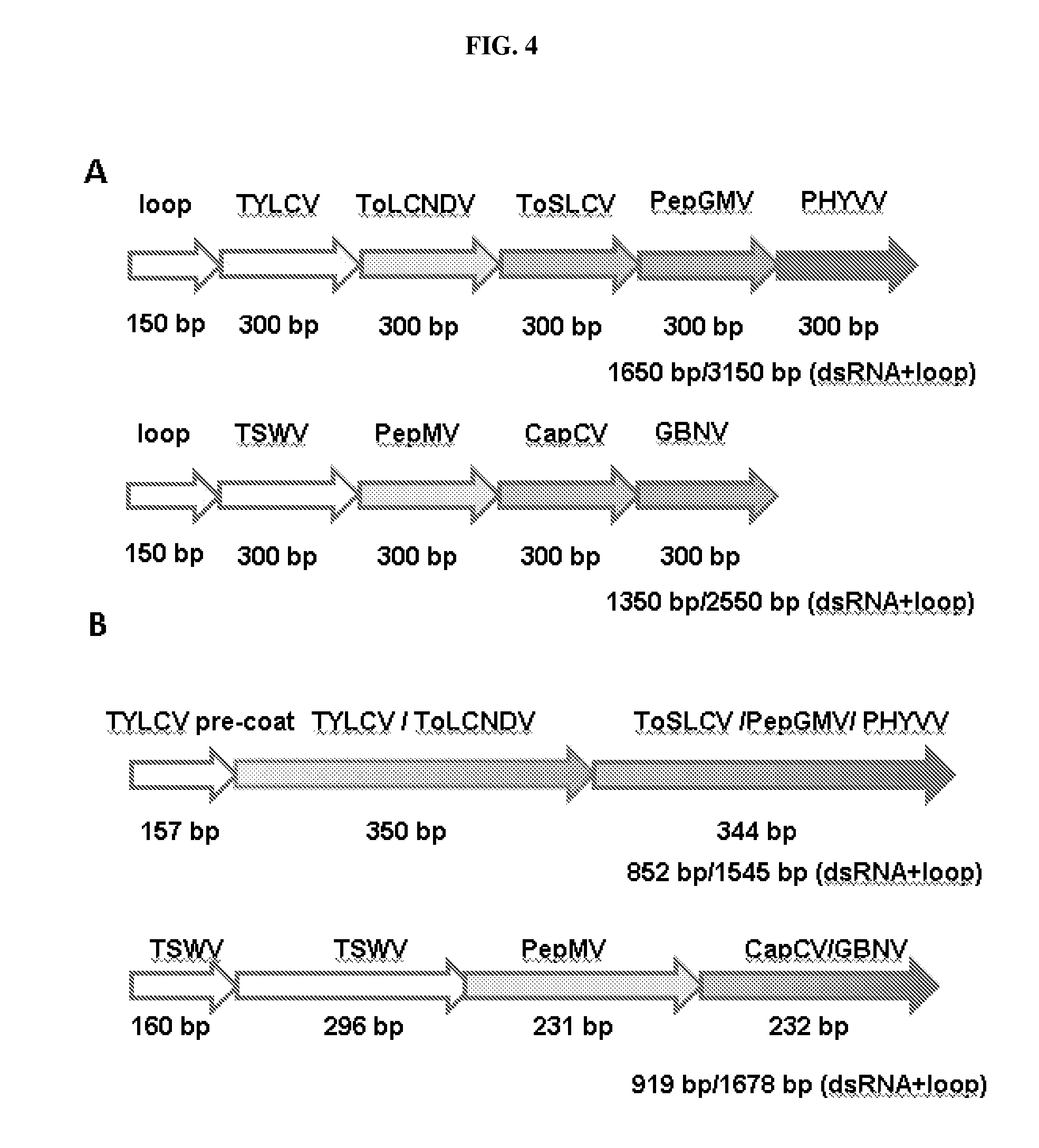
[0010] FIG. 4: Exemplary artificial dsRNA fusion constructs for conferring multiple virus resistance ("MVR").
[0011] FIG. 5: Suitable 21nt sequences (among SEQ ID NOs:1-42) that were analyzed against five targeted Geminiviruses (perfect match: double underline; G:U mis-match: single underline; other mis-match or not utilized: not underlined).
[0012] FIG. 6: Suitable 21nt sequences (among SEQ ID NOs:43-70) that were analyzed against tospoviruses (perfect match: double underline; G:U mis-match: single underline; other mis-match or not utilized: not underlined).
[0013] FIGS. 7A, 7B: Suitable 21nt sequences (among SEQ ID NOs:71-154) that were analyzed against targeted potexviruses (perfect match: double underline; G:U mis-match: single underline; other mis-match or not utilized: not underlined).
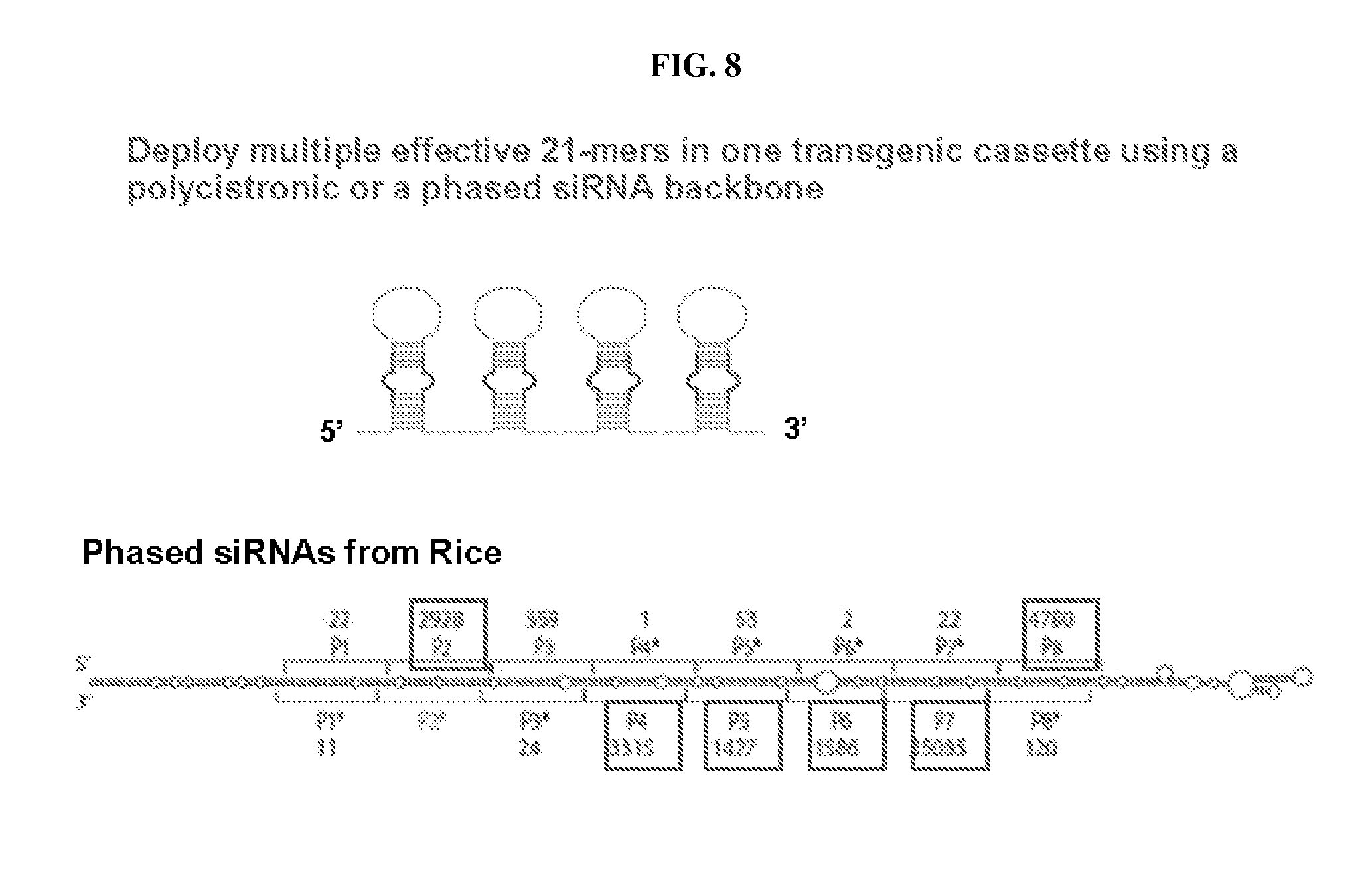
[0014] FIG. 8: Schematic of exemplary construct for deploying multiple engineered miRNAs in one transgenic cassette, such as with phased siRNAs.
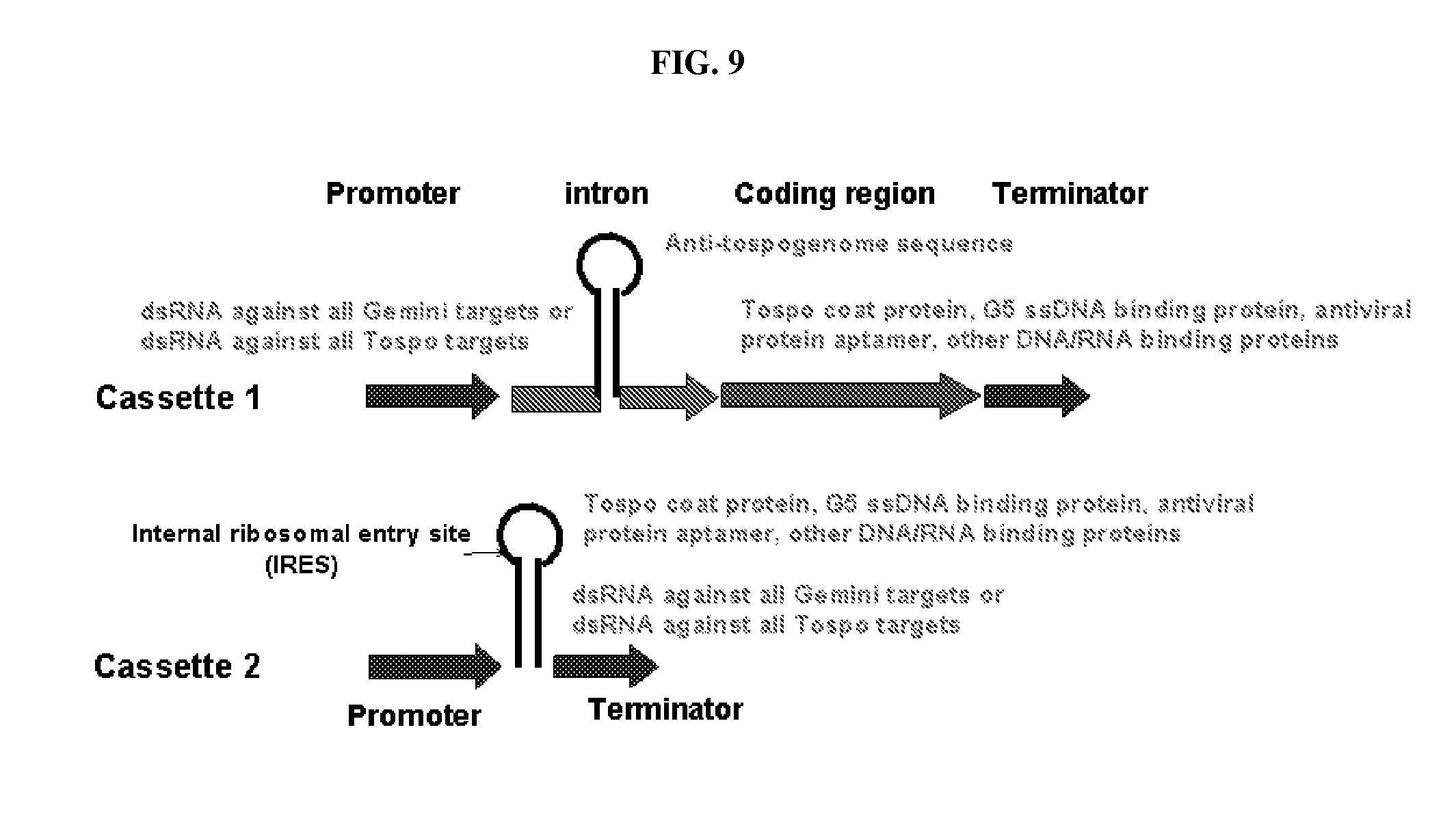
[0015] FIG. 9: Schematic diagram illustrating expression cassette for deploying multiple modes of action for virus resistance.
[0016] FIG. 10: Additional exemplary constructs for deploying multiple engineered miRNAs in a transgenic cassette, as well as for expressing miRNA along with dsRNA.
[0017] FIGS. 11A, 11B: Scanning of regions of the tospovirus genome to define segments which may be expressed as dsRNA with anti-viral efficacy. X axis represents individual events and target regions (CP: coat protein; GP: envelope glycoprotein; and RdRP: RNA-dependent RNA polymerase). Y axis represents % of transgenic R.sub.1 plants displaying virus resistance. FIG. 11A: TSWV results; FIG. 11B: CaCV and GBNV results. "CP4+" refers to presence of the selectable marker gene linked to the dsRNA-encoding sequence, in R.sub.1 plants.
[0018] FIG. 12: Regions of the PepMV genome tested for effectiveness in generating dsRNA-mediated resistance against this potexvirus (CP: coat protein; Mov: movement protein; RdRP, or RdR: RNA-dependent RNA polymerase; TGB: Triple gene block protein).
[0019] FIGS. 13A, 13B: Regions of geminivirus genome assayed for effectiveness in generating dsRNA-mediated resistance against this virus group. (CP: coat protein; Rep: replication protein). Other transgenic plants contain a glyphosate resistance gene. "0%-100%" denotes the percentage of R.sub.1 plants that are selectable-marker positive and virus resistant. FIG. 13A: representative results for TYLCV and ToSLCV; FIG. 13B: representative results for PepGMV, PHYVV, and ToLCNDV. For PHYVV and ToLCNDV events, "Spc" refers to the presence of a selectable marker gene conferring spectinomycin resistance. Other events were transformed with a construct comprising a selectable marker gene conferring glyphosate resistance ("CP4 positive").
[0020] FIG. 14: Schematic diagram illustrating representative expression cassettes for targeting of multiple viruses in multiple virus families.
[0021] FIG. 15: Results of using an artificial dsRNA fusion construct targeting CP expression of tospoviruses and PepMV (potexvirus), for multiple virus resistance as discussed in Example 3. Construct used is schematically shown in FIG. 4B, bottom construct: TSWV (160 bp), TSWV (296 bp), PepMV (231 bp), CaCV/GBNV (232 bp).
[0022] FIG. 16: Depicts resistance observed against inoculated CaCV in inoculated CP4 positive R.sub.1 plants transformed with a construct comprising tospovirus terminal repeat sequences (SEQ ID NOs:167, 168, 376, 377, 378, as found in SEQ ID NO:455).
SUMMARY OF THE INVENTION
[0023] The present invention provides methods and compositions for obtaining plants resistant to multiple plant viruses. In one aspect, the present invention provides a tomato plant comprising resistance to a plurality of plant virus species. In certain embodiments, the resistance is provided by at least two different modes of action selected from the group consisting of dsRNA, miRNA, and inhibition of virion assembly. In other embodiments, the resistance is provided by at least three different modes of action. The resistance of the tomato plant may comprise resistance against begomovirus, tospovirus or potexvirus.
[0024] In certain embodiments, the resistance provided to at least one of the plant virus species is provided by expression of a nucleic acid construct that produces dsRNA. In some embodiments the resistance provided to at least one of the plant virus species is provided by expression of a dsRNA fusion construct. In some embodiments of the invention, the dsRNA interferes with expression of a virus coat protein gene, a virus movement protein gene or a virus replication gene. In particular embodiments, the nucleic acid construct which produces dsRNA comprises a sequence selected from the group consisting of SEQ ID NOs:379-455.
[0025] In other embodiments, the resistance provided to at least one of the plant virus species is provided by expression of a nucleic acid construct that produces miRNA. Thus, in certain embodiments, the resistance against a begomovirus or tospovirus is provided by a sequence encoded by a stacked miRNA expression cassette. In yet other embodiments, the miRNA interferes with expression of a virus coat protein gene, a virus movement protein gene or a virus replication gene. In particular embodiments, the miRNA comprises a sequence selected from the group consisting of SEQ ID NOs:1, 7, 13, 19, 25, 31, 37, 43, 47, 51, 55, 59, 63, 67, 71, 85, 99, 113, 127, and 141.
[0026] In certain embodiments, the tomato plant comprises (a) resistance against a begomovirus which is provided by expression of dsRNA which interferes with expression of a begomovirus replication gene; (b) resistance against a tospovirus or potexvirus which is provided by expression of a dsRNA which interferes with expression of a virus coat protein gene or virus movement protein gene; (c) resistance against a potexvirus which is provided by expression of a nucleic acid construct which produces miRNA; or (d) resistance against a begomovirus or tospovirus which is provided by a sequence encoded by a stacked miRNA expression cassette.
[0027] In other embodiments, a tomato plant is provided wherein resistance provided to at least one of the plant virus species is provided by expression of a tospovirus genome segment terminal sequence that inhibits virion assembly. In certain embodiments, resistance provided to at least one of the plant virus species is provided by inhibiting virion assembly, wherein virion assembly is inhibited by a sequence comprised within a nucleic acid construct comprising a first nucleic acid segment and a second nucleic acid segment, wherein the first and second segments are substantially inverted repeats of each other and are linked together by a third nucleic acid segment, and wherein the third segment comprises at least one terminal sequence of a tospovirus genome segment that inhibits virion assembly. In particular embodiments, the third nucleic acid comprises a tospovirus genome terminal sequence selected from the group consisting of: a terminal sequence of a CaCV or GBNV L genome segment, a terminal sequence of a CaCV or GBNV M genome segment, a terminal sequence of a CaCV or GBNV S genome segment, a tospovirus genome terminal repeat sequence, a nucleic acid sequence comprising SEQ ID NO: 167, a nucleic acid sequence comprising SEQ ID NO:168, a nucleic acid sequence comprising SEQ ID NO:376, a nucleic acid sequence comprising SEQ ID NO:377, a nucleic acid sequence comprising SEQ ID NO:378, and a nucleic acid sequence comprising SEQ ID NO: 455.
[0028] In certain embodiments, the tomato plant comprises resistance to viruses of at least two of the Geminiviridae, Bunyaviridae and Flexiviridae families. Thus, in some embodiments the viruses are selected from the genera Potexvirus, Tospovirus, and Begomovirus. In particular embodiments, the viruses are selected from the group consisting of: a) at least one of TYLCV, ToSLCV, ToLCNDV, PHYVV, PepGMV; b) one or more of TSWV, GBNV, CaCV; and c) PepMV. In a more particular embodiment, the potexvirus is Pepino mosaic virus. In certain embodiments, the begomovirus is TYLCV, ToLCNDV, PHYVV, ToSLCV, or PepGMV. In some embodiments, the tospovirus is CaCV, GBNV, or TSWV. In particular embodiments, the begomovirus is TYLCV and the potexvirus is Pepino mosaic virus; or the tospovirus is TSWV and the potexvirus is Pepino mosaic virus; or the wherein the begomovirus is TYLCV, the potexvirus is Pepino mosaic virus, and the tospovirus is TSWV.
[0029] In some embodiments, the tomato plant may comprise a sequence selected from the group consisting of SEQ ID NOs:156, 158, 160, 162, 164, 166, and 363-375. In those or other embodiments, the tomato plant comprises, or further comprises, a sequence selected from the group consisting of: SEQ ID NOs:1, 7, 13, 19, 25, 31, 37, 43, 47, 51, 55, 59, 63, 67, 71, 85, 99, 113, 127, 141, and 379-454. Thus, a tomato plant of the invention may comprise: (a) at least one sequence selected from the group consisting of SEQ ID NOs:379-454 and at least one sequence selected from the group consisting of SEQ ID NOs:167, 168, 376, 377, 378, and 455; (b) at least one sequence selected from the group consisting of SEQ ID NOs:379-454 and at least one sequence selected from the group consisting of SEQ ID NOs:1, 7, 13, 19, 25, 31, 37, 43, 47, 51, 55, 59, 63, 67, 71, 85, 99, 113, 127, and 141; or (c) at least one sequence selected from the group consisting of SEQ ID NOs:1, 7, 13, 19, 25, 31, 37, 43, 47, 51, 55, 59, 63, 67, 71, 85, 99, 113, 127, and 141 and at least one sequence selected from the group consisting of SEQ ID NOs:167, 168, 376, 377, 378, and 455.
