Microorganisms For The Production Of 1,4-butanediol, 4-hydroxybutanal, 4-hydroxybutyryl-coa, Putrescine And Related Compounds, And Methods Related Thereto
HASELBECK; Robert ; et al.
U.S. patent application number 15/173427 was filed with the patent office on 2016-12-29 for microorganisms for the production of 1,4-butanediol, 4-hydroxybutanal, 4-hydroxybutyryl-coa, putrescine and related compounds, and methods related thereto. The applicant listed for this patent is Genomatica, Inc.. Invention is credited to Anthony P. BURGARD, Robert HASELBECK, Wei NIU, John D. TRAWICK.
| Application Number | 20160376614 15/173427 |
| Document ID | / |
| Family ID | 43876524 |
| Filed Date | 2016-12-29 |






View All Diagrams
| United States Patent Application | 20160376614 |
| Kind Code | A1 |
| HASELBECK; Robert ; et al. | December 29, 2016 |
MICROORGANISMS FOR THE PRODUCTION OF 1,4-BUTANEDIOL, 4-HYDROXYBUTANAL, 4-HYDROXYBUTYRYL-COA, PUTRESCINE AND RELATED COMPOUNDS, AND METHODS RELATED THERETO
Abstract
The invention provides non-naturally occurring microbial organisms comprising a 1,4-butanediol (BDO), 4-hydroxybutyryl-CoA, 4-hydroxybutanal or putrescine pathway comprising at least one exogenous nucleic acid encoding a BDO, 4-hydroxybutyryl-CoA, 4-hydroxybutanal or putrescine pathway enzyme expressed in a sufficient amount to produce BDO, 4-hydroxybutyryl-CoA, 4-hydroxybutanal or putrescine and further optimized for expression of BDO. The invention additionally provides methods of using such microbial organisms to produce BDO, 4-hydroxybutyryl-CoA, 4-hydroxybutanal or putrescine.
| Inventors: | HASELBECK; Robert; (San Diego, CA) ; TRAWICK; John D.; (La Mesa, CA) ; NIU; Wei; (Lincoln, NE) ; BURGARD; Anthony P.; (Bellefonte, PA) | ||||||||||
| Applicant: |
|
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Family ID: | 43876524 | ||||||||||
| Appl. No.: | 15/173427 | ||||||||||
| Filed: | June 3, 2016 |
Related U.S. Patent Documents
| Application Number | Filing Date | Patent Number | ||
|---|---|---|---|---|
| 13737866 | Jan 9, 2013 | |||
| 15173427 | ||||
| 13109732 | May 17, 2011 | 8377667 | ||
| 13737866 | ||||
| 12904130 | Oct 13, 2010 | 8377666 | ||
| 13109732 | ||||
| 61251287 | Oct 13, 2009 | |||
| Current U.S. Class: | 435/147 |
| Current CPC Class: | C12P 13/001 20130101; C12P 7/42 20130101; C12P 7/24 20130101; C12P 19/40 20130101; C12P 7/18 20130101 |
| International Class: | C12P 7/18 20060101 C12P007/18; C12P 7/24 20060101 C12P007/24; C12P 7/42 20060101 C12P007/42; C12P 13/00 20060101 C12P013/00 |
Claims
1. A non-naturally occurring microbial organism, comprising a 4-hydroxybutanal pathway comprising at least one exogenous nucleic acid encoding a 4-hydroxybutanal pathway enzyme expressed in a sufficient amount to produce 4-hydroxybutanal, said 4-hydroxybutanal pathway comprising: succinyl-CoA reductase (aldehyde forming); 4-hydroxybutyrate dehydrogenase; and 4-hydroxybutyrate reductase; alpha-ketoglutarate decarboxylase; 4-hydroxybutyrate dehydrogenase; and 4-hydroxybutyrate reductase; succinate reductase; 4-hydroxybutyrate dehydrogenase, and 4-hydroxybutyrate reductase; alpha-ketoglutarate decarboxylase, or glutamate dehydrogenase or glutamate transaminase and glutamate decarboxylase and 4-aminobutyrate dehydrogenase or 4-aminobutyrate transaminase; 4-hydroxybutyrate dehydrogenase; and 4-hydroxybutyrate reductase; or alpha-ketoglutarate reductase; 5-hydroxy-2-oxopentanoate dehydrogenase; and 5-hydroxy-2-oxopentanoate decarboxylase; succinyl-CoA reductase (aldehyde forming); 4-hydroxybutyrate dehydrogenase; 4-hydroxybutyryl-CoA transferase or 4-hydroxybutyryl-CoA hydrolase or 4-hydroxybutyryl-CoA synthetase; and 4-hydroxybutyryl-CoA reductase (aldehyde forming); succinyl-CoA reductase (aldehyde forming); 4-hydroxybutyrate dehydrogenase; 4-hydroxybutyrate kinase; phosphotrans-4-hydroxybutyrylase; and 4-hydroxybutyryl-CoA reductase (aldehyde forming); or succinyl-CoA reductase (aldehyde forming); 4-hydroxybutyrate dehydrogenase; 4-hydroxybutyrate kinase; and 4-hydroxybutyryl-phosphate reductase.
2. The non-naturally occurring microbial organism of claim 1, wherein said microbial organism comprises three exogenous nucleic acids encoding succinyl-CoA reductase (aldehyde forming); 4-hydroxybutyrate dehydrogenase; and 4-hydroxybutyrate reductase.
3. The non-naturally occurring microbial organism of claim 1, wherein said at least one exogenous nucleic acid is a heterologous nucleic acid.
4. A method for producing 4-hydroxybutanal, comprising culturing the non-naturally occurring microbial organism of claim 1 under conditions and for a sufficient period of time to produce 4-hydroxybutanal.
5. The method of claim 4, wherein said non-naturally occurring microbial organism is in a substantially anaerobic culture medium.
6. The non-naturally occurring microbial organism of claim 1, said 4-hydroxybutanal pathway comprising alpha-ketoglutarate decarboxylase; 4-hydroxybutyrate dehydrogenase; and 4-hydroxybutyrate reductase.
7. The non-naturally occurring microbial organism of claim 6, wherein said microbial organism comprises three exogenous nucleic acids encoding alpha-ketoglutarate decarboxylase; 4-hydroxybutyrate dehydrogenase; and 4-hydroxybutyrate reductase.
8. The non-naturally occurring microbial organism of claim 6, wherein said at least one exogenous nucleic acid is a heterologous nucleic acid.
9-10. (canceled)
11. The non-naturally occurring microbial organism of claim 1, said 4-hydroxybutanal pathway comprising succinate reductase; 4-hydroxybutyrate dehydrogenase, and 4-hydroxybutyrate reductase.
12. The non-naturally occurring microbial organism of claim 11, wherein said microbial organism comprises three exogenous nucleic acids encoding succinate reductase; 4-hydroxybutyrate dehydrogenase, and 4-hydroxybutyrate reductase.
13. The non-naturally occurring microbial organism of claim 11, wherein said at least one exogenous nucleic acid is a heterologous nucleic acid.
14-15. (canceled)
16. The non-naturally occurring microbial organism of claim 1, said 4-hydroxybutanal pathway comprising alpha-ketoglutarate decarboxylase, or glutamate dehydrogenase or glutamate transaminase and glutamate decarboxylase and 4-aminobutyrate dehydrogenase or 4-aminobutyrate transaminase; 4-hydroxybutyrate dehydrogenase; and 4-hydroxybutyrate reductase.
17. The non-naturally occurring microbial organism of claim 1, said 4-hydroxybutanal pathway comprising alpha-ketoglutarate reductase; 5-hydroxy-2-oxopentanoate dehydrogenase; and 5-hydroxy-2-oxopentanoate decarboxylase.
18. A non-naturally occurring microbial organism, comprising a microbial organism having a putrescine pathway comprising at least one exogenous nucleic acid encoding a putrescine pathway enzyme expressed in a sufficient amount to produce putrescine, said putrescine pathway comprising: succinate reductase; 4-aminobutyrate dehydrogenase or 4-aminobutyrate transaminase; 4-aminobutyrate reductase; and putrescine dehydrogenase or putrescine transaminase; alpha-ketoglutarate decarboxylase; 4-aminobutyrate dehydrogenase or 4-aminobutyrate transaminase; 4-aminobutyrate reductase; and putrescine dehydrogenase or putrescine transaminase; glutamate dehydrogenase or glutamate transaminase; glutamate decarboxylase; 4-aminobutyrate reductase; and putrescine dehydrogenase or putrescine transaminase; or alpha-ketoglutarate reductase; 5-amino-2-oxopentanoate dehydrogenase or 5-amino-2-oxopentanoate transaminase; 5-amino-2-oxopentanoate decarboxylase; and putrescine dehydrogenase or putrescine transaminase.
19. The non-naturally occurring microbial organism of claim 18, said putrescine pathway comprising alpha-ketoglutarate decarboxylase; 4-aminobutyrate dehydrogenase or 4-aminobutyrate transaminase; 4-aminobutyrate reductase; and putrescine dehydrogenase or putrescine transaminase.
20. The non-naturally occurring microbial organism of claim 18, said putrescine pathway comprising glutamate dehydrogenase or glutamate transaminase; glutamate decarboxylase; 4-aminobutyrate reductase; and putrescine dehydrogenase or putrescine transaminase.
21. The non-naturally occurring microbial organism of claim 18, said putrescine pathway comprising alpha-ketoglutarate reductase; 5-amino-2-oxopentanoate dehydrogenase or 5-amino-2-oxopentanoate transaminase; 5-amino-2-oxopentanoate decarboxylase; and putrescine dehydrogenase or putrescine transaminase.
Description
[0001] This application is a continuation of application Ser. No. 13/737,866, filed Jan. 9, 2013, which is a continuation of application Ser. No. 13/109,732, filed May 17, 2011, now U.S. Pat. No. 8,377,667, which is a continuation of application Ser. No. 12/904,130, filed Oct. 13, 2010, now U.S. Pat. No. 8,377,666, which claims the benefit of priority of U.S. Provisional Application No. 61/251,287, filed Oct. 13, 2009, each of which the entire contents are incorporated herein by reference.
[0002] Incorporated herein by reference is the Sequence Listing being submitted via EFS-Web as an ASCII text file named 12956-399-999_Sequence_Listing.txt, created Jun. 2, 2016, and being 168,480 bytes in size.
BACKGROUND OF THE INVENTION
[0003] This invention relates generally to in silico design of organisms and engineering of organisms, more particularly to organisms having 1,4-butanediol, 4-hydroxybutyryl-CoA, 4-hydroxybutanal or putrescine biosynthesis capability.
[0004] The compound 4-hydroxybutanoic acid (4-hydroxybutanoate, 4-hydroxybutyrate, 4-HB) is a 4-carbon carboxylic acid that has industrial potential as a building block for various commodity and specialty chemicals. In particular, 4-HB has the potential to serve as a new entry point into the 1,4-butanediol family of chemicals, which includes solvents, resins, polymer precursors, and specialty chemicals. 1,4-Butanediol (BDO) is a polymer intermediate and industrial solvent with a global market of about 3 billion lb/year. BDO is currently produced from petrochemical precursors, primarily acetylene, maleic anhydride, and propylene oxide.
[0005] For example, acetylene is reacted with 2 molecules of formaldehyde in the Reppe synthesis reaction (Kroschwitz and Grant, Encyclopedia of Chem. Tech., John Wiley and Sons, Inc., New York (1999)), followed by catalytic hydrogenation to form 1,4-butanediol. It has been estimated that 90% of the acetylene produced in the U.S. is consumed for butanediol production. Alternatively, it can be formed by esterification and catalytic hydrogenation of maleic anhydride, which is derived from butane. Downstream, butanediol can be further transformed; for example, by oxidation to .gamma.-butyrolactone, which can be further converted to pyrrolidone and N-methyl-pyrrolidone, or hydrogenolysis to tetrahydrofuran. These compounds have varied uses as polymer intermediates, solvents, and additives, and have a combined market of nearly 2 billion lb/year.
[0006] It is desirable to develop a method for production of these chemicals by alternative means that not only substitute renewable for petroleum-based feedstocks, and also use less energy- and capital-intensive processes. The Department of Energy has proposed 1,4-diacids, and particularly succinic acid, as key biologically-produced intermediates for the manufacture of the butanediol family of products (DOE Report, "Top Value-Added Chemicals from Biomass", 2004). However, succinic acid is costly to isolate and purify and requires high temperatures and pressures for catalytic reduction to butanediol.
[0007] Thus, there exists a need for alternative means for effectively producing commercial quantities of 1,4-butanediol and its chemical precursors. The present invention satisfies this need and provides related advantages as well.
SUMMARY OF INVENTION
[0008] The invention provides non-naturally occurring microbial organisms containing a 1,4-butanediol (BDO), 4-hydroxybutanal (4-HBal), 4-hydroxybutyryl-CoA (4-HBCoA) and/or putrescine pathway comprising at least one exogenous nucleic acid encoding a BDO, 4-HBal and/or putrescine pathway enzyme expressed in a sufficient amount to produce BDO, 4-HBal, 4-HBCoA and/or putrescine. The microbial organisms can be further optimized for expression of BDO, 4-HBal, 4-HBCoA and/or putrescine. The invention additionally provides methods of using such microbial organisms to produce BDO, 4-HBal, 4-HBCoA and/or putrescine.
BRIEF DESCRIPTION OF THE DRAWINGS
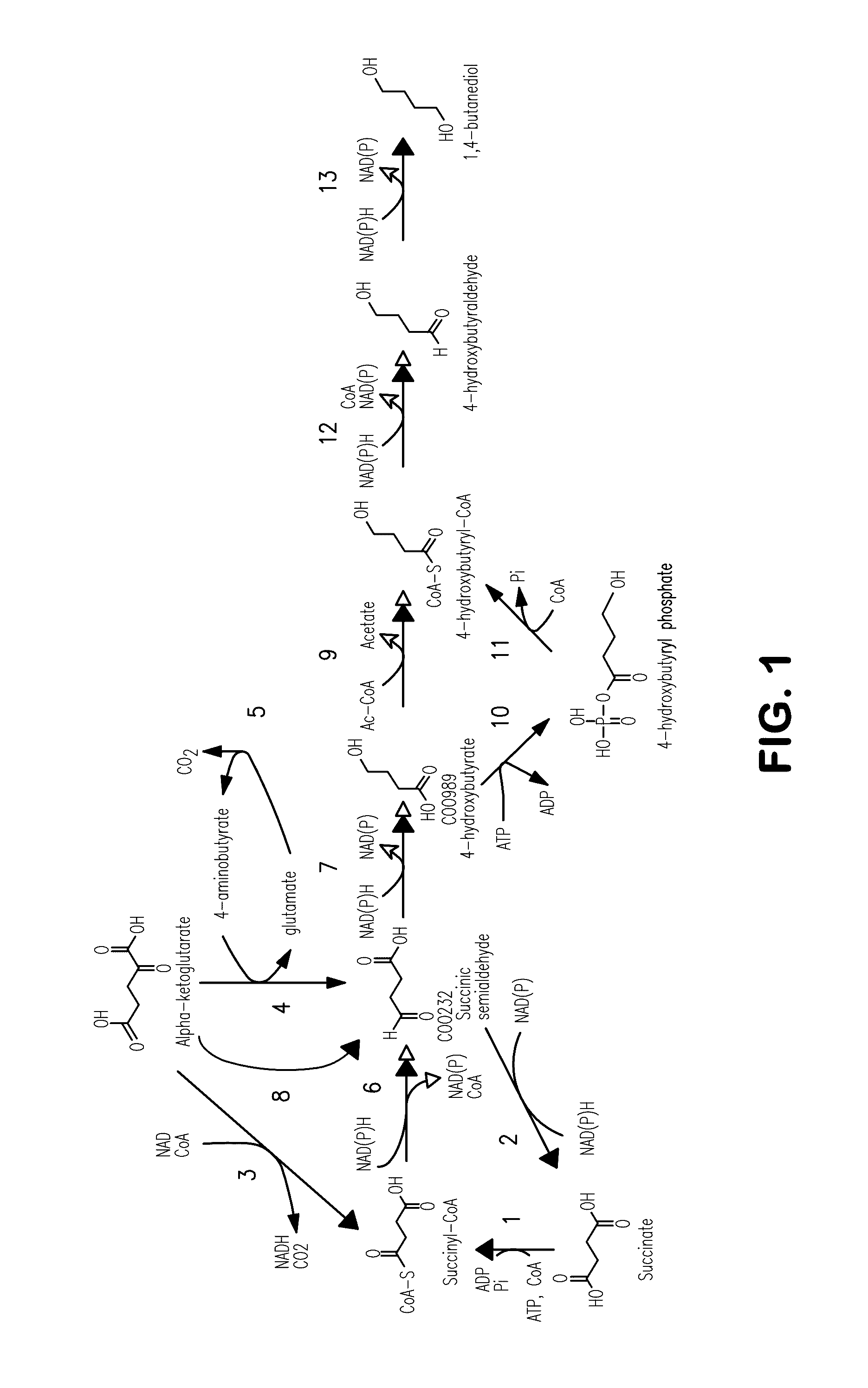
[0009] FIG. 1 is a schematic diagram showing biochemical pathways to 4-hydroxybutyurate (4-HB) and to 1,4-butanediol production. The first 5 steps are endogenous to E. coli, while the remainder can be expressed heterologously. Enzymes catalyzing the biosynthetic reactions are: (1) succinyl-CoA synthetase; (2) CoA-independent succinic semialdehyde dehydrogenase; (3) .alpha.-ketoglutarate dehydrogenase; (4) glutamate:succinate semialdehyde transaminase; (5) glutamate decarboxylase; (6) CoA-dependent succinic semialdehyde dehydrogenase; (7) 4-hydroxybutanoate dehydrogenase; (8) .alpha.-ketoglutarate decarboxylase; (9) 4-hydroxybutyryl CoA:acetyl-CoA transferase; (10) butyrate kinase; (11) phosphotransbutyrylase; (12) aldehyde dehydrogenase; (13) alcohol dehydrogenase.
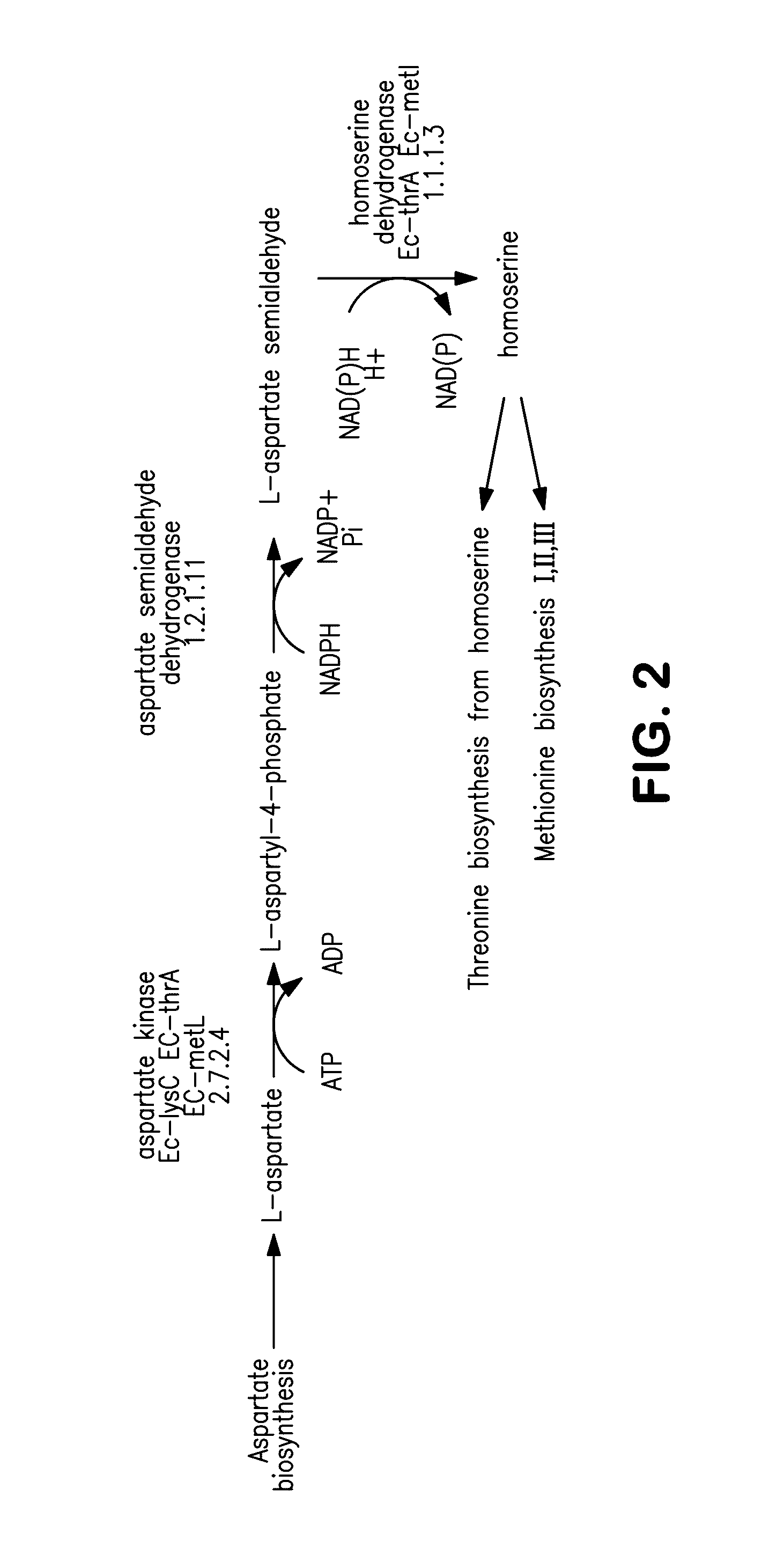
[0010] FIG. 2 is a schematic diagram showing homoserine biosynthesis in E. coli.
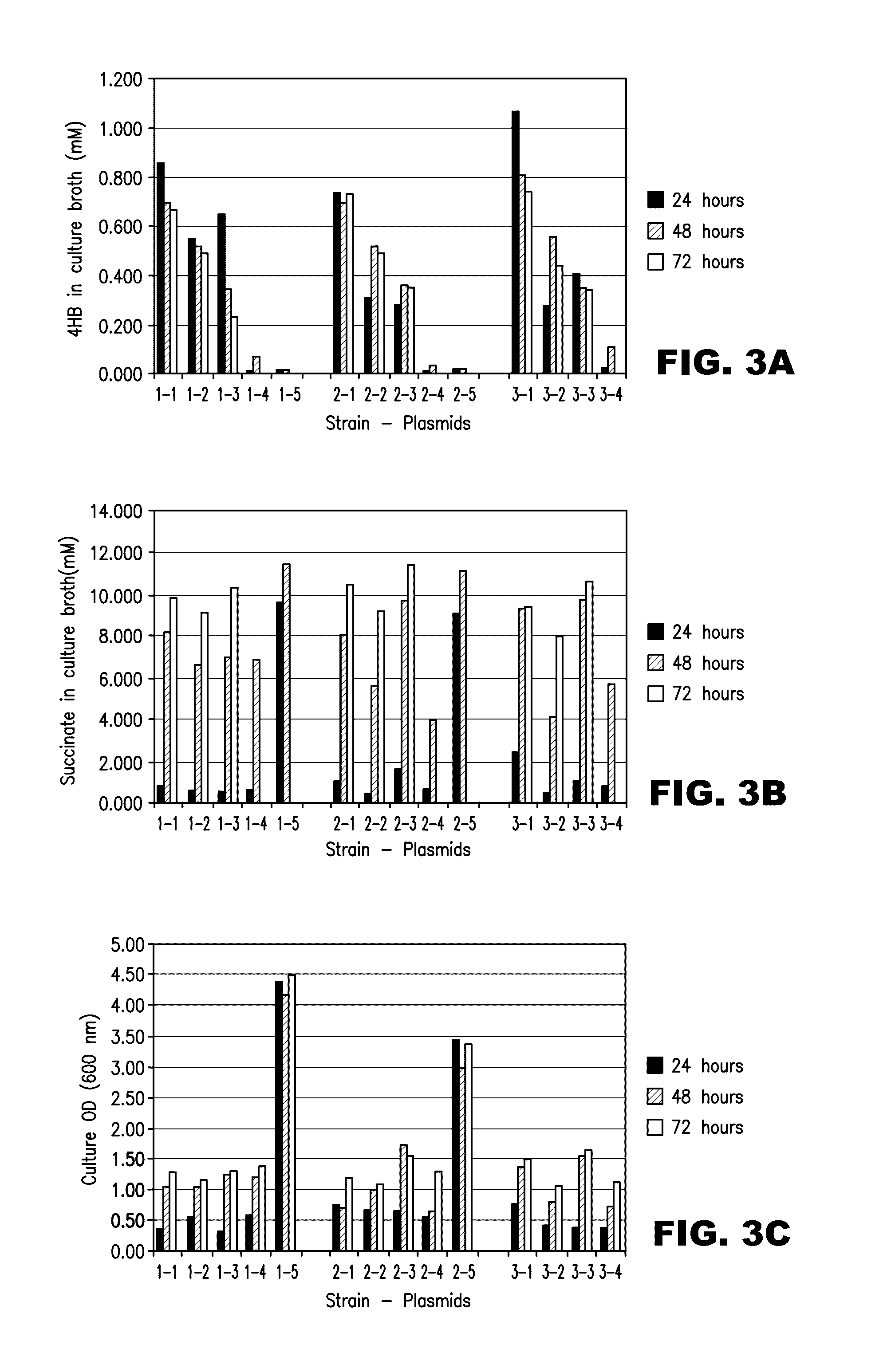
[0011] FIGS. 3A-3C show the production of 4-HB in glucose minimal medium using E. coli strains harboring plasmids expressing various combinations of 4-HB pathway genes. FIG. 3A: 4-HB concentration in culture broth; FIG. 3B: succinate concentration in culture broth; FIG. 3C: culture OD, measured at 600 nm. Clusters of bars represent the 24 hour, 48 hour, and 72 hour (if measured) timepoints. The codes along the x-axis indicate the strain/plasmid combination used. The first index refers to the host strain: 1, MG1655 lacI.sup.Q; 2, MG1655 .DELTA.gabD lacI.sup.Q; 3, MG1655 .DELTA.gabD .DELTA.aldA lacI.sup.Q. The second index refers to the plasmid combination used: 1, pZE13-0004-0035 and pZA33-0036; 2, pZE13-0004-0035 and pZA33-0010n; 3, pZE13-0004-0008 and pZA33-0036; 4, pZE13-0004-0008 and pZA33-0010n; 5, Control vectors pZE13 and pZA33.
[0012] FIG. 4 shows the production of 4-HB from glucose in E. coli strains expressing .alpha.-ketoglutarate decarboxylase from Mycobacterium tuberculosis. Strains 1-3 contain pZE13-0032 and pZA33-0036. Strain 4 expresses only the empty vectors pZE13 and pZA33. Host strains are as follows: 1 and 4, MG1655 lacI.sup.Q; 2, MG1655 .DELTA.gabD lacI.sup.Q; 3, MG1655 .DELTA.gabD .DELTA.aldA lacI.sup.Q. The bars refer to concentration at 24 and 48 hours.
[0013] FIG. 5 shows the production of BDO from 10 mM 4-HB in recombinant E. coli strains. Numbered positions correspond to experiments with MG1655 lacI.sup.Q containing pZA33-0024, expressing cat2 from P. gingivalis, and the following genes expressed on pZE13: 1, none (control); 2, 0002; 3, 0003; 4, 0003n; 5, 0011; 6, 0013; 7, 0023; 8, 0025; 9, 0008n; 10, 0035. Gene numbers are defined in Table 6. For each position, the bars refer to aerobic, microaerobic, and anaerobic conditions, respectively. Microaerobic conditions were created by sealing the culture tubes but not evacuating them.
[0014] FIGS. 6A-6H shows the mass spectrum of 4-HB and BDO produced by MG1655 lacI.sup.Q pZE13-0004-0035-0002 pZA33-0034-0036 grown in M9 minimal medium supplemented with 4 g/L unlabeled glucose (FIGS. 6A, 6C, 6E and 6G) uniformly labeled .sup.13C-glucose (FIGS. 6B, 6D, 6F and 6H). FIGS. 6A and 6B, mass 116 characteristic fragment of derivatized BDO, containing 2 carbon atoms; FIGS. 6C and 6D, mass 177 characteristic fragment of derivatized BDO, containing 1 carbon atom; FIGS. 6E and 6F, mass 117 characteristic fragment of derivatized 4-HB, containing 2 carbon atoms; FIGS. 6G and 6H, mass 233 characteristic fragment of derivatized 4-HB, containing 4 carbon atoms.