[0030] In yet other embodiments, the tomato plant comprises at least one heterologous nucleic acid sequence that confers viral resistance selected from the group consisting of a) a nucleic acid sequence that encodes an RNA sequence that is complementary to all or a part of a first target gene; b) a nucleic acid sequence that comprises multiple copies of at least one anti-sense DNA segment that is anti-sense to at least one segment of said at least one first target gene; c) a nucleic acid sequence that comprises a sense DNA segment from at least one target gene; d) a nucleic acid sequence that comprises multiple copies of at least one sense DNA segment of a target gene; e) a nucleic acid sequence that transcribes to RNA for suppressing a target gene by forming double-stranded RNA and that comprises at least one segment that is anti-sense to all or a portion of the target gene and at least one sense DNA segment that comprises a segment of said target gene; f) a nucleic acid sequence that transcribes to RNA for suppressing a target gene by forming a single double-stranded RNA that comprises multiple serial anti-sense DNA segments that are anti-sense to at least one segment of the target gene and multiple serial sense DNA segments that comprise at least one segment of said target gene; g) a nucleic acid sequence that transcribes to RNA for suppressing a target gene by forming multiple double strands of RNA and comprises multiple segments that are anti-sense to at least one segment of said target gene and multiple sense DNA segments of the target gene, and wherein said multiple anti-sense DNA segments and said multiple sense DNA segments are arranged in a series of inverted repeats; h) a nucleic acid sequence that comprises nucleotides derived from a plant miRNA; and i) a nucleic acid sequence encoding at least one tospovirus terminal sequence that interferes with virion assembly. The invention also provides a plant wherein expression of the at least one heterologous nucleic acid sequence results in resistance to two or more viruses selected from the group consisting of: tospoviruses, begomoviruses, and potexviruses. The plant may also further comprise a non-transgenic plant virus resistance trait.
[0031] In another aspect of the invention, a transgenic seed is provided, of any generation of the tomato plant comprising resistance to a plurality of plant virus species, wherein the resistance is provided by at least two different modes of action selected from the group consisting of dsRNA, miRNA, and inhibition of virion assembly.
[0032] In yet another aspect, the invention provides a method for conferring resistance in a tomato plant to a plurality of plant virus species, the method comprising expressing in the plant at least two nucleic acid sequences that collectively provide resistance to said plurality of plant virus species, wherein at least 2 different modes of action are utilized to provide such resistance, comprising expression of at least two sequences selected from the group consisting of: dsRNA, miRNA, and a sequence which interferes with virion assembly. In certain embodiments, the resistance comprises resistance against a begomovirus, tospovirus or potexvirus.
[0033] The resistance may be provided to at least one of the plant virus species by expression of a nucleic acid construct that produces dsRNA. In particular embodiments, resistance provided to at least one of the plant virus species is provided by expression of a dsRNA fusion construct. In more particular embodiments, the dsRNA interferes with expression of a virus coat protein gene, a virus movement protein gene or a virus replication gene. In yet more particular embodiments, the nucleic acid construct comprises a sequence selected from the group consisting of SEQ ID NOs:379-455.
[0034] In other embodiments, resistance provided to at least one of the plant virus species is provided by expression of a nucleic acid construct that produces miRNA. In one embodiment, it is contemplated that resistance against a begomovirus or tospovirus is provided by a sequence encoded by a stacked miRNA expression cassette. The produced miRNA may further interfere with expression of a virus coat protein gene, a virus movement protein gene or a virus replication gene. In particular embodiments, the miRNA comprises a sequence selected from the group consisting of SEQ ID NOs:1, 7, 13, 19, 25, 31, 37, 43, 47, 51, 55, 59, 63, 67, 71, 85, 99, 113, 127, and 141.
[0035] Thus, in certain embodiments, (a) resistance against a begomovirus is provided by expression of dsRNA which interferes with expression of a begomovirus replication gene; (b) resistance against a tospovirus or potexvirus is provided by expression of a dsRNA which interferes with expression of a virus coat protein gene or virus movement protein gene; (c) resistance against a potexvirus is provided by expression of a nucleic acid construct which produces miRNA; or (d) resistance against a begomovirus or tospovirus is provided by a sequence encoded by a stacked miRNA expression cassette.
[0036] In some embodiments resistance provided to at least one of the plant virus species is provided by expression of a tospovirus genome segment terminal sequence that inhibits virion assembly. In certain embodiments, resistance is provided to at least one of said plant virus species by inhibiting virion assembly, wherein virion assembly is inhibited by a sequence comprised within a nucleic acid construct comprising a first nucleic acid segment and a second nucleic acid segment, wherein the first and second segments are substantially inverted repeats of each other and are linked together by a third nucleic acid segment, and wherein the third segment comprises at least one terminal sequence of a tospovirus genome segment, expression of which inhibits virion assembly. Further, in particular embodiments, the third nucleic acid may comprise a tospovirus genome terminal sequence selected from the group consisting of: a terminal sequence of a CaCV or GBNV L genome segment, a terminal sequence of a CaCV or GBNV M genome segment, a terminal sequence of a CaCV or GBNV S genome segment, and a tospovirus genome terminal repeat sequence. In more particular embodiments, the terminal sequence or terminal repeat sequence comprises SEQ ID NO:167, SEQ ID NO:168, SEQ ID NO:376, SEQ ID NO:377, or SEQ ID NO: 378.
[0037] In other embodiments of the invention, the plurality of plant virus species are selected from at least two of the Geminiviridae, Bunyaviridae and Flexiviridae families. Thus, the viruses may be selected from the genera Potexvirus, Tospovirus, and Begomovirus. In certain embodiments, the viruses are selected from the group consisting of: a) one or more of TYLCV, ToSLCV, ToLCNDV, PHYVV, PepGMV; b) one or more of TSWV, GBNV, CaCV; and c) PepMV.
[0038] In other embodiments, the nucleic acid sequence comprises at least one gene suppression element for suppressing at least one first target gene. For instance, the method may comprise expressing in the plant: a) at least one sequence selected from the group consisting of SEQ ID NOs:379-454 and at least one sequence selected from the group consisting of SEQ ID NOs:167, 168, 376, 377, 378, and 455; (b) at least one sequence selected from the group consisting of SEQ ID NOs:379-454 and at least one sequence selected from the group consisting of SEQ ID NOs:1, 7, 13, 19, 25, 31, 37, 43, 47, 51, 55, 59, 63, 67, 71, 85, 99, 113, 127, and 141; or (c)at least one sequence selected from the group consisting of SEQ ID NOs:1, 7, 13, 19, 25, 31, 37, 43, 47, 51, 55, 59, 63, 67, 71, 85, 99, 113, 127, and 141 and at least one sequence selected from the group consisting of SEQ ID NOs:167, 168, 376, 377, 378, and 455.
[0039] Another aspect of the invention provides a transgenic cell of the tomato plant comprising resistance to a plurality of plant virus species, wherein the resistance is provided by at least two different modes of action selected from the group consisting of dsRNA, miRNA, and inhibition of virion assembly.
[0040] The term "about" is used to indicate that a value includes the standard deviation of error for the device or method being employed to determine the value. The use of the term "or" in the claims is used to mean "and/or" unless explicitly indicated to refer to alternatives only or the alternatives are mutually exclusive, although the disclosure supports a definition that refers to only alternatives and to "and/or." When not used in conjunction closed wording in the claims or specifically noted otherwise, the words "a" and "an" denote "one or more." The term "conferred by a transgene," for example, thus encompasses one ore more transgene(s).
[0041] The terms "comprise," "have" and "include" are open-ended linking verbs. Any forms or tenses of one or more of these verbs, such as "comprises," "comprising," "has," "having," "includes" and "including," are also open-ended. For example, any method that "comprises," "has" or "includes" one or more steps is not limited to possessing only those one or more steps and also covers other unlisted steps. Similarly, any plant that "comprises," "has" or "includes" one or more traits is not limited to possessing only those one or more traits and covers other unlisted traits.
[0042] Other objects, features and advantages of the present invention will become apparent from the following detailed description. It should be understood, however, that the detailed description and any specific examples provided, while indicating specific embodiments of the invention, are given by way of illustration only, since various changes and modifications within the spirit and scope of the invention will become apparent to those skilled in the art from this detailed description.
DETAILED DESCRIPTION OF THE INVENTION
[0043] The following is a detailed description of the invention provided to aid those skilled in the art in practicing the present invention. Those of ordinary skill in the art may make modifications and variations in the embodiments described herein without departing from the spirit or scope of the present invention.
[0044] The present invention provides methods and compositions for genetic control of virus diseases in plants, including Solanaceous plants such as tomato (i.e. Lycopersicon or Solanum sp.), pepper (i.e. Capsicum sp.), petunia (i.e. Petunia sp.), and potato and eggplant (i.e. Solanum sp). In one embodiment RNA-mediated gene suppression can be conferred by the expression of an inverted-repeat transgene cassette that generates a population of small interfering RNAs (siRNAs) derived from the dsRNA region of a transgene transcript. Another RNA-mediated approach for gene suppression is by expression of one or more miRNA segments that "target" specific transcripts and lead to their degradation. Thus approaches including engineering dsRNA, miRNA, ta-siRNA and/or phased siRNA may be utilized in accordance with the invention. For instance, begomovirus-derived, or other virus-derived sequences targeting replication, coat protein, and C2 and/or C3 proteins may also be utilized. Likewise, for control of potexviruses such as Pepino mosaic virus, sequences targeting portions of coat (capsid) protein ("CP"), replication protein such as RNA-dependent RNA polymerase ("RdRP"), and/or one or more movement protein(s) ("MP") which may include a triple gene block ("TGB") or a "30K" MP may be used. For control of tospoviruses, sequences targeting, for instance, the coat protein ("CP", also termed the nucleocapsid, "N" protein), RdRP, movement protein ("NsM"), and/or non-structural glyocoprotein(s) (encoded by "G1" or G2" genes) may similarly be utilized. Such sequences may correspond exactly to sequences from one or more viral isolates, or may be variants, for instance designed to increase their antiviral efficacy, or avoid non-target effects.
[0045] In certain embodiments, multiple virus resistance ("MVR") is achieved by utilizing dsRNA and/or miRNA expressed from a single transformed construct, or more than one construct. Further, a single construct may comprise one or more expression cassettes that produce dsRNA and/or miRNA that targets one or more functions necessary for plant viral infection, multiplication, and/or transmission, as well as, in certain embodiments, one or more expression cassettes that produce at least one mRNA that encodes a protein, or portion of a protein, being targeted. Thus, resistance to multiple plant viruses may be achieved in a single transgenic "event." RNA mediated resistance may further be enhanced by protein based approaches utilizing aptamer(s) that inhibit replication, expression or mutation in replicase or replication associated proteins, ssDNA binding proteins such as m13-G5 (e.g. U.S. Pat. No. 6,852,907; Padidam et al., 1999), for geminivirus resistance, or a peptide aptamer that interferes with geminivirus replication may also be employed (e.g. Lopez-Ochoa et al., 2006).
[0046] It is also contemplated that inhibition of virion assembly, for instance by a nucleic-acid based approach, may be utilized as a mode of action in providing virus resistance to a tomato plant. This inhibition of virion assembly may be provided, for instance, by use of a tospovirus terminal sequence, such as a terminal repeat sequence. By "inhibition of virion assembly" is meant interference with the interaction between viral capsid proteins and nucleic acid(s) which together may form a viral particle ("virion"). Such interference may occur, for instance, by expression of a sequence that can serve as an artificial substrate competing for reverse transcriptase, and/or may occur by interference with proper circularization of replicating viral genome components.
[0047] Additionally, classical genetic resistance loci for tolerance in tomato, peppers, and other Solanaceous plants may be utilized, for instance through classical breeding approaches. In certain embodiments, protein-based approaches using tospovirus "N" gene (nucleocapsid; coat protein) plus/minus inverted repeats and a potexvirus (e.g. Pepino mosaic virus) coat protein (CP) and replicase for resistance, are also provided.
[0048] Methods of gene suppression may include use of anti-sense, co-suppression, and RNA interference. Anti-sense gene suppression in plants is described by Shewmaker et al. in U.S. Pat. Nos. 5,107,065, 5,453,566, and 5,759,829. Gene suppression in bacteria using DNA which is complementary to mRNA encoding the gene to be suppressed is disclosed by Inouye et al. in U.S. Pat. Nos. 5,190,931, 5,208,149, and 5,272,065. RNA interference or RNA-mediated gene suppression has been described by, e.g., Redenbaugh et al., 1992; Chuang et al., 2000; and Wesley et al., 2001.
[0049] Several cellular pathways involved in RNA-mediated gene suppression have been described, each distinguished by a characteristic pathway and specific components. See, for example, reviews by Brodersen and Voinnet (2006), and Tomari and Zamore (2005). The siRNA pathway involves the non-phased cleavage of a double-stranded RNA to small interfering RNAs ("siRNAs"). The microRNA pathway involves microRNAs ("miRNAs"), non-protein coding RNAs generally of between about 19 to about 25 nucleotides (commonly about 20-24 nucleotides in plants) that guide cleavage in trans of target transcripts, negatively regulating the expression of genes involved in various regulation and development pathways; see Ambros et al. (2003). Plant miRNAs have been defined by a set of characteristics including a paired stem-loop precursor that is processed by DCL1 to a single specific .about.21-nucleotide miRNA, expression of a single pair of miRNA and miRNA* species from the double-stranded RNA precursor with two-nucleotide 3' overhangs, and silencing of specific targets in trans (see Bartel (2004); Kim (2005); Jones-Rhoades et al. (2006); Ambros et al. (2003)). In the trans-acting siRNA ("ta-siRNA") pathway, miRNAs serve to guide in-phase processing of siRNA primary transcripts in a process that requires an RNA-dependent RNA polymerase for production of a double-stranded RNA precursor; trans-acting siRNAs are defined by lack of secondary structure, a miRNA target site that initiates production of double-stranded RNA, requirements of DCL4 and an RNA-dependent RNA polymerase (RDR6), and production of multiple perfectly phased .about.21-nt small RNAs with perfectly matched duplexes with 2-nucleotide 3' overhangs (see Allen et al., 2005).
[0050] Many microRNA genes (MIR genes) have been identified and made publicly available in a database ("miRBase", available online at www.microrna.sanger.ac.uk/sequences; also see Griffiths-Jones et al. (2003)). Additional MIR genes and mature miRNAs are also described in U.S. Patent Application Publications 2005/0120415 and 2005/144669, which are incorporated by reference herein. MIR gene families appear to be substantial, estimated to account for 1% of at least some genomes and capable of influencing or regulating expression of about a third of all genes (see, for example, Tomari et al. (2005); Tang (2005); and Kim (2005)). MIR genes have been reported to occur in intergenic regions, both isolated and in clusters in the genome, but can also be located entirely or partially within introns of other genes (both protein-coding and non-protein-coding). For a recent review of miRNA biogenesis, see Kim (2005). Transcription of MIR genes can be, at least in some cases, under control of a MIR gene's own promoter. The primary transcript, termed a "pri-miRNA", can be quite large (several kilobases) and can be polycistronic, containing one or more pre-miRNAs (fold-back structures containing a stem-loop arrangement that is processed to the mature miRNA) as well as the usual 5' "cap" and polyadenylated tail of an mRNA. See, for example, FIG. 1 in Kim (2005).
[0051] A "phased small RNA locus," which transcribes to an RNA transcript forming a single foldback structure that is cleaved in phase in vivo into multiple small double-stranded RNAs (termed "phased small RNAs") capable of suppressing a target gene may also be employed (e.g. U.S. Patent Application Publication 20080066206). In contrast to siRNAs, a phased small RNA transcript is cleaved in phase. In contrast to miRNAs, a phased small RNA transcript is cleaved by DCL4 or a DCL4-like orthologous ribonuclease (not DCL1) to produce multiple abundant small RNAs capable of silencing a target gene. In contrast to the ta-siRNA pathway, the phased small RNA locus transcribes to an RNA transcript that forms hybridized RNA independently of an RNA-dependent RNA polymerase and without a miRNA target site that initiates production of double-stranded RNA. Novel recombinant DNA constructs that are designed based on a phased small RNA locus are useful for suppression of one or multiple target genes, without the use of miRNAs, ta-siRNAs, or expression vectors designed to form a hairpin structure for processing to siRNAs. Furthermore, the recognition sites corresponding to a phased small RNA are useful for suppression of a target sequence in a cell or tissue where the appropriate phased small RNA is expressed endogenously or as a transgene.
A. Virus Targets
[0052] In accordance with the invention, methods and compositions are provided for conferring resistance to multiple viruses to plants, including Solanaceous plants such as tomatoes. Viruses to which resistance may be targeted in the present invention include, without limitation, two or more viruses from among the geminiviruses, tospoviruses, and potexviruses. FIG. 1 illustrates the genome organization of representatives of these viral genera.
[0053] 1. Begomoviruses
[0054] The Geminiviridae are a large, diverse family of plant viruses that infect a broad variety of plants and cause significant crop losses worldwide. They are characterized by twin icosahedral capsids and circular ssDNA genomes that replicate through dsDNA intermediates. Geminiviruses ("begomovirus" and "geminivirus" are used interchangeably herein) depend on the plant nuclear DNA and RNA polymerases for replication and transcription. These viruses contribute only a few factors for their replication and transcription. The family Geminiviridae contains three main genera (formerly termed "subgroups") that differ with respect to insect vector, host range, and genome structure.
[0055] Geminiviridae Subgroup I (genus Mastrevirus) includes leafhopper-transmitted viruses that generally infect monocot plants and have single-component genomes.
[0056] Geminiviridae Subgroup III (genus Begomovirus) includes whitefly-transmitted viruses that infect dicot plants and most commonly have bipartite genomes.
[0057] Geminiviridae Subgroup II (genus Curtovirus) viruses are transmitted by leafhoppers and have single-component genomes like Subgroup I, but infect dicot plants like subgroup III.
[0058] 2. Tospoviruses
[0059] Viruses in the genus Tospovirus cause significant worldwide crop losses. The genus name is derived from Tomato spotted wilt virus ("TSWV"). The Spotted Wilt Disease of tomato was first observed in Australia in 1915 and was later shown to be of viral origin. Until the early 1990s TSWV was considered to be the sole member of the tomato spotted wilt group of plant viruses. The identification and characterization of several similar viruses, including Impatiens necrotic spot virus (INSV), Capsicum chlorosis virus ("CaCV"), Peanut bud necrosis virus (also known as Groundnut bud necrosis virus, "GBNV"), and Tomato chlorotic spot virus led to the creation of the plant-infecting Tospovirus genus within the Bunyaviridae family. This family includes a large group of predominantly animal-infecting viruses. More than twenty tospoviruses have since been identified and characterized and previously unknown species of the genus continue to be described on a regular basis.
[0060] Tospoviruses have a tripartite RNA genome of ambisense polarity. The three portions of the genome are termed the "L" segment, the "M" segment, and the "S" segment. A consensus terminal sequence of each portion of the RNA genome is found, defined by segments UCUCGUUAGC (SEQ ID NO:167) at the 3'end and AGAGCAAUCG (SEQ ID NO:168) at the 5'end. The largest RNA, the "L segment," encodes replicase. The medium size RNA, "M segment," encodes glycoproteins G1 and G2 in the complementary-sense RNA and a nonstructural protein, NSm, in the genome-sense RNA. The smallest segment, "S segment," encodes the nucleocapsid protein (N) in the complementary-sense RNA and a cell-to-cell movement, NSs, in the genome-sense RNA. The virus is transmitted by thrips in the genera Frankliniella (five species) and Thrips (three species). Mechanical transmission of the virus is also possible. TSWV can infect more than 925 plant species belonging to 70 botanical families, whereas the other tospovirus species have much narrower host ranges.
[0061] 3. Potexviruses
[0062] The Pepino mosaic virus (PepMV) is a representative potexvirus from among the Flexiviridae, and is highly contagious with a significant potential to cause damage in protected tomato production. Significant crop losses are possible if action is not taken to eliminate infection. The virus is readily spread via contaminated tools, human hands or clothing, and by direct plant-to-plant contact. It can also be transmitted by grafting or when taking cuttings from infected mother plants. The use of coat protein-mediated resistance may provide good resistance. However, including inverted repeats to the CP may enhance resistant line production.
B. Nucleic Acid Compositions and Constructs
[0063] The invention provides recombinant DNA constructs and methods for use in achieving resistance to multiple (i.e. more than one) viral species and strains from among the begomoviruses, tospoviruses, and potexviruses in transgenic plants. In certain embodiments, resistance is conferred to 2, 3, 4, 5, 6, 7, 8, or more viral species selected from at least two of the following groups: begomoviruses, tospoviruses, and potexviruses. The resistance may be directed by production of siRNA or miRNA, and may also be complemented by protein based approaches such as resistance mediated by expressed coat protein or replicase, mutated forms of replicases, and production of aptamers. Genetically based tolerance (i.e. as identified in a classical breeding approach) may also be utilized.
[0064] As used herein, the term "nucleic acid" refers to a single or double-stranded polymer of deoxyribonucleotide or ribonucleotide bases read from the 5' to the 3' end. The "nucleic acid" may also optionally contain non-naturally occurring or altered nucleotide bases that permit correct read through by a polymerase and do not reduce expression of a polypeptide encoded by that nucleic acid. The term "nucleotide sequence" or "nucleic acid sequence" refers to both the sense and antisense strands of a nucleic acid as either individual single strands or in the duplex. The term "ribonucleic acid" (RNA) is inclusive of dsRNA (double stranded RNA), siRNA (small interfering RNA), shRNA (small hairpin RNA), mRNA (messenger RNA), miRNA (micro-RNA), tRNA (transfer RNA, whether charged or discharged with a corresponding acylated amino acid), and cRNA (complementary RNA) and the term "deoxyribonucleic acid" (DNA) is inclusive of cDNA and genomic DNA and DNA-RNA hybrids. The words "nucleic acid segment," "nucleotide sequence segment," or more generally "segment" will be understood by those in the art as a functional term that includes both genomic sequences, ribosomal RNA sequences, transfer RNA sequences, messenger RNA sequences, operon sequences and smaller engineered nucleotide sequences that express or may be adapted to express, proteins, polypeptides or peptides.
[0065] As used herein, the term "substantially homologous" or "substantial homology," with reference to a nucleic acid sequence, includes a nucleotide sequence that hybridizes under stringent conditions to any of SEQ ID NOs:169-455, or a portion or complement thereof, are those that allow an antiparallel alignment to take place between the two sequences, and the two sequences are then able, under stringent conditions, to form hydrogen bonds with corresponding bases on the opposite strand to form a duplex molecule that is sufficiently stable under conditions of appropriate stringency, including high stringency, to be detectable using methods well known in the art. Substantially homologous sequences may have from about 70% to about 80% sequence identity, or more preferably from about 80% to about 85% sequence identity, or most preferable from about 90% to about 95% sequence identity, to about 99% sequence identity, to the referent nucleotide sequences as set forth the sequence listing, or the complements thereof.
[0066] As used herein, the term "ortholog" refers to a gene in two or more species that has evolved from a common ancestral nucleotide sequence, and may retain the same function in the two or more species.
[0067] As used herein, the term "sequence identity," "sequence similarity" or "homology" is used to describe sequence relationships between two or more nucleotide sequences. The percentage of "sequence identity" between two sequences is determined by comparing two optimally aligned sequences over a comparison window such as the full length of a referenced SEQ ID NO, wherein the portion of the sequence in the comparison window may comprise additions or deletions (i.e., gaps) as compared to the reference sequence (which does not comprise additions or deletions) for optimal alignment of the two sequences. The percentage is calculated by determining the number of positions at which the identical nucleic acid base or amino acid residue occurs in both sequences to yield the number of matched positions, dividing the number of matched positions by the total number of positions in the window of comparison, and multiplying the result by 100 to yield the percentage of sequence identity. A sequence that is identical at every position in comparison to a reference sequence is said to be identical to the reference sequence and vice-versa. A first nucleotide sequence when observed in the 5' to 3' direction is said to be a "complement" of, or complementary to, a second or reference nucleotide sequence observed in the 3' to 5' direction if the first nucleotide sequence exhibits complete complementarity with the second or reference sequence. As used herein, nucleic acid sequence molecules are said to exhibit "complete complementarity" when every nucleotide of one of the sequences read 5' to 3' is complementary to every nucleotide of the other sequence when read 3' to 5'. A nucleotide sequence that is complementary to a reference nucleotide sequence will exhibit a sequence identical to the reverse complement sequence of the reference nucleotide sequence. These terms and descriptions are well defined in the art and are easily understood by those of ordinary skill in the art.
[0068] As used herein, a "comparison window" refers to a conceptual segment of at least 6 contiguous positions, usually about 50 to about 100, more usually about 100 to about 150, in which a sequence is compared to a reference sequence of the same number of contiguous positions after the two sequences are optimally aligned. The comparison window may comprise additions or deletions (i.e. gaps) of about 20% or less as compared to the reference sequence (which does not comprise additions or deletions) for optimal alignment of the two sequences Those skilled in the art should refer to the detailed methods used for sequence alignment, such as in the Wisconsin Genetics Software Package Release 7.0 (Genetics Computer Group, 575 Science Drive Madison, Wis., USA).
[0069] The present invention provides one or more DNA sequences capable of being expressed as an RNA transcript in a cell or microorganism to inhibit target gene expression of at least one plant virus. The sequences comprise a DNA molecule coding for one or more different nucleotide sequences, wherein each of the different nucleotide sequences comprises a sense nucleotide sequence and an antisense nucleotide sequence. The sequences may be connected by a spacer sequence. The spacer sequence can constitute part of the sense nucleotide sequence or the antisense nucleotide sequence or an unrelated nucleotide sequence and forms within the dsRNA molecule between the sense and antisense sequences. The spacer sequence may comprise, for example, a sequence of nucleotides of at least about 10-100 nucleotides in length, or alternatively at least about 100-200 nucleotides in length, at least 200-400 about nucleotides in length, or at least about 400-500 nucleotides in length. The sense nucleotide sequence or the antisense nucleotide sequence may be substantially identical to the nucleotide sequence of the target gene or a derivative thereof or a complementary sequence thereto. The dsDNA molecule may be placed operably under the control of one or more promoter sequences that function in the cell, tissue or organ of the host expressing the dsDNA to produce RNA molecules. As used herein, "expressing" or "expression" and the like refer to transcription of a RNA molecule from a transcribed polynucleotide. The RNA molecule may or may not be translated into a polypeptide sequence.
[0070] The invention also provides a DNA sequence for expression in a cell of a plant that, upon expression of the DNA to RNA and contact with a plant virus achieves suppression of a target viral gene or viral replication or symptomatology (i.e. expression of symptoms). Methods to express a gene suppression molecule in plants are known (e.g. US Publication 2006/0200878 A1; US Publication 2006/0174380; US Publication 2008/0066206; Niu et al., 2006), and may be used to express a nucleotide sequence of the present invention.
[0071] Non-constitutive promoters suitable for use with recombinant DNA constructs of the invention include spatially specific promoters, developmentally specific promoters, and inducible promoters. Spatially specific promoters can include organelle-, cell-, tissue-, or organ-specific promoters (e. g., a plastid-specific, a root-specific, a pollen-specific, or a seed-specific promoter for suppressing expression of the first target RNA in plastids, roots, pollen, or seeds, respectively). In many cases a seed-specific, embryo-specific, aleurone-specific, or endosperm-specific promoter is especially useful. Developmentally specific promoters can include promoters that tend to promote expression during certain developmental stages in a plant's growth cycle, or at different seasons in a year. Inducible promoters include promoters induced by chemicals or by environmental conditions such as, but not limited to, biotic or abiotic stress (e. g., water deficit or drought, heat, cold, high or low nutrient or salt levels, high or low light levels, or pest or pathogen infection). Also of interest are microRNA promoters, especially those having a developmentally specific, spatially specific, or inducible expression pattern. An expression-specific promoter can also include promoters that are generally constitutively expressed but at differing degrees or "strengths" of expression, including promoters commonly regarded as "strong promoters" or as "weak promoters."
[0072] Thus, a gene sequence or fragment for plant virus control according to the invention may be cloned downstream of a promoter or promoters which are operable in a transgenic plant cell and therein expressed to produce mRNA in the transgenic plant cell that form dsRNA molecules. Numerous examples of plant expressible promoters are known in the art (e.g. CaMV 35S; FMV 35S; PC1SV (e.g. U.S. Pat. No. 5,850,019); ScBV; AtAct7, among others). Promoters useful for expression of polypeptides in plants include those that are inducible, viral, synthetic, or constitutive as described in Odell et al. (1985), and/or promoters that are temporally regulated, spatially regulated, and spatio-temporally regulated. A number of organ-specific promoters have been identified and are known in the art (e.g. U.S. Pat. Nos. 5,110,732; 5,837,848; 5,459,252; 6,229,067; Hirel et al. 1992). The dsRNA molecules contained in plant tissues are expressed in a plant so that the intended suppression of the targeted virus gene expression is achieved. The cauliflower mosaic virus 35S promoter, an archetypal strong promoter common in transgenic plant applications, or a related promoter such as the E35S or the FMV promoter, may be employed for driving virus resistance genes. Promoters have also been identified that direct tissue specific gene expression.
[0073] A transgene transcription unit includes DNA sequence encoding a gene of interest. A gene of interest can include any coding or non-coding sequence from a virus species. Non-limiting examples of a non-coding sequence to be expressed by a transgene transcription unit include, but not limited to, 5' untranslated regions, promoters, enhancers, or other non-coding transcriptional regions, 3' untranslated regions, terminators, intron, microRNAs, microRNA precursor DNA sequences, small interfering RNAs, RNA components of ribosomes or ribozymes, small nucleolar RNAs, RNA aptamers capable of binding to a ligand, and other non-coding RNAs.
[0074] Non-limiting examples of a gene of interest further include, but are not limited to, translatable (coding) sequence, such as genes encoding transcription factors and genes encoding enzymes involved in the biosynthesis or catabolism of molecules of interest (such as amino acids, fatty acids and other lipids, sugars and other carbohydrates, biological polymers, and secondary metabolites including alkaloids, terpenoids, polyketides, non-ribosomal peptides, and secondary metabolites of mixed biosynthetic origin). A gene of interest can be a gene native to the cell (e. g., a plant cell) in which the recombinant DNA construct of the invention is to be transcribed, or can be a non-native gene. A gene of interest can be a marker gene, for example, a selectable marker gene encoding antibiotic, antifungal, or herbicide resistance, or a marker gene encoding an easily detectable trait (e.g., in a plant cell, phytoene synthase or other genes imparting a particular pigment to the plant), or a gene encoding a detectable molecule, such as a fluorescent protein, luciferase, or a unique polypeptide or nucleic acid "tag" detectable by protein or nucleic acid detection methods, respectively). Selectable markers are genes of interest of particular utility in identifying successful processing of constructs of the invention.
[0075] Genes of interest include those genes that may be described as "target genes." The target gene can include a single gene or part of a single gene that is targeted for suppression, or can include, for example, multiple consecutive segments of a target gene, multiple non-consecutive segments of a target gene, multiple alleles of a target gene, or multiple target genes from one or more species. The target gene can be translatable (coding) sequence, or can be non-coding sequence (such as non-coding regulatory sequence), or both. The transgene transcription unit can further include 5' or 3' sequence or both as required for transcription of the transgene. In other embodiments (e. g., where it is desirable to suppress a target gene across multiple strains or species, for instance of viruses), it may be desirable to design the recombinant DNA construct to be processed to a mature miRNA for suppressing a target gene sequence common to the multiple strains or species in which the target gene is to be silenced. Thus, the miRNA processed from the recombinant DNA construct can be designed to be specific for one taxon (for example, specific to a genus, family, but not for other taxa.
[0076] The nucleic acid molecules or fragments of the nucleic acid molecules or other nucleic acid molecules in the sequence listing are capable of specifically hybridizing to other nucleic acid molecules under certain circumstances. As used herein, two nucleic acid molecules are said to be capable of specifically hybridizing to one another if the two molecules are capable of forming an anti-parallel, double-stranded nucleic acid structure. A nucleic acid molecule is said to be the complement of another nucleic acid molecule if they exhibit complete complementarity. Two molecules are said to be "minimally complementary" if they can hybridize to one another with sufficient stability to permit them to remain annealed to one another under at least conventional "low-stringency" conditions. Similarly, the molecules are said to be complementary if they can hybridize to one another with sufficient stability to permit them to remain annealed to one another under conventional "high-stringency" conditions. Conventional stringency conditions are described by Sambrook, et al. (1989), and by Haymes et al. (1985).
[0077] Departures from complete complementarity are therefore permissible for, as long as such departures do not completely preclude the capacity of the molecules to form a double-stranded structure. Thus, in order for a nucleic acid molecule or a fragment of the nucleic acid molecule to serve as a primer or probe it needs only be sufficiently complementary in sequence to be able to form a stable double-stranded structure under the particular solvent and salt concentrations employed.
[0078] Appropriate stringency conditions that promote DNA hybridization are, for example, for applications requiring high selectivity, a relatively low salt and/or high temperature condition, such as provided by about 0.02 M to about 0.15 M NaCl at temperatures of about 50.degree. C. to about 70.degree. C. A high stringency condition, for example, is to wash the hybridization filter at least twice with high-stringency wash buffer (0.2.times.SSC or 1.times.SSC, 0.1% SDS, 65.degree. C.). Other conditions, such as 6.0.times.sodium chloride/sodium citrate (SSC) at about 45.degree. C., followed by a wash of 2.0.times.SSC at 50.degree. C., are also known to those skilled in the art or can be found in Current Protocols in Molecular Biology (1989). For example, the salt concentration in the wash step can be selected from a low stringency of about 2.0.times.SSC at 50.degree. C. to a high stringency of about 0.2.times.SSC at 50.degree. C. In addition, the temperature in the wash step can be increased from low stringency conditions at room temperature, about 22.degree. C., to high stringency conditions at about 65.degree. C. Both temperature and salt may be varied, or either the temperature or the salt concentration may be held constant while the other variable is changed. A nucleic acid for use in the present invention may specifically hybridize to one or more of nucleic acid molecules from a plant virus selected from the group consisting of a tospovirus, a begomovirus, and a potexvirus, or complements thereof under such conditions. In specific embodiments, a nucleic acid for use in the present invention will exhibit at least from about 80%, or at least from about 90%, or at least from about 95%, or at least from about 98% or even about 100% sequence identity with one or more nucleic acid molecules as set forth in the sequence listing, or a complement thereof.
[0079] Nucleic acids of the present invention may also be synthesized, either completely or in part, especially where it is desirable to provide plant-preferred sequences, by methods known in the art. Thus, all or a portion of the nucleic acids of the present invention may be synthesized using codons preferred by a selected host. Species-preferred codons may be determined, for example, from the codons used most frequently in the proteins expressed in a particular host species. Other modifications of the nucleotide sequences may result in mutants having slightly altered activity.
[0080] DsRNA or siRNA nucleotide sequences comprise double strands of polymerized ribonucleotide and may include modifications to either the phosphate-sugar backbone or the nucleoside. Modifications in RNA structure may be tailored to allow specific genetic inhibition. In one embodiment, the dsRNA molecules may be modified through an enzymatic process so that siRNA molecules may be generated. Alternatively, a construct may be engineered to express a nucleotide segment for use in an miRNA- or siRNA-mediated resistance approach. The siRNA can efficiently mediate the down-regulation effect for some target genes in some pathogens. This enzymatic process may be accomplished by utilizing an RNAse III enzyme or a DICER enzyme, present in the cells of an insect, a vertebrate animal, a fungus or a plant in the eukaryotic RNAi pathway (Elbashir et al., 2001; Hamilton and Baulcombe, 1999). This process may also utilize a recombinant DICER or RNAse III introduced into the cells of a target insect through recombinant DNA techniques that are readily known to the skilled in the art. Both the DICER enzyme and RNAse III, being naturally occurring in a pathogen or being made through recombinant DNA techniques, cleave larger dsRNA strands into smaller oligonucleotides. The DICER enzymes specifically cut the dsRNA molecules into siRNA pieces each of which is about 19-25 nucleotides in length while the RNAse III enzymes normally cleave the dsRNA molecules into 12-15 base-pair siRNA. The siRNA molecules produced by the either of the enzymes have 2 to 3 nucleotide 3' overhangs, and 5' phosphate and 3' hydroxyl termini. The siRNA molecules generated by RNAse III enzyme are the same as those produced by Dicer enzymes in the eukaryotic RNAi pathway and are hence then targeted and degraded by an inherent cellular RNA-degrading mechanism after they are subsequently unwound, separated into single-stranded RNA and hybridize with the RNA sequences transcribed by the target gene. This process results in the effective degradation or removal of the RNA sequence encoded by the nucleotide sequence of the target gene in the pathogen. The outcome is the silencing of a particularly targeted nucleotide sequence within the pathogen. Detailed descriptions of enzymatic processes can be found in Hannon (2002).
[0081] A nucleotide sequence of the present invention can be recorded on computer readable media. As used herein, "computer readable media" refers to any tangible medium of expression that can be read and accessed directly by a computer. Such media include, but are not limited to: magnetic storage media, such as floppy discs, hard disc, storage medium, and magnetic tape:
[0082] optical storage media such as CD-ROM; electrical storage media such as RAM and ROM; optical character recognition formatted computer files, and hybrids of these categories such as magnetic/optical storage media. A skilled artisan can readily appreciate that any of the presently known computer readable mediums can be used to create a manufacture comprising computer readable medium having recorded thereon a nucleotide sequence of the present invention.
[0083] As used herein, "recorded" refers to a process for storing information on computer readable medium. A skilled artisan can readily adopt any of the presently known methods for recording information on computer readable medium to generate media comprising the nucleotide sequence information of the present invention. A variety of data storage structures are available to a skilled artisan for creating a computer readable medium having recorded thereon a nucleotide sequence of the present invention. The choice of the data storage structure will generally be based on the means chosen to access the stored information. In addition, a variety of data processor programs and formats can be used to store the nucleotide sequence information of the present invention on computer readable medium. The sequence information can be represented in a word processing text file, formatted in commercially-available software such as WordPerfect and Microsoft Word, or represented in the form of an ASCII text file, stored in a database application, such as DB2, Sybase, Oracle, or the like. The skilled artisan can readily adapt any number of data processor structuring formats (e.g. text file or database) in order to obtain computer readable medium having recorded thereon the nucleotide sequence information of the present invention.
[0084] Computer software is publicly available which allows a skilled artisan to access sequence information provided in a computer readable medium. Software that implements the BLAST (Altschul et al., 1990) and BLAZE (Brutlag, et al., 1993) search algorithms on a Sybase system can be used to identify open reading frames (ORFs) within sequences such as the Unigenes and EST's that are provided herein and that contain homology to ORFs or proteins from other organisms. Such ORFs are protein-encoding fragments within the sequences of the present invention and are useful in producing commercially important proteins such as enzymes used in amino acid biosynthesis, metabolism, transcription, translation, RNA processing, nucleic acid and a protein degradation, protein modification, and DNA replication, restriction, modification, recombination, and repair.
[0085] As used herein, a "target," a "target structural motif," or a "target motif," refers to any rationally selected sequence or combination of sequences in which the sequences or sequence(s) are chosen based on a three-dimensional configuration that is formed upon the folding of the target motif or the nucleotide sequence thereof, as appropriate. There are a variety of target motifs known in the art.