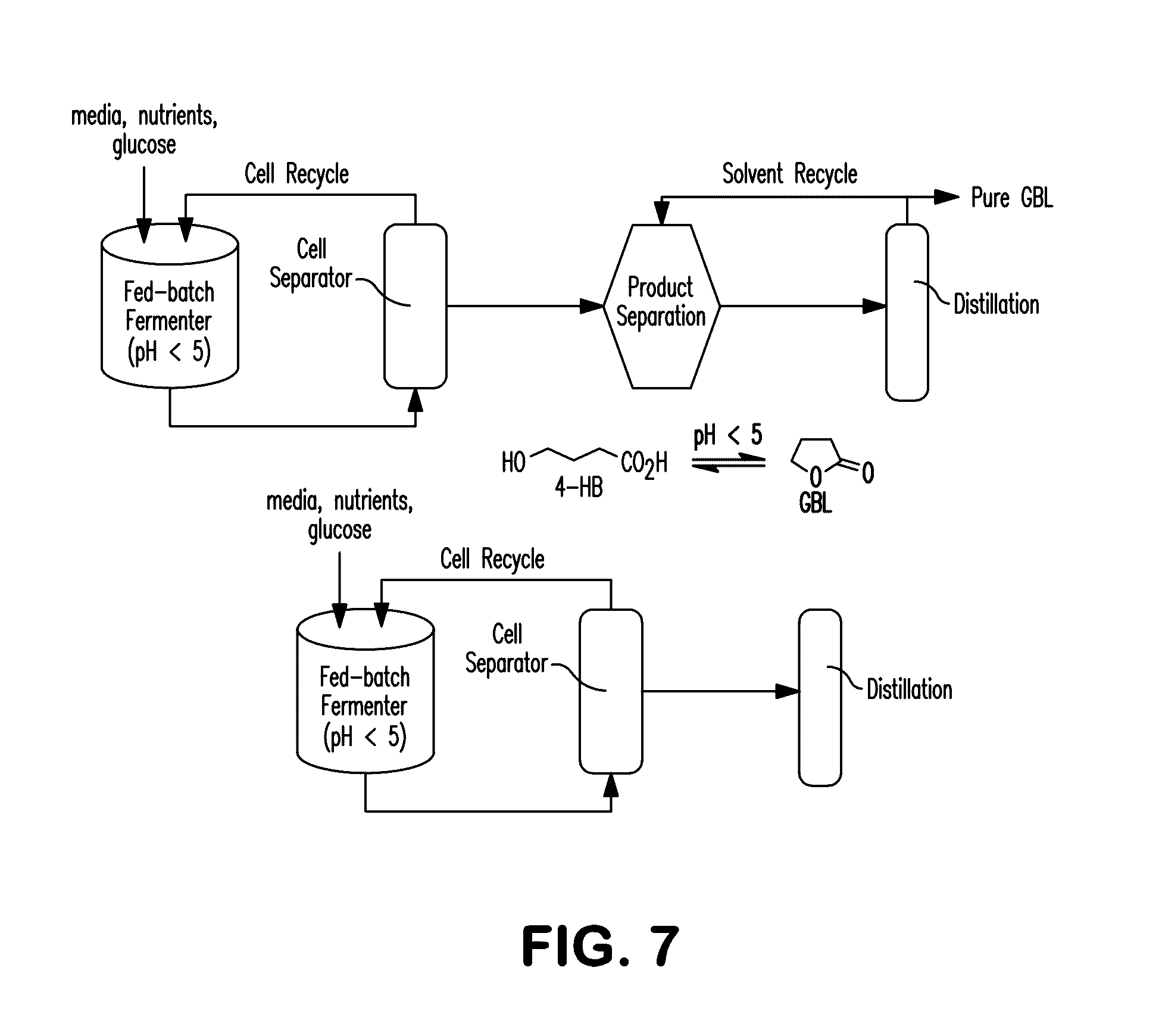
[0015] FIG. 7 is a schematic process flow diagram of bioprocesses for the production of .gamma.-butyrolactone. Panel (a) illustrates fed-batch fermentation with batch separation and panel (b) illustrates fed-batch fermentation with continuous separation.
[0016] FIGS. 8A and 8B show exemplary 1,4-butanediol (BDO) pathways. FIG. 8A shows BDO pathways from succinyl-CoA. FIG. 8B shows BDO pathways from alpha-ketoglutarate.
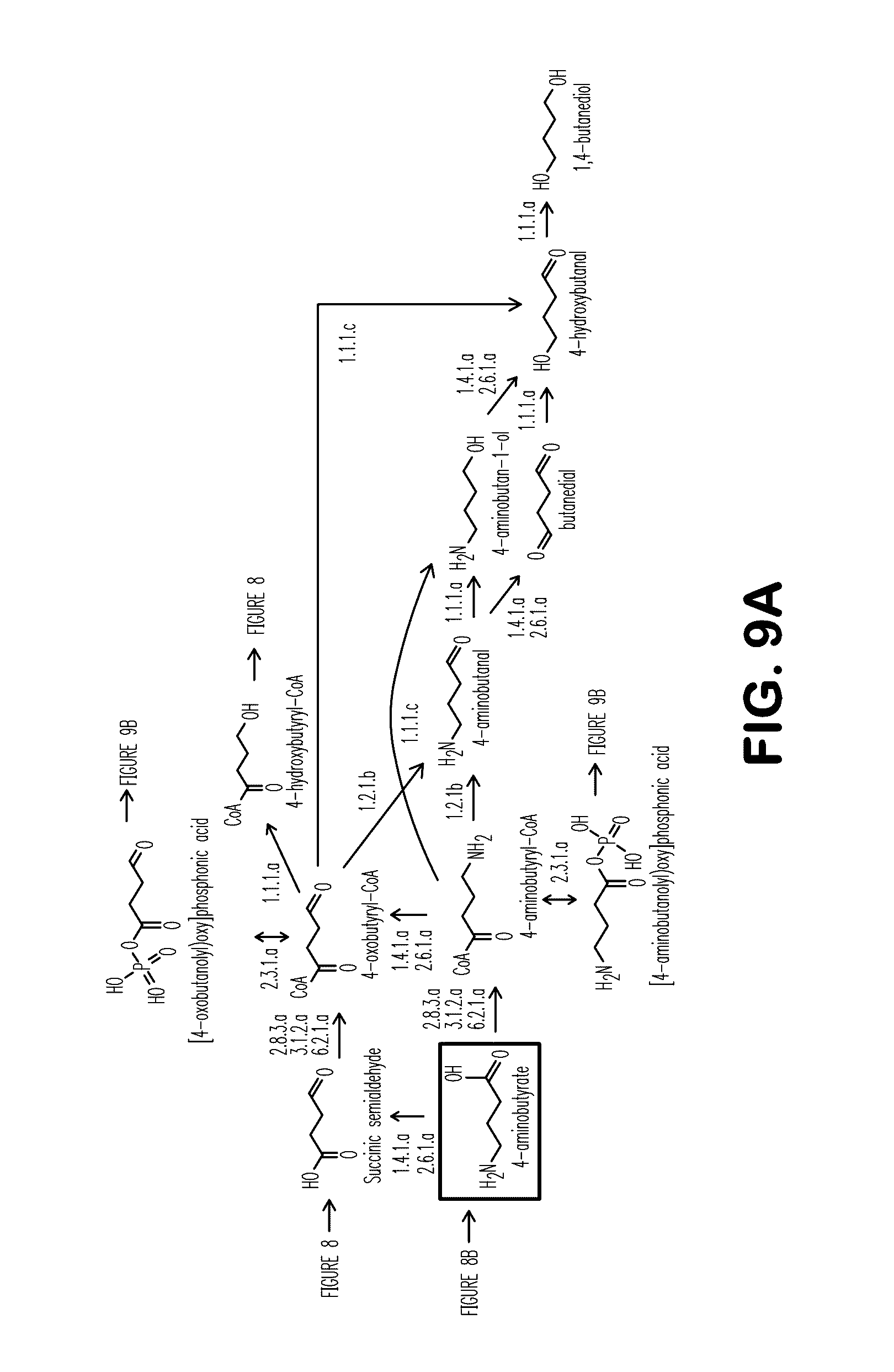
[0017] FIGS. 9A-9C show exemplary BDO pathways. FIGS. 9A and 9B show pathways from 4-aminobutyrate. FIG. 9C shows a pathway from acetoactyl-CoA to 4-aminobutyrate.
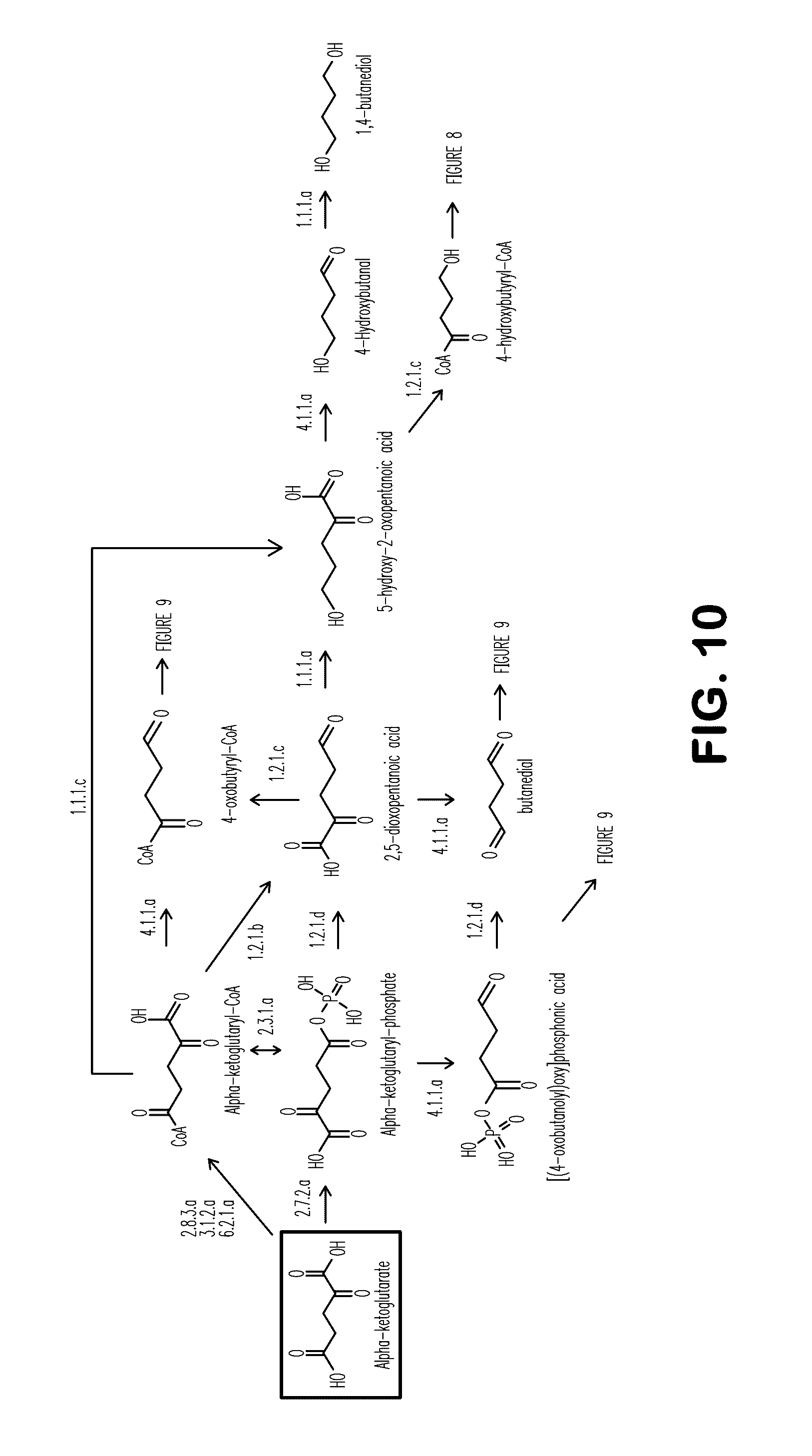
[0018] FIG. 10 shows exemplary BDO pathways from alpha-ketoglutarate.
[0019] FIG. 11 shows exemplary BDO pathways from glutamate.
[0020] FIG. 12 shows exemplary BDO pathways from acetoacetyl-CoA.
[0021] FIG. 13 shows exemplary BDO pathways from homoserine.
[0022] FIGS. 14A-14C show the nucleotide and amino acid sequences of E. coli succinyl-CoA synthetase. FIG. 14A shows the nucleotide sequence (SEQ ID NO:46) of the E. coli sucCD operon. FIGS. 14B (SEQ ID NO:47) and 14C (SEQ ID NO:48) show the amino acid sequences of the succinyl-CoA synthetase subunits encoded by the sucCD operon.
[0023] FIGS. 15A and 15B show the nucleotide and amino acid sequences of Mycobacterium bovis alpha-ketoglutarate decarboxylase. FIG. 15A shows the nucleotide sequence (SEQ ID NO:49) of Mycobacterium bovis sucA gene. FIG. 15B shows the amino acid sequence (SEQ ID NO:50) of M. bovis alpha-ketoglutarate decarboxylase.
[0024] FIG. 16 shows biosynthesis in E. coli of 4-hydroxybutyrate from glucose in minimal medium via alpha-ketoglutarate under anaerobic (microaerobic) conditions. The host strain is ECKh-401. The experiments are labeled based on the upstream pathway genes present on the plasmid pZA33 as follows: 1) 4hbd-sucA; 2) sucCD-sucD-4hbd; 3) sucCD-sucD-4hbd-sucA.
[0025] FIG. 17 shows biosynthesis in E. coli of 4-hydroxybutyrate from glucose in minimal medium via succinate and alpha-ketoglutarate. The host strain is wild-type MG1655. The experiments are labeled based on the genes present on the plasmids pZE13 and pZA33 as follows: 1) empty control vectors 2) empty pZE13, pZA33-4hbd; 3) pZE13-sucA, pZA33-4hbd.
[0026] FIG. 18A shows the nucleotide sequence (SEQ ID NO:51) of CoA-dependent succinate semialdehyde dehydrogenase (sucD) from Porphyromonas gingivalis, and FIG. 18B shows the encoded amino acid sequence (SEQ ID NO:52).
[0027] FIG. 19A shows the nucleotide sequence (SEQ ID NO:53) of 4-hydroxybutyrate dehydrogenase (4hbd) from Porphymonas gingivalis, and FIG. 19B shows the encoded amino acid sequence (SEQ ID NO:54).
[0028] FIG. 20A shows the nucleotide sequence (SEQ ID NO:55) of 4-hydroxybutyrate CoA transferase (cat2) from Porphyromonas gingivalis, and FIG. 20B shows the encoded amino acid sequence (SEQ ID NO:56).
[0029] FIG. 21A shows the nucleotide sequence (SEQ ID NO:57) of phosphotransbutyrylase (ptb) from Clostridium acetobutylicum, and FIG. 21B shows the encoded amino acid sequence (SEQ ID NO:58).
[0030] FIG. 22A shows the nucleotide sequence (SEQ ID NO:59) of butyrate kinase (buk1) from Clostridium acetobutylicum, and FIG. 22B shows the encoded amino acid sequence (SEQ ID NO:60).
[0031] FIGS. 23A-23D show alternative nucleotide sequences for C. acetobutylicum 020 (phosphtransbutyrylase) with altered codons for more prevalent E. coli codons relative to the C. acetobutylicum native sequence. FIGS. 23A-23D (020A-020D, SEQ ID NOS:61-64, respectively) contain sequences with increasing numbers of rare E. coli codons replaced by more prevalent codons (A<B<C<D).
[0032] FIGS. 24A-24D show alternative nucleotide sequences for C. acetobuytlicum 021 (butyrate kinase) with altered codons for more prevalent E. coli codons relative to the C. acetobutylicum native sequence. FIGS. 24A-24D (021A-021B, SEQ ID NOS:65-68, respectively) contain sequences with increasing numbers of rare E. coli codons replaced by more prevalent codons (A<B<C<D).
[0033] FIGS. 25A and 25B show improved expression of butyrate kinase (BK) and phosphotransbutyrylase (PTB) with optimized codons for expression in E. coli. FIG. 25A shows sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) stained for proteins with Coomassie blue; lane 1, control vector with no insert; lane 2, expression of C. acetobutylicum native sequences in E. coli; lane 3, expression of 020B-021B codon optimized PTB-BK; lane 4, expression of 020C-021C codon optimized PTB-BK. The positions of BK and PTB are shown. FIG. 25B shows the BK and PTB activities of native C. acetobutylicum sequence (2021n) compared to codon optimized 020B-021B (2021B) and 020C-021C (2021C).
[0034] FIG. 26 shows production of BDO and gamma-butyrylactone (GBL) in various strains expressing BDO producing enzymes: Cat2 (034); 2021n; 2021B; 2021C.
[0035] FIG. 27A shows the nucleotide sequence (SEQ ID NO:69) of the native Clostridium biejerinckii ald gene (025n), and FIG. 27B shows the encoded amino acid sequence (SEQ ID NO:70).
[0036] FIGS. 28A-28D show alternative gene sequences for the Clostridium beijerinckii ald gene (025A-025D, SEQ ID NOS:71-74, respectively), in which increasing numbers of rare codons are replaced by more prevalent codons (A<B<C<D).
[0037] FIG. 29 shows expression of native C. beijerinckii ald gene and codon optimized variants; no ins (control with no insert), 025n, 025A, 025B, 025C, 025D.
[0038] FIGS. 30A and 30B show BDO or BDO and ethanol production in various strains. FIG. 30A shows BDO production in strains containing the native C. beijerinckii ald gene (025n) or variants with optimized codons for expression in E. coli (025A-025D). FIG. 30B shows production of ethanol and BDO in strains expressing the C. acetobutylicum AdhE2 enzyme (002C) compared to the codon optimized variant 025B. The third set shows expression of P. gingivalis sucD (035). In all cases, P. gingivalis Cat2 (034) is also expressed.
[0039] FIG. 31A shows the nucleotide sequence (SEQ ID NO:75) of the adh1 gene from Geobacillus thermoglucosidasius, and FIG. 31B shows the encoded amino acid sequence (SEQ ID NO:76).
[0040] FIG. 32A shows the expression of the Geobacillus thermoglucosidasius adh1 gene in E. coli. Either whole cell lysates or supernatants were analyzed by SDS-PAGE and stained with Coomassie blue for plasmid with no insert, plasmid with 083 (Geotrichum capitatum N-benzyl-3-pyrrolidinol dehydrogenase) and plasmid with 084 (Geobacillus thermoglucosidasius adh1) inserts. FIG. 32B shows the activity of 084 with butyraldehyde (diamonds) or 4-hydroxybutyraldehyde (squares) as substrates.
[0041] FIG. 33 shows the production of BDO in various strains: plasmid with no insert; 025B, 025B-026n; 025B-026A; 025B-026B; 025B-026C; 025B-050; 025B-052; 025B-053; 025B-055; 025B-057; 025B-058; 025B-071; 025B-083; 025B-084; PTSlacO-025B; PTSlacO-025B-026n.
[0042] FIG. 34 shows a plasmid map for the vector pRE118-V2.
[0043] FIG. 35 shows the sequence (SEQ ID NO:77) of the ECKh-138 region encompassing the aceF and lpdA genes. The K. pneumonia lpdA gene is underlined, and the codon changed in the Glu354Lys mutant shaded.
[0044] FIG. 36 shows the protein sequence comparison of the native E. coli lpdA (SEQ ID NO:78) and the mutant K. pneumonia lpdA (SEQ ID NO:79).
[0045] FIG. 37 shows 4-hydroxybutyrate (left bars) and BDO (right bars) production in the strains AB3, MG1655 .DELTA.ldhA and ECKh-138. All strains expressed E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd on the medium copy plasmid pZA33, and P. gingivalis Cat2, C. acetobutylicum AdhE2 on the high copy plasmid pZE13.
[0046] FIG. 38 shows the nucleotide sequence (SEQ ID NO:80) of the 5' end of the aceE gene fused to the pflB-p6 promoter and ribosome binding site (RBS). The 5' italicized sequence shows the start of the aroP gene, which is transcribed in the opposite direction from the pdh operon. The 3' italicized sequence shows the start of the aceE gene. In upper case: pflB RBS. Underlined: FNR binding site. In bold: pflB-p6 promoter sequence.
[0047] FIG. 39 shows the nucleotide sequence (SEQ ID NO:81) in the aceF-lpdA region in the strain ECKh-456.
[0048] FIG. 40 shows the production of 4-hydroxybutyrate, BDO and pyruvate (left to right bars, respectively) for each of strains ECKh-439, ECKh-455 and ECKh-456.
[0049] FIG. 41A shows a schematic of the recombination sites for deletion of the mdh gene. FIG. 41B shows the sequence (nucleotide sequence, SEQ ID NO:82; amino acid sequence, SEQ ID NO:83) of the PCR product of the amplification of chloramphenicol resistance gene (CAT) flanked by FRT sites and homology regions from the mdh gene from the plasmid pKD3.
[0050] FIG. 42 shows the sequence (SEQ ID NO:84) of the arcA deleted region in strain ECKh-401.
[0051] FIG. 43 shows the sequence (SEQ ID NO:85) of the region encompassing a mutated gltA gene of strain ECKh-422.
[0052] FIGS. 44A and 44B show the citrate synthase activity of wild type gltA gene product and the R163L mutant. The assay was performed in the absence (diamonds) or presence of 0.4 mM NADH (squares).
[0053] FIG. 45 shows the 4-hydroxybutyrate (left bars) and BDO (right bars) production in strains ECKh-401 and ECKh-422, both expressing genes for the complete BDO pathway on plasmids.
[0054] FIG. 46 shows central metabolic fluxes and associated 95% confidence intervals from metabolic labeling experiments. Values are molar fluxes normalized to a glucose uptake rate of 1 mmol/hr. The result indicates that carbon flux is routed through citrate synthase in the oxidative direction and that most of the carbon enters the BDO pathway rather than completing the TCA cycle.
[0055] FIGS. 47A and 47B show extracellular product formation for strains ECKh-138 and ECKh-422, both expressing the entire BDO pathway on plasmids. The products measured were acetate (Ace), pyruvate (Pyr), 4-hydroxybutyrate (4HB), 1,4-butanediol (BDO), ethanol (EtOH), and other products, which include gamma-butyrolactone (GBL), succinate, and lactate.
[0056] FIG. 48 shows the sequence (SEQ ID NO:86) of the region following replacement of PEP carboxylase (ppc) by H. influenzae phosphoenolpyruvate carboxykinase (pepck). The pepck coding region is underlined.
[0057] FIG. 49 shows growth of evolved pepCK strains grown in minimal medium containing 50 mM NaHCO.sub.3.
[0058] FIG. 50 shows product formation in strain ECKh-453 expressing P. gingivalis Cat2 and C. beijerinckii Ald on the plasmid pZS*13. The products measured were 1,4-butanediol (BDO), pyruvate, 4-hydroxybutyrate (4HB), acetate, .gamma.-butyrolactone (GBL) and ethanol.
[0059] FIG. 51 shows BDO production of two strains, ECKh-453 and ECKh-432. Both contain the plasmid pZS*13 expressing P. gingivalis Cat2 and C. beijerinckii Ald. The cultures were grown under microaerobic conditions, with the vessels punctured with 27 or 18 gauge needles, as indicated.
[0060] FIG. 52 shows the nucleotide sequence (SEQ ID NO:87) of the genomic DNA of strain ECKh-426 in the region of insertion of a polycistronic DNA fragment containing a promoter, sucCD gene, sucD gene, 4hbd gene and a terminator sequence.
[0061] FIG. 53 shows the nucleotide sequence (SEQ ID NO:88) of the chromosomal region of strain ECKh-432 in the region of insertion of a polycistronic sequence containing a promoter, sucA gene, C. kluyveri 4hbd gene and a terminator sequence.
[0062] FIG. 54 shows BDO synthesis from glucose in minimal medium in the ECKh-432 strain having upstream BDO pathway encoding genes integrated into the chromosome and containing a plasmid harboring downstream BDO pathway genes.
[0063] FIG. 55 shows a PCR product (SEQ ID NO:89) containing the non-phosphotransferase (non-PTS) sucrose utilization genes flanked by regions of homology to the rrnC region.
[0064] FIG. 56 shows a schematic diagram of the integrations site in the rrnC operon.
[0065] FIG. 57 shows average product concentration, normalized to culture OD600, after 48 hours of growth of strain ECKh-432 grown on glucose and strain ECKh-463 grown on sucrose. Both contain the plasmid pZS*13 expressing P. gingivalis Cat2 and C. beijerinckii Ald. The data is for 6 replicate cultures of each strain. The products measured were 1,4-butanediol (BDO), 4-hydroxybutyrate (4HB), .gamma.-butyrolactone (GBL), pyruvate (PYR) and acetate (ACE) (left to right bars, respectively).
[0066] FIG. 58 shows exemplary pathways to 1,4-butanediol from succcinyl-CoA and alpha-ketoglutarate. Abbreviations: A) Succinyl-CoA reductase (aldehyde forming), B) Alpha-ketoglutarate decarboxylase, C) 4-Hydroxybutyrate dehydrogenase, D) 4-Hydroxybutyrate reductase, E) 1,4-Butanediol dehydrogenase.
[0067] FIG. 59A shows the nucleotide sequence (SEQ ID NO:90) of carboxylic acid reductase from Nocardia iowensis (GNM_720), and FIG. 59B shows the encoded amino acid sequence (SEQ ID NO:91).
[0068] FIG. 60A shows the nucleotide sequence (SEQ ID NO:92) of phosphpantetheine transferase, which was codon optimized, and FIG. 60B shows the encoded amino acid sequence (SEQ ID NO:93).
[0069] FIG. 61 shows a plasmid map of plasmid pZS*-13S-720 721opt.
[0070] FIGS. 62A and 62B show pathways to 1,4-butanediol from succinate, succcinyl-CoA, and alpha-ketoglutarate. Abbreviations: A) Succinyl-CoA reductase (aldehyde forming), B) Alpha-ketoglutarate decarboxylase, C) 4-Hydroxybutyrate dehydrogenase, D) 4-Hydroxybutyrate reductase, E) 1,4-Butanediol dehydrogenase, F) Succinate reductase, G) Succinyl-CoA transferase, H) Succinyl-CoA hydrolase, I) Succinyl-CoA synthetase (or Succinyl-CoA ligase), J) Glutamate dehydrogenase, K) Glutamate transaminase, L) Glutamate decarboxylase, M) 4-aminobutyrate dehydrogenase, N) 4-aminobutyrate transaminase, O) 4-Hydroxybutyrate kinase, P) Phosphotrans-4-hydroxybutyrylase, Q) 4-Hydroxybutyryl-CoA reductase (aldehyde forming), R) 4-hydroxybutyryl-phosphate reductase, S) Succinyl-CoA reductase (alcohol forming), T) 4-Hydroxybutyryl-CoA transferase, U) 4-Hydroxybutyryl-CoA hydrolase, V) 4-Hydroxybutyryl-CoA synthetase (or 4-Hydroxybutyryl-CoA ligase), W) 4-Hydroxybutyryl-CoA reductase (alcohol forming), X) Alpha-ketoglutarate reductase, Y) 5-Hydroxy-2-oxopentanoate dehydrogenase, Z) 5-Hydroxy-2-oxopentanoate decarboxylase, AA) 5-hydroxy-2-oxopentanoate dehydrogenase (decarboxylation).
[0071] FIG. 63 shows pathways to putrescine from succinate, succcinyl-CoA, and alpha-ketoglutarate. Abbreviations: A) Succinyl-CoA reductase (aldehyde forming), B) Alpha-ketoglutarate decarboxylase, C) 4-Aminobutyrate reductase, D) Putrescine dehydrogenase, E) Putrescine transaminase, F) Succinate reductase, G) Succinyl-CoA transferase, H) Succinyl-CoA hydrolase, I) Succinyl-CoA synthetase (or Succinyl-CoA ligase), J) Glutamate dehydrogenase, K) Glutamate transaminase, L) Glutamate decarboxylase, M) 4-Aminobutyrate dehydrogenase, N) 4-Aminobutyrate transaminase, O) Alpha-ketoglutarate reductase, P) 5-Amino-2-oxopentanoate dehydrogenase, Q) 5-Amino-2-oxopentanoate transaminase, R) 5-Amino-2-oxopentanoate decarboxylase, S) Ornithine dehydrogenase, T) Ornithine transaminase, U) Ornithine decarboxylase.
DETAILED DESCRIPTION OF THE INVENTION
[0072] The present invention is directed to the design and production of cells and organisms having biosynthetic production capabilities for 4-hydroxybutanoic acid (4-HB), .gamma.-butyrolactone, 1,4-butanediol (BDO), 4-hydroxybutanal (4-HBal), 4-hydroxybutyryl-CoA (4-HBCoA) and/or putrescine. The invention, in particular, relates to the design of microbial organisms capable of producing BDO, 4-HBal, 4-HBCoA and/or putrescine by introducing one or more nucleic acids encoding a BDO, 4-HBal, 4-HBCoA and/or putrescine pathway enzyme.