C. Nucleic Acid Expression and Target Gene Suppression
[0086] The present invention provides, as an example, a transformed host plant of a pathogenic target organism, transformed plant cells and transformed plants and their progeny. The transformed plant cells and transformed plants may be engineered to express one or more of the dsRNA, miRNA, or mRNA sequences, under the control of a heterologous promoter, described herein to provide a pathogen-protective effect. These sequences may be used for gene suppression in a pathogen, thereby reducing the level or incidence of disease caused by the pathogen on a protected transformed host organism. As used herein the words "gene suppression" are intended to refer to any of the well-known methods for reducing the levels of protein produced as a result of gene transcription to mRNA and subsequent translation of the mRNA.
[0087] Gene suppression is also intended to mean the reduction of protein expression from a gene or a coding sequence including posttranscriptional gene suppression and transcriptional suppression. Posttranscriptional gene suppression is mediated by the homology between of all or a part of a mRNA transcribed from a gene or coding sequence targeted for suppression and the corresponding double stranded RNA used for suppression, and refers to the substantial and measurable reduction of the amount of available mRNA available in the cell for binding by ribosomes or the prevention of translation by the ribosomes. The transcribed RNA can be in the sense orientation to effect what is called co-suppression, in the anti-sense orientation to effect what is called anti-sense suppression, or in both orientations producing a dsRNA to effect what is called RNA interference (RNAi).
[0088] Gene suppression can also be effective against target genes in a plant virus that may contact plant material containing gene suppression agents, specifically designed to inhibit or suppress the expression of one or more homologous or complementary sequences of the virus. Post-transcriptional gene suppression by anti-sense or sense oriented RNA to regulate gene expression in plant cells is disclosed in U.S. Pat. Nos. 5,107,065, 5,759,829, 5,283,184, and 5,231,020. The use of dsRNA to suppress genes in plants is disclosed in WO 99/53050, WO 99/49029, U.S. Publication No. 2003/017596, U.S. Patent Application Publication 2004/0029283.
[0089] A beneficial method of post transcriptional gene suppression versus a plant virus employs both sense-oriented and anti-sense-oriented, transcribed RNA which is stabilized, e.g., as a hairpin or stem and loop structure (e.g. U.S. Publication 2007/0259785). A DNA construct for effecting post transcriptional gene suppression may be one in which a first segment encodes an RNA exhibiting an anti-sense orientation exhibiting substantial identity to a segment of a gene targeted for suppression, which is linked to a second segment encoding an RNA exhibiting substantial complementarity to the first segment. Such a construct forms a stem and loop structure by hybridization of the first segment with the second segment and a loop structure from the nucleotide sequences linking the two segments (see WO94/01550, WO98/05770, US 2002/0048814, and US 2003/0018993). Co-expression with an additional target gene segment may also be employed, as noted above (e.g. WO05/019408).
[0090] According to one embodiment of the present invention, there is provided an exogenous nucleotide sequence (i.e. not naturally found in the genome of the host plant cell), for which expression results in transcription of a RNA sequence that is substantially similar in sequence to a RNA molecule of a targeted gene of a plant virus, selected from the group consisting of a tospovirus, a begomovirus, and a potexvirus, that comprises an RNA sequence encoded by a nucleotide sequence within the genome of the virus. By substantially similar is meant that the exogenous RNA sequence is capable of effecting RNA-mediated gene suppression of a target sequence in a viral genome. Thus, a down-regulation of the expression of the nucleotide sequence corresponding to the target gene is effected.
[0091] In certain embodiments of the invention, expression of a fragment of at least 21 contiguous nucleotides of a nucleic acid sequence of any of SEQ ID NOs:169-455, or complements thereof, may be utilized, including expression of a fragment of up to 21, 36, 60, 100, 550, or 1000 contiguous nucleotides, or sequences displaying 90-100% identity with such sequences, or their complements. In specific embodiments, a nucleotide provided by the invention may comprise a sequence selected from among SEQ ID NOs: 1, 7, 13, 19, 25, 31, 37, 43, 47, 51, 55, 59, 63, 67, 71, 85, 99, 113, 127, and 141. In other specific embodiments, a nucleotide provided by the invention may comprise a sequence selected from among SEQ ID NOs: 379-455. In yet other embodiments, a nucleotide provided by the invention may be described as comprising one or more of nucleotides 1-21, 22-50, 51-100, 101-150, 151-200, 201-250, 251-300, 301-350, 351-400, 401-450, 451-500, 501-550, 551-600, 601-650, 651-700, 701-750, 751-800, 801-850, 851-900, 901-950, 951-1000, 1001-1050, 1051-1100, 1101-1150, 1151-1200, 1201-1250, 1251-1300, 1301-1350, 1351-1400, 1401-1450, 1451-1500, 1501-1550, 1551-1600, 1601-1650, 1651-1700, 1701-1750, 1751-1800, 1801-1850, 1851-1900, 1901-1950, 1951-2000, 2001-2050, 2051-2100, 23-75, 76-125, 126-175, 176-225, 226-275, 276-325, 326-375, 376-425, 426-475, 476-525, 526-575, 576-625, 626-675, 676-725, 726-775, 776-825, 826-875, 876-925, 926-975, 976-1025, 1026-1075, 1076-1125, 1126-1175, 1176-1225, 1226-1275, 1276-1325, 1326-1375, 1376-1425, 1426-1475, 1476-1525, 1526-1575, 1576-1625, 1626-1675, 1676-1725, 1726-1775, 1776-1825, 1826-1875, 1876-1925, 1926-1975, 1976-2025, 2026-2075, 2076-2125, 1-550, 200-750, 300-850, 400-950, 500-1050, 600-1150, 700-1250, 800-1350, 900-1450, 1000-1550, 1100-1650, 1200-1750, 1300-1850, 1400-1950, 1500-2050, up to the full length of the sequence, of one or more of any of SEQ ID NOs:169-455. A sequence complementary to all or a portion of any one or more of SEQ ID NOs:169-455, or SEQ ID NOs: 1, 7, 13, 19, 25, 31, 37, 43, 47, 51, 55, 59, 63, 67, 71, 85, 99, 113, 127, and 141, wherein expression of said sequence suppresses the expression of any one or more of gene(s) encoded by a nucleotide sequence of SEQ ID NOs:169-455, is contemplated. The sequences arrayed for expression to produce dsRNA can be combined with: (1) sequences designed for production of miRNA, including in stacked miRNA cassettes; and/or (2) sequences for inhibition of viral assembly, in order to synergistically inhibit target viruses. Methods for selecting specific sub-sequences as targets for miRNA or siRNA-mediated gene suppression are known in the art (e.g. Reynolds et al., 2004).
[0092] Inhibition of a target gene using dsRNA technology of the present invention is sequence-specific in that nucleotide sequences corresponding to the duplex region of the RNA are targeted for suppression. RNA containing a nucleotide sequences identical to a portion of the target gene transcript is usually preferred for inhibition. RNA sequences with insertions, deletions, and single point mutations relative to the target sequence have also been found to be effective for inhibition. In performance of the present invention, the inhibitory dsRNA and the portion of the target gene may share at least from about 80% sequence identity, or from about 90% sequence identity, or from about 95% sequence identity, or from about 99% sequence identity, or even about 100% sequence identity. Alternatively, the duplex region of the RNA may be defined functionally as a nucleotide sequence that is capable of hybridizing with a portion of the target gene transcript. A less than full length sequence exhibiting a greater homology compensates for a longer less homologous sequence. The length of the identical nucleotide sequences may be at least about 18, 21, 25, 50, 100, 200, 300, 400, 500 or at least about 1000 bases. Normally, a sequence of greater than 20-100 nucleotides should be used, though a sequence of greater than about 200-300 nucleotides may be preferred, and a sequence of greater than about 500-1000 nucleotides may be especially preferred depending on the size of the target gene. The invention has the advantage of being able to tolerate sequence variations that might be expected due to genetic mutation, strain polymorphism, or evolutionary divergence. The introduced nucleic acid molecule may not need to be absolutely homologous to the target sequence, and it may not need to be full length relative to either the primary transcription product or fully processed mRNA of the target gene. Therefore, those skilled in the art need to realize that, as disclosed herein, 100% sequence identity between the RNA and the target gene is not required to practice the present invention.
[0093] Inhibition of target gene expression may be quantified by measuring either the endogenous target RNA or the protein produced by translation of the target RNA and the consequences of inhibition can be confirmed by examination of the outward properties of the cell or organism. Techniques for quantifying RNA and proteins are well known to one of ordinary skill in the art.
[0094] In certain embodiments gene expression is inhibited by at least 10%, by at least 33%, by at least 50%, or by at least 80%. In particular embodiments of the invention gene expression is inhibited by at least 80%, by at least 90%, by at least 95%, or by at least 99% within host cells infected by the virus, such a significant inhibition takes place. Significant inhibition is intended to refer to sufficient inhibition that results in a detectable phenotype (e.g., reduction of symptom expression, etc.) or a detectable decrease in RNA and/or protein corresponding to the target gene being inhibited.
[0095] DsRNA molecules may be synthesized either in vivo or in vitro. The dsRNA may be formed by a single self-complementary RNA strand or from two complementary RNA strands. Endogenous RNA polymerase of the cell may mediate transcription in vivo, or cloned RNA polymerase can be used for transcription in vivo or in vitro. Inhibition may be targeted by specific transcription in an organ, tissue, or cell type; stimulation of an environmental condition (e.g., infection, stress, temperature, chemical inducers); and/or engineering transcription at a developmental stage or age. The RNA strands may or may not be polyadenylated; the RNA strands may or may not be capable of being translated into a polypeptide by a cell's translational apparatus.
[0096] As used herein, the term "disease control agent," or "gene suppression agent" refers, in certain embodiments, to a particular RNA molecule consisting of a first RNA segment and a second RNA segment linked by a third RNA segment. The first and the second RNA segments lie within the length of the RNA molecule and are substantially inverted repeats of each other and are linked together by the third RNA segment. The complementarity between the first and the second RNA segments results in the ability of the two segments to hybridize in vivo and in vitro to form a double stranded molecule, i.e., a stem, linked together at one end of each of the first and second segments by the third segment which forms a loop, so that the entire structure forms into a stem and loop structure, or even more tightly hybridizing structures may form into a stem-loop knotted structure. The first and the second segments correspond invariably, but not necessarily respectively, to a sense and an antisense sequence homologous with respect to the target RNA transcribed from the target gene in the target virus that is intended to be suppressed by the dsRNA molecule.
[0097] As used herein, the term "genome" as it applies to a plant virus or a host encompasses not only viral DNA or RNA, or chromosomal DNA found within the nucleus, but organelle DNA found within subcellular components of the cell. The DNA's of the present invention introduced into plant cells can therefore be either chromosomally integrated or organelle-localized. The term "genome" as it applies to bacteria encompasses both the chromosome and plasmids within a bacterial host cell. The DNA's of the present invention introduced into bacterial host cells can therefore be either chromosomally integrated or plasmid-localized.
[0098] It is envisioned that the compositions of the present invention can be incorporated within the seeds of a plant species either as a product of expression from a recombinant gene incorporated into a genome of the plant cells. The plant cell containing a recombinant gene is considered herein to be a transgenic event.
D. Recombinant Vectors and Host Cell Transformation
[0099] A recombinant DNA vector may, for example, be a linear or a closed circular plasmid. The vector system may be a single vector or plasmid or two or more vectors or plasmids that together contain the total DNA to be introduced into the genome of the bacterial host. In addition, a bacterial vector may be an expression vector. Nucleic acid molecules as set forth in the sequence listing, or complements or fragments thereof, can, for example, be suitably inserted into a vector under the control of a suitable promoter that functions in one or more microbial hosts to drive expression of a linked coding sequence or other DNA sequence. Many vectors are available for this purpose, and selection of the appropriate vector will depend mainly on the size of the nucleic acid to be inserted into the vector and the particular host cell to be transformed with the vector. Each vector contains various components depending on its function (amplification of DNA or expression of DNA) and the particular host cell with which it is compatible. The vector components for bacterial transformation generally include, but are not limited to, one or more of the following: a signal sequence, an origin of replication, one or more selectable marker genes, and an inducible promoter allowing the expression of exogenous DNA.
[0100] Expression and cloning vectors generally contain a selection gene, also referred to as a selectable marker. This gene encodes a protein necessary for the survival or growth of transformed host cells grown in a selective culture medium. Typical selection genes encode proteins that (a) confer resistance to antibiotics or other toxins, e.g., ampicillin, neomycin, methotrexate, or tetracycline, (b) complement auxotrophic deficiencies, or (c) supply critical nutrients not available from complex media, e.g., the gene encoding D-alanine racemase for Bacilli. Those cells that are successfully transformed with a heterologous protein or fragment thereof produce a protein conferring drug resistance and thus survive the selection regimen.
[0101] An expression vector for producing a mRNA can also contain an inducible promoter that is recognized by a host organism and is operably linked to the nucleic acid. The term "operably linked," as used in reference to a regulatory sequence and a structural nucleotide sequence, means that the regulatory sequence causes regulated expression of the linked structural nucleotide sequence. "Regulatory sequences" or "control elements" refer to nucleotide sequences located upstream (5' noncoding sequences), within, or downstream (3' non-translated sequences) of a structural nucleotide sequence, and which influence the timing and level or amount of transcription, RNA processing or stability, or translation of the associated structural nucleotide sequence. Regulatory sequences may include promoters, translation leader sequences, introns, enhancers, stem-loop structures, repressor binding sequences, and polyadenylation recognition sequences and the like.
[0102] Construction of suitable vectors containing one or more of the above-listed components employs standard recombinant DNA techniques. Isolated plasmids or DNA fragments are cleaved, tailored, and re-ligated in the form desired to generate the plasmids required. Examples of available bacterial expression vectors include, but are not limited to, the multifunctional E. coli cloning and expression vectors such as Bluescript.TM. (Stratagene, La Jolla, Calif.), in which, for example, a nucleic acid, or fragment thereof may be ligated into the vector in frame with sequences for the amino-terminal Met and the subsequent 7 residues of .beta.-galactosidase so that a hybrid protein is produced; pIN vectors (Van Heeke and Schuster, 1989); and the like.
[0103] The present invention also contemplates transformation of a nucleotide sequence of the present invention into a plant to achieve virus-inhibitory levels of expression of one or more RNA molecules. A transformation vector can be readily prepared using methods available in the art. The transformation vector comprises one or more nucleotide sequences that is/are capable of being transcribed to an RNA molecule and that is/are substantially homologous and/or complementary to one or more nucleotide sequences encoded by the genome of the target virus or viruses, such that upon contact of the RNA transcribed from the one or more nucleotide sequences by the target plant-parasitic virus, there is down-regulation of expression of at least one of the respective nucleotide sequences of the genome of the virus.
[0104] The transformation vector may be termed a dsDNA construct and may also be defined as a recombinant molecule, a disease control agent, a genetic molecule or a chimeric genetic construct. A chimeric genetic construct of the present invention may comprise, for example, nucleotide sequences encoding one or more antisense transcripts, one or more sense transcripts, one or more of each of the aforementioned, wherein all or part of a transcript there from is homologous to all or part of an RNA molecule comprising an RNA sequence encoded by a nucleotide sequence within the genome of a plant virus.
[0105] In one embodiment a plant transformation vector comprises an isolated and purified DNA molecule comprising a heterologous promoter operatively linked to one or more nucleotide sequences of the present invention. The nucleotide sequence includes a segment coding all or part of an RNA present within a targeted RNA transcript and may comprise inverted repeats of all or a part of a targeted RNA. The DNA molecule comprising the expression vector may also contain a functional intron sequence positioned either upstream of the coding sequence or even within the coding sequence, and may also contain a five prime (5') untranslated leader sequence (i.e., a UTR or 5'-UTR) positioned between the promoter and the point of translation initiation.
[0106] A plant transformation vector may contain sequences from more than one gene, thus allowing production of more than one dsRNA, miRNA, or siRNA for inhibiting expression of genes of the more than one target virus. For instance a vector or construct may comprise up to about 8 or 10, or more, nucleic acid segments for transcription of antiviral sequences, as shown in FIG. 10. One skilled in the art will readily appreciate that segments of DNA whose sequence corresponds to that present in different genes can be combined into a single composite DNA segment for expression in a transgenic plant. Alternatively, a plasmid of the present invention already containing at least one DNA segment can be modified by the sequential insertion of additional DNA segments between the enhancer and promoter and terminator sequences. In the disease control agent of the present invention designed for the inhibition of multiple genes, the genes to be inhibited can be obtained from the same plant viral strain or species in order to enhance the effectiveness of the control agent. In certain embodiments, the genes can be derived from different plant viruses in order to broaden the range of viruses against which the agent(s) is/are effective. When multiple genes are targeted for suppression or a combination of expression and suppression, a polycistronic DNA element can be fabricated as illustrated and disclosed in U.S. Publication No. 2004/0029283.
[0107] A recombinant DNA vector or construct of the present invention may comprise a selectable marker that confers a selectable phenotype on plant cells. Selectable markers may also be used to select for plants or plant cells that contain the exogenous nucleic acids encoding polypeptides or proteins of the present invention. The marker may encode biocide resistance, antibiotic resistance (e.g., kanamycin, G418 bleomycin, hygromycin, etc.), or herbicide resistance (e.g., glyphosate, etc.). Examples of selectable markers include, but are not limited to, a neo gene which codes for kanamycin resistance and can be selected for using kanamycin, G418, etc., a bar gene which codes for bialaphos resistance; a mutant EPSP synthase gene which encodes glyphosate resistance; a nitrilase gene which confers resistance to bromoxynil; a mutant acetolactate synthase gene (ALS) which confers imidazolinone or sulfonylurea resistance; and a methotrexate resistant DHFR gene. Multiple selectable markers are available that confer resistance to ampicillin, bleomycin, chloramphenicol, gentamycin, hygromycin, kanamycin, lincomycin, methotrexate, phosphinothricin, puromycin, spectinomycin, rifampicin, and tetracycline, and the like. Examples of such selectable markers are illustrated in U.S. Pat. Nos. 5,550,318; 5,633,435; 5,780,708 and 6,118,047.
[0108] A recombinant vector or construct of the present invention may also include a screenable marker. Screenable markers may be used to monitor expression. Exemplary screenable markers include a .beta.-glucuronidase or uidA gene (GUS) which encodes an enzyme for which various chromogenic substrates are known (Jefferson et al., 1987); one or more of the various fluorescent proteins (FP) genes such as green fluorescent protein (GFP), red fluorescent protein (RFP) or any one of a large family of proteins which typically fluoresce at a characteristic wavelength; an R-locus gene, which encodes a product that regulates the production of anthocyanin pigments (red color) in plant tissues (Dellaporta et al., 1988); a .beta.-lactamase gene (Sutcliffe et al., 1978), a gene which encodes an enzyme for which various chromogenic substrates are known (e.g., PADAC, a chromogenic cephalosporin); a luciferase gene (Ow et al., 1986) a xylE gene (Zukowski et al., 1983) which encodes a catechol dioxygenase that can convert chromogenic catechols; an .alpha.-amylase gene (Ikatu et al., 1990); a tyrosinase gene (Katz et al., 1983) which encodes an enzyme capable of oxidizing tyrosine to DOPA and dopaquinone which in turn condenses to melanin; an .alpha.-galactosidase, which catalyzes a chromogenic .alpha.-galactose substrate.
[0109] Plant transformation vectors for use with the present invention may for instance include those derived from a Ti plasmid of Agrobacterium tumefaciens (e.g. U.S. Pat. Nos. 4,536,475, 4,693,977, 4,886,937, 5,501,967 and EP 0 122 791). Agrobacterium rhizogenes plasmids (or "Ri") are also useful and known in the art. Other preferred plant transformation vectors include those disclosed, e.g., by Herrera-Estrella (1983); Bevan (1983), Klee (1985) and EP 0 120 516.
[0110] In certain embodiments, a functional recombinant DNA may be introduced at a non-specific location in a plant genome. In special cases it may be useful to insert a recombinant DNA construct by site-specific integration. Several site-specific recombination systems exist which are known to function in plants include cre-lox as disclosed in U.S. Pat. No. 4,959,317 and FLP-FRT as disclosed in U.S. Pat. No. 5,527,695.
[0111] Suitable methods for transformation of host cells for use with the current invention are believed to include virtually any method by which DNA can be introduced into a cell (see, for example, Miki et al., 1993), such as by transformation of protoplasts (U.S. Pat. No. 5,508,184; Omirulleh et al., 1993), by desiccation/inhibition-mediated DNA uptake (Potrykus et al., 1985), by electroporation (U.S. Pat. No. 5,384,253), by agitation with silicon carbide fibers (Kaeppler et al., 1990; U.S. Pat. Nos. 5,302,523; and 5,464,765), by Agrobacterium-mediated transformation (U.S. Pat. Nos. 5,563,055; 5,591,616; 5,693,512; 5,824,877; 5,981,840; 6,384,301) and by acceleration of DNA coated particles (U.S. Pat. Nos. 5,015,580; 5,550,318; 5,538,880; 6,160,208; 6,399,861; 6,403,865; Padgette et al. 1995), etc. Through the application of techniques such as these, the cells of virtually any species may be stably transformed. In the case of multicellular species, the transgenic cells may be regenerated into transgenic organisms.
[0112] The most widely utilized method for introducing an expression vector into plants is based on the natural transformation system of Agrobacterium (for example, Horsch et al., 1985). A. tumefaciens and A. rhizogenes are plant pathogenic soil bacteria which genetically transform plant cells. The Ti and Ri plasmids of A. tumefaciens and A. rhizogenes, respectively, carry genes responsible for genetic transformation of the plant. Descriptions of Agrobacterium vector systems and methods for Agrobacterium-mediated gene transfer are provided by numerous references, including Gruber et al. 1993; Miki et al., 1993, Moloney et al., 1989, and U.S. Pat. Nos: 4,940,838 and 5,464,763. Other bacteria such as Sinorhizobium, Rhizobium, and Mesorhizobium that interact with plants naturally can be modified to mediate gene transfer to a number of diverse plants. These plant-associated symbiotic bacteria can be made competent for gene transfer by acquisition of both a disarmed Ti plasmid and a suitable binary vector (Broothaerts et al., 2005). Methods for introducing virus sequences to plants may also be used (e.g. Grimsley, 1990, Boulton, 1996).
[0113] Methods for the creation of transgenic plants and expression of heterologous nucleic acids in plants in particular are known and may be used with the nucleic acids provided herein to prepare transgenic plants that exhibit reduced susceptibility to plant-pathogenic viruses. Plant transformation vectors can be prepared, for example, by inserting the dsRNA producing nucleic acids disclosed herein into plant transformation vectors and introducing these into plants. One known vector system has been derived by modifying the natural gene transfer system of Agrobacterium tumefaciens. The natural system comprises large Ti (tumor-inducing)-plasmids containing a large segment, known as T-DNA, which is transferred to transformed plants. Another segment of the Ti plasmid, the vir region, is responsible for T-DNA transfer. The T-DNA region is bordered by terminal repeats. In the modified binary vectors the tumor-inducing genes have been deleted and the functions of the vir region are utilized to transfer foreign DNA bordered by the T-DNA border sequences. The T-region may also contain a selectable marker for efficient recovery of transgenic plants and cells, and a multiple cloning site for inserting sequences for transfer such as a dsRNA encoding nucleic acid.
[0114] Protocols for transformation of tomato cells are known in the art (e.g. McCormick, 1991). Alternate plant transformation protocols are discussed in Boulton (1996), and Grimsley (1990). Transformation and regeneration protocols for other plants, such as pepper, are known in the art (e.g. Christopher and Rajam, 1996; U.S. Pat. No. 5,262,316; Liu et al. 1990). For instance, such a protocol for transformation of tomato could include well known steps of seed sterilization, seed germination and growth, explanting of seedlings, Agrobacterium culture growth and preparation, co-cultivation, selection, and regeneration.
E. Transgenic Plants and Cells
[0115] A transgenic plant formed using Agrobacterium transformation methods typically may contain a single simple recombinant DNA sequence inserted into one chromosome, referred to as a transgenic event. Such transgenic plants can be referred to as being heterozygous for the inserted exogenous sequence. A transgenic plant homozygous with respect to a transgene can be obtained by sexually mating (selfing) an independent segregant transgenic plant that contains a single exogenous gene sequence to itself, for example an F0 plant, to produce F1 seed. One fourth of the F1 seed produced will be homozygous with respect to the transgene. Germinating F1 seed results in plants that can be tested for heterozygosity, typically using a SNP assay or a thermal amplification assay that allows for the distinction between heterozygotes and homozygotes (i.e., a zygosity assay). Crossing a heterozygous plant with itself or another heterozygous plant results in heterozygous progeny, as well as homozygous transgenic and homozygous null progeny.
[0116] In addition to direct transformation of a plant with a recombinant DNA construct, transgenic plants can be prepared by crossing a first plant having a recombinant DNA construct with a second plant lacking the construct. For example, recombinant DNA for gene suppression can be introduced into first plant line that is amenable to transformation to produce a transgenic plant that can be crossed with a second plant line to introgress the recombinant DNA for gene suppression into the second plant line.
F. Virus Resistance Screens
[0117] Inoculation and disease testing with viruses such as TSWV, TYLCV, and PepMV was performed using either mechanical transmission or agroinfection (e.g. Boulton, 1996; Grimsley, 1990). TSWV infection was accomplished mechanically, essentially as described by Kumar et al., (1993) with some modifications. Inoculation with begomovirus (e.g. TYLCV) and disease testing of plants was accomplished by agroinfection as follows: Seeds of tomato (Lycopersicon esculentum) cvs. (Resistant (R): HP919; Intermediate Resistant (IR): Hilario; Susceptible (S): Arletta) were sown and approximately 20 plants for each inoculation, as well as a non-inoculated or mock-inoculated control, were grown for 7-10 days, to the cotyledon stage with no primary leaf visible. The lower sides of the cotyledons were then infiltrated using a needleless syringe, and additional inoculations were subsequently performed by injection into stems at the 2-3 true leaf stage, approximately 1-2 weeks later. Infiltration or injection utilized a transformed A. tumefaciens strain, induced with acetosyringone, containing an infections clone of the virus that had been grown in YEB broth with antibiotics for selection of the pBIN vector for about 15 hours at 28.degree. C. on a shaker (170 RPM) from inoculation of a fresh culture. Following growth at 28.degree. C., the culture was spun down and resuspended in 10 ml MMA (Per 1 liter: 20 g sucrose, 5 g MS salts, 1.95 g MES, pH 5.6 with NaOH, and 1 ml of 200 mM acetosyringone stock; dH.sub.2O to 1 liter). Plants were grown at 20-25.degree. C. (day/night), 16 hours light in the greenhouse or growth chamber. After 6 weeks, symptoms were scored as follows:
[0118] 1 no symptoms visible (resistant)
[0119] 3. very mild symptoms (resistant)
[0120] 5. medium symptoms
[0121] 7. strong symptoms
[0122] 9. very strong symptoms
[0123] Inoculation with the potexvirus Pepino mosaic virus (PepMV) was accomplished as follows: Virus inoculum was prepared by mechanically infecting tomato seedlings (susceptible cv. Apollo, 9-12 days old) following dusting of leaves with carborundum powder. Infected tissues were harvested and 1 g of infected tissue was homogenized with 5-10 ml of phosphate buffer (pH 9). This prepared inoculum was then used to mechanically inoculate experimental plants at the cotyledon stage (7-10 days after sowing). Inoculated plants were grown in the greenhouse at 19-23.degree. C. (day/night), with 16 hours of light per 24 hours, in greenhouse with 70% relative humidity. Plants were evaluated at 11-21 days after inoculation, and scored as follows:
[0124] 1. no symptoms visible (resistant)
[0125] 3. some foliar chlorosis
[0126] 5. chlorosis in foliar veins and/or foliar mosaic
[0127] 7. foliar vein chlorosis and spiky leaves
[0128] 9. foliar vein chlorosis and spiky leaves and yellow mosaic
G. RNA Extraction and siRNA Northern Blots
[0129] RNA was extracted for small RNA northern blotting using Trizol.RTM. essentially according to the manufacturer's directions (Invitrogen). Briefly, approximately 100-200 mg of fresh leaf tissue was ground in liquid nitrogen in 1.5 ml centrifuge tube placed on dry ice. Sample was removed from dry ice and 1 ml of Trizol was added under a fume hood. This was mixed well and incubated at room temperature (RT) for 10 minutes. Next, 0.2 ml of chloroform was added, and the sample was shaken by hand for 30 seconds and centrifuged at 13000 rpm in a refrigerated table top centrifuge for 15 minutes at 4.degree. C.
[0130] The aqueous phase was transferred to a new tube, 0.5 ml of isopropanol was added, and tubes were inverted a few times, and then incubated for 10 min at RT followed by centrifugation at 13000 rpm in a refrigerated table top centrifuge for 15 min at 4.degree. C. The supernatant was discarded and the pellet was washed by adding 0.75 ml of 75% ethanol, then centrifuged at 13000 rpm for 10 min at 4.degree. C. The supernatant was discarded and RNA was air dried for 10 minutes at room temperature.
[0131] The pellet was resuspended by adding 100 .mu.l of RNAse free ddH2O and was vortexed for a few seconds. The samples were frozen on dry ice and the RNA concentrated by the use of a speed vacuum for about 20 minutes: this step removes all traces of ethanol. RNA was quantified by spectrophotometer (usually about 80-100 .mu.g are recovered from tomato leaf). About 7 .mu.g of total RNA were loaded for siRNA analysis.
[0132] siRNA Northern blot with DIG labeled probe: Pre-run the 15% TBE-Urea gel (Invitrogen EC68855BOX) for about 30' at 110 volts. Samples were prepared for loading by adding 7 .mu.l of Novex.RTM. TBE-Urea Sample Buffer 2.times. (Invitrogen LC6876) to 7 .mu.l of total RNA (5-7 .mu.g), they are denatured for about 10' at 94.degree. C. and immediately cooled on ice. If sample visualization under UV light was needed, ethidium bromide was added to the sample buffer (1 .mu.l of EtBr @0.624 mg/ml for every 100 .mu.l of buffer). Samples were briefly spun down before loading into the gel wells and electrophoresis was carried on for about 1.5 hrs at 180 Volts in 0.5.times.TBE until the blue dye reached the bottom of the gel. After the run was terminated, the gel was observed under UV lights.
[0133] Transfer of the gel to Nytran Supercharge Nylon membrane (VWR 28151-318) was done in a Transblot semi dry transfer cell (BioRad 170-3940): the membrane was pre-wet in water and equilibrated in 0.5.times.TBE together with 2 pieces of extra-thick paper (BioRad 170-3968). The transfer was set up according to manufacturer's instruction (anode-blotting paper-membrane-gel-blotting paper-cathode) and carried on for 50' at 380 mAmp. After transfer the membrane was left to air dry for about 10' and then the RNA was cross-linked to it in a Stratalinker 1800 (Stratagene).
[0134] Hybridization: The membrane was pre-hybridized at 42.degree. C. for 1 hr with 10 ml of PerfectHyb solution (Sigma H7033) in a hybridization oven. 200 ng of DIG labeled probe, prepared with the PCR DIG labeling mix (Roche 11585550910) following manufacturer's instruction, was denatured for 10' at 94.degree. C., cooled on ice, and added to 10 ml of fresh hybridization solution, which was added to the pre-hybridized membrane and incubated overnight at 42.degree. C. in the hybridization oven.
[0135] Washes and detection: After discarding the hybridization solution, the membranes was briefly rinsed in 2.times.SSC 0.1% SDS; and then washed 2 times in pre-warmed 2.times.SSC 0.1% SDS for 20' at 50.degree. C., 2 times in pre-warmed 1.times.SSC 0.1% SDS for 20' at 50.degree. C., 2 times in pre-warmed 0.5.times.SSC 0.1% SDS for 20' at 50.degree. C. Detection was performed following the instructions provided with the DIG Wash and Block Buffer Set (Roche 11585762001). Briefly, the membrane was rinsed for 2' in 1.times.DIG Washing Buffer before incubating for 1 hr at RT in 100 mL DIG Blocking Solution (10 mL 10.times. Blocking buffer+10 mL 10.times. Maleic Acid+80 mL water). It was then incubated for 1 hr at room temperature in 100 mL fresh DIG Blocking Solution to which 10 .mu.l of Anti-Digoxigenin-AP antibody (Roche 11093274910) had been added. The membrane was washed 15' at RT twice 100 mL 1.times.DIG Washing Buffer and equilibrated with 100 mL of 1.times.DIG Detection Buffer. About 1 mL of CDP-star solution (Roche 12041677001) was distributed across the top of the membrane, which was placed between 2 plastic sheets, incubated at RT for 5' and exposed to a film for variable amounts of time before developing.
H. Validation of Constructs in Transgenic Tomato Plants
[0136] The engineered sequences for generating dsRNA or miRNA were cloned into binary vectors for tomato transformation. R.sub.0 transgenic plants were assayed for the production of the specific .about.21mers and/or virus resistance efficacy by small RNA northern blot analysis and resistance assays as outlined above and as described in Example 7. Other selected dsRNA-generating sequences or miRNA-generating sequences, for instance from among any of SEQ ID NOs:1-154, or from within SEQ ID NOs:169-455 or portions thereof, may be utilized to create additional efficacious constructs. Thus, for instance, MIR backbone sequence(s), for instance MON1 from soybean, MON5 from soybean, MON13 from rice, MON18 from maize, miR159a from maize, or mi167g from maize (SEQ ID NOs:155, 157, 159, 161, 163, or 165) may be utilized to create dsRNA- or miRNA-generating sequence(s). In certain embodiments, the dsRNA-generating sequence or miRNA-generating sequence may comprise any of SEQ ID NOs:1, 7, 13, 19, 25, 31, 37, 43, 47, 55, 59, 63, 67, 71, 85, 99, 113, 127, 141, or 379-455. In particular embodiments, the sequence may comprise any of SEQ ID NOs:156, 158, 160, 162, 164, 166, 363, 364, 365, 366, 367, 368, 369, 370, 371, 372, 373, 374, or 375.
[0137] The present invention includes combinations with other disease control traits in a plant, including non-transgenic approaches, to achieve desired traits for enhanced control of plant disease. Combining disease control traits that employ distinct modes-of-action can provide protected transgenic plants with superior durability over plants harboring a single control trait because of the reduced probability that resistance will develop in the field. Thus, in certain embodiments, at least two or at least three modes of action are employed to confer virus resistance, wherein such modes of action are selected from the group consisting of expression of dsRNA, expression of miRNA, inhibition of virion assembly, phenotypic expression of a non-transgenic virus resistance trait, and transgenic protein expression. In particular embodiments, transgenic protein expression comprises expression of a coat protein-encoding nucleic acid sequence, expression of a movement protein-encoding sequence or expression of a replicase-encoding nucleic acid sequence.
[0138] The invention also relates to commodity products containing one or more of the sequences of the present invention, and produced from a recombinant plant or seed containing one or more of the nucleotide sequences of the present invention are specifically contemplated as embodiments of the present invention. A commodity product containing one or more of the sequences of the present invention is intended to include, but not be limited to, meals, oils, crushed or whole grains or seeds of a plant, or any food product comprising any meal, oil, or crushed or whole grain of a recombinant plant or seed containing one or more of the sequences of the present invention. The detection of one or more of the sequences of the present invention in one or more commodity or commodity products contemplated herein is defacto evidence that the commodity or commodity product is composed of a transgenic plant designed to express one or more of the nucleotides sequences of the present invention for the purpose of controlling plant disease using dsRNA mediated gene suppression methods.
I. Nucleic Acid Compositions
[0139] The present invention provides methods for obtaining a nucleic acid comprising a nucleotide sequence for producing a dsRNA, miRNA, or siRNA. In one embodiment, such a method comprises: (a) analyzing one or more target gene(s) for their expression, function, and phenotype upon dsRNA-mediated, or mi-RNA- or siRNA-mediated suppression of a gene of a plant pathogenic virus; (b) probing a nucleic acid library with a hybridization probe comprising all or a portion of a nucleotide sequence, or a homolog thereof, from a targeted virus that displays an altered phenotype in a dsRNA-mediated suppression analysis; (c) identifying a DNA clone that hybridizes with the hybridization probe; (d) isolating the DNA clone identified in step (b); and (e) sequencing the nucleic acid fragment that comprises the clone isolated in step (d) wherein the sequenced nucleic acid molecule transcribes all or a substantial portion of the RNA nucleotide acid sequence or a homolog thereof. The RNA-mediated resistance approach utilizing dsRNA, miRNA, or siRNA may be supplemented by placing antiviral sequences into the loop of a dsRNA-encoding sequence, or by inserting a sequence that encodes an efficacious dsRNA or miRNA into an intron of a polypeptide expression cassette (intronic dsRNA/miRNA; Frizzi et al., 2008) (e.g. FIG. 9). For example, viral protein(s) such as coat protein and/or replicase may be expressed in a transgenic plant that also expresses an efficacious dsRNA, miRNA, or siRNA, without use of an additional transgene cassette, by inserting protein coding sequence into the loop of a dsRNA.
[0140] In another embodiment, a method of the present invention for obtaining a nucleic acid fragment comprising a nucleotide sequence for producing a substantial portion of a dsRNA or siRNA comprises: (a) synthesizing a first and a second oligonucleotide primers corresponding to a portion of one of the nucleotide sequences from a targeted pathogen; and (b) amplifying a nucleic acid insert present in a cloning vector using the first and second oligonucleotide primers of step (a) wherein the amplified nucleic acid molecule transcribes a substantial portion of the a substantial portion of a dsRNA or siRNA of the present invention.
[0141] In practicing the present invention, target genes may be derived from a begomovirus, tospovirus, or potexvirus. It is contemplated that several criteria may be employed in the selection of preferred target genes. Such sequences may be identified by aligning, for instance, begomovirus or tospovirus sequences from multiple strains and/or species. A bioinformatics approach has identified numerous 21-mers that are mostly conserved in more than 100 begomovirus strains and species. In some instances, mismatches within a particular 21-mer are allowed for broader targeting.
[0142] As used herein, the term "derived from" refers to a specified nucleotide sequence that may be obtained from a particular specified source or species, albeit not necessarily directly from that specified source or species.
[0143] In one embodiment, a gene is selected that results in suppression of viral replication and/or symptomatology. Other target genes for use in the present invention may include, for example, those that play important roles in viral transmission, movement within a plant, or virion assembly (e.g. tospovirus terminal sequences, comprising the terminal repeat sequences). According to one aspect of the present invention for virus control, the target sequences may essentially be derived from the targeted plant viruses. Some of the exemplary target sequences cloned from a begomovirus, tospovirus, or potexvirus may be found in the Sequence Listing, for instance in SEQ ID NOs: 1, 7, 13, 19, 25, 31, 37, 43, 47, 51, 55, 59, 63, 67, 71, 85, 99, 113, 127, and 141, as well as within SEQ ID NOs:169-455.
[0144] For the purpose of the present invention, the dsRNA molecules may be obtained by polymerase chain (PCR) amplification of a target gene sequences derived from a gDNA or cDNA library or portions thereof. The DNA library may be prepared using methods known to the ordinary skilled in the art and DNA/RNA may be extracted. Genomic DNA or cDNA libraries generated from a target organism may be used for PCR amplification for production of the dsRNA or siRNA.
[0145] The target gene sequences may be then be PCR amplified and sequenced using the methods readily available in the art. One skilled in the art may be able to modify the PCR conditions to ensure optimal PCR product formation. The confirmed PCR product may be used as a template for in vitro transcription to generate sense and antisense RNA with the included minimal promoters. In one embodiment, the present invention comprises isolated and purified nucleotide sequences that may be used as plant-virus control agents. The isolated and purified nucleotide sequences may comprise those as set forth in the sequence listing.