[0073] In one embodiment, the invention utilizes in silico stoichiometric models of Escherichia coli metabolism that identify metabolic designs for biosynthetic production of 4-hydroxybutanoic acid (4-HB), 1,4-butanediol (BDO), 4-HBal, 4-HBCoA and/or putrescine. The results described herein indicate that metabolic pathways can be designed and recombinantly engineered to achieve the biosynthesis of 4-HBal, 4-HBCoA or 4-HB and downstream products such as 1,4-butanediol or putrescine in Escherichia coli and other cells or organisms. Biosynthetic production of 4-HB, 4-HBal, 4-HBCoA, BDO and/or putrescine, for example, for the in silico designs can be confirmed by construction of strains having the designed metabolic genotype. These metabolically engineered cells or organisms also can be subjected to adaptive evolution to further augment 4-HB, 4-HBal, 4-HBCoA, BDO and/or putrescine biosynthesis, including under conditions approaching theoretical maximum growth.
[0074] In certain embodiments, the 4-HB, 4-HBal, 4-HBCoA, BDO and/or putrescine biosynthesis characteristics of the designed strains make them genetically stable and particularly useful in continuous bioprocesses. Separate strain design strategies were identified with incorporation of different non-native or heterologous reaction capabilities into E. coli or other host organisms leading to 4-HB and 1,4-butanediol producing metabolic pathways from either CoA-independent succinic semialdehyde dehydrogenase, succinyl-CoA synthetase and CoA-dependent succinic semialdehyde dehydrogenase, or glutamate: succinic semialdehyde transaminase. In silico metabolic designs were identified that resulted in the biosynthesis of 4-HB in both E. coli and yeast species from each of these metabolic pathways. The 1,4-butanediol intermediate .gamma.-butyrolactone can be generated in culture by spontaneous cyclization under conditions at pH<7.5, particularly under acidic conditions, such as below pH 5.5, for example, pH<7, pH<6.5, pH<6, and particularly at pH<5.5 or lower.
[0075] Strains identified via the computational component of the platform can be put into actual production by genetically engineering any of the predicted metabolic alterations which lead to the biosynthetic production of 4-HB, 1,4-butanediol or other intermediate and/or downstream products. In yet a further embodiment, strains exhibiting biosynthetic production of these compounds can be further subjected to adaptive evolution to further augment product biosynthesis. The levels of product biosynthesis yield following adaptive evolution also can be predicted by the computational component of the system.
[0076] In other specific embodiments, microbial organisms were constructed to express a 4-HB biosynthetic pathway encoding the enzymatic steps from succinate to 4-HB and to 4-HB-CoA. Co-expression of succinate coenzyme A transferase, CoA-dependent succinic semialdehyde dehydrogenase, NAD-dependent 4-hydroxybutyrate dehydrogenase and 4-hydroxybutyrate coenzyme A transferase in a host microbial organism resulted in significant production of 4-HB compared to host microbial organisms lacking a 4-HB biosynthetic pathway. In a further specific embodiment, 4-HB-producing microbial organisms were generated that utilized .alpha.-ketoglutarate as a substrate by introducing nucleic acids encoding .alpha.-ketoglutarate decarboxylase and NAD-dependent 4-hydroxybutyrate dehydrogenase.
[0077] In another specific embodiment, microbial organisms containing a 1,4-butanediol (BDO) biosynthetic pathway were constructed that biosynthesized BDO when cultured in the presence of 4-HB. The BDO biosynthetic pathway consisted of a nucleic acid encoding either a multifunctional aldehyde/alcohol dehydrogenase or nucleic acids encoding an aldehyde dehydrogenawse and an alcohol dehydrogenase. To support growth on 4-HB substrates, these BDO-producing microbial organisms also expressed 4-hydroxybutyrate CoA transferase or 4-butyrate kinase in conjunction with phosphotranshydroxybutyrlase. In yet a further specific embodiment, microbial organisms were generated that synthesized BDO through exogenous expression of nucleic acids encoding a functional 4-HB biosynthetic pathway and a functional BDO biosynthetic pathway. The 4-HB biosynthetic pathway consisted of succinate coenzyme A transferase, CoA-dependent succinic semialdehyde dehydrogenase, NAD-dependent 4-hydroxybutyrate dehydrogenase and 4-hydroxybutyrate coenzyme A transferase. The BDO pathway consisted of a multifunctional aldehyde/alcohol dehydrogenase. Further described herein are additional pathways for production of BDO (see FIGS. 8-13).
[0078] In a further embodiment, described herein is the cloning and expression of a carboxylic acid reductase enzyme that functions in a 4-hydroxybutanal, 4-hydroxybutyryl-CoA or 1,4-butanediol metabolic pathway. Advantages of employing a carboxylic acid reductase as opposed to an acyl-CoA reductase to form 4-hydroxybutyraldehyde (4-hydroxybutanal) include lower ethanol and GBL byproduct formation accompanying the production of BDO. Also disclosed herein is the application of carboxylic acid reductase as part of additional numerous pathways to produce 1,4-butanediol and putrescine from the tricarboxylic acid (TCA) cycle metabolites, for example, succinate, succinyl-CoA, and/or alpha-ketoglutarate.
[0079] As used herein, the term "non-naturally occurring" when used in reference to a microbial organism or microorganism of the invention is intended to mean that the microbial organism has at least one genetic alteration not normally found in a naturally occurring strain of the referenced species, including wild-type strains of the referenced species. Genetic alterations include, for example, modifications introducing expressible nucleic acids encoding metabolic polypeptides, other nucleic acid additions, nucleic acid deletions and/or other functional disruption of the microbial organism's genetic material. Such modifications include, for example, coding regions and functional fragments thereof, for heterologous, homologous or both heterologous and homologous polypeptides for the referenced species. Additional modifications include, for example, non-coding regulatory regions in which the modifications alter expression of a gene or operon. Exemplary metabolic polypeptides include enzymes or proteins within a biosynthetic pathway for a 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine family of compounds.
[0080] A metabolic modification refers to a biochemical reaction that is altered from its naturally occurring state. Therefore, non-naturally occurring microorganisms can have genetic modifications to nucleic acids encoding metabolic polypeptides or, functional fragments thereof. Exemplary metabolic modifications are disclosed herein.
[0081] As used herein, the term "isolated" when used in reference to a microbial organism is intended to mean an organism that is substantially free of at least one component as the referenced microbial organism is found in nature. The term includes a microbial organism that is removed from some or all components as it is found in its natural environment. The term also includes a microbial organism that is removed from some or all components as the microbial organism is found in non-naturally occurring environments. Therefore, an isolated microbial organism is partly or completely separated from other substances as it is found in nature or as it is grown, stored or subsisted in non-naturally occurring environments. Specific examples of isolated microbial organisms include partially pure microbes, substantially pure microbes and microbes cultured in a medium that is non-naturally occurring.
[0082] As used herein, the terms "microbial," "microbial organism" or "microorganism" is intended to mean any organism that exists as a microscopic cell that is included within the domains of archaea, bacteria or eukarya. Therefore, the term is intended to encompass prokaryotic or eukaryotic cells or organisms having a microscopic size and includes bacteria, archaea and eubacteria of all species as well as eukaryotic microorganisms such as yeast and fungi. The term also includes cell cultures of any species that can be cultured for the production of a biochemical.
[0083] As used herein, the term "4-hydroxybutanoic acid" is intended to mean a 4-hydroxy derivative of butyric acid having the chemical formula C.sub.4H.sub.8O.sub.3 and a molecular mass of 104.11 g/mol (126.09 g/mol for its sodium salt). The chemical compound 4-hydroxybutanoic acid also is known in the art as 4-HB, 4-hydroxybutyrate, gamma-hydroxybutyric acid or GHB. The term as it is used herein is intended to include any of the compound's various salt forms and include, for example, 4-hydroxybutanoate and 4-hydroxybutyrate. Specific examples of salt forms for 4-HB include sodium 4-HB and potassium 4-HB. Therefore, the terms 4-hydroxybutanoic acid, 4-HB, 4-hydroxybutyrate, 4-hydroxybutanoate, gamma-hydroxybutyric acid and GHB as well as other art recognized names are used synonymously herein.
[0084] As used herein, the term "monomeric" when used in reference to 4-HB is intended to mean 4-HB in a non-polymeric or underivatized form. Specific examples of polymeric 4-HB include poly-4-hydroxybutanoic acid and copolymers of, for example, 4-HB and 3-HB. A specific example of a derivatized form of 4-HB is 4-HB-CoA. Other polymeric 4-HB forms and other derivatized forms of 4-HB also are known in the art.
[0085] As used herein, the term ".gamma.-butyrolactone" is intended to mean a lactone having the chemical formula C.sub.4H.sub.6O.sub.2 and a molecular mass of 86.089 g/mol. The chemical compound .gamma.-butyrolactone also is know in the art as GBL, butyrolactone, 1,4-lactone, 4-butyrolactone, 4-hydroxybutyric acid lactone, and gamma-hydroxybutyric acid lactone. The term as it is used herein is intended to include any of the compound's various salt forms.
[0086] As used herein, the term "1,4-butanediol" is intended to mean an alcohol derivative of the alkane butane, carrying two hydroxyl groups which has the chemical formula C.sub.4H.sub.10O.sub.2 and a molecular mass of 90.12 g/mol. The chemical compound 1,4-butanediol also is known in the art as BDO and is a chemical intermediate or precursor for a family of compounds referred to herein as BDO family of compounds.
[0087] As used herein, the term "4-hydroxybutanal" is intended to mean an aledehyde having the chemical formula C.sub.4H.sub.8O.sub.2 and a molecular mass of 88.10512 g/mol. The chemical compound 4-hydroxybutanal (4-HBal) is also known in the art as 4-hydroxybutyraldehyde.
[0088] As used herein, the term "putrescine" is intended to mean a diamine having the chemical formula C.sub.4H.sub.12N.sub.2 and a molecular mass of 88.15148 g/mol. The chemical compound putrescine is also known in the art as 1,4-butanediamine, 1,4-diaminobutane, butylenediamine, tetramethylenediamine, tetramethyldiamine, and 1,4-butylenediamine.
[0089] As used herein, the term "tetrahydrofuran" is intended to mean a heterocyclic organic compound corresponding to the fully hydrogenated analog of the aromatic compound furan which has the chemical formula C.sub.4H.sub.8O and a molecular mass of 72.11 g/mol. The chemical compound tetrahydrofuran also is known in the art as THF, tetrahydrofuran, 1,4-epoxybutane, butylene oxide, cyclotetramethylene oxide, oxacyclopentane, diethylene oxide, oxolane, furanidine, hydrofuran, tetra-methylene oxide. The term as it is used herein is intended to include any of the compound's various salt forms.
[0090] As used herein, the term "CoA" or "coenzyme A" is intended to mean an organic cofactor or prosthetic group (nonprotein portion of an enzyme) whose presence is required for the activity of many enzymes (the apoenzyme) to form an active enzyme system. Coenzyme A functions in certain condensing enzymes, acts in acetyl or other acyl group transfer and in fatty acid synthesis and oxidation, pyruvate oxidation and in other acetylation.
[0091] As used herein, the term "substantially anaerobic" when used in reference to a culture or growth condition is intended to mean that the amount of oxygen is less than about 10% of saturation for dissolved oxygen in liquid media. The term also is intended to include sealed chambers of liquid or solid medium maintained with an atmosphere of less than about 1% oxygen.
[0092] The non-naturally occurring microbal organisms of the invention can contain stable genetic alterations, which refers to microorganisms that can be cultured for greater than five generations without loss of the alteration. Generally, stable genetic alterations include modifications that persist greater than 10 generations, particularly stable modifications will persist more than about 25 generations, and more particularly, stable genetic modifications will be greater than 50 generations, including indefinitely.
[0093] Those skilled in the art will understand that the genetic alterations, including metabolic modifications exemplified herein are described with reference to a suitable source or host organism such as E. coli, yeast, or other organisms disclosed herein and their corresponding metabolic reactions or a suitable source organism for desired genetic material such as genes encoding enzymes for their corresponding metabolic reactions for a desired metabolic pathway. However, given the complete genome sequencing of a wide variety of organisms and the high level of skill in the area of genomics, those skilled in the art will readily be able to apply the teachings and guidance provided herein to essentially all other organisms. For example, the E. coli metabolic alterations exemplified herein can readily be applied to other species by incorporating the same or analogous encoding nucleic acid from species other than the referenced species. Such genetic alterations include, for example, genetic alterations of species homologs, in general, and in particular, orthologs, paralogs or nonorthologous gene displacements.
[0094] An ortholog is a gene or genes that are related by vertical descent and are responsible for substantially the same or identical functions in different organisms. For example, mouse epoxide hydrolase and human epoxide hydrolase can be considered orthologs for the biological function of hydrolysis of epoxides. Genes are related by vertical descent when, for example, they share sequence similarity of sufficient amount to indicate they are homologous, or related by evolution from a common ancestor. Genes can also be considered orthologs if they share three-dimensional structure but not necessarily sequence similarity, of a sufficient amount to indicate that they have evolved from a common ancestor to the extent that the primary sequence similarity is not identifiable. Genes that are orthologous can encode proteins with sequence similarity of about 25% to 100% amino acid sequence identity. Genes encoding proteins sharing an amino acid similarity less that 25% can also be considered to have arisen by vertical descent if their three-dimensional structure also shows similarities. Members of the serine protease family of enzymes, including tissue plasminogen activator and elastase, are considered to have arisen by vertical descent from a common ancestor.
[0095] Orthologs include genes or their encoded gene products that through, for example, evolution, have diverged in structure or overall activity. For example, where one species encodes a gene product exhibiting two functions and where such functions have been separated into distinct genes in a second species, the three genes and their corresponding products are considered to be orthologs. For the production, including growth-coupled production, of a biochemical product, those skilled in the art will understand that the orthologous gene harboring the metabolic activity to be introduced or disrupted is to be chosen for construction of the non-naturally occurring microorganism. An example of orthologs exhibiting separable activities is where distinct activities have been separated into distinct gene products between two or more species or within a single species. A specific example is the separation of elastase proteolysis and plasminogen proteolysis, two types of serine protease activity, into distinct molecules as plasminogen activator and elastase. A second example is the separation of mycoplasma 5'-3' exonuclease and Drosophila DNA polymerase III activity. The DNA polymerase from the first species can be considered an ortholog to either or both of the exonuclease or the polymerase from the second species and vice versa.
[0096] In contrast, paralogs are homologs related by, for example, duplication followed by evolutionary divergence and have similar or common, but not identical functions. Paralogs can originate or derive from, for example, the same species or from a different species. For example, microsomal epoxide hydrolase (epoxide hydrolase I) and soluble epoxide hydrolase (epoxide hydrolase II) can be considered paralogs because they represent two distinct enzymes, co-evolved from a common ancestor, that catalyze distinct reactions and have distinct functions in the same species. Paralogs are proteins from the same species with significant sequence similarity to each other suggesting that they are homologous, or related through co-evolution from a common ancestor. Groups of paralogous protein families include HipA homologs, luciferase genes, peptidases, and others.
[0097] A nonorthologous gene displacement is a nonorthologous gene from one species that can substitute for a referenced gene function in a different species. Substitution includes, for example, being able to perform substantially the same or a similar function in the species of origin compared to the referenced function in the different species. Although generally, a nonorthologous gene displacement will be identifiable as structurally related to a known gene encoding the referenced function, less structurally related but functionally similar genes and their corresponding gene products nevertheless will still fall within the meaning of the term as it is used herein. Functional similarity requires, for example, at least some structural similarity in the active site or binding region of a nonorthologous gene product compared to a gene encoding the function sought to be substituted. Therefore, a nonorthologous gene includes, for example, a paralog or an unrelated gene.
[0098] Therefore, in identifying and constructing the non-naturally occurring microbial organisms of the invention having 4-HB, GBL, 4-HBal, 4-HBCoA, BDO and/or putrescine biosynthetic capability, those skilled in the art will understand with applying the teaching and guidance provided herein to a particular species that the identification of metabolic modifications can include identification and inclusion or inactivation of orthologs. To the extent that paralogs and/or nonorthologous gene displacements are present in the referenced microorganism that encode an enzyme catalyzing a similar or substantially similar metabolic reaction, those skilled in the art also can utilize these evolutionally related genes.
[0099] Orthologs, paralogs and nonorthologous gene displacements can be determined by methods well known to those skilled in the art. For example, inspection of nucleic acid or amino acid sequences for two polypeptides will reveal sequence identity and similarities between the compared sequences. Based on such similarities, one skilled in the art can determine if the similarity is sufficiently high to indicate the proteins are related through evolution from a common ancestor. Algorithms well known to those skilled in the art, such as Align, BLAST, Clustal W and others compare and determine a raw sequence similarity or identity, and also determine the presence or significance of gaps in the sequence which can be assigned a weight or score. Such algorithms also are known in the art and are similarly applicable for determining nucleotide sequence similarity or identity. Parameters for sufficient similarity to determine relatedness are computed based on well known methods for calculating statistical similarity, or the chance of finding a similar match in a random polypeptide, and the significance of the match determined. A computer comparison of two or more sequences can, if desired, also be optimized visually by those skilled in the art. Related gene products or proteins can be expected to have a high similarity, for example, 25% to 100% sequence identity. Proteins that are unrelated can have an identity which is essentially the same as would be expected to occur by chance, if a database of sufficient size is scanned (about 5%). Sequences between 5% and 24% may or may not represent sufficient homology to conclude that the compared sequences are related. Additional statistical analysis to determine the significance of such matches given the size of the data set can be carried out to determine the relevance of these sequences.
[0100] Exemplary parameters for determining relatedness of two or more sequences using the BLAST algorithm, for example, can be as set forth below. Briefly, amino acid sequence alignments can be performed using BLASTP version 2.0.8 (Jan. 5, 1999) and the following parameters: Matrix: 0 BLOSUM62; gap open: 11; gap extension: 1; x_dropoff: 50; expect: 10.0; wordsize: 3; filter: on. Nucleic acid sequence alignments can be performed using BLASTN version 2.0.6 (Sep. 16, 1998) and the following parameters: Match: 1; mismatch: -2; gap open: 5; gap extension: 2; x_dropoff: 50; expect: 10.0; wordsize: 11; filter: off. Those skilled in the art will know what modifications can be made to the above parameters to either increase or decrease the stringency of the comparison, for example, and determine the relatedness of two or more sequences.
[0101] Disclosed herein are non-naturally occurring microbial biocatalyst or microbial organisms including a microbial organism having a 4-hydroxybutanoic acid (4-HB) biosynthetic pathway that includes at least one exogenous nucleic acid encoding 4-hydroxybutanoate dehydrogenase, CoA-independent succinic semialdehyde dehydrogenase, succinyl-CoA synthetase, CoA-dependent succinic semialdehyde dehydrogenase, glutamate: succinic semialdehyde transaminase, alpha-ketoglutarate decarboxylase, or glutamate decarboxylase, wherein the exogenous nucleic acid is expressed in sufficient amounts to produce monomeric 4-hydroxybutanoic acid (4-HB). 4-hydroxybutanoate dehydrogenase is also referred to as 4-hydroxybutyrate dehydrogenase or 4-HB dehydrogenase. Succinyl-CoA synthetase is also referred to as succinyl-CoA synthase or succinyl-CoA ligase.
[0102] Also disclosed herein is a non-naturally occurring microbial biocatalyst or microbial organism including a microbial organism having a 4-hydroxybutanoic acid (4-HB) biosynthetic pathway having at least one exogenous nucleic acid encoding 4-hydroxybutanoate dehydrogenase, succinyl-CoA synthetase, CoA-dependent succinic semialdehyde dehydrogenase, or .alpha.-ketoglutarate decarboxylase, wherein the exogenous nucleic acid is expressed in sufficient amounts to produce monomeric 4-hydroxybutanoic acid (4-HB).
[0103] The non-naturally occurring microbial biocatalysts or microbial organisms can include microbial organisms that employ combinations of metabolic reactions for biosynthetically producing the compounds of the invention. The biosynthesized compounds can be produced intracellularly and/or secreted into the culture medium. Exemplary compounds produced by the non-naturally occurring microorganisms include, for example, 4-hydroxybutanoic acid, 1,4-butanediol and .gamma.-butyrolactone.
[0104] In one embodiment, a non-naturally occurring microbial organism is engineered to produce 4-HB. This compound is one useful entry point into the 1,4-butanediol family of compounds. The biochemical reactions for formation of 4-HB from succinate, from succinate through succinyl-CoA or from .alpha.-ketoglutarate are shown in steps 1-8 of FIG. 1.
[0105] It is understood that any combination of appropriate enzymes of a BDO, 4-HBal, 4-HBCoA and/or putrescine pathway can be used so long as conversion from a starting component to the BDO, 4-HBal, 4-HBCoA and/or putrescine product is achieved. Thus, it is understood that any of the metabolic pathways disclosed herein can be utilized and that it is well understood to those skilled in the art how to select appropriate enzymes to achieve a desired pathway, as disclosed herein.
[0106] In another embodiment, disclosed herein is a non-naturally occurring microbial organism, comprising a microbial organism having a 1,4-butanediol (BDO) pathway comprising at least one exogenous nucleic acid encoding a BDO pathway enzyme expressed in a sufficient amount to produce BDO, the BDO pathway comprising 4-aminobutyrate CoA transferase, 4-aminobutyryl-CoA hydrolase, 4-aminobutyrate-CoA ligase, 4-aminobutyryl-CoA oxidoreductase (deaminating), 4-aminobutyryl-CoA transaminase, or 4-hydroxybutyryl-CoA dehydrogenase (see Example VII Table 17). The BDO pathway further can comprise 4-hydroxybutyryl-CoA reductase (alcohol forming), 4-hydroxybutyryl-CoA reductase, or 1,4-butanediol dehydrogenase.
[0107] It is understood by those skilled in the art that various combinations of the pathways can be utilized, as disclosed herein. For example, in a non-naturally occurring microbial organism, the nucleic acids can encode 4-aminobutyrate CoA transferase, 4-aminobutyryl-CoA hydrolase, or 4-aminobutyrate-CoA ligase; 4-aminobutyryl-CoA oxidoreductase (deaminating) or 4-aminobutyryl-CoA transaminase; and 4-hydroxybutyryl-CoA dehydrogenase. Other exemplary combinations are specifically describe below and further can be found in FIGS. 8-13. For example, the BDO pathway can further comprise 4-hydroxybutyryl-CoA reductase (alcohol forming), 4-hydroxybutyryl-CoA reductase, or 1,4-butanediol dehydrogenase.
[0108] Additionally disclosed herein is a non-naturally occurring microbial organism, comprising a microbial organism having a BDO pathway comprising at least one exogenous nucleic acid encoding a BDO pathway enzyme expressed in a sufficient amount to produce BDO, the BDO pathway comprising 4-aminobutyrate CoA transferase, 4-aminobutyryl-CoA hydrolase, 4-aminobutyrate-CoA ligase, 4-aminobutyryl-CoA reductase (alcohol forming), 4-aminobutyryl-CoA reductase, 4-aminobutan-1-ol dehydrogenase, 4-aminobutan-1-ol oxidoreductase (deaminating) or 4-aminobutan-1-ol transaminase (see Example VII and Table 18), and can further comprise 1,4-butanediol dehydrogenase. For example, the exogenous nucleic acids can encode 4-aminobutyrate CoA transferase, 4-aminobutyryl-CoA hydrolase, or 4-aminobutyrate-CoA ligase; 4-aminobutyryl-CoA reductase (alcohol forming); and 4-aminobutan-1-ol oxidoreductase (deaminating) or 4-aminobutan-1-ol transaminase. In addition, the nucleic acids can encode. 4-aminobutyrate CoA transferase, 4-aminobutyryl-CoA hydrolase, or 4-aminobutyrate-CoA ligase; 4-aminobutyryl-CoA reductase; 4-aminobutan-1-ol dehydrogenase; and 4-aminobutan-1-ol oxidoreductase (deaminating) or 4-aminobutan-1-ol transaminase.
[0109] Also disclosed herein is a non-naturally occurring microbial organism, comprising a microbial organism having a BDO pathway comprising at least one exogenous nucleic acid encoding a BDO pathway enzyme expressed in a sufficient amount to produce BDO, the BDO pathway comprising 4-aminobutyrate kinase, 4-aminobutyraldehyde dehydrogenase (phosphorylating), 4-aminobutan-1-ol dehydrogenase, 4-aminobutan-1-ol oxidoreductase (deaminating), 4-aminobutan-1-ol transaminase, [(4-aminobutanolyl)oxy]phosphonic acid oxidoreductase (deaminating), [(4-aminobutanolyl)oxy]phosphonic acid transaminase, 4-hydroxybutyryl-phosphate dehydrogenase, or 4-hydroxybutyraldehyde dehydrogenase (phosphorylating) (see Example VII and Table 19). For example, the exogenous nucleic acids can encode 4-aminobutyrate kinase; 4-aminobutyraldehyde dehydrogenase (phosphorylating); 4-aminobutan-1-ol dehydrogenase; and 4-aminobutan-1-ol oxidoreductase (deaminating) or 4-aminobutan-1-ol transaminase. Alternatively, the exogenous nucleic acids can encode 4-aminobutyrate kinase; [(4-aminobutanolyl)oxy]phosphonic acid oxidoreductase (deaminating) or [(4-aminobutanolyl)oxy]phosphonic acid transaminase; 4-hydroxybutyryl-phosphate dehydrogenase; and 4-hydroxybutyraldehyde dehydrogenase (phosphorylating).
[0110] Additionally disclosed herein is a non-naturally occurring microbial organism, comprising a microbial organism having a BDO pathway comprising at least one exogenous nucleic acid encoding a BDO pathway enzyme expressed in a sufficient amount to produce BDO, the BDO pathway comprising alpha-ketoglutarate 5-kinase, 2,5-dioxopentanoic semialdehyde dehydrogenase (phosphorylating), 2,5-dioxopentanoic acid reductase, alpha-ketoglutarate CoA transferase, alpha-ketoglutaryl-CoA hydrolase, alpha-ketoglutaryl-CoA ligase, alpha-ketoglutaryl-CoA reductase, 5-hydroxy-2-oxopentanoic acid dehydrogenase, alpha-ketoglutaryl-CoA reductase (alcohol forming), 5-hydroxy-2-oxopentanoic acid decarboxylase, or 5-hydroxy-2-oxopentanoic acid dehydrogenase (decarboxylation) (see Example VIII and Table 20). The BDO pathway can further comprise 4-hydroxybutyryl-CoA reductase (alcohol forming), 4-hydroxybutyryl-CoA reductase, or 1,4-butanediol dehydrogenase. For example, the exogenous nucleic acids can encode alpha-ketoglutarate 5-kinase; 2,5-dioxopentanoic semialdehyde dehydrogenase (phosphorylating); 2,5-dioxopentanoic acid reductase; and 5-hydroxy-2-oxopentanoic acid decarboxylase. Alternatively, the exogenous nucleic acids can encode alpha-ketoglutarate 5-kinase; 2,5-dioxopentanoic semialdehyde dehydrogenase (phosphorylating); 2,5-dioxopentanoic acid reductase; and 5-hydroxy-2-oxopentanoic acid dehydrogenase (decarboxylation). Alternatively, the exogenous nucleic acids can encode alpha-ketoglutarate CoA transferase, alpha-ketoglutaryl-CoA hydrolase, or alpha-ketoglutaryl-CoA ligase; alpha-ketoglutaryl-CoA reductase, 5-hydroxy-2-oxopentanoic acid dehydrogenase; and 5-hydroxy-2-oxopentanoic acid decarboxylase. In another embodiment, the exogenous nucleic acids can encode alpha-ketoglutarate CoA transferase, alpha-ketoglutaryl-CoA hydrolase, or alpha-ketoglutaryl-CoA ligase; alpha-ketoglutaryl-CoA reductase, 5-hydroxy-2-oxopentanoic acid dehydrogenase, and 5-hydroxy-2-oxopentanoic acid dehydrogenase (decarboxylation). Alternatively, the exogenous nucleic acids can encode alpha-ketoglutarate CoA transferase, alpha-ketoglutaryl-CoA hydrolase, or alpha-ketoglutaryl-CoA ligase; alpha-ketoglutaryl-CoA reductase (alcohol forming); and 5-hydroxy-2-oxopentanoic acid decarboxylase. In yet another embodiment, the exogenous nucleic acids can encode alpha-ketoglutarate CoA transferase, alpha-ketoglutaryl-CoA hydrolase, or alpha-ketoglutaryl-CoA ligase; alpha-ketoglutaryl-CoA reductase (alcohol forming); and 5-hydroxy-2-oxopentanoic acid dehydrogenase (decarboxylation).