[0146] As used herein, the phrase "coding sequence," "structural nucleotide sequence" or "structural nucleic acid molecule" refers to a nucleotide sequence that is translated into a polypeptide, usually via mRNA, when placed under the control of appropriate regulatory sequences. The boundaries of the coding sequence are determined by a translation start codon at the 5'-terminus and a translation stop codon at the 3'-terminus. A coding sequence can include, but is not limited to, genomic DNA, cDNA, EST and recombinant nucleotide sequences.
[0147] The term " recombinant DNA" or "recombinant nucleotide sequence" refers to DNA that contains a genetically engineered modification through manipulation via mutagenesis, restriction enzymes, and the like.
EXAMPLES
[0148] The inventors herein have identified a means for controlling virus infections in plants by incorporating into plants engineered miRNAs, ta-siRNAs and/or phased siRNAs. Any one or any combination of these attributes can result in an effective inhibition of plant infection, inhibition of plant disease, and/or reduction in severity of disease symptoms.
Example 1
Targets for Multivirus Resistance
[0149] Sequences of targeted viruses were assembled from Genbank. FIG. 1 schematically shows the genome organization of representative targeted viruses. For targeted begomoviruses, 58 isolates of Tomato yellow leaf curl virus (TYLCV), 30 isolates of Tomato leaf curl New Delhi virus (ToLCNDV), 5 isolates of Tomato severe leaf curl virus (ToSLCV), 5 isolates of Pepper golden mosaic virus (PepGMV), and 2 isolates of Pepper huasteco yellow vein virus (PHYVV) were analyzed. For targeted tospoviruses, Tomato spotted wilt virus (TSWV), Groundnut bud necrosis virus (GBNV) and Capsicum chlorosis virus (CaCV), 6 L segments (TSWV: 4; GBNV: 1; CaCV: 1), 23 M segments (TSWV: 20; GBNV: 2; CaCV: 1), and 45 S segments (TSWV: 41; GBNV: 2; CaCV: 2) were analyzed. For isolates of the targeted potexvirus, 13 isolates of Pepino mosaic virus (PepMV) were analyzed (e.g. Lopez et al., 2005; Cotillon et al., 2002). A bioinformatics approach was utilized to identify approximately 20-24 nucleotide sequences to be used as artificial miRNAs to suppress as many of the targeted viruses as possible.
[0150] The following Table (Table 1) lists sequences analyzed for possible use in designing transgene constructs for generating engineered miRNAs, as well as for identifying sequences for use in dsRNA-mediated viral resistance approaches:
TABLE-US-00001 TABLE 1 Exemplary sequences used for bioinformatics analysis (SEQ ID NOs: 169-362) Virus type Virus name GenBank Accession Begomo- Tomato yellow leaf AF024715, X15656, X76319, virus curl virus (TYLCV) AB014346, AB014347, AB110217, AB110218, AB116629, AB16630, AB116631, AB116632, AB116633, AB116634, AB116635, AB116636, AF071228, AF105975, AF271234, AJ132711, AJ223505, AJ489258, AJ519441, AJ812277, AJ865337, AM282874, AM409201, AM698117, AM698118, AM698119, AY044138, AY134494, AY227892, AY502934, AY530931, AY594174, AY594175. DQ144621, DQ358913, DQ631892, DQ644565, EF051116, EF054893, EF054894, EF060196, EF101929, EF107520, EF110890, EF158044, EF185318, EF210554, EF210555, EF433426, EF523478, EF539831, EU031444, EU143745 Begomo- Tomato leaf curl NC_004611, U15015, U15016, virus New Delhi virus Y16421, AB330079, AB368447, (ToLCNDV) AB368448, AF102276, AF448058, AF448059, AJ620187, AJ875157, AM286433, AM286434, AM292302, AM850115, AY286316, AY428769, AY939926, DQ116880, DQ116883, DQ116885, DQ169056, EF035482, EF043230, EF043231, EF063145, EF068246, EF450316, EF620534, EU309045 Begomo- Tomato severe AF130415, AJ508784, AJ508785, virus leaf curl DQ347946, DQ347947 virus (ToSLCV) Begomoi- Pepper Huasteco NC_001359, X70418, AY044162 virus yellow vein virus (PHYVV) Begomo- Pepper golden NC_004101, U57457, AF149227, virus mosaic virus AY928512, AY928514, AY928516 (PepGMV) Tospo- Tomato spotted NC_002052, D10066, AB190813, virus wilt wirus AB198742, AY070218 (TSWV) (L segment) Tospo- Tomato spotted NC_002050, S48091, AB010996, virus wilt wirus AB190818, AF208497, AF208498, (TSWV) AY744481, AY744482, AY744483, (M segment) AY744484, AY744485, AY744486, AY744487, AY744488, AY744489, AY744490, AY744491, AY744492, AY744493, AY870389, AY870390 Tospo- Tomato spotted NC_002051, D00645, AB088385, virus wilt wirus AB 190819, AF020659, AF020660, (TSWV) AJ418777, AJ418778, AJ418779, (S segment) AJ418780, AJ418781, AY744468, AY744469, AY744470, AY744471, AY744472, AY744473, AY744474, AY744475, AY744476, AY744477, AY744478, AY744479, AY744480, AY870391, AY870392, DQ376177, DQ376178, DQ376179, DQ376180, DQ376181, DQ376182, DQ376183, DQ376184, DQ376185, DQ398945, DQ431237, DQ431238, DQ915946, DQ915947, DQ915948 Tospo- Groundnut bud NC_003614, AF025538 virus necrosis virus (GBNV) (L segment) Tospo- Groundnut bud NC_003620, U42555, AY871097 virus necrosis virus (GBNV) (M segment) Tospo- Groundnut bud NC_003619, U27809, AY871098 virus necrosis virus (GBNV) (S segment) Tospo- Capsicum NC_008302, DQ256124 virus chlorosis virus (CaCV) (L segment) Tospo- Capsicum DQ256125 virus chlorosis virus (CaCV) (M segment) Tospo- Capsicum DQ355974, DQ256123 virus chlorosis virus (CaCV) (S segment) Potex- Pepino mosaic EF599605, AY509926, AJ438767, virus virus (PepMV) DQ000984, DQ000985, AY509927, EF408821, AM491606, AJ606360, AJ606359, AJ606361, AF484251, AM109896
Example 2
Virus Segments for RNAi
[0151] Selected viral genomes were divided to .about.350-500 bp fragments, partially overlapping by about 50 bp (e.g. FIG. 2A-2D). Efficacy data was collected for the ability of single sequences to control particular virus species when expressed individually in transformed plants. In initial studies, segments corresponding to approximately 2.3 kB out of the 2.7 kB geminivirus DNA-A genome were screened (FIG. 2B; segments 1-8). Likewise, as shown in FIG. 2C, segments 1-8 representing approximately 4 kB out of the 6.5 kB PepMV (potexvirus) genome were tested. FIG. 2D shows the portions of the tripartite tospovirus genome that were tested for efficacy: approximately 1.5 kB out of the 8.9 kB L vRNA (i.e. virus RNA) (segments 1-3); approximately 1.5 kB of the 5.4 kB M vRNA (segments 4-6); and approximately 1 kB of the 2.9 kB S vRNA (segments 7-8).
[0152] In one representative study, a nucleic acid segment corresponding to a part of the TSWV coat protein (nucleocapsid) was tested for efficacy when expressed as an inverted repeat (FIG. 3). Virus resistance was found to correlate with siRNA production in tomato plants. Also in the TYLCV resistance screen, R.sub.1 plants were first examined for the presence of the transgene and then infected with the TYLCV infectious clone. As a control, a non-transformed (wild-type) tomato line was included; these wild-type plants typically have less than 5% "resistance", i.e. less than 5% of plants escape infection by inoculation in this assay. In contrast, among CP4 positive plants (containing the selectable marker gene specifying resistance to glyphosate, which is linked to the expression cassette with viral sequence), 56% of R.sub.1 plants comprising an introduced nucleic acid segment targeting the replication protein (3 constructs), 27% comprising an introduced nucleic acid segment targeting the coat protein (3 constructs), and 38% comprising an introduced nucleic acid segment targeting the rest of the coding regions (2 construct) showed resistance to TYLCV.
[0153] Additional studies scanning these and additional portions of the tospovirus genome were then undertaken, in order to identify and define portions of the viral genome which can mediate dsRNA-related virus resistance to representative viruses. These sequences were tested as inverted repeat segments in double stranded (dsRNA)-generating constructs, wherein tested constructs comprised a promoter operably linked to a given sequence in an antisense orientation (e.g. SEQ ID NOs:379-442 as described below), followed by a loop sequence, and then the given sequence in a sense orientation, and a transcriptional terminator. After transcription and base pairing of the inverted repeat sequences, the double stranded RNA regions are cleaved by a Dicer or Dicer-like RNAse to generate the specific antiviral siRNAs which target and interfere with viral gene expression.
[0154] Capsicum chlorosis virus (CaCV), Groundnut bud necrosis virus (GBNV), and Tomato spotted wilt virus (TSWV), among other tospoviruses. Virus resistance tests of transgenic R.sub.1 plants demonstrated that all regions of the tospovirus genome are equally as effective as the target of dsRNA. Thus, significant virus resistance was seen following expression of transcripts which disrupt expression of tospovirus coat protein (CP), as well as the glycoprotein-encoding virus genome segments GP1, GP2, GP3, and RNA-dependent-RNA polymerase (RdRP) protein segments. FIG. 11A-B describes resistance results seen from transgenic plants. With each bar representing an individual transgenic event and target region, for instance 50-80% of R.sub.1 plants expressing a dsRNA targeting the CP region were resistant to CaCV, while .about.25-90% of R.sub.1 plants expressing a dsRNA targeting a virus glycoprotein showed virus resistance, and about 25-80% of R.sub.1 plants expressing a dsRNA targeting a virus RdRP showed resistance (e.g. FIG. 11B). dsRNA-mediated resistance to TSWV was particularly effective, with 100% of R.sub.1 plants displaying virus resistance from all tested events with constructs targeting the coat protein (FIG. 11A). Representative TSWV, CaCV, and GBNV sequences selected for use in generation of dsRNAs targeting tospovirus gene expression are listed in SEQ ID NOs:419-436 (in antisense orientation as listed).
[0155] Additional studies demonstrated that all regions of the PepMV genome are equally effective in generating dsRNA-mediated resistance against this potexvirus. FIG. 12 shows that in most cases, 95-100% of all transgenic R.sub.1 plants comprising virus-encoded segments targeting CP (coat protein), CP/MOV (Coat Protein and Movement Protein sequences), RdRP, TGB (Triple Gene Block motif protein implicated in viral cell-to-cell and long distance movement in a host plant), or TGB/RdRP were resistant to PepMV. Representative PepMV sequences selected for use in generation of dsRNAs targeting potexvirus gene expression are listed in SEQ ID NOs:437-442, in antisense orientation as listed.
[0156] Likewise, regions of the begomovirus (geminivirus) genome were tested for their ability to generate dsRNA-mediated resistance against this virus group. TYLCV, ToSLCV, PepGMV, PHYVV, and ToLCNDV were tested in bioassays as representative begomoviruses. FIGS. 13A-B shows that up to about 70% of transgenic R.sub.1 plants displayed resistance to a given virus when CP was the dsRNA target, while up to about 100% of transgenic R.sub.1 plants displayed resistance when a nucleic acid segment encoding viral replication protein was the dsRNA target. Representative tospovirus sequences selected for use in generation of dsRNAs targeting begomovirus gene expression are listed in SEQ ID NOs:379-418, in antisense orientation as listed.
[0157] Together, numerous effective dsRNA targets for viruses in these three genera (i.e. Tospovirus, Potexvirus, Begomovirus) were identified.
[0158] Based on these RNAi results, sequences targeting efficacious target regions were selected for each virus and fused into two transgenic cassettes on a nucleic acid construct, to target multiple viruses in multiple virus families. An example of this approach is shown in FIG. 14. For begomoviruses, a Rep region was used, while for tospovirus and potexvirus, CP regions were selected since the CP region is relatively more conserved among different strains of the same viruses. Some of the sequences utilized in the cassettes were further modified, in view of expected G::U base pairings, to allow for broadening of protection to related viral strains. Representative sequences selected for use in generation of dsRNAs targeting virus gene expression in the cassettes are listed in SEQ ID NOs:443-446 for "cassette 1" of FIG. 14, and in SEQ ID NOs:447-451 for "cassette 2" of FIG. 14, each in antisense orientation as listed in the sequence listing. Thus, for instance, the nucleic acid segment targeting GBNV-CP1 which is found in cassette 1 (SEQ ID NO:443) is similar to the segment targeting GBNV-CP1 as found in SEQ ID NO:429, but may have modification as noted above, and likewise for the other nucleic acid segments used in the cassettes, relative to the target segments as initially tested for relative efficacy.
Example 3
Artificial dsRNA Fusion Constructs for Multivirus Resistance
[0159] Nucleotide segments for dsRNA expression, from two or three of the different virus groups (geminivirus, tospovirus, potexvirus), were also combined in a single expression cassette and plants were generated from transformed cells and analyzed for resistance to more than one virus. This was accomplished by fusing viral genomic fragments that tested effective for generating siRNAs conferring virus resistance, for instance as shown in Example 2. Analogously to Example 2, these sequences were tested as inverted repeat segments in double stranded (dsRNA)-generating constructs, wherein tested constructs comprised a promoter operably linked to a given sequence in an antisense orientation (e.g. SEQ ID NOs:452-454 as described below), followed by a loop sequence, and then the given sequence in a sense orientation, and a transcriptional terminator. After transcription and base pairing of the inverted repeat sequences, the double stranded RNA regions are cleaved by a Dicer or Dicer-like RNAse to generate the specific antiviral siRNAs which target and interfere with viral gene expression. In this approach, specific dsRNA segments for each of the targeted virus were used for multiple virus resistance (FIG. 4A).
[0160] Alternatively, conserved regions of the various viral genomes may provide a broader spectrum of resistance against different variants within the species or closely related species. Additionally, inverted repeats comprising sequences that are highly similar to, but less than 100% identical to, targeted segments of viral genomes may be designed and tested for efficacy. This approach, i.e. partially similar or "imperfect" inverted repeats, may broaden RNA-mediated protection to related virus strains and species. For instance, since G-U basepairing at the RNA level has a comparable degree of thermodynamic stability as some typical Watson-Crick baseparing (e.g. A-U), some imperfect repeats may nonetheless activate RISC and guide cleavage of both perfect and imperfect viral targets.
[0161] In this instance, such partially similar inverted repeats may also comprise one or more sub-sequences of at least 21 nucleotides in length that display near 100% identity to one or more viral target sequences, in order to stimulate RNAi-mediated virus resistance. Thus for instance, the overall identity of an imperfect repeat segment to one or more viral target(s) may be as low as 75%, but with 2-4 or more sub-sequences of 21-24 nucleotides that display 100% identity to a viral target sequence. Additionally, the presence of such "imperfect" dsRNA may help to increase accumulation of dsRNA in plants and prevent the triggering of transcriptional gene silencing. Thus, nucleotide segments from more than one strain of a virus, or from more than one virus, may be utilized in design and preparation of a fusion construct for dsRNA expression.
[0162] Such sequences may be identified by aligning, for instance, geminivirus or tospovirus sequences from multiple strains and/or species. Table 2 shows percent similarities between geminivirus targets at the whole genome level. Table 3 shows percent similarities between tospovirus targets at the whole genome level.
TABLE-US-00002 TABLE 2 Percent similarities between geminivirus targets (whole genome). TYLCV ToLCNDV PHYVV ToSLCV PepGMV TYLCV -- 71 64 63 60 ToLCNDV 71 -- 62 60 59 PHYVV 64 62 -- ToSLCV 63 60 67 -- 76 PepGMV 60 59 66 76 --
TABLE-US-00003 TABLE 3 Percent similarities between tospovirus targets (whole genome). Segment Virus CapCV GBNV TSWV L vRNA CapCV_L -- 44 45 8.8 kb GBNV_L 44 -- 58 TSWV_L 45 58 -- M vRNA CapCV_M -- 78 53 4.8-5.4 kB GBNV_M 78 -- 54 TSWV_M 53 54 -- S vRNA CapCV_S -- 65 42 2.9-3.3 kB GBNV_S 73 -- 48 TSWV_S 49 49 --
[0163] FIG. 4B schematically illustrates exemplary fusion constructs for dsRNA expression. Coding sequences, including sequences lacking significant identity with human and host plant genomes, were selected for generation of inverted repeats. Viral-derived segments may be oriented, for instance, "antisense-loop-sense" for dsRNA expression. This approach can reduce the size of a transgenic dsRNA cassette required for multiple virus resistance. The schematically described artificial fusion constructs of FIG. 4B are listed as SEQ ID NOs: 452-453. SEQ ID NO:452 artificial coat protein construct comprises sequences as follows: bp 1-157: TYLCV precoat protein for the loop; bp 158-507 (i.e. 350 bp segment) targeting TYLCV and ToLCNDV CP; bp 508-851 (i.e. 344 bp segment) targeting ToSLCV, PepGMV, and PHYVV CP expression. SEQ ID NO:453 comprises segments as follows: bp 1-160 targeting TSWV coat protein (CP) for the loop; bp 161-456 (i.e. 296 bp segment) targeting TSWV CP; bp 457-687 (i.e. 231 bp segment) targeting the PepMV CP; bp 688-919 (i.e. 232 bp segment) targeting the CaCV and GBNV CP. Efficacy results for SEQ ID NO:453, the artificial CP sequence against multiple tospoviruses and PepMV, are shown in FIG. 15. An additional artificial dsRNA fusion construct targeting Rep proteins of five geminiviruses (TYLCV, ToLCNDV, ToSLCV, PepGMV, and PHYVV) is given as SEQ ID NO:454. In this sequence, bp 1-150 target TYLCV Rep protein for the loop; bp 151-500 (i.e. 350 bp segment) target TYLCV and ToLCNDV Rep protein; bp 501-860 (i.e. 360 bp segment) target ToSLCV, PepGMV, and PHYVV Rep protein.
Example 4
Engineered miRNA Approach: Selection of Suitable Sequences
[0164] The bioinformatics approach described in Example 1 was used to identify several suitable sequences, approximately 21 nt in length, that target geminivirus sequences thought to be useful for miRNA-mediated suppression of geminivirus replication and/or symptom expression (Table 4; FIG. 5).
TABLE-US-00004 TABLE 4 Suitable 21 nt sequences against targeted Geminiviruses. SEQ ID NO: Name miRNA Sequence (5'.fwdarw.3') Target 1 Gemini 1 TGTCATCAATGACGTTGTACT Rep 7 Gemini 2 TGGACTTTACATGGGCCTTCA Coat Protein 13 Gemini 3 TACATGCCATATACAATAGCA Coat Protein 19 Gemini 4 TCATAGAAGTAGATCCGGATT Coat Protein 25 Gemini 5 TTCCCCTGTGCGTGAATCCGT C2/C3 31 Gemini 6 TTCCGCCTTTAATTTGGATTG Rep 37 Gemini 7 TTACATGGGCCTTCACAGCCT Coat Protein
[0165] The bioinformatics approach described in Example 1 was used to identify several suitable sequences, approximately 21 nt in length, that target Tospovirus sequences thought to be useful for miRNA-mediated suppression of Tospovirus replication and/or symptom expression (Table 5; FIG. 6).
TABLE-US-00005 TABLE 5 Suitable 21 nt sequences for each genomic segment against three targeted Tospoviruses. SEQ ID NO: Name miRNA Sequence (5'.fwdarw.3') Target 43 TospoL1-1 TTTAGGCATCATATAGATAGCT RdRP 47 Tospo L1-2 TGATTTAGGCATCATATAGAT RdRP 51 Tospo M1 TATCTATATTTTCCATCTACC GP 55 Tospo M2 TTAGTTTGCAGGCTTCAATTA NSm 59 Tospo M3 TTGCATGCTTCAATGAGAGCT NSm 63 Tospo S2 TTGACGTTAGACATGGTGTTT N 67 Tospo S3 TAGAAAGTTTTGAAGTTGAAT N
[0166] The bioinformatics approach described in Example 1 was used to identify several suitable sequences, approximately 21 nt in length, that target Potexvirus sequences thought to be useful for miRNA-mediated suppression of Tospovirus replication and/or symptom expression (Table 6; FIG. 7A-7B).
TABLE-US-00006 TABLE 6 Suitable ~21 nt sequences against targeted potexvirus. SEQ ID NO: Name miRNA Sequence (5'.fwdarw.3') Target 71 PepMV1 TCTTCATTGTAGTTAATGGAG RdPR 85 PepMV2 TTGGAAGAGGAAAAGGTGGTT RdPR 99 Pep MV3 TCAATCATGCACCTCCAGTCG RdPR 113 PepMV4 TAAGTAGCAAGGCCTAGGTGA TGBp1 127 PepMV5 TTTGGAAGTAAATGCAGGCTG TGBp2 141 PepMV6 TAACCCGTTCCAAGGGGAGAAG TGBp3
Example 5
Incorporation of miRNA-Generating Sequences in a MIR Gene Backbone
[0167] The miRNA-generating sequences of Tables 4-6 were incorporated into transgene constructs by gene synthesis. For instance, the wildtype MIR gene backbone "MON1" (see U.S. Publication 2007/0300329) from soybean with the wildtype 21nt miRNA-generating sequence (SEQ ID NO:155 of the present application) was altered to replace the wildtype miRNA-generating sequence with the Gemini1 target (SEQ ID NO:1), resulting in the Gemini1/MON1 construct (SEQ ID NO:156). Thus, a MIR backbone (e.g. the MON1 backbone of SEQ ID NO:155) can be altered by replacing the wildtype 21nt miRNA-generating sequence with any of the miRNA sequences of Tables 4-6.
[0168] Similarly, MONS from soybean (SEQ ID NO:157), MON13 from rice (SEQ ID NO:159), MON18 from maize (SEQ ID NO:161), miR159a from maize (SEQ ID NO:163), or miR167g (SEQ ID NO:165), were used as backbone sequences with miRNA-generating sequences of Tables 4-6. Thus, the following sequences were created (Table 7):
TABLE-US-00007 TABLE 7 Exemplary sequences incorporated into MIR gene backbone. miRNA-generating MIR backbone Incorporated construct sequence (SEQ ID NO) (SEQ ID NO) (SEQ ID NO) Gemini 1 MON1 Gemini1/MON1 (SEQ ID NO: 1) (SEQ ID NO: 155) (SEQ ID NO: 156) Gemini 4 MON5 Gemini 4/MON5 (SEQ ID NO: 19) (SEQ ID NO: 157) (SEQ ID NO: 158) Gemini 7 MON13 Gemini 7/MON13 (SEQ ID NO: 37) (SEQ ID NO: 159) (SEQ ID NO: 160) PepMV5 MON18 PepMV5/MON18 (SEQ ID NO: 127) (SEQ ID NO: 161) (SEQ ID NO: 162) Gemini 6 miR159a Gemini 6/miR159a (SEQ ID NO: 31) (SEQ ID NO: 163) (SEQ ID NO: 164) PepMV6 miR167g PepMV6/miR167g (SEQ ID NO: 141) (SEQ ID NO: 165) (SEQ ID NO: 166) Gemini 2 MON13 Gemini 2/MON13 (SEQ ID NO: 7) (SEQ ID NO: 159) (SEQ ID NO: 367) Gemini 3 MON1 Gemini3/MON1 (SEQ ID NO: 13) (SEQ ID NO: 155) (SEQ ID NO: 368) Gemini 5 miR159a Gemini 5/MiR159a (SEQ ID NO: 25) (SEQ ID NO: 163) (SEQ ID NO: 369) PepMV1 miR159a PepMV1/miR159a (SEQ ID NO: 71) (SEQ ID NO: 163) (SEQ ID NO: 363) PepMV2 MON5 PepMV2/MON5 (SEQ ID NO: 85) (SEQ ID NO: 157) (SEQ ID NO: 364) PepMV3 MON13 PepMV3/MON13 (SEQ ID NO: 99) (SEQ ID NO: 159) (SEQ ID NO: 365) PepMV4 MON1 PepMV4/MON1 (SEQ ID NO: 113) (SEQ ID NO: 155) (SEQ ID NO: 366) TospoL1-1 miR167g TospoL1-1/miR167g (SEQ ID NO: 43) (SEQ ID NO: 165) SEQ ID NO: 370) TospoL1-2 MON1 TospoL1-2/MON1 (SEQ ID NO: 47) (SEQ ID NO: 155) (SEQ ID NO: 371) TospoM2 MON1 TospoM2/MON1 (SEQ ID NO: 55) (SEQ ID NO: 155) (SEQ ID NO: 372) TospoM3 MON13 TospoM3/MON13 (SEQ ID NO: 59) (SEQ ID NO: 159) (SEQ ID NO: 373) TospoS2 MON5 TospoS2/MON5 (SEQ ID NO: 63) (SEQ ID NO: 157) (SEQ ID NO: 374) TospoS3 MON13 TospoS3/MON13 (SEQ ID NO: 67) (SEQ ID NO: 159) (SEQ ID NO: 375)
Other selected miRNA-generating sequences, for instance from among any of SEQ ID NOs:1-154, or from within SEQ ID NOs:169-362 or portions thereof, may be utilized with MIR backbone sequences, for instance MON1 from soybean, MONS from soybean, MON13 from rice, MON18 from maize, miR159a from maize, or mi167g from maize (SEQ ID NOs:155, 157, 159, 161, 163, or 165) to create additional efficacious miRNA-generating constructs. Additionally, the MIR backbone sequences may be modified for instance by replacing the portion of the sequence of the plant-derived backbone sequence which specifies an miRNA with a selected virus-derived or artificial (e.g. consensus) sequence, and/or shortened, e.g. a deletion made at the 3' and/or 5' end, for instance as reflected in the miRNA-generating sequences and MIR backbone portions of SEQ ID NOs:158, 160, 162, 164, 166, 363, 364, 365, 367, 369, 370, 373, 374, and 375, described below.
[0169] 21-mers identified from within SEQ ID NOs:169-362, for instance among or within SEQ ID NOs:1-154, are tested individually, or two or more at a time while present in one expression cassette, for efficacy in transgenic tomato plants against appropriate viruses such as TYLCV and TSWV. Multiple effective 21-mers are expressed in one transgenic cassette using a polycistronic or a phased siRNA backbone (e.g. FIG. 8). Results of virus resistance assays from such testing of 21-mers are given in Example 7.
[0170] Once the efficacious antiviral miRNAs are identified, multiple miRNAs are deployed by expressing as a single transgenic trait, to achieve multiple virus resistance in transgenic plants. This is accomplished by fusing engineered MIR gene(s) described above together (e.g. to create SEQ ID NOs:156, 158, 160, 162, 164, 166, 363-375), or using a ta-siRNA or a phased siRNA gene structure that can deliver multiple antiviral .about.21nt sequences (e.g. FIG. 8). The later approaches are accomplished via a new round of gene synthesis to replace wild-type siRNAs with the efficacious antiviral 21nt sequences in the ta-siRNA or phased siRNA backbone.
Example 6
Supplementing RNA-Based Resistance
[0171] The RNA-mediated resistance approach utilizing dsRNA or miRNA can be supplemented by placing antiviral sequences, e.g. ones that encode a protein, into the loop of a dsRNA-encoding sequence, or by inserting a sequence that encodes an efficacious dsRNA or miRNA into an intron of a polypeptide expression cassette (intronic dsRNA/miRNA; Frizzi et al., 2008) (e.g. FIG. 9). For example, viral protein(s) such as coat protein and/or replicase may be expressed in a transgenic plant that also expresses an efficacious dsRNA, miRNA, or siRNA, without use of an additional transgene cassette, by inserting protein coding sequence into the loop of a dsRNA. A sequence that encodes a peptide aptamer that interferes with geminivirus replication may also be employed (e.g. Lopez-Ochoa et al., 2006).
[0172] Thus, artificial sequences may be expressed along with the dsRNA, miRNA, or siRNA in order to augment the virus resistant phenotype. For instance, an artificial nucleotide loop engineered from the tospovirus genome may be used. The tospovirus (ssRNA) genome comprises a "panhandle" structure due to the presence of conserved terminal repeats of 8-11 nt at both ends of each genome component. These terminal repeats from each component of the genome (SEQ ID NOs:167-168), for instance from GBNV (Groundnut bud necrosis virus) or CaCV (Capsicum chlorosis virus), may be fused and used as part of a loop-forming sequence in a plant transformation construct. Such an artificial sequence may comprise, for instance, SEQ ID NOs:376-378, or SEQ ID NO:455, and can serve as an artificial substrate competing for reverse transcriptase, may interfere with proper circularization of replicating viral genome components, may themselves generate nucleotide segments efficacious via RNAi, or one or more of the above, thus interfering with virion assembly.
[0173] FIG. 16 demonstrates that inclusion of tospovirus terminal sequences in a construct for generating dsRNA results in measurable resistance to a virus such as CaCV. In the tested construct, SEQ ID NOs:376-378, themselves comprising SEQ ID NOs:168, were fused to create SEQ ID NO:455 (i.e. bp 1-247 of SEQ ID NO:455 comprise SEQ ID NO:376; bp 248-303 comprise SEQ ID NO:377, and bp 304-369 comprise SEQ ID NO:378). The construct was tested in both sense and antisense orientations, and the level of virus resistance was compared to that demonstrated by a control tomato plant, non-transgenic but otherwise isogenic.
[0174] Sequences encoding, for instance, coat protein or replicase, as well as artificial sequences described above, may be embedded within an intron sequence as well. Thus, multiple modes of action may be deployed by being engineered into a single expression cassette (e.g. FIG. 9), or more than one expression cassette.
Example 7
Results of Exemplary Virus Resistance Assays Using Engineered miRNAs
[0175] Engineered miRNA-producing constructs as listed in Table 7 were tested for efficacy. Bioassay results for geminivirus and potexvirus assays are shown in Tables 8-9. Virus resistance was observed due to miRNA expression, and correlating to expression and proper processing of a given transgene transcript. For instance, for SEQ ID NO:166, proper processing of PepMV6/mir167g was not observed, nor was virus resistance seen for this construct, thus correlating miRNA production with virus resistance in transgenic R.sub.1 plants. Regarding tospovirus experiments, expression and proper processing of transcripts from SEQ ID NOs:370, 371, 373, and 375 was observed, targeting, respectively, the RdRP, RdRP, NsM, and N genes. CP4-positive R.sub.1 plants displayed reduced symptoms when infected with TSWV. For SEQ ID NOs:372 and 374, targeting, respectively, the NsM and N genes, proper processing of miRNA was not observed, and no reduction in symptoms was noted.
[0176] The synthetic miRNAs were most effective against PepMV, and gave delayed or reduced virus infection symptoms against geminiviruses and tospovirus. The constructs can be combined in stacked miRNA cassettes in order to synergistically inhibit the target viruses. Since multiple miRNAs can be expressed from a single expression cassette, constructs expressing, for instance, 8 or 10 antiviral miRNAs can be created (as shown in FIG. 10A-10B). As an additional approach, antiviral miRNA expression cassettes can be inserted as the "loop" sequence (e.g. FIG. 9) of a dsRNA expression cassette, to combine the dsRNA and miRNA mechanisms for viral control (see FIG. 10C).
TABLE-US-00008 TABLE 8 Results of geminivirus resistance assays utilizing engineered sequences incorporated into MIR gene backbones. Incorporated construct % Virus Resistance.sup.b Target (SEQ ID NO) miRNA Expression.sup.a TYLCV ToLCNDV PepGMV PHYVV Region SEQ ID NO: 156 Gemini1/MON1 yes .sup. 16 .+-. 16% 31 .+-. 17 37 .+-. 13 29 .+-. 16 Rep SEQ ID NO: 367 Gemini 2/MON13 yes 34 .+-. 27 27 .+-. 14 29 .+-. 4 32 .+-. 16 CP SEQ ID NO: 368 Gemini3/MON1 yes 13 .+-. 8 23 .+-. 7 34 .+-. 11 14 .+-. 19 CP SEQ ID NO: 158 Gemini4/MON5 no -- -- -- -- CP SEQ ID NO: 369 Gemini5/miR159a yes 14 .+-. 19 30 .+-. 20 51 .+-. 16 28 .+-. 37 C2/C3 SEQ ID NO: 164 Gemini6/miR159a yes 20 .+-. 13 23 .+-. 12 36 .+-. 10 23 .+-. 19 Rep SEQ ID NO: 160 Gemini 7-1/MON13 yes 7 28 40 11 CP .sup.aproper processing of the miRNA detected in transgenic plants .sup.baverage percentage .+-. s.d. of R1 CP4-positive segregants for resistance to tomato yellow leaf curl virus (TYLCV), Tomato leaf curl New Delhi virus (ToLCNDV), Pepper golden mosaic virus (PePGMV), or Pepper Huasteco yellow vein virus (PHYVV.
TABLE-US-00009 TABLE 9 Results of Pepino mosaic virus (PepMV) resistance assays utilizing engineered sequences incorporated into MIR gene backbones. Incorporated construct (SEQ ID NO) miRNA Expression.sup.a % Virus Resistance.sup.b Target Region 363 PepMV1/miR159a yes Not tested RdRP 364 PepMV2/MON5 yes (weak) 10 .+-. 14 RdRP 365 PepMV3/MON13 yes 100 .+-. 0 RdRP 366 PepMV4/MON1 yes (weak) 44 .+-. 52 TGBp1 162 PepMV5/MON18 yes 99 .+-. 3 TGBp2 166 PepMV6/miR167g no -- TGBp3 .sup.aproper processing of the miRNA detected in transgenic plants .sup.baverage percentage .+-. s.d. of R1 CP4-positive segregants for resistance to PepMV.
[0177] All of the compositions and methods disclosed and claimed herein can be made and executed without undue experimentation in light of the present disclosure. While the compositions and methods of this invention have been described in terms of the foregoing illustrative embodiments, it will be apparent to those of skill in the art that variations, changes, modifications, and alterations may be applied to the composition, methods, and in the steps or in the sequence of steps of the methods described herein, without departing from the true concept, spirit, and scope of the invention. More specifically, it will be apparent that certain agents that are both chemically and physiologically related may be substituted for the agents described herein while the same or similar results would be achieved. All such similar substitutes and modifications apparent to those skilled in the art are deemed to be within the spirit, scope, and concept of the invention as defined by the appended claims.
REFERENCES
[0178] The references listed below are incorporated herein by reference to the extent that they supplement, explain, provide a background, or teach methodology, techniques, and/or compositions employed herein. [0179] U.S. Pat. Nos. 4,536,475; 4,693,977; 4,886,937; 4,940,838; 4,959,317; 5,015,580; 5,107,065; 5,110,732; 5,231,020; 5,262,316; 5,283,184; 5,302,523; 5,384,253; 5,464,763; 5,464,765; 5,501,967; 5,508,184; 5,527,695; 5,459,252; 5,538,880; 5,550,318; 5,563,055; 5,591,616; 5,693,512; 5,633,435; 5,759,829; 5,981,840; 6,118,047; 6,160,208; 6,229,067; 6,384,301; 6,399,861; 6,403,865; 6,852,907. [0180] U.S. Publication 2002/0048814; U.S. Publication 2003/017596; U.S. Publication 2003/018993; U.S. Publication 2004/0029283; U.S. Publications 2005/0120415; U.S. Publication 2005/144669; U.S. Publication 2006/0200878; U.S. Publication 2006/0174380; U.S. Publication 2007/0259785; U.S. Publication 2007/0300329; U.S. Publication 2008/0066206. [0181] Allen et al., Cell 121:207-221, 2005. [0182] Altschul et al., J. Mol. Biol. 215:403-410, 1990. [0183] Ambros et al., RNA 9:277-279, 2003. [0184] Bartel, Cell 116:281-297, 2004. [0185] Bevan, Nature 304:184-187, 1983. [0186] Boulton, "Agrobacterium-Mediated Transfer of geminiviruses to Plant Tissues," in "Methods in Molec. Biol., vol. 49: Plant Gene Transfer and Expression Protocols, ed. H. Jones, Humana Press, Totowa, N.J., 1996. [0187] Brodersen and Voinnet, Trends Genetics, 22:268-280, 2006. [0188] Broothaerts et al., Nature, 433:629-633, 2005. [0189] Brutlag et al., Comp. Chem. 17: 203-207, 1993. [0190] Christopher and Rajam, Pl. Cell Tiss. Org. Cult. 46:245-250, 1996. [0191] Chuang et al., PNAS, 97:4985-4990, 2000. [0192] Cotillon et al., Arch. Virol. 147:2225-2230, 2002. [0193] Dellaporta et al., Stadler Symposium 11:263-282, 1988. [0194] Elbashir et al., Genes & Devel., 15:188-200, 2002. [0195] Frizzi et al., Plant Biotechnol. J., 6:13-21, 2008. [0196] Griffiths-Jones, Nucleic Acids Res., 31:439-441, 2003. [0197] Grimsley, Physiol. Plantarum, 79:147-153, 1990. [0198] Gruber et al., In: Vectors for Plant Transformation, Glick and Thompson (Eds.), CRC Press, Inc., Boca Raton, 89-119, 1993. [0199] Hamilton and Baulcombe, Science, 286:950-952, 1999. [0200] Hannon, Nature 418:244-251, 2002. [0201] Haymes et al., Nucleic Acid Hybridization, A Practical Approach, IRL Press, Washington, D.C. (1985). [0202] Herrera-Estrella Nature 303:209-213, 1983. [0203] Hirel et al., Plant Molecular Biology, 20:207-218, 1992. [0204] Horsch et al., Science, 227:1229, 1985. [0205] Ikatu et al., Bio/Technol. 8:241-242, 1990. [0206] Jefferson et al., EMBO J. 6:3901-3907, 1987. [0207] Jones-Rhoades et al. Annu. Rev. Plant Biol., 57:19-53, 2006. [0208] Kaeppler et al., Plant Cell Reports, 9:415-418, 1990. [0209] Katz et al., J. Gen. Microbiol. 129:2703-2714, 1983. [0210] Kim, Nature Rev. Mol. Cell Biol., 6:376-385, 2005. [0211] Klee, Bio/Technol. 3:637-642, 1985. [0212] Kumar et al., Plant Dis. 77:938-941, 1993. [0213] Liu et al., Plant Cell Rep. 9:360-364, 1990. [0214] Lopez et al., Arch. Virol. 150:619-627, 2005. [0215] Lopez-Ochoa et al., J. Virol., 80:5841, 2006. [0216] McCormick, Plant Tissue Culture Manual B6:1-9, 1991. [0217] Miki et al., In: Methods in Plant Molecular Biology and Biotechnology, Glick and Thompson (Eds.), CRC Press, 67-88, 1993. [0218] Moloney et al., Plant Cell Reports, 8:238, 1989. [0219] Niu et al., Nature Biotechnology 24:1420-1428, 2006. [0220] Odell et al., Nature 313:810-812, 1985. [0221] Omirulleh et al., Plant Mol. Biol., 21:415-28, 1993. [0222] Ow et al., Science 234:856-859, 1986. [0223] Padidam et al., J. Gen. Virol. 73:1609-1616, 1999. [0224] Potrykus et al., Mol. Gen. Genet., 199:183-188, 1985. [0225] Redenbaugh et al. in "Safety Assessment of Genetically Engineered Fruits and Vegetables", CRC Press, 1992. [0226] Reynolds et al., Nat Biotechnol. 22:326-30, 2004. [0227] Sambrook et al., (ed.), Molecular Cloning, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989. [0228] Sutcliffe et al., Proc. Natl. Acad. Sci. (U.S.A.) 75:3737-3741, 1978. [0229] Tang, Trends Biochem. Sci., 30:106-14, 2005. [0230] Tomari and Zamore, Genes & Dev., 19:517-529, 2005. [0231] Tomari et al. Curr. Biol., 15:R61-64, 2005. [0232] Van Heeke and Schuster, J. Biol. Chem. 264:5503-5509, 1989. [0233] Wesley et al., Plant J., 27:581-590, 2001. [0234] WO 98/05770; WO 99/053050; WO 99/049029; WO94/01550; WO 05/019408
Sequence CWU 0 SQTB SEQUENCE LISTING The patent application
contains a lengthy "Sequence Listing" section. A copy of the
"Sequence Listing" is available in electronic form from the USPTO
web site
(http://seqdata.uspto.gov/?pageRequest=docDetail&DocID=US20190249189A1).