[0111] Further disclosed herein is a non-naturally occurring microbial organism, comprising a microbial organism having a BDO pathway comprising at least one exogenous nucleic acid encoding a BDO pathway enzyme expressed in a sufficient amount to produce BDO, the BDO pathway comprising glutamate CoA transferase, glutamyl-CoA hydrolase, glutamyl-CoA ligase, glutamate 5-kinase, glutamate-5-semialdehyde dehydrogenase (phosphorylating), glutamyl-CoA reductase, glutamate-5-semialdehyde reductase, glutamyl-CoA reductase (alcohol forming), 2-amino-5-hydroxypentanoic acid oxidoreductase (deaminating), 2-amino-5-hydroxypentanoic acid transaminase, 5-hydroxy-2-oxopentanoic acid decarboxylase, 5-hydroxy-2-oxopentanoic acid dehydrogenase (decarboxylation) (see Example IX and Table 21). For example, the exogenous nucleic acids can encode glutamate CoA transferase, glutamyl-CoA hydrolase, or glutamyl-CoA ligase; glutamyl-CoA reductase; glutamate-5-semialdehyde reductase; 2-amino-5-hydroxypentanoic acid oxidoreductase (deaminating) or 2-amino-5-hydroxypentanoic acid transaminase; and 5-hydroxy-2-oxopentanoic acid decarboxylase or 5-hydroxy-2-oxopentanoic acid dehydrogenase (decarboxylation). Alternatively, the exogenous nucleic acids can encode glutamate 5-kinase; glutamate-5-semialdehyde dehydrogenase (phosphorylating); glutamate-5-semialdehyde reductase; 2-amino-5-hydroxypentanoic acid oxidoreductase (deaminating) or 2-amino-5-hydroxypentanoic acid transaminase; and 5-hydroxy-2-oxopentanoic acid decarboxylase or 5-hydroxy-2-oxopentanoic acid dehydrogenase (decarboxylation). In still another embodiment, the exogenous nucleic acids can encode glutamate CoA transferase, glutamyl-CoA hydrolase, or glutamyl-CoA ligase; glutamyl-CoA reductase (alcohol forming); 2-amino-5-hydroxypentanoic acid oxidoreductase (deaminating) or 2-amino-5-hydroxypentanoic acid transaminase; and 5-hydroxy-2-oxopentanoic acid decarboxylase or 5-hydroxy-2-oxopentanoic acid dehydrogenase (decarboxylation). In yet another embodiment, the exogenous nucleic acids can encode glutamate 5-kinase; glutamate-5-semialdehyde dehydrogenase (phosphorylating); 2-amino-5-hydroxypentanoic acid oxidoreductase (deaminating) or 2-amino-5-hydroxypentanoic acid transaminase; and 5-hydroxy-2-oxopentanoic acid decarboxylase or 5-hydroxy-2-oxopentanoic acid dehydrogenase (decarboxylation).
[0112] Also disclosed herein is a non-naturally occurring microbial organism, comprising a microbial organism having a BDO pathway comprising at least one exogenous nucleic acid encoding a BDO pathway enzyme expressed in a sufficient amount to produce BDO, the BDO pathway comprising 3-hydroxybutyryl-CoA dehydrogenase, 3-hydroxybutyryl-CoA dehydratase, vinylacetyl-CoA .DELTA.-isomerase, or 4-hydroxybutyryl-CoA dehydratase (see Example X and Table 22). For example, the exogenous nucleic acids can encode 3-hydroxybutyryl-CoA dehydrogenase; 3-hydroxybutyryl-CoA dehydratase; vinylacetyl-CoA .DELTA.-isomerase; and 4-hydroxybutyryl-CoA dehydratase.
[0113] Further disclosed herein is a non-naturally occurring microbial organism, comprising a microbial organism having a BDO pathway comprising at least one exogenous nucleic acid encoding a BDO pathway enzyme expressed in a sufficient amount to produce BDO, the BDO pathway comprising homoserine deaminase, homoserine CoA transferase, homoserine-CoA hydrolase, homoserine-CoA ligase, homoserine-CoA deaminase, 4-hydroxybut-2-enoyl-CoA transferase, 4-hydroxybut-2-enoyl-CoA hydrolase, 4-hydroxybut-2-enoyl-CoA ligase, 4-hydroxybut-2-enoate reductase, 4-hydroxybutyryl-CoA transferase, 4-hydroxybutyryl-CoA hydrolase, 4-hydroxybutyryl-CoA ligase, or 4-hydroxybut-2-enoyl-CoA reductase (see Example XI and Table 23). For example, the exogenous nucleic acids can encode homoserine deaminase; 4-hydroxybut-2-enoyl-CoA transferase, 4-hydroxybut-2-enoyl-CoA hydrolase, 4-hydroxybut-2-enoyl-CoA ligase; 4-hydroxybut-2-enoyl-CoA reductase. Alternatively, the exogenous nucleic acids can encode homoserine CoA transferase, homoserine-CoA hydrolase, or homoserine-CoA ligase; homoserine-CoA deaminase; and 4-hydroxybut-2-enoyl-CoA reductase. In a further embodiment, the exogenous nucleic acids can encode homoserine deaminase; 4-hydroxybut-2-enoate reductase; and 4-hydroxybutyryl-CoA transferase, 4-hydroxybutyryl-CoA hydrolase, or 4-hydroxybutyryl-CoA ligase. Alternatively, the exogenous nucleic acids can encode homoserine CoA transferase, homoserine-CoA hydrolase, or homoserine-CoA ligase; homoserine-CoA deaminase; and 4-hydroxybut-2-enoyl-CoA reductase.
[0114] Further disclosed herein is a non-naturally occurring microbial organism, comprising a microbial organism having a BDO pathway comprising at least one exogenous nucleic acid encoding a BDO pathway enzyme expressed in a sufficient amount to produce BOD, the BDO pathway comprising succinyl-CoA reductase (alcohol forming), 4-hydroxybutyryl-CoA hydrolase, 4-hydroxybutyryl-CoA ligase, 4-hydroxybutanal dehydrogenase (phosphorylating) (see Table 15). Such a BDO pathway can further comprise succinyl-CoA reductase, 4-hydroxybutyrate dehydrogenase, 4-hydroxybutyryl-CoA transferase, 4-hydroxybutyrate kinase, phosphotrans-4-hydroxybutyrylase, 4-hydroxybutyryl-CoA reductase, 4-hydroxybutyryl-CoA reductase (alcohol forming), or 1,4-butanediol dehydrogenase.
[0115] Additionally disclosed herein is a non-naturally occurring microbial organism, comprising a microbial organism having a BDO pathway comprising at least one exogenous nucleic acid encoding a BDO pathway enzyme expressed in a sufficient amount to produce BDO, the BDO pathway comprising glutamate dehydrogenase, 4-aminobutyrate oxidoreductase (deaminating), 4-aminobutyrate transaminase, glutamate decarboxylase, 4-hydroxybutyryl-CoA hydrolase, 4-hydroxybutyryl-CoA ligase, 4-hydroxybutanal dehydrogenase (phosphorylating)(see Table 16). Such a BDO pathway can further comprise alpha-ketoglutarate decarboxylase, 4-hydroxybutyrate dehydrogenase, 4-hydroxybutyryl-CoA transferase, 4-hydroxybutyrate kinase, phosphotrans-4-hydroxybutyrylase, 4-hydroxybutyryl-CoA reductase, 4-hydroxybutyryl-CoA reductase (alcohol forming), or 1,4-butanediol dehydrogenase.
[0116] The pathways described above are merely exemplary. One skilled in the art can readily select appropriate pathways from those disclosed herein to obtain a suitable BDO pathway or other metabolic pathway, as desired.
[0117] The invention provides genetically modified organisms that allow improved production of a desired product such as BDO by increasing the product or decreasing undesirable byproducts. As disclosed herein, the invention provides a non-naturally occurring microbial organism, comprising a microbial organism having a 1,4-butanediol (BDO) pathway comprising at least one exogenous nucleic acid encoding a BDO pathway enzyme expressed in a sufficient amount to produce BDO. In one embodiment, the microbial organism is genetically modified to express exogenous succinyl-CoA synthetase (see Example XII). For example, the succinyl-CoA synthetase can be encoded by an Escherichia coli sucCD genes.
[0118] In another embodiment, the microbial organism is genetically modified to express exogenous alpha-ketoglutarate decarboxylase (see Example XIII) For example, the alpha-ketoglutarate decarboxylase can be encoded by the Mycobacterium bovis sucA gene. In still another embodiment, the microbial organism is genetically modified to express exogenous succinate semialdehyde dehydrogenase and 4-hydroxybutyrate dehydrogenase and optionally 4-hydroxybutyryl-CoA/acetyl-CoA transferase (see Example XIII) For example, the succinate semialdehyde dehydrogenase (CoA-dependent), 4-hydroxybutyrate dehydrogenase and 4-hydroxybutyryl-CoA/acetyl-CoA transferase can be encoded by Porphyromonas gingivalis W83 genes. In an additional embodiment, the microbial organism is genetically modified to express exogenous butyrate kinase and phosphotransbutyrylase (see Example XIII) For example, the butyrate kinase and phosphotransbutyrylase can be encoded by Clostridium acetobutilicum buk1 and ptb genes.
[0119] In yet another embodiment, the microbial organism is genetically modified to express exogenous 4-hydroxybutyryl-CoA reductase (see Example XIII) For example, the 4-hydroxybutyryl-CoA reductase can be encoded by Clostridium beijerinckii ald gene. Additionally, in an embodiment of the invention, the microbial organism is genetically modified to express exogenous 4-hydroxybutanal reductase (see Example XIII) For example, the 4-hydroxybutanal reductase can be encoded by Geobacillus thermoglucosidasius adh1 gene. In another embodiment, the microbial organism is genetically modified to express exogenous pyruvate dehydrogenase subunits (see Example XIV). For example, the exogenous pyruvate dehydrogenase can be NADH insensitive. The pyruvate dehydrogenase subunit can be encoded by the Klebsiella pneumonia lpdA gene. In a particular embodiment, the pyruvate dehydrogenase subunit genes of the microbial organism can be under the control of a pyruvate formate lyase promoter.
[0120] In still another embodiment, the microbial organism is genetically modified to disrupt a gene encoding an aerobic respiratory control regulatory system (see Example XV). For example, the disruption can be of the arcA gene. Such an organism can further comprise disruption of a gene encoding malate dehydrogenase. In a further embodiment, the microbial organism is genetically modified to express an exogenous NADH insensitive citrate synthase (see Example XV). For example, the NADH insensitive citrate synthase can be encoded by gltA, such as an R163L mutant of gltA. In still another embodiment, the microbial organism is genetically modified to express exogenous phosphoenolpyruvate carboxykinase (see Example XVI). For example, the phosphoenolpyruvate carboxykinase can be encoded by an Haemophilus influenza phosphoenolpyruvate carboxykinase gene.
[0121] It is understood that any of a number of genetic modifications, as disclosed herein, can be used alone or in various combinations of one or more of the genetic modifications disclosed herein to increase the production of BDO in a BDO producing microbial organism. In a particular embodiment, the microbial organism can be genetically modified to incorporate any and up to all of the genetic modifications that lead to increased production of BDO. In a particular embodiment, the microbial organism containing a BDO pathway can be genetically modified to express exogenous succinyl-CoA synthetase; to express exogenous alpha-ketoglutarate decarboxylase; to express exogenous succinate semialdehyde dehydrogenase and 4-hydroxybutyrate dehydrogenase and optionally 4-hydroxybutyryl-CoA/acetyl-CoA transferase; to express exogenous butyrate kinase and phosphotransbutyrylase; to express exogenous 4-hydroxybutyryl-CoA reductase; and to express exogenous 4-hydroxybutanal reductase; to express exogenous pyruvate dehydrogenase; to disrupt a gene encoding an aerobic respiratory control regulatory system; to express an exogenous NADH insensitive citrate synthase; and to express exogenous phosphoenolpyruvate carboxykinase. Such strains for improved production are described in Examples XII-XIX. It is thus understood that, in addition to the modifications described above, such strains can additionally include other modifications disclosed herein. Such modifications include, but are not limited to, deletion of endogenous lactate dehydrogenase (ldhA), alcohol dehydrogenase (adhE), and/or pyruvate formate lyase (pflB)(see Examples XII-XIX and Table 28).
[0122] Additionally provided is a microbial organism in which one or more genes encoding the exogenously expressed enzymes are integrated into the fimD locus of the host organism (see Example XVII). For example, one or more genes encoding a BDO pathway enzyme can be integrated into the fimD locus for increased production of BDO. Further provided is a microbial organism expressing a non-phosphotransferase sucrose uptake system that increases production of BDO.
[0123] Although the genetically modified microbial organisms disclosed herein are exemplified with microbial organisms containing particular BDO pathway enzymes, it is understood that such modifications can be incorporated into any microbial organism having a BDO, 4-HBal, 4-HBCoA and/or putrescine pathway suitable for enhanced production in the presence of the genetic modifications. The microbial organisms of the invention can thus have any of the BDO, 4-HBal, 4-HBCoA and/or putrescine pathways disclosed herein. For example, the BDO pathway can comprise 4-hydroxybutanoate dehydrogenase, succinyl-CoA synthetase, CoA-dependent succinic semialdehyde dehydrogenase, 4-hydroxybutyrate:CoA transferase, 4-butyrate kinase, phosphotransbutyrylase, alpha-ketoglutarate decarboxylase, aldehyde dehydrogenase, alcohol dehydrogenase or an aldehyde/alcohol dehydrogenase (see FIG. 1). Alternatively, the BDO pathway can comprise 4-aminobutyrate CoA transferase, 4-aminobutyryl-CoA hydrolase, 4-aminobutyrate-CoA ligase, 4-aminobutyryl-CoA oxidoreductase (deaminating), 4-aminobutyryl-CoA transaminase, or 4-hydroxybutyryl-CoA dehydrogenase (see Table 17). Such a BDO pathway can further comprise 4-hydroxybutyryl-CoA reductase (alcohol forming), 4-hydroxybutyryl-CoA reductase, or 1,4-butanediol dehydrogenase
[0124] Additionally, the BDO pathway can comprise 4-aminobutyrate CoA transferase, 4-aminobutyryl-CoA hydrolase, 4-aminobutyrate-CoA ligase, 4-aminobutyryl-CoA reductase (alcohol forming), 4-aminobutyryl-CoA reductase, 4-aminobutan-1-ol dehydrogenase, 4-aminobutan-1-ol oxidoreductase (deaminating) or 4-aminobutan-1-ol transaminase (see Table 18). Also, the BDO pathway can comprise 4-aminobutyrate kinase, 4-aminobutyraldehyde dehydrogenase (phosphorylating), 4-aminobutan-1-ol dehydrogenase, 4-aminobutan-1-ol oxidoreductase (deaminating), 4-aminobutan-1-ol transaminase, [(4-aminobutanolyl)oxy]phosphonic acid oxidoreductase (deaminating), [(4-aminobutanolyl)oxy]phosphonic acid transaminase, 4-hydroxybutyryl-phosphate dehydrogenase, or 4-hydroxybutyraldehyde dehydrogenase (phosphorylating) (see Table 19). Such a pathway can further comprise 1,4-butanediol dehydrogenase.
[0125] The BDO pathway can also comprise alpha-ketoglutarate 5-kinase, 2,5-dioxopentanoic semialdehyde dehydrogenase (phosphorylating), 2,5-dioxopentanoic acid reductase, alpha-ketoglutarate CoA transferase, alpha-ketoglutaryl-CoA hydrolase, alpha-ketoglutaryl-CoA ligase, alpha-ketoglutaryl-CoA reductase, 5-hydroxy-2-oxopentanoic acid dehydrogenase, alpha-ketoglutaryl-CoA reductase (alcohol forming), 5-hydroxy-2-oxopentanoic acid decarboxylase, or 5-hydroxy-2-oxopentanoic acid dehydrogenase (decarboxylation)(see Table 20). Such a BDO pathway can further comprise 4-hydroxybutyryl-CoA reductase (alcohol forming), 4-hydroxybutyryl-CoA reductase, or 1,4-butanediol dehydrogenase. Additionally, the BDO pathway can comprise glutamate CoA transferase, glutamyl-CoA hydrolase, glutamyl-CoA ligase, glutamate 5-kinase, glutamate-5-semialdehyde dehydrogenase (phosphorylating), glutamyl-CoA reductase, glutamate-5-semialdehyde reductase, glutamyl-CoA reductase (alcohol forming), 2-amino-5-hydroxypentanoic acid oxidoreductase (deaminating), 2-amino-5-hydroxypentanoic acid transaminase, 5-hydroxy-2-oxopentanoic acid decarboxylase, 5-hydroxy-2-oxopentanoic acid dehydrogenase (decarboxylation)(see Table 21). Such a BDO pathway can further comprise 4-hydroxybutyryl-CoA reductase (alcohol forming), 4-hydroxybutyryl-CoA reductase, or 1,4-butanediol dehydrogenase.
[0126] Additionally, the BDO pathway can comprise 3-hydroxybutyryl-CoA dehydrogenase, 3-hydroxybutyryl-CoA dehydratase, vinylacetyl-CoA .DELTA.-isomerase, or 4-hydroxybutyryl-CoA dehydratase (see Table 22). Also, the BDO pathway can comprise homoserine deaminase, homoserine CoA transferase, homoserine-CoA hydrolase, homoserine-CoA ligase, homoserine-CoA deaminase, 4-hydroxybut-2-enoyl-CoA transferase, 4-hydroxybut-2-enoyl-CoA hydrolase, 4-hydroxybut-2-enoyl-CoA ligase, 4-hydroxybut-2-enoate reductase, 4-hydroxybutyryl-CoA transferase, 4-hydroxybutyryl-CoA hydrolase, 4-hydroxybutyryl-CoA ligase, or 4-hydroxybut-2-enoyl-CoA reductase (see Table 23). Such a BDO pathway can further comprise 4-hydroxybutyryl-CoA reductase (alcohol forming), 4-hydroxybutyryl-CoA reductase, or 1,4-butanediol dehydrogenase.
[0127] The BDO pathway can additionally comprise succinyl-CoA reductase (alcohol forming), 4-hydroxybutyryl-CoA hydrolase, 4-hydroxybutyryl-CoA ligase, or 4-hydroxybutanal dehydrogenase (phosphorylating) (see Table 15). Such a pathway can further comprise succinyl-CoA reductase, 4-hydroxybutyrate dehydrogenase, 4-hydroxybutyryl-CoA transferase, 4-hydroxybutyrate kinase, phosphotrans-4-hydroxybutyrylase, 4-hydroxybutyryl-CoA reductase, 4-hydroxybutyryl-CoA reductase (alcohol forming), or 1,4-butanediol dehydrogenase. Also, the BDO pathway can comprise glutamate dehydrogenase, 4-aminobutyrate oxidoreductase (deaminating), 4-aminobutyrate transaminase, glutamate decarboxylase, 4-hydroxybutyryl-CoA hydrolase, 4-hydroxybutyryl-CoA ligase, or 4-hydroxybutanal dehydrogenase (phosphorylating)(see Table 16). Such a BDO pathway can further comprise alpha-ketoglutarate decarboxylase, 4-hydroxybutyrate dehydrogenase, 4-hydroxybutyryl-CoA transferase, 4-hydroxybutyrate kinase, phosphotrans-4-hydroxybutyrylase, 4-hydroxybutyryl-CoA reductase, 4-hydroxybutyryl-CoA reductase (alcohol forming), or 1,4-butanediol dehydrogenase.
[0128] The invention additionally provides a non-naturally occurring microbial organism, comprising a 4-hydroxybutanal pathway comprising at least one exogenous nucleic acid encoding a 4-hydroxybutanal pathway enzyme expressed in a sufficient amount to produce 4-hydroxybutanal, the 4-hydroxybutanal pathway comprising succinyl-CoA reductase (aldehyde forming); 4-hydroxybutyrate dehydrogenase; and 4-hydroxybutyrate reductase (see FIG. 58, steps A-C-D). The invention also provides a non-naturally occurring microbial organism, comprising a 4-hydroxybutanal pathway comprising at least one exogenous nucleic acid encoding a 4-hydroxybutanal pathway enzyme expressed in a sufficient amount to produce 4-hydroxybutanal, the 4-hydroxybutanal pathway comprising alpha-ketoglutarate decarboxylase; 4-hydroxybutyrate dehydrogenase; and 4-hydroxybutyrate reductase (FIG. 58, steps B-C-D).
[0129] The invention further provides a non-naturally occurring microbial organism, comprising a 4-hydroxybutanal pathway comprising at least one exogenous nucleic acid encoding a 4-hydroxybutanal pathway enzyme expressed in a sufficient amount to produce 4-hydroxybutanal, the 4-hydroxybutanal pathway comprising succinate reductase; 4-hydroxybutyrate dehydrogenase, and 4-hydroxybutyrate reductase (see FIG. 62, steps F-C-D). In yet another embodiment, the invention provides a non-naturally occurring microbial organism, comprising a 4-hydroxybutanal pathway comprising at least one exogenous nucleic acid encoding a 4-hydroxybutanal pathway enzyme expressed in a sufficient amount to produce 4-hydroxybutanal, the 4-hydroxybutanal pathway comprising alpha-ketoglutarate decarboxylase, or glutamate dehydrogenase or glutamate transaminase and glutamate decarboxylase and 4-aminobutyrate dehydrogenase or 4-aminobutyrate transaminase; 4-hydroxybutyrate dehydrogenase; and 4-hydroxybutyrate reductase (see FIG. 62, steps B or ((J or K)-L-(M or N))-C-D).
[0130] The invention also provides a non-naturally occurring microbial organism, comprising a 4-hydroxybutanal pathway comprising at least one exogenous nucleic acid encoding a 4-hydroxybutanal pathway enzyme expressed in a sufficient amount to produce 4-hydroxybutanal, the 4-hydroxybutanal pathway comprising alpha-ketoglutarate reductase; 5-hydroxy-2-oxopentanoate dehydrogenase; and 5-hydroxy-2-oxopentanoate decarboxylase (see FIG. 62, steps X-Y-Z). In yet another embodiment, the invention provides a non-naturally occurring microbial organism, comprising a 4-hydroxybutyryl-CoA pathway comprising at least one exogenous nucleic acid encoding a 4-hydroxybutyryl-CoA pathway enzyme expressed in a sufficient amount to produce 4-hydroxybutyryl-CoA, the 4-hydroxybutyryl-CoA pathway comprising alpha-ketoglutarate reductase; 5-hydroxy-2-oxopentanoate dehydrogenase; and 5-hydroxy-2-oxopentanoate dehydrogenase (decarboxylation) (see FIG. 62, steps X-Y-AA).
[0131] The invention additionally provides a non-naturally occurring microbial organism, comprising a putrescine pathway comprising at least one exogenous nucleic acid encoding a putrescine pathway enzyme expressed in a sufficient amount to produce putrescine, the putrescine pathway comprising succinate reductase; 4-aminobutyrate dehydrogenase or 4-aminobutyrate transaminase; 4-aminobutyrate reductase; and putrescine dehydrogenase or putrescine transaminase (see FIG. 63, steps F-M/N-C-D/E). In still another embodiment, the invention provides a non-naturally occurring microbial organism, comprising a putrescine pathway comprising at least one exogenous nucleic acid encoding a putrescine pathway enzyme expressed in a sufficient amount to produce putrescine, the putrescine pathway comprising alpha-ketoglutarate decarboxylase; 4-aminobutyrate dehydrogenase or 4-aminobutyrate transaminase; 4-aminobutyrate reductase; and putrescine dehydrogenase or putrescine transaminase (see FIG. 63, steps B-M/N-C-D/E). The invention additionally provides a non-naturally occurring microbial organism, comprising a putrescine pathway comprising at least one exogenous nucleic acid encoding a putrescine pathway enzyme expressed in a sufficient amount to produce putrescine, the putrescine pathway comprising glutamate dehydrogenase or glutamate transaminase; glutamate decarboxylase; 4-aminobutyrate reductase; and putrescine dehydrogenase or putrescine transaminase (see FIG. 63, steps J/K-L-C-D/E).
[0132] The invention provides in another embodiment a non-naturally occurring microbial organism, comprising a putrescine pathway comprising at least one exogenous nucleic acid encoding a putrescine pathway enzyme expressed in a sufficient amount to produce putrescine, the putrescine pathway comprising alpha-ketoglutarate reductase; 5-amino-2-oxopentanoate dehydrogenase or 5-amino-2-oxopentanoate transaminase; 5-amino-2-oxopentanoate decarboxylase; and putrescine dehydrogenase or putrescine transaminase (see FIG. 63, steps 0-P/Q-R-D/E). Also provided is a non-naturally occurring microbial organism, comprising a putrescine pathway comprising at least one exogenous nucleic acid encoding a putrescine pathway enzyme expressed in a sufficient amount to produce putrescine, the putrescine pathway comprising alpha-ketoglutarate reductase; 5-amino-2-oxopentanoate dehydrogenase or 5-amino-2-oxopentanoate transaminase; ornithine dehydrogenase or ornithine transaminase; and ornithine decarboxylase (see FIG. 63, steps O-P/Q-S/T-U).
[0133] In an additional embodiment, the invention provides a non-naturally occurring microbial organism having a 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway, wherein the non-naturally occurring microbial organism comprises at least one exogenous nucleic acid encoding an enzyme or protein that converts a substrate of any of the pathways disclosed herein (see, for example, the Examples and FIGS. 1, 8-13, 58, 62 and 63). In an exmemplary embodiment for producing BDO, the microbial organism can convert a substrate to a product selected from the group consisting of succinate to succinyl-CoA; succinyl-CoA to succinic semialdehyde; succinic semialdehyde to 4-hydroxybutrate; 4-hydroxybutyrate to 4-hydroxybutyryl-phosphate; 4-hydroxybutyryl-phosphate to 4-hydroxtbutyryl-CoA; 4-hydroxybutyryl-CoA to 4-hydroxybutanal; and 4-hydroxybutanal to 1,4-butanediol. In a pathway for producing 4-HBal, a microbial organism can convert, for example, succinate to succinic semialdehyde; succinic semialdehyde to 4-hydroxybutyrate; and 4-hydroxybutyrate to 4-hydroxybutanal. Such an organism can additionally include activity to convert 4-hydroxybutanal to 1,4-butanediol in order to produce BDO. Yet another pathway for producing 4-HBal can be, for example, alpha-ketoglutarate to succinic semialdehyde; succinic semialdehyde to 4-hydroxybutyrate; and 4-hydroxybutyrate to 4-hydroxybutanal. An alternative pathway for producing 4-HBal can be, for example, alpha-ketoglutarate to 2,5-dioxopentanoic acid; 2,5-dioxopentanoic acid to 5-hydroxy-2-oxopentanooic acid; and 5-hydroxy-2-oxopentanoic acid to 4-hydroxybutanal. An exemplary 4-hydroxybutyryl-CoA pathway can be, for example, alpha-ketoglutarate to 2,5-dioxopentanoic acid; 2,5-dioxopentanoic acid to 5-hydroxy-2-oxopentanoic acid; and 5-hydroxy-2-oxopentanoic acid to 4-hydroxybutyryl-CoA. An exemplary putrescine pathway can be, for example, succinate to succinyl-CoA; succinyl-CoA to succinic semialdehyde; succinic semialdehyde to 4-aminobutyrate; 4-aminobutyrate to 4-aminobutanal; and 4-aminobutanal to putrescine. An alternative putrescine pathway can be, for example, succinate to succinic semialdehyde; succinic semialdehyde to 4-aminobutyrate; 4-aminobutyrate to 4-aminobutanal; and 4-aminobutanal to putrescine. One skilled in the art will understand that these are merely exemplary and that any of the substrate-product pairs disclosed herein suitable to produce a desired product and for which an appropriate activity is available for the conversion can be readily determined by one skilled in the art based on the teachings herein. Thus, the invention provides a non-naturally occurring microbial organism containing at least one exogenous nucleic acid encoding an enzyme or protein, where the enzyme or protein converts the substrates and products of a pathway (see FIGS. 1, 8-13, 58, 62 and 63).
[0134] While generally described herein as a microbial organism that contains a 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway, it is understood that the invention additionally provides a non-naturally occurring microbial organism comprising at least one exogenous nucleic acid encoding a 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway enzyme or protein expressed in a sufficient amount to produce an intermediate of a 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway. For example, as disclosed herein, 4-HB, 4-HBal, 4-HBCoA, BDO and putrescine pathways are exemplified in FIGS. 1, 8-13, 58, 62 and 63. Therefore, in addition to a microbial organism containing, for example, a BDO pathway that produces BDO, the invention additionally provides a non-naturally occurring microbial organism comprising at least one exogenous nucleic acid encoding a BDO pathway enzyme, where the microbial organism produces a BDO pathway intermediate as a product rather than an intermediate of the pathway. In one exemplary embodiment as shown in FIG. 62, for example, the invention provides a microbial organism that produces succinyl-CoA, succinic semialdehyde, 4-hydroxybutyrate, 4-hydroxybutyryl-phosphate, 4-hydroxybutyryl-CoA, or 4-hydroxybutanal as a product rather than an intermediate. Another exemplary embodiment includes, for example, a microbial organism that produces alpha-ketoglutarate, 2,5-dioxopentanoic acid, 5-hydroxy-2-oxopentanoic acid, or 4-hydroxybutanal as a product rather than an intermediate. An exemplary embodiment in a putrescine pathway includes, for example, a microbial organism that produces glutamate, 4-aminobutyrate, or 4-aminobutanal as a product rather than an intermediate. An alternative embodiment in a putrescine pathway can be, for example, a microbial organism that produces 2,5-dioxopentanoate, 5-amino-2-oxopentanoate, or ornithine as a product rather than an intermediate.
[0135] It is understood that any of the pathways disclosed herein, as described in the Examples and exemplified in the Figures, including the pathways of FIGS. 1, 8-13, 58, 62 and 63, can be utilized to generate a non-naturally occurring microbial organism that produces any pathway intermediate or product, as desired. As disclosed herein, such a microbial organism that produces an intermediate can be used in combination with another microbial organism expressing downstream pathway enzymes to produce a desired product. However, it is understood that a non-naturally occurring microbial organism that produces a 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway intermediate can be utilized to produce the intermediate as a desired product.
[0136] The invention is described herein with general reference to the metabolic reaction, reactant or product thereof, or with specific reference to one or more nucleic acids or genes encoding an enzyme associated with or catalyzing the referenced metabolic reaction, reactant or product. Unless otherwise expressly stated herein, those skilled in the art will understand that reference to a reaction also constitutes reference to the reactants and products of the reaction. Similarly, unless otherwise expressly stated herein, reference to a reactant or product also references the reaction and that reference to any of these metabolic constitutes also references the gene or genes encoding the enzymes that catalyze the referenced reaction, reactant or product. Likewise, given the well known fields of metabolic biochemistry, enzymology and genomics, reference herein to a gene or encoding nucleic acid also constitutes a reference to the corresponding encoded enzyme and the reaction it catalyzes as well as the reactants and products of the reaction.
[0137] The production of 4-HB via biosynthetic modes using the microbial organisms of the invention is particularly useful because it can produce monomeric 4-HB. The non-naturally occurring microbial organisms of the invention and their biosynthesis of 4-HB and BDO family compounds also is particularly useful because the 4-HB product can be (1) secreted; (2) can be devoid of any derivatizations such as Coenzyme A; (3) avoids thermodynamic changes during biosynthesis; (4) allows direct biosynthesis of BDO, and (5) allows for the spontaneous chemical conversion of 4-HB to .gamma.-butyrolactone (GBL) in acidic pH medium. This latter characteristic also is particularly useful for efficient chemical synthesis or biosynthesis of BDO family compounds such as 1,4-butanediol and/or tetrahydrofuran (THF), for example.
[0138] Microbial organisms generally lack the capacity to synthesize 4-HB and therefore any of the compounds disclosed herein to be within the 1,4-butanediol family of compounds or known by those in the art to be within the 1,4-butanediol family of compounds. Moreover, organisms having all of the requisite metabolic enzymatic capabilities are not known to produce 4-HB from the enzymes described and biochemical pathways exemplified herein. Rather, with the possible exception of a few anaerobic microorganisms described further below, the microorganisms having the enzymatic capability to use 4-HB as a substrate to produce, for example, succinate. In contrast, the non-naturally occurring microbial organisms of the invention can generate 4-HB, 4-HBal, 4-HBCoA, BDO and/or putrescine as a product. As described above, the biosynthesis of 4-HB in its monomeric form is not only particularly useful in chemical synthesis of BDO family of compounds, it also allows for the further biosynthesis of BDO family compounds and avoids altogether chemical synthesis procedures.
[0139] The non-naturally occurring microbial organisms of the invention that can produce 4-HB, 4-HBal, 4-HBCoA, BDO and/or putrescine are produced by ensuring that a host microbial organism includes functional capabilities for the complete biochemical synthesis of at least one 4-HB, 4-HBal, 4-HBCoA, BDO and/or putrscine biosynthetic pathway of the invention. Ensuring at least one requisite 4-HB, 4-HBal, 4-HBCoA or BDO biosynthetic pathway confers 4-HB biosynthesis capability onto the host microbial organism.
[0140] Several 4-HB biosynthetic pathways are exemplified herein and shown for purposes of illustration in FIG. 1. Additional 4-HB and BDO pathways are described in FIGS. 8-13. One 4-HB biosynthetic pathway includes the biosynthesis of 4-HB from succinate (the succinate pathway). The enzymes participating in this 4-HB pathway include CoA-independent succinic semialdehyde dehydrogenase and 4-hydroxybutanoate dehydrogenase. In this pathway, CoA-independent succinic semialdehyde dehydrogenase catalyzes the reverse reaction to the arrow shown in FIG. 1. Another 4-HB biosynthetic pathway includes the biosynthesis from succinate through succinyl-CoA (the succinyl-CoA pathway). The enzymes participating in this 4-HB pathway include succinyl-CoA synthetase, CoA-dependent succinic semialdehyde dehydrogenase and 4-hydroxybutanoate dehydrogenase. Three other 4-HB biosynthetic pathways include the biosynthesis of 4-HB from .alpha.-ketoglutarate (the .alpha.-ketoglutarate pathways). Hence, a third 4-HB biosynthetic pathway is the biosynthesis of succinic semialdehyde through glutamate: succinic semialdehyde transaminase, glutamate decarboxylase and 4-hydroxybutanoate dehydrogenase. A fourth 4-HB biosynthetic pathway also includes the biosynthesis of 4-HB from .alpha.-ketoglutarate, but utilizes .alpha.-ketoglutarate decarboxylase to catalyze succinic semialdehyde synthesis. 4-hydroxybutanoate dehydrogenase catalyzes the conversion of succinic semialdehyde to 4-HB. A fifth 4-HB biosynthetic pathway includes the biosynthesis from .alpha.-ketoglutarate through succinyl-CoA and utilizes .alpha.-ketoglutarate dehydrogenase to produce succinyl-CoA, which funnels into the succinyl-CoA pathway described above. Each of these 4-HB biosynthetic pathways, their substrates, reactants and products are described further below in the Examples. As described herein, 4-HB can further be biosynthetically converted to BDO by inclusion of appropriate enzymes to produce BDO (see Example). Thus, it is understood that a 4-HB pathway can be used with enzymes for converting 4-HB to BDO to generate a BDO pathway.
[0141] The non-naturally occurring microbial organisms of the invention can be produced by introducing expressible nucleic acids encoding one or more of the enzymes participating in one or more 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine biosynthetic pathways. Depending on the host microbial organism chosen for biosynthesis, nucleic acids for some or all of a particular 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine biosynthetic pathway can be expressed. For example, if a chosen host is deficient in one or more enzymes in a desired biosynthetic pathway, for example, the succinate to 4-HB pathway, then expressible nucleic acids for the deficient enzyme(s), for example, both CoA-independent succinic semialdehyde dehydrogenase and 4-hydroxybutanoate dehydrogenase in this example, are introduced into the host for subsequent exogenous expression. Alternatively, if the chosen host exhibits endogenous expression of some pathway enzymes, but is deficient in others, then an encoding nucleic acid is needed for the deficient enzyme(s) to achieve 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine biosynthesis. For example, if the chosen host exhibites endogenous CoA-independent succinic semialdehyde dehydrogenase, but is deficient in 4-hydroxybutanoate dehydrogenase, then an encoding nucleic acid is needed for this enzyme to achieve 4-HB biosynthesis. Thus, a non-naturally occurring microbial organism of the invention can be produced by introducing exogenous enzyme or protein activities to obtain a desired biosynthetic pathway or a desired biosynthetic pathway can be obtained by introducing one or more exogenous enzyme or protein activities that, together with one or more endogenous enzymes or proteins, produces a desired product such as 4-HB, 4-HBal, 4-HBCoA, BDO and/or putrescine.
[0142] In like fashion, where 4-HB biosynthesis is selected to occur through the succinate to succinyl-CoA pathway (the succinyl-CoA pathway), encoding nucleic acids for host deficiencies in the enzymes succinyl-CoA synthetase, CoA-dependent succinic semialdehyde dehydrogenase and/or 4-hydroxybutanoate dehydrogenase are to be exogenously expressed in the recipient host. Selection of 4-HB biosynthesis through the .alpha.-ketoglutarate to succinic semialdehyde pathway (the .alpha.-ketoglutarate pathway) can utilize exogenous expression for host deficiencies in one or more of the enzymes for glutamate:succinic semialdehyde transaminase, glutamate decarboxylase and/or 4-hydroxybutanoate dehydrogenase, or .alpha.-ketoglutarate decarboxylase and 4-hydroxybutanoate dehydrogenase. One skilled in the art can readily determine pathway enzymes for production of 4-HB or BDO, as disclosed herein.
[0143] Depending on the 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine biosynthetic pathway constituents of a selected host microbial organism, the non-naturally occurring microbial organisms of the invention will include at least one exogenously expressed 4-HB, 4-HB, 4-HBCoA, BDO or putrescine pathway-encoding nucleic acid and up to all encoding nucleic acids for one or more 4-HB or BDO biosynthetic pathways. For example, 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine biosynthesis can be established in a host deficient in a pathway enzyme or protein through exogenous expression of the corresponding encoding nucleic acid. In a host deficient in all enzymes or proteins of a 4-HB, 4-HB, 4-HBCoA, BDO or putrescine pathway, exogenous expression of all enzyme or proteins in the pathway can be included, although it is understood that all enzymes or proteins of a pathway can be expressed even if the host contains at least one of the pathway enzymes or proteins. For example, 4-HB biosynthesis can be established from all five pathways in a host deficient in 4-hydroxybutanoate dehydrogenase through exogenous expression of a 4-hydroxybutanoate dehydrogenase encoding nucleic acid. In contrast, 4-HB biosynthesis can be established from all five pathways in a host deficient in all eight enzymes through exogenous expression of all eight of CoA-independent succinic semialdehyde dehydrogenase, succinyl-CoA synthetase, CoA-dependent succinic semialdehyde dehydrogenase, glutamate: succinic semialdehyde transaminase, glutamate decarboxylase, .alpha.-ketoglutarate decarboxylase, .alpha.-ketoglutarate dehydrogenase and 4-hydroxybutanoate dehydrogenase.
[0144] Given the teachings and guidance provided herein, those skilled in the art will understand that the number of encoding nucleic acids to introduce in an expressible form will, at least, parallel the 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway deficiencies of the selected host microbial organism. Therefore, a non-naturally occurring microbial organism of the invention can have one, two, three, four, five, six, seven, eight or up to all nucleic acids encoding the enzymes disclosed herein constituting one or more 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine biosynthetic pathways. In some embodiments, the non-naturally occurring microbial organisms also can include other genetic modifications that facilitate or optimize 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine biosynthesis or that confer other useful functions onto the host microbial organism. One such other functionality can include, for example, augmentation of the synthesis of one or more of the 4-HB pathway precursors such as succinate, succinyl-CoA, .alpha.-ketoglutarate, 4-aminobutyrate, glutamate, acetoacetyl-CoA, and/or homoserine.
[0145] Generally, a host microbial organism is selected such that it produces the precursor of a 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway, either as a naturally produced molecule or as an engineered product that either provides de novo production of a desired precursor or increased production of a precursor naturally produced by the host microbial organism. For example, succinyl-CoA, .alpha.-ketoglutarate, 4-aminobutyrate, glutamate, acetoacetyl-CoA, and homoserine are produced naturally in a host organism such as E. coli. A host organism can be engineered to increase production of a precursor, as disclosed herein. In addition, a microbial organism that has been engineered to produce a desired precursor can be used as a host organism and further engineered to express enzymes or proteins of a 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway.
[0146] In some embodiments, a non-naturally occurring microbial organism of the invention is generated from a host that contains the enzymatic capability to synthesize 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine. In this specific embodiment it can be useful to increase the synthesis or accumulation of a 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway product to, for example, drive 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway reactions toward 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine production. Increased synthesis or accumulation can be accomplished by, for example, overexpression of nucleic acids encoding one or more of the 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway enzymes disclosed herein. Over expression of the 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway enzyme or enzymes can occur, for example, through exogenous expression of the endogenous gene or genes, or through exogenous expression of the heterologous gene or genes. Therefore, naturally occurring organisms can be readily generated to be non-naturally 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine producing microbial organisms of the invention through overexpression of one, two, three, four, five, six and so forth up to all nucleic acids encoding 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine biosynthetic pathway enzymes. In addition, a non-naturally occurring organism can be generated by mutagenesis of an endogenous gene that results in an increase in activity of an enzyme in the 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine biosynthetic pathway.
[0147] In particularly useful embodiments, exogenous expression of the encoding nucleic acids is employed. Exogenous expression confers the ability to custom tailor the expression and/or regulatory elements to the host and application to achieve a desired expression level that is controlled by the user. However, endogenous expression also can be utilized in other embodiments such as by removing a negative regulatory effector or induction of the gene's promoter when linked to an inducible promoter or other regulatory element. Thus, an endogenous gene having a naturally occurring inducible promoter can be up-regulated by providing the appropriate inducing agent, or the regulatory region of an endogenous gene can be engineered to incorporate an inducible regulatory element, thereby allowing the regulation of increased expression of an endogenous gene at a desired time. Similarly, an inducible promoter can be included as a regulatory element for an exogenous gene introduced into a non-naturally occurring microbial organism (see Examples).
[0148] "Exogenous" as it is used herein is intended to mean that the referenced molecule or the referenced activity is introduced into the host microbial organism. The molecule can be introduced, for example, by introduction of an encoding nucleic acid into the host genetic material such as by integration into a host chromosome or as non-chromosomal genetic material such as a plasmid. Therefore, the term as it is used in reference to expression of an encoding nucleic acid refers to introduction of the encoding nucleic acid in an expressible form into the microbial organism. When used in reference to a biosynthetic activity, the term refers to an activity that is introduced into the host reference organism. The source can be, for example, a homologous or heterologous encoding nucleic acid that expresses the referenced activity following introduction into the host microbial organism. Therefore, the term "endogenous" refers to a referenced molecule or activity that is present in the host. Similarly, the term when used in reference to expression of an encoding nucleic acid refers to expression of an encoding nucleic acid contained within the microbial organism. The term "heterologous" refers to a molecule or activity derived from a source other than the referenced species whereas "homologous" refers to a molecule or activity derived from the host microbial organism. Accordingly, exogenous expression of an encoding nucleic acid of the invention can utilize either or both a heterologous or homologous encoding nucleic acid.
[0149] It is understood that when more than one exogenous nucleic acid is included in a microbial organism that the more than one exogenous nucleic acids refers to the referenced encoding nucleic acid or biosynthetic activity, as discussed above. It is further understood, as disclosed herein, that such more than one exogenous nucleic acids can be introduced into the host microbial organism on separate nucleic acid molecules, on polycistronic nucleic acid molecules, or a combination thereof, and still be considered as more than one exogenous nucleic acid. For example, as disclosed herein a microbial organism can be engineered to express two or more exogenous nucleic acids encoding a desired pathway enzyme or protein. In the case where two exogenous nucleic acids encoding a desired activity are introduced into a host microbial organism, it is understood that the two exogenous nucleic acids can be introduced as a single nucleic acid, for example, on a single plasmid, on separate plasmids, can be integrated into the host chromosome at a single site or multiple sites, and still be considered as two exogenous nucleic acids. Similarly, it is understood that more than two exogenous nucleic acids can be introduced into a host organism in any desired combination, for example, on a single plasmid, on separate plasmids, can be integrated into the host chromosome at a single site or multiple sites, and still be considered as two or more exogenous nucleic acids, for example three exogenous nucleic acids. Thus, the number of referenced exogenous nucleic acids or biosynthetic activities refers to the number of encoding nucleic acids or the number of biosynthetic activities, not the number of separate nucleic acids introduced into the host organism.
[0150] Sources of encoding nucleic acids for a 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway enzyme can include, for example, any species where the encoded gene product is capable of catalyzing the referenced reaction. Such species include both prokaryotic and eukaryotic organisms including, but not limited to, bacteria, including archaea and eubacteria, and eukaryotes, including yeast, plant, insect, animal, and mammal, including human. Exemplary species for such sources include, for example, Escherichia coli, Saccharomyces cerevisiae, Saccharomyces kluyveri, Clostridium kluyveri, Clostridium acetobutylicum, Clostridium beijerinckii, Clostridium saccharoperbutylacetonicum, Clostridium perfringens, Clostridium difficile, Clostridium botulinum, Clostridium tyrobutyricum, Clostridium tetanomorphum, Clostridium tetani, Clostridium propionicum, Clostridium aminobutyricum, Clostridium subterminale, Clostridium sticklandii, Ralstonia eutropha, Mycobacterium bovis, Mycobacterium tuberculosis, Porphyromonas gingivalis, Arabidopsis thaliana, Thermus thermophilus, Pseudomonas species, including Pseudomonas aeruginosa, Pseudomonas putida, Pseudomonas stutzeri, Pseudomonas fluorescens, Homo sapiens, Oryctolagus cuniculus, Rhodobacter spaeroides, Thermoanaerobacter brockii, Metallosphaera sedula, Leuconostoc mesenteroides, Chloroflexus aurantiacus, Roseiflexus castenholzii, Erythrobacter, Simmondsia chinensis, Acinetobacter species, including Acinetobacter calcoaceticus and Acinetobacter baylyi, Porphyromonas gingivalis, Sulfolobus tokodaii, Sulfolobus solfataricus, Sulfolobus acidocaldarius, Bacillus subtilis, Bacillus cereus, Bacillus megaterium, Bacillus brevis, Bacillus pumilus, Rattus norvegicus, Klebsiella pneumonia, Klebsiella oxytoca, Euglena gracilis, Treponema denticola, Moorella thermoacetica, Thermotoga maritima, Halobacterium salinarum, Geobacillus stearothermophilus, Aeropyrum pernix, Sus scrofa, Caenorhabditis elegans, Corynebacterium glutamicum, Acidaminococcus fermentans, Lactococcus lactis, Lactobacillus plantarum, Streptococcus thermophilus, Enterobacter aerogenes, Candida, Aspergillus terreus, Pedicoccus pentosaceus, Zymomonas mobilus, Acetobacter pasteurians, Kluyveromyces lactis, Eubacterium barkeri, Bacteroides capillosus, Anaerotruncus colihominis, Natranaerobius thermophilusm, Campylobacter jejuni, Haemophilus influenzae, Serratia marcescens, Citrobacter amalonaticus, Myxococcus xanthus, Fusobacterium nuleatum, Penicillium chrysogenum, marine gamma proteobacterium, butyrate-producing bacterium, Nocardia iowensis, Nocardia farcinica, Streptomyces griseus, Schizosaccharomyces pombe, Geobacillus thermoglucosidasius, Salmonella typhimurium, Vibrio cholera, Heliobacter pylori, Nicotiana tabacum, Oryza sativa, Haloferax mediterranei, Agrobacterium tumefaciens, Achromobacter denitrificans, Fusobacterium nucleatum, Streptomyces clavuligenus, Acinetobacter baumanii, Mus musculus, Lachancea kluyveri, Trichomonas vaginalis, Trypanosoma brucei, Pseudomonas stutzeri, Bradyrhizobium japonicum, Mesorhizobium loti, Bos taurus, Nicotiana glutinosa, Vibrio vulnificus, Selenomonas ruminantium, Vibrio parahaemolyticus, Archaeoglobus fulgidus, Haloarcula marismortui, Pyrobaculum aerophilum, Mycobacterium smegmatis MC2 155, Mycobacterium avium subsp. paratuberculosis K-10, Mycobacterium marinum M, Tsukamurella paurometabola DSM 20162, Cyanobium PCC7001, Dictyostelium discoideum AX4, and others disclosed herein (see Examples). For example, microbial organisms having 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine biosynthetic production are exemplified herein with reference to E. coli and yeast hosts. However, with the complete genome sequence available for now more than 550 species (with more than half of these available on public databases such as the NCBI), including 395 microorganism genomes and a variety of yeast, fungi, plant, and mammalian genomes, the identification of genes encoding the requisite 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine biosynthetic activity for one or more genes in related or distant species, including for example, homologues, orthologs, paralogs and nonorthologous gene displacements of known genes, and the interchange of genetic alterations between organisms is routine and well known in the art. Accordingly, the metabolic alterations allowing biosynthesis of 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine and other compounds of the invention described herein with reference to a particular organism such as E. coli or yeast can be readily applied to other microorganisms, including prokaryotic and eukaryotic organisms alike. Given the teachings and guidance provided herein, those skilled in the art will know that a metabolic alteration exemplified in one organism can be applied equally to other organisms.
[0151] In some instances, such as when an alternative 4-HB, 4-HBal, BDO or putrescine biosynthetic pathway exists in an unrelated species, 4-HB, 4-HBal, BDO or putrescine biosynthesis can be conferred onto the host species by, for example, exogenous expression of a paralog or paralogs from the unrelated species that catalyzes a similar, yet non-identical metabolic reaction to replace the referenced reaction. Because certain differences among metabolic networks exist between different organisms, those skilled in the art will understand that the actual gene usage between different organisms may differ. However, given the teachings and guidance provided herein, those skilled in the art also will understand that the teachings and methods of the invention can be applied to all microbial organisms using the cognate metabolic alterations to those exemplified herein to construct a microbial organism in a species of interest that will synthesize 4-HB, such as monomeric 4-HB, 4-HBal, BDO or putrescine.
[0152] Host microbial organisms can be selected from, and the non-naturally occurring microbial organisms generated in, for example, bacteria, yeast, fungus or any of a variety of other microorganisms applicable to fermentation processes. Exemplary bacteria include species selected from Escherichia coli, Klebsiella oxytoca, Anaerobiospirillum succiniciproducens, Actinobacillus succinogenes, Mannheimia succiniciproducens, Rhizobium etli, Bacillus subtilis, Corynebacterium glutamicum, Gluconobacter oxydans, Zymomonas mobilis, Lactococcus lactis, Lactobacillus plantarum, Streptomyces coelicolor, Clostridium acetobutylicum, Pseudomonas fluorescens, and Pseudomonas putida. Exemplary yeasts or fungi include species selected from Saccharomyces cerevisiae, Schizosaccharomyces pombe, Kluyveromyces lactis, Kluyveromyces marxianus, Aspergillus terreus, Aspergillus niger, Pichia pastoris, Rhizopus arrhizus, Rhizopus oryzae, and the like. E. coli is a particularly useful host organisms since it is a well characterized microbial organism suitable for genetic engineering. Other particularly useful host organisms include yeast such as Saccharomyces cerevisiae. It is understood that any suitable microbial host organism can be used to introduce metabolic and/or genetic modifications to produce a desired product.
[0153] Methods for constructing and testing the expression levels of a non-naturally occurring 4-HB-, 4-HBal-, 4-HBCoA-, BDO-, or putrescine-producing host can be performed, for example, by recombinant and detection methods well known in the art. Such methods can be found described in, for example, Sambrook et al., Molecular Cloning: A Laboratory Manual, Third Ed., Cold Spring Harbor Laboratory, New York (2001); Ausubel et al., Current Protocols in Molecular Biology, John Wiley and Sons, Baltimore, Md. (1999). 4-HB and GBL can be separated by, for example, HPLC using a Spherisorb 5 ODS1 column and a mobile phase of 70% 10 mM phosphate buffer (pH=7) and 30% methanol, and detected using a UV detector at 215 nm (Hennessy et al. 2004, J. Forensic Sci. 46(6):1-9). BDO is detected by gas chromatography or by HPLC and refractive index detector using an Aminex HPX-87H column and a mobile phase of 0.5 mM sulfuric acid (Gonzalez-Pajuelo et al., Met. Eng. 7:329-336 (2005)).
[0154] Exogenous nucleic acid sequences involved in a pathway for production of 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine can be introduced stably or transiently into a host cell using techniques well known in the art including, but not limited to, conjugation, electroporation, chemical transformation, transduction, transfection, and ultrasound transformation. For exogenous expression in E. coli or other prokaryotic cells, some nucleic acid sequences in the genes or cDNAs of eukaryotic nucleic acids can encode targeting signals such as an N-terminal mitochondrial or other targeting signal, which can be removed before transformation into prokaryotic host cells, if desired. For example, removal of a mitochondrial leader sequence led to increased expression in E. coli (Hoffmeister et al., J. Biol. Chem. 280:4329-4338 (2005)). For exogenous expression in yeast or other eukaryotic cells, genes can be expressed in the cytosol without the addition of leader sequence, or can be targeted to mitochondrion or other organelles, or targeted for secretion, by the addition of a suitable targeting sequence such as a mitochondrial targeting or secretion signal suitable for the host cells. Thus, it is understood that appropriate modifications to a nucleic acid sequence to remove or include a targeting sequence can be incorporated into an exogenous nucleic acid sequence to impart desirable properties. Furthermore, genes can be subjected to codon optimization with techniques well known in the art to achieve optimized expression of the proteins.
[0155] An expression vector or vectors can be constructed to harbor one or more 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine biosynthetic pathway and/or one or more biosynthetic encoding nucleic acids as exemplified herein operably linked to expression control sequences functional in the host organism. Expression vectors applicable for use in the microbial host organisms of the invention include, for example, plasmids, phage vectors, viral vectors, episomes and artificial chromosomes, including vectors and selection sequences or markers operable for stable integration into a host chromosome. Additionally, the expression vectors can include one or more selectable marker genes and appropriate expression control sequences. Selectable marker genes also can be included that, for example, provide resistance to antibiotics or toxins, complement auxotrophic deficiencies, or supply critical nutrients not in the culture media. Expression control sequences can include constitutive and inducible promoters, transcription enhancers, transcription terminators, and the like which are well known in the art. When two or more exogenous encoding nucleic acids are to be co-expressed, both nucleic acids can be inserted, for example, into a single expression vector or in separate expression vectors. For single vector expression, the encoding nucleic acids can be operationally linked to one common expression control sequence or linked to different expression control sequences, such as one inducible promoter and one constitutive promoter. The transformation of exogenous nucleic acid sequences involved in a metabolic or synthetic pathway can be confirmed using methods well known in the art. Such methods include, for example, nucleic acid analysis such as Northern blots or polymerase chain reaction (PCR) amplification of mRNA, or immunoblotting for expression of gene products, or other suitable analytical methods to test the expression of an introduced nucleic acid sequence or its corresponding gene product. It is understood by those skilled in the art that the exogenous nucleic acid is expressed in a sufficient amount to produce the desired product, and it is further understood that expression levels can be optimized to obtain sufficient expression using methods well known in the art and as disclosed herein.
[0156] The non-naturally occurring microbial organisms of the invention are constructed using methods well known in the art as exemplified herein to exogenously express at least one nucleic acid encoding a 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway enzyme in sufficient amounts to produce 4-HB, such as monomeric 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine. It is understood that the microbial organisms of the invention are cultured under conditions sufficient to produce 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine. Exemplary levels of expression for 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine enzymes in each pathway are described further below in the Examples. Following the teachings and guidance provided herein, the non-naturally occurring microbial organisms of the invention can achieve biosynthesis of 4-HB, such as monomeric 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine resulting in intracellular concentrations between about 0.1-200 mM or more, for example, 0.1-25 mM or more. Generally, the intracellular concentration of 4-HB, such as monomeric 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine is between about 3-150 mM or more, particularly about 5-125 mM or more, and more particularly between about 8-100 mM, for example, about 3-20 mM, particularly between about 5-15 mM and more particularly between about 8-12 mM, including about 10 mM, 20 mM, 50 MM, 80 mM or more. Intracellular concentrations between and above each of these exemplary ranges also can be achieved from the non-naturally occurring microbial organisms of the invention. In particular embodiments, the microbial organisms of the invention, particularly strains such as those disclosed herein (see Examples XII-XIX and Table 28), can provide improved production of a desired product such as 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine by increasing the production of 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine and/or decreasing undesirable byproducts. Such production levels include, but are not limited to, those disclosed herein and including from about 1 gram to about 25 grams per liter, for example about 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, or even higher amounts of product per liter.