An electronic copy of the "Sequence Listing" will also be available
from the USPTO upon request and payment of the fee set forth in 37
CFR 1.19(b)(3).
0 SQTB SEQUENCE LISTING The patent application contains a lengthy
"Sequence Listing" section. A copy of the "Sequence Listing" is
available in electronic form from the USPTO web site
(http://seqdata.uspto.gov/?pageRequest=docDetail&DocID=US20190249189A1).
An electronic copy of the "Sequence Listing" will also be available
from the USPTO upon request and payment of the fee set forth in 37
CFR 1.19(b)(3).
References
D00001
D00002
D00003
D00004
D00005
D00006
D00007
D00008
D00009
D00010
D00011
D00012
D00013

D00014
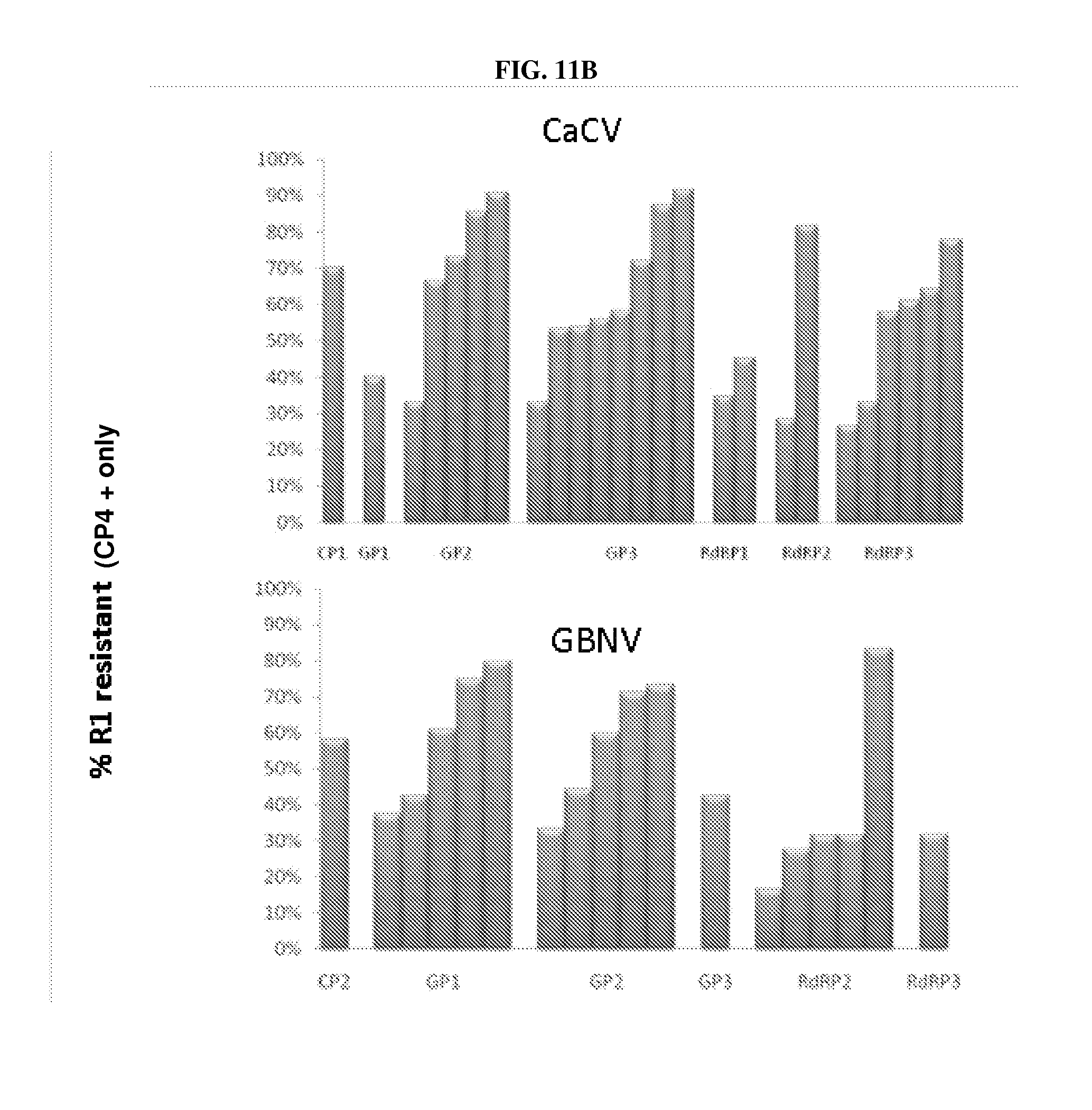
D00015
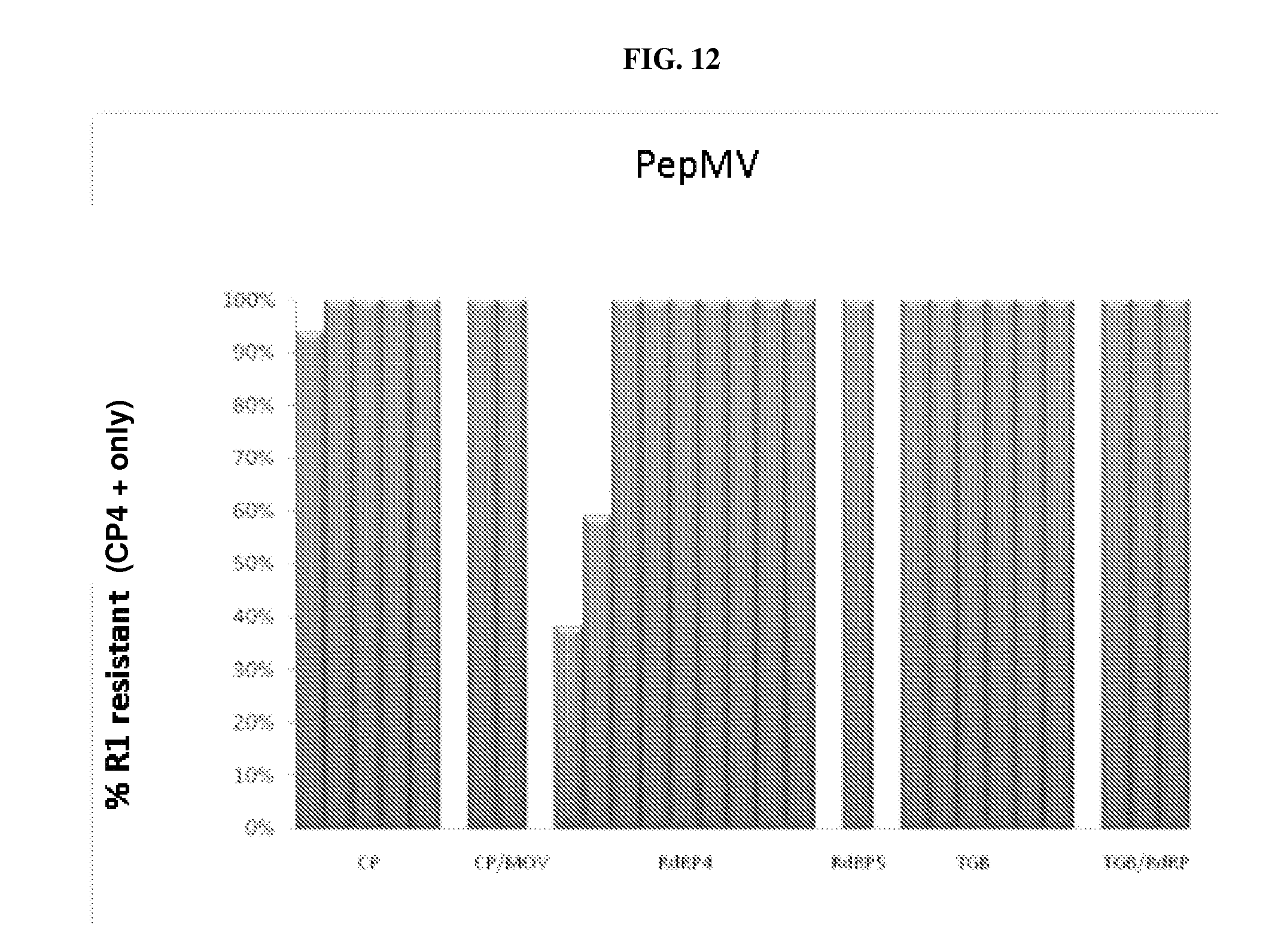
D00016
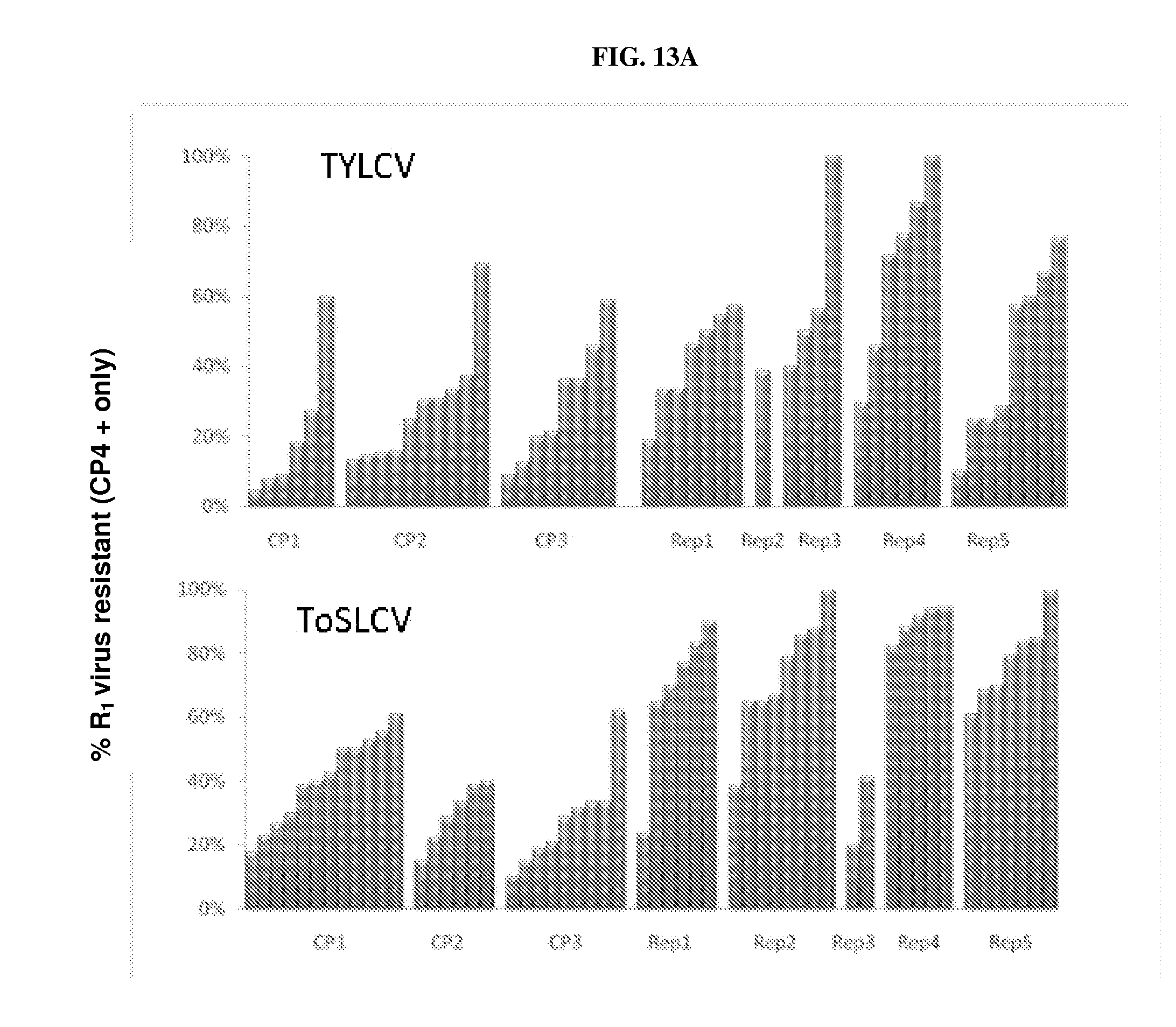
D00017
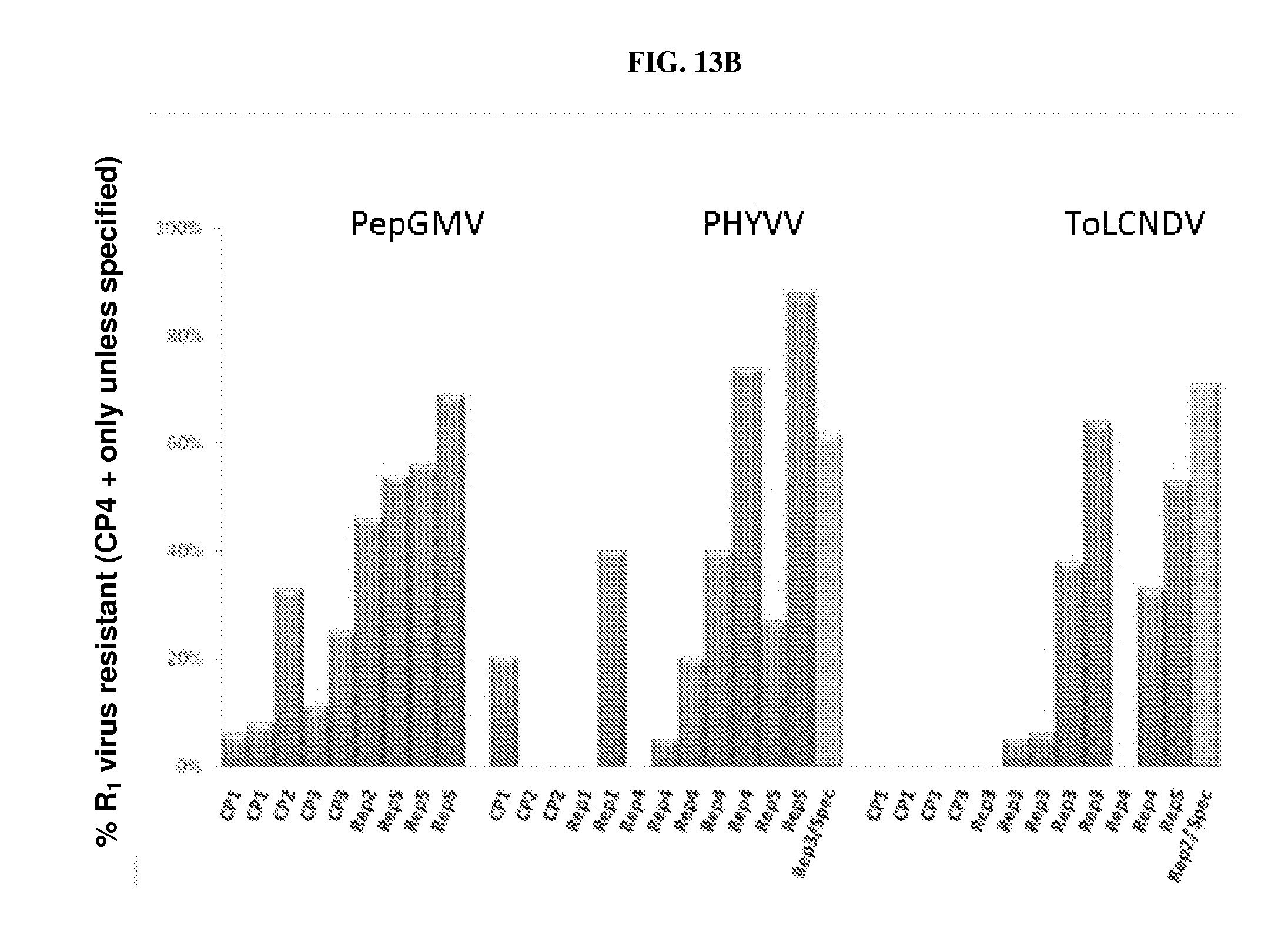
D00018
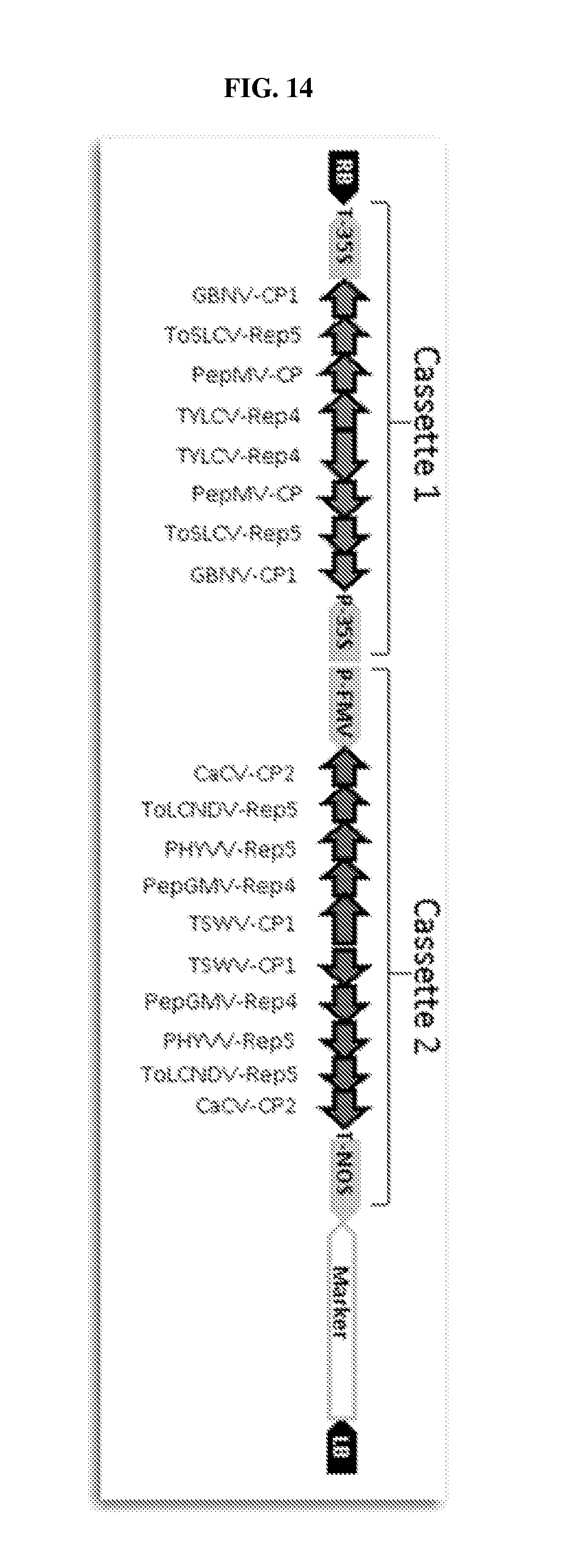
D00019

D00020

XML
uspto.report is an independent third-party trademark research tool that is not affiliated, endorsed, or sponsored by the United States Patent and Trademark Office (USPTO) or any other governmental organization. The information provided by uspto.report is based on publicly available data at the time of writing and is intended for informational purposes only.
While we strive to provide accurate and up-to-date information, we do not guarantee the accuracy, completeness, reliability, or suitability of the information displayed on this site. The use of this site is at your own risk. Any reliance you place on such information is therefore strictly at your own risk.
All official trademark data, including owner information, should be verified by visiting the official USPTO website at www.uspto.gov. This site is not intended to replace professional legal advice and should not be used as a substitute for consulting with a legal professional who is knowledgeable about trademark law.