[0157] In addition to the culturing and fermentation conditions disclosed herein, growth condition for achieving biosynthesis of BDO, 4-HB, 4-HBCoA, 4-HBal and/or putrescine can include the addition of an osmoprotectant to the culturing conditions. In certain embodiments, the non-naturally occurring microbial organisms of the invention can be sustained, cultured or fermented as described herein in the presence of an osmoprotectant. Briefly, an osmoprotectant refers to a compound that acts as an osmolyte and helps a microbial organism as described herein survive osmotic stress. Osmoprotectants include, but are not limited to, betaines, amino acids, and the sugar trehalose. Non-limiting examples of such are glycine betaine, praline betaine, dimethylthetin, dimethyl slfonioproprionate, 3-dimethylsulfonio-2-methylproprionate, pipecolic acid, dimethylsulfonioacetate, choline, L-carnitine and ectoine. In one aspect, the osmoprotectant is glycine betaine. It is understood to one of ordinary skill in the art that the amount and type of osmoprotectant suitable for protecting a microbial organism described herein from osmotic stress will depend on the microbial organism used. The amount of osmoprotectant in the culturing conditions can be, for example, no more than about 0.1 mM, no more than about 0.5 mM, no more than about 1.0 mM, no more than about 1.5 mM, no more than about 2.0 mM, no more than about 2.5 mM, no more than about 3.0 mM, no more than about 5.0 mM, no more than about 7.0 mM, no more than about 10 mM, no more than about 50 mM, no more than about 100 mM or no more than about 500 mM.
[0158] In some embodiments, culture conditions include anaerobic or substantially anaerobic growth or maintenance conditions. Exemplary anaerobic conditions have been described previously and are well known in the art. Exemplary anaerobic conditions for fermentation processes are described herein and are described, for example, in U.S. publication 2009/0047719, filed Aug. 10, 2007. Any of these conditions can be employed with the non-naturally occurring microbial organisms as well as other anaerobic conditions well known in the art. Under such anaerobic conditions, the 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine producers can synthesize 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine at intracellular concentrations of 5-10 mM or more as well as all other concentrations exemplified herein. It is understood that, even though the above description refers to intracellular concentrations, 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine producing microbial organisms can produce 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine intracellularly and/or secrete the product into the culture medium.
[0159] The culture conditions can include, for example, liquid culture procedures as well as fermentation and other large scale culture procedures. As described herein, particularly useful yields of the biosynthetic products of the invention can be obtained under anaerobic or substantially anaerobic culture conditions.
[0160] As described herein, one exemplary growth condition for achieving biosynthesis of 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine includes anaerobic culture or fermentation conditions. In certain embodiments, the non-naturally occurring microbial organisms of the invention can be sustained, cultured or fermented under anaerobic or substantially anaerobic conditions. Briefly, anaerobic conditions refers to an environment devoid of oxygen. Substantially anaerobic conditions include, for example, a culture, batch fermentation or continuous fermentation such that the dissolved oxygen concentration in the medium remains between 0 and 10% of saturation. Substantially anaerobic conditions also includes growing or resting cells in liquid medium or on solid agar inside a sealed chamber maintained with an atmosphere of less than 1% oxygen. The percent of oxygen can be maintained by, for example, sparging the culture with an N.sub.2/CO.sub.2 mixture or other suitable non-oxygen gas or gases.
[0161] The invention also provides a non-naturally occurring microbial biocatalyst including a microbial organism having 4-hydroxybutanoic acid (4-HB) and 1,4-butanediol (BDO) biosynthetic pathways that include at least one exogenous nucleic acid encoding 4-hydroxybutanoate dehydrogenase, CoA-independent succinic semialdehyde dehydrogenase, succinyl-CoA synthetase, CoA-dependent succinic semialdehyde dehydrogenase, 4-hydroxybutyrate:CoA transferase, glutamate: succinic semialdehyde transaminase, glutamate decarboxylase, CoA-independent aldehyde dehydrogenase, CoA-dependent aldehyde dehydrogenase or alcohol dehydrogenase, wherein the exogenous nucleic acid is expressed in sufficient amounts to produce 1,4-butanediol (BDO). 4-Hydroxybutyrate:CoA transferase also is known as 4-hydroxybutyryl CoA:acetyl-CoA transferase. Additional 4-HB or BDO pathway enzymes are also disclosed herein (see Examples and FIGS. 8-13).
[0162] The invention further provides non-naturally occurring microbial biocatalyst including a microbial organism having 4-hydroxybutanoic acid (4-HB) and 1,4-butanediol (BDO) biosynthetic pathways, the pathways include at least one exogenous nucleic acid encoding 4-hydroxybutanoate dehydrogenase, succinyl-CoA synthetase, CoA-dependent succinic semialdehyde dehydrogenase, 4-hydroxybutyrate:CoA transferase, 4-butyrate kinase, phosphotransbutyrylase, .alpha.-ketoglutarate decarboxylase, aldehyde dehydrogenase, alcohol dehydrogenase or an aldehyde/alcohol dehydrogenase, wherein the exogenous nucleic acid is expressed in sufficient amounts to produce 1,4-butanediol (BDO).
[0163] Non-naturally occurring microbial organisms also can be generated which biosynthesize BDO. As with the 4-HB producing microbial organisms of the invention, the BDO producing microbial organisms also can produce intracellularly or secret the BDO into the culture medium. Following the teachings and guidance provided previously for the construction of microbial organisms that synthesize 4-HB, additional BDO pathways can be incorporated into the 4-HB producing microbial organisms to generate organisms that also synthesize BDO and other BDO family compounds. The chemical synthesis of BDO and its downstream products are known. The non-naturally occurring microbial organisms of the invention capable of BDO biosynthesis circumvent these chemical synthesis using 4-HB as an entry point as illustrated in FIG. 1. As described further below, the 4-HB producers also can be used to chemically convert 4-HB to GBL and then to BDO or THF, for example. Alternatively, the 4-HB producers can be further modified to include biosynthetic capabilities for conversion of 4-HB and/or GBL to BDO.
[0164] The additional BDO pathways to introduce into 4-HB producers include, for example, the exogenous expression in a host deficient background or the overexpression of one or more of the enzymes exemplified in FIG. 1 as steps 9-13. One such pathway includes, for example, the enzyme activies necessary to carryout the reactions shown as steps 9, 12 and 13 in FIG. 1, where the aldehyde and alcohol dehydrogenases can be separate enzymes or a multifunctional enzyme having both aldehyde and alcohol dehydrogenase activity. Another such pathway includes, for example, the enzyme activities necessary to carry out the reactions shown as steps 10, 11, 12 and 13 in FIG. 1, also where the aldehyde and alcohol dehydrogenases can be separate enzymes or a multifunctional enzyme having both aldehyde and alcohol dehydrogenase activity. Accordingly, the additional BDO pathways to introduce into 4-HB producers include, for example, the exogenous expression in a host deficient background or the overexpression of one or more of a 4-hydroxybutyrate:CoA transferase, butyrate kinase, phosphotransbutyrylase, CoA-independent aldehyde dehydrogenase, CoA-dependent aldehyde dehydrogenase or an alcohol dehydrogenase. In the absence of endogenous acyl-CoA synthetase capable of modifying 4-HB, the non-naturally occurring BDO producing microbial organisms can further include an exogenous acyl-CoA synthetase selective for 4-HB, or the combination of multiple enzymes that have as a net reaction conversion of 4-HB into 4-HB-CoA. As exemplified further below in the Examples, butyrate kinase and phosphotransbutyrylase exhibit BDO pathway activity and catalyze the conversions illustrated in FIG. 1 with a 4-HB substrate. Therefore, these enzymes also can be referred to herein as 4-hydroxybutyrate kinase and phosphotranshydroxybutyrylase respectively.
[0165] Exemplary alcohol and aldehyde dehydrogenases that can be used for these in vivo conversions from 4-HB to BDO are listed below in Table 1.
TABLE-US-00001 TABLE 1 Alcohol and Aldehyde Dehydrogenases for Conversion of 4-HB to BDO. ALCOHOL DEHYDROGENASES ec: 1.1.1.1 alcohol dehydrogenase ec: 1.1.1.2 alcohol dehydrogenase (NADP+) ec: 1.1.1.4 (R,R)-butanediol dehydrogenase ec: 1.1.1.5 acetoin dehydrogenase ec: 1.1.1.6 glycerol dehydrogenase ec: 1.1.1.7 propanediol-phosphate dehydrogenase ec: 1.1.1.8 glycerol-3-phosphate dehydrogenase (NAD+) ec: 1.1.1.11 D-arabinitol 4-dehydrogenase ec: 1.1.1.12 L-arabinitol 4-dehydrogenase ec: 1.1.1.13 L-arabinitol 2-dehydrogenase ec: 1.1.1.14 L-iditol 2-dehydrogenase ec: 1.1.1.15 D-iditol 2-dehydrogenase ec: 1.1.1.16 galactitol 2-dehydrogenase ec: 1.1.1.17 mannitol-l-phosphate 5- dehydrogenase ec: 1.1.1.18 inositol 2-dehydrogenase ec: 1.1.1.21 aldehyde reductase ec: 1.1.1.23 histidinol dehydrogenase ec: 1.1.1.26 glyoxylate reductase ec: 1.1.1.27 L-lactate dehydrogenase ec: 1.1.1.28 D-lactate dehydrogenase ec: 1.1.1.29 glycerate dehydrogenase ec: 1.1.1.30 3-hydroxybutyrate dehydrogenase ec: 1.1.1.31 3-hydroxyisobutyrate dehydrogenase ec: 1.1.1.35 3-hydroxyacyl-CoA dehydrogenase ec: 1.1.1.36 acetoacetyl-CoA reductase ec: 1.1.1.37 malate dehydrogenase ec: 1.1.1.38 malate dehydrogenase (oxaloacetate- decarboxylating) ec: 1.1.1.39 malate dehydrogenase (decarboxylating) ec: 1.1.1.40 malate dehydrogenase (oxaloacetate- decarboxylating) (NADP+) ec: 1.1.1.41 isocitrate dehydrogenase (NAD+) ec: 1.1.1.42 isocitrate dehydrogenase (NADP+) ec: 1.1.1.54 allyl-alcohol dehydrogenase ec: 1.1.1.55 lactaldehyde reductase (NADPH) ec: 1.1.1.56 ribitol 2-dehydrogenase ec: 1.1.1.59 3-hydroxypropionate dehydrogenase ec: 1.1.1.60 2-hydroxy-3-oxopropionate reductase ec: 1.1.1.61 4-hydroxybutyrate dehydrogenase ec: 1.1.1.66 omega-hydroxydecanoate dehydrogenase ec: 1.1.1.67 mannitol 2-dehydrogenase ec: 1.1.1.71 alcohol dehydrogenase [NAD(P)+] ec: 1.1.1.72 glycerol dehydrogenase (NADP+) ec: 1.1.1.73 octanol dehydrogenase ec: 1.1.1.75 (R)-aminopropanol dehydrogenase ec: 1.1.1.76 (S,S)-butanediol dehydrogenase ec: 1.1.1.77 lactaldehyde reductase ec: 1.1.1.78 methylglyoxal reductase (NADH- dependent) ec: 1.1.1.79 glyoxylate reductase (NADP+) ec: 1.1.1.80 isopropanol dehydrogenase (NADP+) ec: 1.1.1.81 hydroxypyruvate reductase ec: 1.1.1.82 malate dehydrogenase (NADP+) ec: 1.1.1.83 D-malate dehydrogenase (decarboxylating) ec: 1.1.1.84 dimethylmalate dehydrogenase ec: 1.1.1.85 3-isopropylmalate dehydrogenase ec: 1.1.1.86 ketol-acid reductoisomerase ec: 1.1.1.87 homoisocitrate dehydrogenase ec: 1.1.1.88 hydroxymethylglutaryl-CoA reductase ec: 1.1.1.90 aryl-alcohol dehydrogenase ec: 1.1.1.91 aryl-alcohol dehydrogenase (NADP+) ec: 1.1.1.92 oxaloglycolate reductase (decarboxylating) ec: 1.1.1.94 glycerol-3-phosphate dehydrogenase [NAD(P)+] ec: 1.1.1.95 phosphoglycerate dehydrogenase ec: 1.1.1.97 3-hydroxybenzyl-alcohol dehydrogenase ec: 1.1.1.101 acylglycerone-phosphate reductase ec: 1.1.1.103 L-threonine 3-dehydrogenase ec: 1.1.1.104 4-oxoproline reductase ec: 1.1.1.105 retinol dehydrogenase ec: 1.1.1.110 indolelactate dehydrogenase ec: 1.1.1.112 indanol dehydrogenase ec: 1.1.1.113 L-xylose 1-dehydrogenase ec: 1.1.1.129 L-threonate 3-dehydrogenase ec: 1.1.1.137 ribitol-5-phosphate 2-dehydrogenase ec: 1.1.1.138 mannitol 2-dehydrogenase (NADP+) ec: 1.1.1.140 sorbitol-6-phosphate 2- dehydrogenase ec: 1.1.1.142 D-pinitol dehydrogenase ec: 1.1.1.143 sequoyitol dehydrogenase ec: 1.1.1.144 perillyl-alcohol dehydrogenase ec: 1.1.1.156 glycerol 2-dehydrogenase (NADP+) ec: 1.1.1.157 3-hydroxybutyryl-CoA dehydrogenase ec: 1.1.1.163 cyclopentanol dehydrogenase ec: 1.1.1.164 hexadecanol dehydrogenase ec: 1.1.1.165 2-alkyn-1-ol dehydrogenase ec: 1.1.1.166 hydroxycyclohexanecarboxylate dehydrogenase ec: 1.1.1.167 hydroxymalonate dehydrogenase ec: 1.1.1.174 cyclohexane-1,2-diol dehydrogenase ec: 1.1.1.177 glycerol-3-phosphate 1- dehydrogenase (NADP+) ec: 1.1.1.178 3-hydroxy-2-methylbutyryl-CoA dehydrogenase ec: 1.1.1.185 L-glycol dehydrogenase ec: 1.1.1.190 indole-3-acetaldehyde reductase (NADH) ec: 1.1.1.191 indole-3-acetaldehyde reductase (NADPH) ec: 1.1.1.192 long-chain-alcohol dehydrogenase ec: 1.1.1.194 coniferyl-alcohol dehydrogenase ec: 1.1.1.195 cinnamyl-alcohol dehydrogenase ec: 1.1.1.198 (+)-borneol dehydrogenase ec: 1.1.1.202 1,3-propanediol dehydrogenase ec: 1.1.1.207 (-)-menthol dehydrogenase ec: 1.1.1.208 (+)-neomenthol dehydrogenase ec: 1.1.1.216 farnesol dehydrogenase ec: 1.1.1.217 benzyl-2-methyl-hydroxybutyrate dehydrogenase ec: 1.1.1.222 (R)-4-hydroxyphenyllactate dehydrogenase ec: 1.1.1.223 isopiperitenol dehydrogenase ec: 1.1.1.226 4-hydroxycyclohexanecarboxylate dehydrogenase ec: 1.1.1.229 diethyl 2-methyl-3-oxosuccinate reductase ec: 1.1.1.237 hydroxyphenylpyruvate reductase ec: 1.1.1.244 methanol dehydrogenase ec: 1.1.1.245 cyclohexanol dehydrogenase ec: 1.1.1.250 D-arabinitol 2-dehydrogenase ec: 1.1.1.251 galactitol 1-phosphate 5- dehydrogenase ec: 1.1.1.255 mannitol dehydrogenase ec: 1.1.1.256 fluoren-9-ol dehydrogenase ec: 1.1.1.257 4-(hydroxymethyl)benzenesulfonate dehydrogenase ec: 1.1.1.258 6-hydroxyhexanoate dehydrogenase ec: 1.1.1.259 3-hydroxypimeloyl-CoA dehydrogenase ec: 1.1.1.261 glycerol-1-phosphate dehydrogenase [NAD(P)+] ec: 1.1.1.265 3-methylbutanal reductase ec: 1.1.1.283 methylglyoxal reductase (NADPH- dependent) ec: 1.1.1.286 isocitrate-homoisocitrate dehydrogenase ec: 1.1.1.287 D-arabinitol dehydrogenase (NADP+) butanol dehydrogenase ALDEHYDE DEHYDROGENASES ec: 1.2.1.2 formate dehydrogenase ec: 1.2.1.3 aldehyde dehydrogenase (NAD+) ec: 1.2.1.4 aldehyde dehydrogenase (NADP+) ec: 1.2.1.5 aldehyde dehydrogenase [NAD(P)+] ec: 1.2.1.7 benzaldehyde dehydrogenase (NADP+) ec: 1.2.1.8 betaine-aldehyde dehydrogenase ec: 1.2.1.9 glyceraldehyde-3-phosphate dehydrogenase (NADP+) ec: 1.2.1.10 acetaldehyde dehydrogenase (acetylating) ec: 1.2.1.11 aspartate-semialdehyde dehydrogenase ec: 1.2.1.12 glyceraldehyde-3-phosphate dehydrogenase (phosphorylating) ec: 1.2.1.13 glyceraldehyde-3-phosphate dehydrogenase (NADP+) (phosphorylating) ec: 1.2.1.15 malonate-semialdehyde dehydrogenase ec: 1.2.1.16 succinate-semialdehyde dehydrogenase [NAD(P)+] ec: 1.2.1.17 glyoxylate dehydrogenase (acylating) ec: 1.2.1.18 malonate-semialdehyde dehydrogenase (acetylating) ec: 1.2.1.19 aminobutyraldehyde dehydrogenase ec: 1.2.1.20 glutarate-semialdehyde dehydrogenase ec: 1.2.1.21 glycolaldehyde dehydrogenase ec: 1.2.1.22 lactaldehyde dehydrogenase ec: 1.2.1.23 2-oxoaldehyde dehydrogenase (NAD+) ec: 1.2.1.24 succinate-semialdehyde dehydrogenase ec: 1.2.1.25 2-oxoisovalerate dehydrogenase (acylating) ec: 1.2.1.26 2,5-dioxovalerate dehydrogenase ec: 1.2.1.27 methylmalonate-semialdehyde dehydrogenase (acylating) ec: 1.2.1.28 benzaldehyde dehydrogenase (NAD+) ec: 1.2.1.29 aryl-aldehyde dehydrogenase ec: 1.2.1.30 aryl-aldehyde dehydrogenase (NADP+) ec: 1.2.1.31 L-aminoadipate-semialdehyde dehydrogenase ec: 1.2.1.32 aminomuconate-semialdehyde dehydrogenase ec: 1.2.1.36 retinal dehydrogenase ec: 1.2.1.39 phenylacetaldehyde dehydrogenase ec: 1.2.1.41 glutamate-5-semialdehyde dehydrogenase ec: 1.2.1.42 hexadecanal dehydrogenase (acylating) ec: 1.2.1.43 formate dehydrogenase (NADP+) ec: 1.2.1.45 4-carboxy-2-hydroxymuconate-6- semialdehyde dehydrogenase ec: 1.2.1.46 formaldehyde dehydrogenase ec: 1.2.1.47 4-trimethylammoniobutyraldehyde dehydrogenase ec: 1.2.1.48 long-chain-aldehyde dehydrogenase ec: 1.2.1.49 2-oxoaldehyde dehydrogenase (NADP+) ec: 1.2.1.51 pyruvate dehydrogenase (NADP+) ec: 1.2.1.52 oxoglutarate dehydrogenase (NADP+) ec: 1.2.1.53 4-hydroxyphenylacetaldehyde dehydrogenase ec: 1.2.1.57 butanal dehydrogenase ec: 1.2.1.58 phenylglyoxylate dehydrogenase (acylating) ec: 1.2.1.59 glyceraldehyde-3-phosphate dehydrogenase (NAD(P)+) (phosphorylating) ec: 1.2.1.62 4-formylbenzenesulfonate dehydrogenase ec: 1.2.1.63 6-oxohexanoate dehydrogenase ec: 1.2.1.64 4-hydroxybenzaldehyde dehydrogenase ec: 1.2.1.65 salicylaldehyde dehydrogenase ec: 1.2.1.66 mycothiol-dependent formaldehyde dehydrogenase ec: 1.2.1.67 vanillin dehydrogenase ec: 1.2.1.68 coniferyl-aldehyde dehydrogenase ec: 1.2.1.69 fluoroacetaldehyde dehydrogenase ec: 1.2.1.71 succinylglutamate-semialdehyde dehydrogenase
[0166] Other exemplary enzymes and pathways are disclosed herein (see Examples). Furthermore, it is understood that enzymes can be utilized for carry out reactions for which the substrate is not the natural substrate. While the activity for the non-natural substrate may be lower than the natural substrate, it is understood that such enzymes can be utilized, either as naturally occurring or modified using the directed evolution or adaptive evolution, as disclosed herein (see also Examples).
[0167] BDO production through any of the pathways disclosed herein are based, in part, on the identification of the appropriate enzymes for conversion of precursors to BDO. A number of specific enzymes for several of the reaction steps have been identified. For those transformations where enzymes specific to the reaction precursors have not been identified, enzyme candidates have been identified that are best suited for catalyzing the reaction steps. Enzymes have been shown to operate on a broad range of substrates, as discussed below. In addition, advances in the field of protein engineering also make it feasible to alter enzymes to act efficiently on substrates, even if not a natural substrate. Described below are several examples of broad-specificity enzymes from diverse classes suitable for a BDO pathway as well as methods that have been used for evolving enzymes to act on non-natural substrates.
[0168] A key class of enzymes in BDO pathways is the oxidoreductases that interconvert ketones or aldehydes to alcohols (1.1.1). Numerous exemplary enzymes in this class can operate on a wide range of substrates. An alcohol dehydrogenase (1.1.1.1) purified from the soil bacterium Brevibacterium sp KU 1309 (Hirano et al., J. Biosc. Bioeng. 100:318-322 (2005)) was shown to operate on a plethora of aliphatic as well as aromatic alcohols with high activities. Table 2 shows the activity of the enzyme and its K.sub.m on different alcohols. The enzyme is reversible and has very high activity on several aldehydes also (Table 3).
TABLE-US-00002 TABLE 2 Relative activities of an alcohol dehydrogenase from Brevibacterium sp KU to oxidize various alcohols. Relative Activity K.sub.m Substrate (0%) (mM) 2-Phenylethanol 100* 0.025 (S)-2-Phenylpropanol 156 0.157 (R)-2-Phenylpropanol 63 0.020 Bynzyl alcohol 199 0.012 3-Phenylpropanol 135 0.033 Ethanol 76 1-Butanol 111 1-Octanol 101 1-Dodecanol 68 1-Phenylethanol 46 2-Propanol 54 *The activity of 2-phenylethanol, corresponding to 19.2 U/mg, was taken as 100%.
TABLE-US-00003 TABLE 3 Relative activities of an alcohol dehydrogenase from Brevibacterium sp KU 1309 to reduce various carbonyl compounds. Relative Activity K.sub.m Substrate (%) (mM) Phenylacetaldehyde 100 0.261 2-Phenylpropionaldehyde 188 0.864 1-Octylaldehyde 87 Acetophenone 0
[0169] Lactate dehydrogenase (1.1.1.27) from Ralstonia eutropha is another enzyme that has been demonstrated to have high activities on several 2-oxoacids such as 2-oxobutyrate, 2-oxopentanoate and 2-oxoglutarate (a C5 compound analogous to 2-oxoadipate) (Steinbuchel and Schlegel, Eur. J. Biochem. 130:329-334 (1983)). Column 2 in Table 4 demonstrates the activities of ldhA from R. eutropha (formerly A. eutrophus) on different substrates (Steinbuchel and Schlegel, supra, 1983).
TABLE-US-00004 TABLE 4 The in vitro activity of R. eutropha ldhA (Steinbuchel and Schlegel, supra, 1983) on different substrates and compared with that on pyruvate. Activity (%) of L(+)-lactate L(+)-lactate D(-)-lactate dehydro- dehydro- dehydro- genase from genase from genase from Substrate A. eutrophus rabbit muscle L. leichmanii Glyoxylate 8.7 23.9 5.0 Pyruvate 100.0 100.0 100.0 2-Oxobutyrate 107.0 18.6 1.1 2-Oxovalerate 125.0 0.7 0.0 3-Methyl-2- 28.5 0.0 0.0 oxobutyrate 3-Methyl-2- 5.3 0.0 0.0 oxovalerate 4-Methyl-2- 39.0 1.4 1.1 oxopentanoate Oxaloacetate 0.0 33.1 23.1 2-Oxoglutarate 79.6 0.0 0.0 3-Fluoropyruvate 33.6 74.3 40.0
[0170] Oxidoreductases that can convert 2-oxoacids to their acyl-CoA counterparts (1.2.1) have been shown to accept multiple substrates as well. For example, branched-chain 2-keto-acid dehydrogenase complex (BCKAD), also known as 2-oxoisovalerate dehydrogenase (1.2.1.25), participates in branched-chain amino acid degradation pathways, converting 2-keto acids derivatives of valine, leucine and isoleucine to their acyl-CoA derivatives and CO.sub.2. In some organisms including Rattus norvegicus (Paxton et al., Biochem. J. 234:295-303 (1986)) and Saccharomyces cerevisiae (Sinclair et al., Biochem. Mol Biol. Int. 32:911-922 (1993), this complex has been shown to have a broad substrate range that includes linear oxo-acids such as 2-oxobutanoate and alpha-ketoglutarate, in addition to the branched-chain amino acid precursors.
[0171] Members of yet another class of enzymes, namely aminotransferases (2.6.1), have been reported to act on multiple substrates. Aspartate aminotransferase (aspAT) from Pyrococcus fursious has been identified, expressed in E. coli and the recombinant protein characterized to demonstrate that the enzyme has the highest activities towards aspartate and alpha-ketoglutarate but lower, yet significant activities towards alanine, glutamate and the aromatic amino acids (Ward et al., Archaea 133-141 (2002)). In another instance, an aminotransferase identified from Leishmania mexicana and expressed in E. coli (Vernal et al., FEMS Microbiol. Lett. 229:217-222 (2003)) was reported to have a broad substrate specificity towards tyrosine (activity considered 100% on tyrosine), phenylalanine (90%), tryptophan (85%), aspartate (30%), leucine (25%) and methionine (25%), respectively (Vernal et al., Mol. Biochem. Parasitol 96:83-92 (1998)). Similar broad specificity has been reported for a tyrosine aminotransferase from Trypanosoma cruzi, even though both of these enzymes have a sequence homology of only 6%. The latter enzyme can accept leucine, methionine as well as tyrosine, phenylalanine, tryptophan and alanine as efficient amino donors (Nowicki et al., Biochim. Biophys. Acta 1546: 268-281 (2001)).
[0172] CoA transferases (2.8.3) have been demonstrated to have the ability to act on more than one substrate. Specifically, a CoA transferase was purified from Clostridium acetobutylicum and was reported to have the highest activities on acetate, propionate, and butyrate. It also had significant activities with valerate, isobutyrate, and crotonate (Wiesenborn et al., Appl. Environ. Microbiol. 55:323-329 (1989)). In another study, the E. coli enzyme acyl-CoA:acetate-CoA transferase, also known as acetate-CoA transferase (EC 2.8.3.8), has been shown to transfer the CoA moiety to acetate from a variety of branched and linear acyl-CoA substrates, including isobutyrate (Matthies and Schink, App. Environm. Microbiol. 58:1435-1439 (1992)), valerate (Vanderwinkel et al., Biochem. Biophys. Res Commun. 33:902-908 (1968b)) and butanoate (Vanderwinkel et al., Biochem. Biophys. Res Commun. 33:902-908 (1968a).
[0173] Other enzyme classes additionally support broad substrate specificity for enzymes. Some isomerases (5.3.3) have also been proven to operate on multiple substrates. For example, L-rhamnose isomerase from Pseudomonas stutzeri catalyzes the isomerization between various aldoalses and ketoses (Yoshida et al., J. Mol. Biol. 365:1505-1516 (2007)). These include isomerization between L-rhamnose and L-rhamnulose, L-mannose and L-fructose, L-xylose and L-xylulose, D-ribose and D-ribulose, and D-allose and D-psicose.
[0174] In yet another class of enzymes, the phosphotransferases (2.7.1), the homoserine kinase (2.7.1.39) from E. coli that converts L-homoserine to L-homoserine phosphate, was found to phosphorylate numerous homoserine analogs. In these substrates, the carboxyl functional group at the R-position had been replaced by an ester or by a hydroxymethyl group (Huo and Viola, Biochemistry 35:16180-16185 (1996)). Table 5 demonstrates the broad substrate specificity of this kinase.
TABLE-US-00005 TABLE 5 The substrate specificity of homoserine kinase. K.sub.m Substrate k.sub.cat % k.sub.cat (mM) k.sub.cat/K.sub.m L-homoserine 18.3 .+-. 0.1 100 0.14 .+-. 0.04 184 .+-. 17 D-homoserine 8.3 .+-. 1.1 32 31.8 .+-. 7.2 0.26 .+-. 0.03 L-aspartate 2.1 .+-. 0.1 8.2 0.28 .+-. 0.02 7.5 .+-. 0.3 .beta.-semialdehyde L-2-amino-1,4- 2.0 .+-. 0.5 7.9 11.6 .+-. 6.5 0.17 .+-. 0.06 butanediol L-2-amino-5- 2.5 .+-. 0.4 9.9 1.1 .+-. 0.5 2.3 .+-. 0.3 hydroxyvalerate L-homoserine 14.7 .+-. 2.6 80 4.9 .+-. 2.0 3.0 .+-. 0.6 methyl ester L-homoserine 13.6 .+-. 0.8 74 1.9 .+-. 0.5 7.2 .+-. 1.7 ethyl ester L-homoserine 13.6 .+-. 1.4 74 1.2 .+-. 0.5 11.3 .+-. 1.1 isopropyl ester L-homoserine 14.0 .+-. 0.4 76 3.5 .+-. 0.4 4.0 .+-. 1.2 n-propyl ester L-homoserine 16.4 .+-. 0.8 84 6.9 .+-. 1.1 2.4 .+-. 0.3 isobutyl ester L-homserine 29.1 .+-. 1.2 160 5.8 .+-. 0.8 5.0 .+-. 0.5 n-butyl ester
[0175] Another class of enzymes useful in BDO pathways is the acid-thiol ligases (6.2.1). Like enzymes in other classes, certain enzymes in this class have been determined to have broad substrate specificity. For example, acyl CoA ligase from Pseudomonas putida has been demonstrated to work on several aliphatic substrates including acetic, propionic, butyric, valeric, hexanoic, heptanoic, and octanoic acids and on aromatic compounds such as phenylacetic and phenoxyacetic acids (Fernandez-Valverde et al., Appl. Environ. Microbiol. 59:1149-1154 (1993)). A related enzyme, malonyl CoA synthetase (6.3.4.9) from Rhizobium trifolii could convert several diacids, namely, ethyl-, propyl-, allyl-, isopropyl-, dimethyl-, cyclopropyl-, cyclopropylmethylene-, cyclobutyl-, and benzyl-malonate into their corresponding monothioesters (Pohl et al., J. Am. Chem. Soc. 123:5822-5823 (2001)). Similarly, decarboxylases (4.1.1) have also been found with broad substrate ranges. Pyruvate decarboxylase (PDC), also termed keto-acid decarboxylase, is a key enzyme in alcoholic fermentation, catalyzing the decarboxylation of pyruvate to acetaldehyde. The enzyme isolated from Saccharomyces cerevisiae has a broad substrate range for aliphatic 2-keto acids including 2-ketobutyrate, 2-ketovalerate, and 2-phenylpyruvate (Li and Jordan, Biochemistry 38:10004-10012 (1999)). Similarly, benzoylformate decarboxylase has a broad substrate range and has been the target of enzyme engineering studies. The enzyme from Pseudomonas putida has been extensively studied and crystal structures of this enzyme are available (Polovnikova et al., Biochemistry 42:1820-1830 (2003); Hasson et al., Biochemistry 37:9918-9930 (1998)). Branched chain alpha-ketoacid decarboxylase (BCKA) has been shown to act on a variety of compounds varying in chain length from 3 to 6 carbons (Oku and Kaneda, J. Biol. Chem. 263:18386-18396 (1998); Smit et al., Appl. Environ. Microbiol. 71:303-311 (2005b)). The enzyme in Lactococcus lactis has been characterized on a variety of branched and linear substrates including 2-oxobutanoate, 2-oxohexanoate, 2-oxopentanoate, 3-methyl-2-oxobutanoate, 4-methyl-2-oxobutanoate and isocaproate (Smit et al., Appl. Environ. Microbiol. 71:303-311 (2005a).
[0176] Interestingly, enzymes known to have one dominant activity have also been reported to catalyze a very different function. For example, the cofactor-dependent phosphoglycerate mutase (5.4.2.1) from Bacillus stearothermophilus and Bacillus subtilis is known to function as a phosphatase as well (Rigden et al., Protein Sci. 10:1835-1846 (2001)). The enzyme from B. stearothermophilus is known to have activity on several substrates, including 3-phosphoglycerate, alpha-napthylphosphate, p-nitrophenylphosphate, AMP, fructose-6-phosphate, ribose-5-phosphate and CMP.
[0177] In contrast to these examples where the enzymes naturally have broad substrate specificities, numerous enzymes have been modified using directed evolution to broaden their specificity towards their non-natural substrates. Alternatively, the substrate preference of an enzyme has also been changed using directed evolution. Therefore, it is feasible to engineer a given enzyme for efficient function on a natural, for example, improved efficiency, or a non-natural substrate, for example, increased efficiency. For example, it has been reported that the enantioselectivity of a lipase from Pseudomonas aeruginosa was improved significantly (Reetz et al., Agnew. Chem. Int. Ed Engl. 36:2830-2832 (1997)). This enzyme hydrolyzed p-nitrophenyl 2-methyldecanoate with only 2% enantiomeric excess (ee) in favor of the (S)-acid. However, after four successive rounds of error-prone mutagenesis and screening, a variant was produced that catalyzed the requisite reaction with 81% ee (Reetz et al., Agnew. Chem. Int. Ed Engl. 36:2830-2832 (1997)).
[0178] Directed evolution methods have been used to modify an enzyme to function on an array of non-natural substrates. The substrate specificity of the lipase in P. aeruginosa was broadened by randomization of amino acid residues near the active site. This allowed for the acceptance of alpha-substituted carboxylic acid esters by this enzyme (Reetz et al., Agnew. Chem. Int. Ed Engl. 44:4192-4196 (2005)). In another successful modification of an enzyme, DNA shuffling was employed to create an Escherichia coli aminotransferase that accepted .beta.-branched substrates, which were poorly accepted by the wild-type enzyme (Yano et al., Proc. Nat. Acad. Sci. U.S.A. 95:5511-5515 (1998)). Specifically, at the end of four rounds of shuffling, the activity of aspartate aminotransferase for valine and 2-oxovaline increased by up to five orders of magnitude, while decreasing the activity towards the natural substrate, aspartate, by up to 30-fold. Recently, an algorithm was used to design a retro-aldolase that could be used to catalyze the carbon-carbon bond cleavage in a non-natural and non-biological substrate, 4-hydroxy-4-(6-methoxy-2-naphthyl)-2-butanone (Jiang et al., Science 319:1387-1391 (2008)). These algorithms used different combinations of four different catalytic motifs to design new enzyme, and 20 of the selected designs for experimental characterization had four-fold improved rates over the uncatalyzed reaction (Jiang et al., Science 319:1387-1391 (2008)). Thus, not only are these engineering approaches capable of expanding the array of substrates on which an enzyme can act, but they allow the design and construction of very efficient enzymes. For example, a method of DNA shuffling (random chimeragenesis on transient templates or RACHITT) was reported to lead to an engineered monooxygenase that had an improved rate of desulfurization on complex substrates as well as 20-fold faster conversion of a non-natural substrate (Coco et al., Nat. Biotechnol. 19:354-359 (2001)). Similarly, the specific activity of a sluggish mutant triosephosphate isomerase enzyme was improved up to 19-fold from 1.3 fold (Hermes et al., Proc. Nat. Acad. Sci. U.S.A. 87:696-700 1990)). This enhancement in specific activity was accomplished by using random mutagenesis over the whole length of the protein and the improvement could be traced back to mutations in six amino acid residues.
[0179] The effectiveness of protein engineering approaches to alter the substrate specificity of an enzyme for a desired substrate has also been demonstrated in several studies. Isopropylmalate dehydrogenase from Thermus thermophilus was modified by changing residues close to the active site so that it could now act on malate and D-lactate as substrates (Fujita et al., Biosci. Biotechnol. Biochem. 65:2695-2700 (2001)). In this study as well as in others, it was pointed out that one or a few residues could be modified to alter the substrate specificity. For example, the dihydroflavonol 4-reductase for which a single amino acid was changed in the presumed substrate-binding region could preferentially reduce dihydrokaempferol (Johnson et al., Plant. J. 25:325-333 (2001)). The substrate specificity of a very specific isocitrate dehydrogenase from Escherichia coli was changed form isocitrate to isopropylmalate by changing one residue in the active site (Doyle et al., Biochemistry 40:4234-4241 (2001)). Similarly, the cofactor specificity of a NAD.sup.+-dependent 1,5-hydroxyprostaglandin dehydrogenase was altered to NADP.sup.+ by changing a few residues near the N-terminal end (Cho et al., Arch. Biochem. Biophys. 419:139-146 (2003)). Sequence analysis and molecular modeling analysis were used to identify the key residues for modification, which were further studied by site-directed mutagenesis.
[0180] Numerous examples exist spanning diverse classes of enzymes where the function of enzyme was changed to favor one non-natural substrate over the natural substrate of the enzyme. A fucosidase was evolved from a galactosidase in E. coli by DNA shuffling and screening (Zhang et al., Proc. Natl Acad. Sci. U.S.A. 94:4504-4509 (1997)). Similarly, aspartate aminotransferase from E. coli was converted into a tyrosine aminotransferase using homology modeling and site-directed mutagenesis (Onuffer and Kirsch, Protein Sci., 4:1750-1757 (1995)). Site-directed mutagenesis of two residues in the active site of benzoylformate decarboxylase from P. putida reportedly altered the affinity (K.sub.m) towards natural and non-natural substrates (Siegert et al., Protein Eng Des Sel 18:345-357 (2005)). Cytochrome c peroxidase (CCP) from Saccharomyces cerevisiae was subjected to directed molecular evolution to generate mutants with increased activity against the classical peroxidase substrate guaiacol, thus changing the substrate specificity of CCP from the protein cytochrome c to a small organic molecule. After three rounds of DNA shuffling and screening, mutants were isolated which possessed a 300-fold increased activity against guaiacol and up to 1000-fold increased specificity for this substrate relative to that for the natural substrate (Iffland et al., Biochemistry 39:10790-10798 (2000)).
[0181] In some cases, enzymes with different substrate preferences than either of the parent enzymes have been obtained. For example, biphenyl-dioxygenase-mediated degradation of polychlorinated biphenyls was improved by shuffling genes from two bacteria, Pseudomonas pseudoalcaligens and Burkholderia cepacia (Kumamaru et al., Nat. Biotechnol. 16:663-666 (1998)). The resulting chimeric biphenyl oxygenases showed different substrate preferences than both the parental enzymes and enhanced the degradation activity towards related biphenyl compounds and single aromatic ring hydrocarbons such as toluene and benzene which were originally poor substrates for the enzyme.
[0182] In addition to changing enzyme specificity, it is also possible to enhance the activities on substrates for which the enzymes naturally have low activities. One study demonstrated that amino acid racemase from P. putida that had broad substrate specificity (on lysine, arginine, alanine, serine, methionine, cysteine, leucine and histidine among others) but low activity towards tryptophan could be improved significantly by random mutagenesis (Kino et al., Appl. Microbiol. Biotechnol. 73:1299-1305 (2007)). Similarly, the active site of the bovine BCKAD was engineered to favor alternate substrate acetyl-CoA (Meng and Chuang, Biochemistry 33:12879-12885 (1994)). An interesting aspect of these approaches is that even if random methods have been applied to generate these mutated enzymes with efficacious activities, the exact mutations or structural changes that confer the improvement in activity can be identified. For example, in the aforementioned study, the mutations that facilitated improved activity on tryptophan was traced back to two different positions.
[0183] Directed evolution has also been used to express proteins that are difficult to express. For example, by subjecting horseradish peroxidase to random mutagenesis and gene recombination, mutants were identified that had more than 14-fold higher activity than the wild type (Lin et al., Biotechnol. Prog. 15:467-471 (1999)).
[0184] Another example of directed evolution shows the extensive modifications to which an enzyme can be subjected to achieve a range of desired functions. The enzyme lactate dehydrogenase from Bacillus stearothermophilus was subjected to site-directed mutagenesis, and three amino acid substitutions were made at sites that were believed to determine the specificity towards different hydroxyacids (Clarke et al., Biochem. Biophys. Res. Commun. 148:15-23 (1987)). After these mutations, the specificity for oxaloacetate over pyruvate was increased to 500 in contrast to the wild type enzyme that had a catalytic specificity for pyruvate over oxaloacetate of 1000. This enzyme was further engineered using site-directed mutagenesis to have activity towards branched-chain substituted pyruvates (Wilks et al., Biochemistry 29:8587-8591 (1990)). Specifically, the enzyme had a 55-fold improvement in K.sub.cat for alpha-ketoisocaproate. Three structural modifications were made in the same enzyme to change its substrate specificity from lactate to malate. The enzyme was highly active and specific towards malate (Wilks et al., Science 242:1541-1544 (1988)). The same enzyme from B. stearothermophilus was subsequently engineered to have high catalytic activity towards alpha-keto acids with positively charged side chains, such as those containing ammonium groups (Hogan et al., Biochemistry 34:4225-4230 (1995)). Mutants with acidic amino acids introduced at position 102 of the enzyme favored binding of such side chain ammonium groups. The results obtained proved that the mutants showed up to 25-fold improvements in k.sub.cat/K.sub.m values for omega-amino-alpha-keto acid substrates. Interestingly, this enzyme was also structurally modified to function as a phenyllactate dehydrogenase instead of a lactate dehydrogenase (Wilks et al., Biochemistry 31:7802-7806 1992). Restriction sites were introduced into the gene for the enzyme which allowed a region of the gene to be excised. This region coded for a mobile surface loop of the polypeptide (residues 98-110) which normally seals the active site from bulk solvent and is a major determinant of substrate specificity. The variable length and sequence loops were inserted so that hydroxyacid dehydrogenases with altered substrate specificities were generated. With one longer loop construction, activity with pyruvate was reduced one-million-fold but activity with phenylpyruvate was largely unaltered. A switch in specificity (k.sub.cat/K.sub.m) of 390,000-fold was achieved. The 1700:1 selectivity of this enzyme for phenylpyruvate over pyruvate is that required in a phenyllactate dehydrogenase. The studies described above indicate that various approaches of enzyme engineering can be used to obtain enzymes for the BDO pathways as disclosed herein.
[0185] As disclosed herein, biosynthetic pathways to 1,4-butanediol from a number of central metabolic intermediates are can be utilized, including acetyl-CoA, succinyl-CoA, alpha-ketoglutarate, glutamate, 4-aminobutyrate, and homoserine. Acetyl-CoA, succinyl-CoA and alpha-ketoglutarate are common intermediates of the tricarboxylic acid (TCA) cycle, a series of reactions that is present in its entirety in nearly all living cells that utilize oxygen for cellular respiration and is present in truncated forms in a number of anaerobic organisms. Glutamate is an amino acid that is derived from alpha-ketoglutarate via glutamate dehydrogenase or any of a number of transamination reactions (see FIG. 8B). 4-aminobutyrate can be formed by the decarboxylation of glutamate (see FIG. 8B) or from acetoacetyl-CoA via the pathway disclosed in FIG. 9C. Acetoacetyl-CoA is derived from the condensation of two acetyl-CoA molecules by way of the enzyme, acetyl-coenzyme A acetyltransferase, or equivalently, acetoacetyl-coenzyme A thiolase. Homoserine is an intermediate in threonine and methionine metabolism, formed from oxaloacetate via aspartate. The conversion of oxaloacetate to homoserine requires one NADH, two NADPH, and one ATP.
[0186] Pathways other than those exemplified above also can be employed to generate the biosynthesis of BDO in non-naturally occurring microbial organisms. In one embodiment, biosynthesis can be achieved using a L-homoserine to BDO pathway (see FIG. 13). This pathway has a molar yield of 0.90 mol/mol glucose, which appears restricted by the availability of reducing equivalents. A second pathway synthesizes BDO from acetoacetyl-CoA and is capable of achieving the maximum theoretical yield of 1.091 mol/mol glucose (see FIG. 9). Implementation of either pathway can be achieved by introduction of two exogenous enzymes into a host organism such as E. coli, and both pathways can additionally complement BDO production via succinyl-CoA. Pathway enzymes, thermodynamics, theoretical yields and overall feasibility are described further below.
[0187] A homoserine pathway also can be engineered to generate BDO-producing microbial organisms. Homoserine is an intermediate in threonine and methionine metabolism, formed from oxaloacetate via aspartate. The conversion of oxaloacetate to homoserine requires one NADH, two NADPH, and one ATP (FIG. 2). Once formed, homoserine feeds into biosynthetic pathways for both threonine and methionine. In most organisms, high levels of threonine or methionine feedback to repress the homoserine biosynthesis pathway (Caspi et al., Nucleic Acids Res. 34:D511-D516 (1990)).
[0188] The transformation of homoserine to 4-hydroxybutyrate (4-HB) can be accomplished in two enzymatic steps as described herein. The first step of this pathway is deamination of homoserine by a putative ammonia lyase. In step 2, the product alkene, 4-hydroxybut-2-enoate is reduced to 4-HB by a putative reductase at the cost of one NADH. 4-HB can then be converted to BDO.
[0189] Enzymes available for catalyzing the above transformations are disclosed herein. For example, the ammonia lyase in step 1 of the pathway closely resembles the chemistry of aspartate ammonia-lyase (aspartase). Aspartase is a widespread enzyme in microorganisms, and has been characterized extensively (Viola, R. E., Mol. Biol. 74:295-341 (2008)). The crystal structure of the E. coli aspartase has been solved (Shi et al., Biochemistry 36:9136-9144 (1997)), so it is therefore possible to directly engineer mutations in the enzyme's active site that would alter its substrate specificity to include homoserine. The oxidoreductase in step 2 has chemistry similar to several well-characterized enzymes including fumarate reductase in the E. coli TCA cycle. Since the thermodynamics of this reaction are highly favorable, an endogenous reductase with broad substrate specificity will likely be able to reduce 4-hydroxybut-2-enoate. The yield of this pathway under anaerobic conditions is 0.9 mol BDO per mol glucose.
[0190] The succinyl-CoA pathway was found to have a higher yield due to the fact that it is more energetically efficient. The conversion of one oxaloacetate molecule to BDO via the homoserine pathway will require the expenditure of 2 ATP equivalents. Because the conversion of glucose to two oxaloacetate molecules can generate a maximum of 3 ATP molecules assuming PEP carboxykinase to be reversible, the overall conversion of glucose to BDO via homoserine has a negative energetic yield. As expected, if it is assumed that energy can be generated via respiration, the maximum yield of the homoserine pathway increases to 1.05 mol/mol glucose which is 96% of the succinyl-CoA pathway yield. The succinyl-CoA pathway can channel some of the carbon flux through pyruvate dehydrogenase and the oxidative branch of the TCA cycle to generate both reducing equivalents and succinyl-CoA without an energetic expenditure. Thus, it does not encounter the same energetic difficulties as the homoserine pathway because not all of the flux is channeled through oxaloacetate to succinyl-CoA to BDO. Overall, the homoserine pathway demonstrates a high-yielding route to BDO.
[0191] An acetoacetate pathway also can be engineered to generate BDO-producing microbial organisms. Acetoacetate can be formed from acetyl-CoA by enzymes involved in fatty acid metabolism, including acetyl-CoA acetyltransferase and acetoacetyl-CoA transferase. Biosynthetic routes through acetoacetate are also particularly useful in microbial organisms that can metabolize single carbon compounds such as carbon monoxide, carbon dioxide or methanol to form acetyl-CoA.
[0192] A three step route from acetoacetyl-CoA to 4-aminobutyrate (see FIG. 9C) can be used to synthesize BDO through acetoacetyl-CoA. 4-Aminobutyrate can be converted to succinic semialdehyde as shown in FIG. 8B. Succinic semialdehyde, which is one reduction step removed from succinyl-CoA or one decarboxylation step removed from .alpha.-ketoglutarate, can be converted to BDO following three reductions steps (FIG. 1). Briefly, step 1 of this pathway involves the conversion of acetoacetyl-CoA to acetoacetate by, for example, the E. coli acetoacetyl-CoA transferase encoded by the atoA and atoD genes (Hanai et al., Appl. Environ. Microbiol. 73: 7814-7818 (2007)). Step 2 of the acetoacetyl-CoA biopathway entails conversion of acetoacetate to 3-aminobutanoate by an .omega.-aminotransferase. The .omega.-amino acid:pyruvate aminotransferase (.omega.-APT) from Alcaligens denitrificans was overexpressed in E. coli and shown to have a high activity toward 3-aminobutanoate in vitro (Yun et al., Appl. Environ. Microbiol. 70:2529-2534 (2004)).
[0193] In step 2, a putative aminomutase shifts the amine group from the 3- to the 4-position of the carbon backbone. An aminomutase performing this function on 3-aminobutanoate has not been characterized, but an enzyme from Clostridium sticklandii has a very similar mechanism. The enzyme, D-lysine-5,6-aminomutase, is involved in lysine biosynthesis.
[0194] The synthetic route to BDO from acetoacetyl-CoA passes through 4-aminobutanoate, a metabolite in E. coli that is normally formed from decarboxylation of glutamate. Once formed, 4-aminobutanoate can be converted to succinic semialdehyde by 4-aminobutanoate transaminase (2.6.1.19), an enzyme which has been biochemically characterized.
[0195] One consideration for selecting candidate enzymes in this pathway is the stereoselectivity of the enzymes involved in steps 2 and 3. The .omega.-ABT in Alcaligens denitrificans is specific to the L-stereoisomer of 3-aminobutanoate, while D-lysine-5,6-aminomutase likely requires the D-stereoisomer. If enzymes with complementary stereoselectivity are not initially found or engineered, a third enzyme can be added to the pathway with racemase activity that can convert L-3-aminobutanoate to D-3-aminobutanoate. While amino acid racemases are widespread, whether these enzymes can function on .omega.-amino acids is not known.
[0196] The maximum theoretical molar yield of this pathway under anaerobic conditions is 1.091 mol/mol glucose. In order to generate flux from acetoacetyl-CoA to BDO it was necessary to assume that acetyl-CoA:acetoacetyl-CoA transferase is reversible. The function of this enzyme in E. coli is to metabolize short-chain fatty acids by first converting them into thioesters.
[0197] While the operation of acetyl-CoA:acetoacetyl-CoA transferase in the acetate-consuming direction has not been demonstrated experimentally in E. coli, studies on similar enzymes in other organisms support the assumption that this reaction is reversible. The enzyme butyryl-CoA:acetate:CoA transferase in gut microbes Roseburia sp. and F. prasnitzii operates in the acetate utilizing direction to produce butyrate (Duncan et al., Appl. Environ. Microbiol 68:5186-5190 (2002)). Another very similar enzyme, acetyl: succinate CoA-transferase in Trypanosoma brucei, also operates in the acetate utilizing direction. This reaction has a .DELTA..sub.rxnG close to equilibrium, so high concentrations of acetate can likely drive the reaction in the direction of interest. At the maximum theoretical BDO production rate of 1.09 mol/mol glucose simulations predict that E. coli can generate 1.098 mol ATP per mol glucose with no fermentation byproducts. This ATP yield should be sufficient for cell growth, maintenance, and production. The acetoacetatyl-CoA biopathway is a high-yielding route to BDO from acetyl-CoA.
[0198] Therefore, in addition to any of the various modifications exemplified previously for establishing 4-HB biosynthesis in a selected host, the BDO producing microbial organisms can include any of the previous combinations and permutations of 4-HB pathway metabolic modifications as well as any combination of expression for CoA-independent aldehyde dehydrogenase, CoA-dependent aldehyde dehydrogenase or an alcohol dehydrogenase or other enzymes disclosed herein to generate biosynthetic pathways for GBL and/or BDO. Therefore, the BDO producers of the invention can have exogenous expression of, for example, one, two, three, four, five, six, seven, eight, nine, or up to all enzymes corresponding to any of the 4-HB pathway and/or any of the BDO pathway enzymes disclosed herein.
[0199] Design and construction of the genetically modified microbial organisms is carried out using methods well known in the art to achieve sufficient amounts of expression to produce BDO. In particular, the non-naturally occurring microbial organisms of the invention can achieve biosynthesis of BDO resulting in intracellular concentrations between about 0.1-200 mM or more, such as about 0.1-25 mM or more, as discussed above. For example, the intracellular concentration of BDO is between about 3-20 mM, particularly between about 5-15 mM and more particularly between about 8-12 mM, including about 10 mM or more. Intracellular concentrations between and above each of these exemplary ranges also can be achieved from the non-naturally occurring microbial organisms of the invention. As with the 4-HB producers, the BDO producers also can be sustained, cultured or fermented under anaerobic conditions.
[0200] The invention further provides a method for the production of 4-HB. The method includes culturing a non-naturally occurring microbial organism having a 4-hydroxybutanoic acid (4-HB) biosynthetic pathway comprising at least one exogenous nucleic acid encoding 4-hydroxybutanoate dehydrogenase, CoA-independent succinic semialdehyde dehydrogenase, succinyl-CoA synthetase, CoA-dependent succinic semialdehyde dehydrogenase, glutamate:succinic semialdehyde transaminase, .alpha.-ketoglutarate decarboxylase, or glutamate decarboxylase under substantially anaerobic conditions for a sufficient period of time to produce monomeric 4-hydroxybutanoic acid (4-HB). The method can additionally include chemical conversion of 4-HB to GBL and to BDO or THF, for example.
[0201] Additionally provided is a method for the production of 4-HB. The method includes culturing a non-naturally occurring microbial organism having a 4-hydroxybutanoic acid (4-HB) biosynthetic pathway including at least one exogenous nucleic acid encoding 4-hydroxybutanoate dehydrogenase, succinyl-CoA synthetase, CoA-dependent succinic semialdehyde dehydrogenase or .alpha.-ketoglutarate decarboxylase under substantially anaerobic conditions for a sufficient period of time to produce monomeric 4-hydroxybutanoic acid (4-HB). The 4-HB product can be secreted into the culture medium.
[0202] Further provided is a method for the production of BDO. The method includes culturing a non-naturally occurring microbial biocatalyst or microbial organism, comprising a microbial organism having 4-hydroxybutanoic acid (4-HB) and 1,4-butanediol (BDO) biosynthetic pathways, the pathways including at least one exogenous nucleic acid encoding 4-hydroxybutanoate dehydrogenase, succinyl-CoA synthetase, CoA-dependent succinic semialdehyde dehydrogenase, 4-hydroxybutyrate:CoA transferase, 4-hydroxybutyrate kinase, phosphotranshydroxybutyrylase, .alpha.-ketoglutarate decarboxylase, aldehyde dehydrogenase, alcohol dehydrogenase or an aldehyde/alcohol dehydrogenase for a sufficient period of time to produce 1,4-butanediol (BDO). The BDO product can be secreted into the culture medium.
[0203] Additionally provided are methods for producing BDO by culturing a non-naturally occurring microbial organism having a BDO pathway of the invention. The BDO pathway can comprise at least one exogenous nucleic acid encoding a BDO pathway enzyme expressed in a sufficient amount to produce BDO, under conditions and for a sufficient period of time to produce BDO, the BDO pathway comprising 4-aminobutyrate CoA transferase, 4-aminobutyryl-CoA hydrolase, 4-aminobutyrate-CoA ligase, 4-aminobutyryl-CoA oxidoreductase (deaminating), 4-aminobutyryl-CoA transaminase, or 4-hydroxybutyryl-CoA dehydrogenase (see Example VII and Table 17).
[0204] Alternatively, the BDO pathway can compare at least one exogenous nucleic acid encoding a BDO pathway enzyme expressed in a sufficient amount to produce BDO, under conditions and for a sufficient period of time to produce BDO, the BDO pathway comprising 4-aminobutyrate CoA transferase, 4-aminobutyryl-CoA hydrolase, 4-aminobutyrate-CoA ligase, 4-aminobutyryl-CoA reductase (alcohol forming), 4-aminobutyryl-CoA reductase, 4-aminobutan-1-ol dehydrogenase, 4-aminobutan-1-ol oxidoreductase (deaminating) or 4-aminobutan-1-ol transaminase (see Example VII and Table 18).
[0205] In addition, the invention provides a method for producing BDO, comprising culturing a non-naturally occurring microbial organism having a BDO pathway, the pathway comprising at least one exogenous nucleic acid encoding a BDO pathway enzyme expressed in a sufficient amount to produce BDO, under conditions and for a sufficient period of time to produce BDO, the BDO pathway comprising 4-aminobutyrate kinase, 4-aminobutyraldehyde dehydrogenase (phosphorylating), 4-aminobutan-1-ol dehydrogenase, 4-aminobutan-1-ol oxidoreductase (deaminating), 4-aminobutan-1-ol transaminase, [(4-aminobutanolyl)oxy]phosphonic acid oxidoreductase (deaminating), [(4-aminobutanolyl)oxy]phosphonic acid transaminase, 4-hydroxybutyryl-phosphate dehydrogenase, or 4-hydroxybutyraldehyde dehydrogenase (phosphorylating) (see Example VII and Table 19).
[0206] The invention further provides a method for producing BDO, comprising culturing a non-naturally occurring microbial organism having a BDO pathway, the pathway comprising at least one exogenous nucleic acid encoding a BDO pathway enzyme expressed in a sufficient amount to produce BDO, under conditions and for a sufficient period of time to produce BDO, the BDO pathway comprising alpha-ketoglutarate 5-kinase, 2,5-dioxopentanoic semialdehyde dehydrogenase (phosphorylating), 2,5-dioxopentanoic acid reductase, alpha-ketoglutarate CoA transferase, alpha-ketoglutaryl-CoA hydrolase, alpha-ketoglutaryl-CoA ligase, alpha-ketoglutaryl-CoA reductase, 5-hydroxy-2-oxopentanoic acid dehydrogenase, alpha-ketoglutaryl-CoA reductase (alcohol forming), 5-hydroxy-2-oxopentanoic acid decarboxylase, or 5-hydroxy-2-oxopentanoic acid dehydrogenase (decarboxylation)(see Example VIII and Table 20).
[0207] The invention additionally provides a method for producing BDO, comprising culturing a non-naturally occurring microbial organism having a BDO pathway, the pathway comprising at least one exogenous nucleic acid encoding a BDO pathway enzyme expressed in a sufficient amount to produce BDO, under conditions and for a sufficient period of time to produce BDO, the BDO pathway comprising glutamate CoA transferase, glutamyl-CoA hydrolase, glutamyl-CoA ligase, glutamate 5-kinase, glutamate-5-semialdehyde dehydrogenase (phosphorylating), glutamyl-CoA reductase, glutamate-5-semialdehyde reductase, glutamyl-CoA reductase (alcohol forming), 2-amino-5-hydroxypentanoic acid oxidoreductase (deaminating), 2-amino-5-hydroxypentanoic acid transaminase, 5-hydroxy-2-oxopentanoic acid decarboxylase, 5-hydroxy-2-oxopentanoic acid dehydrogenase (decarboxylation)(see Example IX and Table 21).
[0208] The invention additionally includes a method for producing BDO, comprising culturing a non-naturally occurring microbial organism having a BDO pathway, the pathway comprising at least one exogenous nucleic acid encoding a BDO pathway enzyme expressed in a sufficient amount to produce BDO, under conditions and for a sufficient period of time to produce BDO, the BDO pathway comprising 3-hydroxybutyryl-CoA dehydrogenase, 3-hydroxybutyryl-CoA dehydratase, vinylacetyl-CoA .DELTA.-isomerase, or 4-hydroxybutyryl-CoA dehydratase (see Example X and Table 22).
[0209] Also provided is a method for producing BDO, comprising culturing a non-naturally occurring microbial organism having a BDO pathway, the pathway comprising at least one exogenous nucleic acid encoding a BDO pathway enzyme expressed in a sufficient amount to produce BDO, under conditions and for a sufficient period of time to produce BDO, the BDO pathway comprising homoserine deaminase, homoserine CoA transferase, homoserine-CoA hydrolase, homoserine-CoA ligase, homoserine-CoA deaminase, 4-hydroxybut-2-enoyl-CoA transferase, 4-hydroxybut-2-enoyl-CoA hydrolase, 4-hydroxybut-2-enoyl-CoA ligase, 4-hydroxybut-2-enoate reductase, 4-hydroxybutyryl-CoA transferase, 4-hydroxybutyryl-CoA hydrolase, 4-hydroxybutyryl-CoA ligase, or 4-hydroxybut-2-enoyl-CoA reductase (see Example XI and Table 23).
[0210] The invention additionally provides a method for producing BDO, comprising culturing a non-naturally occurring microbial organism having a BDO pathway, the pathway comprising at least one exogenous nucleic acid encoding a BDO pathway enzyme expressed in a sufficient amount to produce BDO, under conditions and for a sufficient period of time to produce BDO, the BDO pathway comprising succinyl-CoA reductase (alcohol forming), 4-hydroxybutyryl-CoA hydrolase, 4-hydroxybutyryl-CoA ligase, 4-hydroxybutanal dehydrogenase (phosphorylating). Such a BDO pathway can further comprise succinyl-CoA reductase, 4-hydroxybutyrate dehydrogenase, 4-hydroxybutyryl-CoA transferase, 4-hydroxybutyrate kinase, phosphotrans-4-hydroxybutyrylase, 4-hydroxybutyryl-CoA reductase, 4-hydroxybutyryl-CoA reductase (alcohol forming), or 1,4-butanediol dehydrogenase.
[0211] Also provided is a method for producing BDO, comprising culturing a non-naturally occurring microbial organism having a BDO pathway, the pathway comprising at least one exogenous nucleic acid encoding a BDO pathway enzyme expressed in a sufficient amount to produce BDO, under conditions and for a sufficient period of time to produce BDO, the BDO pathway comprising glutamate dehydrogenase, 4-aminobutyrate oxidoreductase (deaminating), 4-aminobutyrate transaminase, glutamate decarboxylase, 4-hydroxybutyryl-CoA hydrolase, 4-hydroxybutyryl-CoA ligase, 4-hydroxybutanal dehydrogenase (phosphorylating).
[0212] The invention additionally provides methods of producing a desired product using the genetically modified organisms disclosed herein that allow improved production of a desired product such as BDO by increasing the product or decreasing undesirable byproducts. Thus, the invention provides a method for producing 1,4-butanediol (BDO), comprising culturing the non-naturally occurring microbial organisms disclosed herein under conditions and for a sufficient period of time to produce BDO. In one embodiment, the invention provides a method of producing BDO using a non-naturally occurring microbial organism, comprising a microbial organism having a 1,4-butanediol (BDO) pathway comprising at least one exogenous nucleic acid encoding a BDO pathway enzyme expressed in a sufficient amount to produce BDO. In one embodiment, the microbial organism is genetically modified to express exogenous succinyl-CoA synthetase (see Example XII). For example, the succinyl-CoA synthetase can be encoded by an Escherichia coli sucCD genes.
[0213] In another embodiment, the microbial organism is genetically modified to express exogenous alpha-ketoglutarate decarboxylase (see Example XIII) For example, the alpha-ketoglutarate decarboxylase can be encoded by the Mycobacterium bovis sucA gene. In still another embodiment, the microbial organism is genetically modified to express exogenous succinate semialdehyde dehydrogenase and 4-hydroxybutyrate dehydrogenase and optionally 4-hydroxybutyryl-CoA/acetyl-CoA transferase (see Example XIII) For example, the succinate semialdehyde dehydrogenase (CoA-dependent), 4-hydroxybutyrate dehydrogenase and 4-hydroxybutyryl-CoA/acetyl-CoA transferase can be encoded by Porphyromonas gingivalis W83 genes. In an additional embodiment, the microbial organism is genetically modified to express exogenous butyrate kinase and phosphotransbutyrylase (see Example XIII) For example, the butyrate kinase and phosphotransbutyrylase can be encoded by Clostridium acetobutilicum buk1 and ptb genes.
[0214] In yet another embodiment, the microbial organism is genetically modified to express exogenous 4-hydroxybutyryl-CoA reductase (see Example XIII) For example, the 4-hydroxybutyryl-CoA reductase can be encoded by Clostridium beijerinckii ald gene. Additionally, in an embodiment of the invention, the microbial organism is genetically modified to express exogenous 4-hydroxybutanal reductase (see Example XIII) For example, the 4-hydroxybutanal reductase can be encoded by Geobacillus thermoglucosidasius adh1 gene. In another embodiment, the microbial organism is genetically modified to express exogenous pyruvate dehydrogenase subunits (see Example XIV). For example, the exogenous pyruvate dehydrogenase can be NADH insensitive. The pyruvate dehydrogenase subunit can be encoded by the Klebsiella pneumonia lpdA gene. In a particular embodiment, the pyruvate dehydrogenase subunit genes of the microbial organism can be under the control of a pyruvate formate lyase promoter.
[0215] In still another embodiment, the microbial organism is genetically modified to disrupt a gene encoding an aerobic respiratory control regulatory system (see Example XV). For example, the disruption can be of the arcA gene. Such an organism can further comprise disruption of a gene encoding malate dehydrogenase. In a further embodiment, the microbial organism is genetically modified to express an exogenous NADH insensitive citrate synthase (see Example XV). For example, the NADH insensitive citrate synthase can be encoded by gltA, such as an R163L mutant of gltA. In still another embodiment, the microbial organism is genetically modified to express exogenous phosphoenolpyruvate carboxykinase (see Example XVI). For example, the phosphoenolpyruvate carboxykinase can be encoded by an Haemophilus influenza phosphoenolpyruvate carboxykinase gene. It is understood that strains exemplified herein for improved production of BDO can similarly be used, with appropriate modifications, to produce other desired products, for example, 4-hydroxybutyrate or other desired products disclosed herein.
[0216] The invention additionally provides a method for producing 4-hydroxybutanal by culturing a non-naturally occurring microbial organism, comprising a 4-hydroxybutanal pathway comprising at least one exogenous nucleic acid encoding a 4-hydroxybutanal pathway enzyme expressed in a sufficient amount to produce 4-hydroxybutanal, the 4-hydroxybutanal pathway comprising succinyl-CoA reductase (aldehyde forming); 4-hydroxybutyrate dehydrogenase; and 4-hydroxybutyrate reductase (see FIG. 58, steps A-C-D). The invention also provides a method for producing 4-hydroxybutanal by culturing a non-naturally occurring microbial organism, comprising a 4-hydroxybutanal pathway comprising at least one exogenous nucleic acid encoding a 4-hydroxybutanal pathway enzyme expressed in a sufficient amount to produce 4-hydroxybutanal, the 4-hydroxybutanal pathway comprising alpha-ketoglutarate decarboxylase; 4-hydroxybutyrate dehydrogenase; and 4-hydroxybutyrate reductase (FIG. 58, steps B-C-D).
[0217] The invention further provides a method for producing 4-hydroxybutanal by culturing a non-naturally occurring microbial organism, comprising a 4-hydroxybutanal pathway comprising at least one exogenous nucleic acid encoding a 4-hydroxybutanal pathway enzyme expressed in a sufficient amount to produce 4-hydroxybutanal, the 4-hydroxybutanal pathway comprising succinate reductase; 4-hydroxybutyrate dehydrogenase, and 4-hydroxybutyrate reductase (see FIG. 62, steps F-C-D). In yet another embodiment, the invention provides a method for producing 4-hydroxybutanal by culturing a non-naturally occurring microbial organism, comprising a 4-hydroxybutanal pathway comprising at least one exogenous nucleic acid encoding a 4-hydroxybutanal pathway enzyme expressed in a sufficient amount to produce 4-hydroxybutanal, the 4-hydroxybutanal pathway comprising alpha-ketoglutarate decarboxylase, or glutamate dehydrogenase or glutamate transaminase and glutamate decarboxylase and 4-aminobutyrate dehydrogenase or 4-aminobutyrate transaminase; 4-hydroxybutyrate dehydrogenase; and 4-hydroxybutyrate reductase (see FIG. 62, steps B or ((J or K)-L-(M or N))-C-D).
[0218] The invention also provides a method for producing 4-hydroxybutanal by culturing a non-naturally occurring microbial organism, comprising a 4-hydroxybutanal pathway comprising at least one exogenous nucleic acid encoding a 4-hydroxybutanal pathway enzyme expressed in a sufficient amount to produce 4-hydroxybutanal, the 4-hydroxybutanal pathway comprising alpha-ketoglutarate reductase; 5-hydroxy-2-oxopentanoate dehydrogenase; and 5-hydroxy-2-oxopentanoate decarboxylase (see FIG. 62, steps X-Y-Z). The invention further provides a method for producing 4-hydroxybutyryl-CoA by culturing a non-naturally occurring microbial organism, comprising a 4-hydroxybutyryl-CoA pathway comprising at least one exogenous nucleic acid encoding a 4-hydroxybutyryl-CoA pathway enzyme expressed in a sufficient amount to produce 4-hydroxybutyryl-CoA, the 4-hydroxybutyryl-CoA pathway comprising alpha-ketoglutarate reductase; 5-hydroxy-2-oxopentanoate dehydrogenase; and 5-hydroxy-2-oxopentanoate dehydrogenase (decarboxylation) (see FIG. 62, steps X-Y-AA).
[0219] The invention additionally provides a method for producing putrescine by culturing a non-naturally occurring microbial organism, comprising a putrescine pathway comprising at least one exogenous nucleic acid encoding a putrescine pathway enzyme expressed in a sufficient amount to produce putrescine, the putrescine pathway comprising succinate reductase; 4-aminobutyrate dehydrogenase or 4-aminobutyrate transaminase; 4-aminobutyrate reductase; and putrescine dehydrogenase or putrescine transaminase (see FIG. 63, steps F-M/N-C-D/E). In still another embodiment, the invention provides a method for producing putrescine by culturing a non-naturally occurring microbial organism, comprising a putrescine pathway comprising at least one exogenous nucleic acid encoding a putrescine pathway enzyme expressed in a sufficient amount to produce putrescine, the putrescine pathway comprising alpha-ketoglutarate decarboxylase; 4-aminobutyrate dehydrogenase or 4-aminobutyrate transaminase; 4-aminobutyrate reductase; and putrescine dehydrogenase or putrescine transaminase (see FIG. 63, steps B-M/N-C-D/E). The invention additionally provides a method for producing putrescine by culturing a non-naturally occurring microbial organism, comprising a putrescine pathway comprising at least one exogenous nucleic acid encoding a putrescine pathway enzyme expressed in a sufficient amount to produce putrescine, the putrescine pathway comprising glutamate dehydrogenase or glutamate transaminase; glutamate decarboxylase; 4-aminobutyrate reductase; and putrescine dehydrogenase or putrescine transaminase (see FIG. 63, steps J/K-L-C-D/E).
[0220] The invention provides in another embodiment a method for producing putrescine by culturing a non-naturally occurring microbial organism, comprising a putrescine pathway comprising at least one exogenous nucleic acid encoding a putrescine pathway enzyme expressed in a sufficient amount to produce putrescine, the putrescine pathway comprising alpha-ketoglutarate reductase; 5-amino-2-oxopentanoate dehydrogenase or 5-amino-2-oxopentanoate transaminase; 5-amino-2-oxopentanoate decarboxylase; and putrescine dehydrogenase or putrescine transaminase (see FIG. 63, steps O-P/Q-R-D/E). Also provided is a method for producing putrescine by culturing a non-naturally occurring microbial organism, comprising a putrescine pathway comprising at least one exogenous nucleic acid encoding a putrescine pathway enzyme expressed in a sufficient amount to produce putrescine, the putrescine pathway comprising alpha-ketoglutarate reductase; 5-amino-2-oxopentanoate dehydrogenase or 5-amino-2-oxopentanoate transaminase; ornithine dehydrogenase or ornithine transaminase; and ornithine decarboxylase (see FIG. 63, steps O-P/Q-S/T-U). It is understood that a microbial organism comprising any of the pathways disclosed herein can be used to produce a desired product or intermediate, including 4-HB, 4-HBal, BDO or putrescine.
[0221] It is understood that, in methods of the invention, any of the one or more exogenous nucleic acids can be introduced into a microbial organism to produce a non-naturally occurring microbial organism of the invention. The nucleic acids can be introduced so as to confer, for example, a 4-HB, BDO, THF or GBL biosynthetic pathway onto the microbial organism. Alternatively, encoding nucleic acids can be introduced to produce an intermediate microbial organism having the biosynthetic capability to catalyze some of the required reactions to confer 4-HB, BDO, THF or GBL biosynthetic capability. For example, a non-naturally occurring microbial organism having a 4-HB biosynthetic pathway can comprise at least two exogenous nucleic acids encoding desired enzymes, such as the combination of 4-hydroxybutanoate dehydrogenase and .alpha.-ketoglutarate decarboxylase; 4-hydroxybutanoate dehydrogenase and CoA-independent succinic semialdehyde dehydrogenase; 4-hydroxybutanoate dehydrogenase and CoA-dependent succinic semialdehyde dehydrogenase; CoA-dependent succinic semialdehyde dehydrogenase and succinyl-CoA synthetase; succinyl-CoA synthetase and glutamate decarboxylase, and the like. Thus, it is understood that any combination of two or more enzymes of a biosynthetic pathway can be included in a non-naturally occurring microbial organism of the invention. Similarly, it is understood that any combination of three or more enzymes of a biosynthetic pathway can be included in a non-naturally occurring microbial organism of the invention, for example, 4-hydroxybutanoate dehydrogenase, .alpha.-ketoglutarate decarboxylase and CoA-dependent succinic semialdehyde dehydrogenase; CoA-independent succinic semialdehyde dehydrogenase and succinyl-CoA synthetase; 4-hydroxybutanoate dehydrogenase, CoA-dependent succinic semialdehyde dehydrogenase and glutamate:succinic semialdehyde transaminase, and so forth, as desired, so long as the combination of enzymes of the desired biosynthetic pathway results in production of the corresponding desired product.
[0222] Similarly, for example, with respect to any one or more exogenous nucleic acids introduced to confer BDO production, a non-naturally occurring microbial organism having a BDO biosynthetic pathway can comprise at least two exogenous nucleic acids encoding desired enzymes, such as the combination of 4-hydroxybutanoate dehydrogenase and .alpha.-ketoglutarate decarboxylase; 4-hydroxybutanoate dehydrogenase and 4-hydroxybutyryl CoA:acetyl-CoA transferase; 4-hydroxybutanoate dehydrogenase and butyrate kinase; 4-hydroxybutanoate dehydrogenase and phosphotransbutyrylase; 4-hydroxybutyryl CoA:acetyl-CoA transferase and aldehyde dehydrogenase; 4-hydroxybutyryl CoA:acetyl-CoA transferase and alcohol dehydrogenase; 4-hydroxybutyryl CoA:acetyl-CoA transferase and an aldehyde/alcohol dehydrogenase, 4-aminobutyrate-CoA transferase and 4-aminobutyryl-CoA transaminase; 4-aminobutyrate kinase and 4-aminobutan-1-ol oxidoreductase (deaminating), and the like. Thus, it is understood that any combination of two or more enzymes of a biosynthetic pathway can be included in a non-naturally occurring microbial organism of the invention. Similarly, it is understood that any combination of three or more enzymes of a biosynthetic pathway can be included in a non-naturally occurring microbial organism of the invention, for example, 4-hydroxybutanoate dehydrogenase, .alpha.-ketoglutarate decarboxylase and 4-hydroxybutyryl CoA:acetyl-CoA transferase; 4-hydroxybutanoate dehydrogenase, butyrate kinase and phosphotransbutyrylase; 4-hydroxybutanoate dehydrogenase, 4-hydroxybutyryl CoA:acetyl-CoA transferase and aldehyde dehydrogenase; 4-hydroxybutyryl CoA:acetyl-CoA transferase, aldehyde dehydrogenase and alcohol dehydrogenase; butyrate kinase, phosphotransbutyrylase and an aldehyde/alcohol dehydrogenase; 4-aminobutyryl-CoA hydrolase, 4-aminobutyryl-CoA reductase and 4-amino butan-1-ol transaminase; 3-hydroxybutyryl-CoA dehydrogenase, 3-hydroxybutyryl-CoA dehydratase and 4-hydroxybutyryl-CoA dehydratase, and the like. Similarly, any combination of four, five or more enzymes of a biosynthetic pathway as disclosed herein can be included in a non-naturally occurring microbial organism of the invention, as desired, so long as the combination of enzymes of the desired biosynthetic pathway results in production of the corresponding desired product.
[0223] Any of the non-naturally occurring microbial organisms described herein can be cultured to produce and/or secrete the biosynthetic products of the invention. For example, the 4-HB producers can be cultured for the biosynthetic production of 4-HB. The 4-HB can be isolated or be treated as described below to generate GBL, THF and/or BDO. Similarly, the BDO producers can be cultured for the biosynthetic production of BDO. The BDO can be isolated or subjected to further treatments for the chemical synthesis of BDO family compounds, as disclosed herein.
[0224] The growth medium can include, for example, any carbohydrate source which can supply a source of carbon to the non-naturally occurring microorganism. Such sources include, for example, sugars such as glucose, sucrose, xylose, arabinose, galactose, mannose, fructose and starch. Other sources of carbohydrate include, for example, renewable feedstocks and biomass. Exemplary types of biomasses that can be used as feedstocks in the methods of the invention include cellulosic biomass, hemicellulosic biomass and lignin feedstocks or portions of feedstocks. Such biomass feedstocks contain, for example, carbohydrate substrates useful as carbon sources such as glucose, sucrose, xylose, arabinose, galactose, mannose, fructose and starch. Given the teachings and guidance provided herein, those skilled in the art will understand that renewable feedstocks and biomass other than those exemplified above also can be used for culturing the microbial organisms of the invention for the production of 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine and other compounds of the invention.
[0225] Accordingly, given the teachings and guidance provided herein, those skilled in the art will understand that a non-naturally occurring microbial organism can be produced that secretes the biosynthesized compounds of the invention when grown on a carbon source such as a carbohydrate. Such compounds include, for example, 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine and any of the intermediates metabolites in the 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathways and/or the combined 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathways. All that is required is to engineer in one or more of the enzyme activities shown in FIG. 1 to achieve biosynthesis of the desired compound or intermediate including, for example, inclusion of some or all of the 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine biosynthetic pathways. Accordingly, the invention provides a non-naturally occurring microbial organism that secretes 4-HB when grown on a carbohydrate, secretes BDO when grown on a carbohydrate and/or secretes any of the intermediate metabolites shown in FIG. 1, 8-13, 58, 62 or 63 when grown on a carbohydrate. A BDO producing microbial organisms of the invention can initiate synthesis from, for example, succinate, succinyl-CoA, .alpha.-ketogluterate, succinic semialdehyde, 4-HB, 4-hydroxybutyrylphosphate, 4-hydroxybutyryl-CoA (4-HB-CoA) and/or 4-hydroxybutyraldehyde.
[0226] In some embodiments, culture conditions include anaerobic or substantially anaerobic growth or maintenance conditions. Exemplary anaerobic conditions have been described previously and are well known in the art. Exemplary anaerobic conditions for fermentation processes are described below in the Examples. Any of these conditions can be employed with the non-naturally occurring microbial organisms as well as other anaerobic conditions well known in the art. Under such anaerobic conditions, the 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine producers can synthesize monomeric 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine, respectively, at intracellular concentrations of 5-10 mM or more as well as all other concentrations exemplified previously.
[0227] A number of downstream compounds also can be generated for the 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine producing non-naturally occurring microbial organisms of the invention. With respect to the 4-HB producing microbial organisms of the invention, monomeric 4-HB and GBL exist in equilibrium in the culture medium. The conversion of 4-HB to GBL can be efficiently accomplished by, for example, culturing the microbial organisms in acid pH medium. A pH less than or equal to 7.5, in particular at or below pH 5.5, spontaneously converts 4-HB to GBL.
[0228] The resultant GBL can be separated from 4-HB and other components in the culture using a variety of methods well known in the art. Such separation methods include, for example, the extraction procedures exemplified in the Examples as well as methods which include continuous liquid-liquid extraction, pervaporation, membrane filtration, membrane separation, reverse osmosis, electrodialysis, distillation, crystallization, centrifugation, extractive filtration, ion exchange chromatography, size exclusion chromatography, adsorption chromatography, and ultrafiltration. All of the above methods are well known in the art. Separated GBL can be further purified by, for example, distillation.
[0229] Another down stream compound that can be produced from the 4-HB producing non-naturally occurring microbial organisms of the invention includes, for example, BDO. This compound can be synthesized by, for example, chemical hydrogenation of GBL. Chemical hydrogenation reactions are well known in the art. One exemplary procedure includes the chemical reduction of 4-HB and/or GBL or a mixture of these two components deriving from the culture using a heterogeneous or homogeneous hydrogenation catalyst together with hydrogen, or a hydride-based reducing agent used stoichiometrically or catalytically, to produce 1,4-butanediol.
[0230] Other procedures well known in the art are equally applicable for the above chemical reaction and include, for example, WO No. 82/03854 (Bradley, et al.), which describes the hydrogenolysis of gamma-butyrolactone in the vapor phase over a copper oxide and zinc oxide catalyst. British Pat. No. 1,230,276, which describes the hydrogenation of gamma-butyrolactone using a copper oxide-chromium oxide catalyst. The hydrogenation is carried out in the liquid phase. Batch reactions also are exemplified having high total reactor pressures. Reactant and product partial pressures in the reactors are well above the respective dew points. British Pat. No. 1,314,126, which describes the hydrogenation of gamma-butyrolactone in the liquid phase over a nickel-cobalt-thorium oxide catalyst. Batch reactions are exemplified as having high total pressures and component partial pressures well above respective component dew points. British Pat. No. 1,344,557, which describes the hydrogenation of gamma-butyrolactone in the liquid phase over a copper oxide-chromium oxide catalyst. A vapor phase or vapor-containing mixed phase is indicated as suitable in some instances. A continuous flow tubular reactor is exemplified using high total reactor pressures. British Pat. No. 1,512,751, which describes the hydrogenation of gamma-butyrolactone to 1,4-butanediol in the liquid phase over a copper oxide-chromium oxide catalyst. Batch reactions are exemplified with high total reactor pressures and, where determinable, reactant and product partial pressures well above the respective dew points. U.S. Pat. No. 4,301,077, which describes the hydrogenation to 1,4-butanediol of gamma-butyrolactone over a Ru--Ni--Co--Zn catalyst. The reaction can be conducted in the liquid or gas phase or in a mixed liquid-gas phase. Exemplified are continuous flow liquid phase reactions at high total reactor pressures and relatively low reactor productivities. U.S. Pat. No. 4,048,196, which describes the production of 1,4-butanediol by the liquid phase hydrogenation of gamma-butyrolactone over a copper oxide-zinc oxide catalyst. Further exemplified is a continuous flow tubular reactor operating at high total reactor pressures and high reactant and product partial pressures. And U.S. Pat. No. 4,652,685, which describes the hydrogenation of lactones to glycols.
[0231] A further downstream compound that can be produced form the 4-HB producing microbial organisms of the invention includes, for example, THF. This compound can be synthesized by, for example, chemical hydrogenation of GBL. One exemplary procedure well known in the art applicable for the conversion of GBL to THF includes, for example, chemical reduction of 4-HB and/or GBL or a mixture of these two components deriving from the culture using a heterogeneous or homogeneous hydrogenation catalyst together with hydrogen, or a hydride-based reducing agent used stoichiometrically or catalytically, to produce tetrahydrofuran. Other procedures well know in the art are equally applicable for the above chemical reaction and include, for example, U.S. Pat. No. 6,686,310, which describes high surface area sol-gel route prepared hydrogenation catalysts. Processes for the reduction of maleic acid to tetrahydrofuran (THF) and 1,4-butanediol (BDO) and for the reduction of gamma butyrolactone to tetrahydrofuran and 1,4-butanediol also are described.
[0232] The culture conditions can include, for example, liquid culture procedures as well as fermentation and other large scale culture procedures. As described further below in the Examples, particularly useful yields of the biosynthetic products of the invention can be obtained under anaerobic or substantially anaerobic culture conditions.
[0233] Suitable purification and/or assays to test for the production of 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine can be performed using well known methods. Suitable replicates such as triplicate cultures can be grown for each engineered strain to be tested. For example, product and byproduct formation in the engineered production host can be monitored. The final product and intermediates, and other organic compounds, can be analyzed by methods such as HPLC (High Performance Liquid Chromatography), GC-MS (Gas Chromatography-Mass Spectroscopy) and LC-MS (Liquid Chromatography-Mass Spectroscopy) or other suitable analytical methods using routine procedures well known in the art. The release of product in the fermentation broth can also be tested with the culture supernatant. Byproducts and residual glucose can be quantified by HPLC using, for example, a refractive index detector for glucose and alcohols, and a UV detector for organic acids (Lin et al., Biotechnol. Bioeng. 90:775-779 (2005)), or other suitable assay and detection methods well known in the art. The individual enzyme or protein activities from the exogenous DNA sequences can also be assayed using methods well known in the art.
[0234] The 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine product can be separated from other components in the culture using a variety of methods well known in the art. Such separation methods include, for example, extraction procedures as well as methods that include continuous liquid-liquid extraction, pervaporation, membrane filtration, membrane separation, reverse osmosis, electrodialysis, distillation, crystallization, centrifugation, extractive filtration, ion exchange chromatography, size exclusion chromatography, adsorption chromatography, and ultrafiltration. All of the above methods are well known in the art.
[0235] The invention further provides a method of manufacturing 4-HB. The method includes fermenting a non-naturally occurring microbial organism having a 4-hydroxybutanoic acid (4-HB) biosynthetic pathway comprising at least one exogenous nucleic acid encoding 4-hydroxybutanoate dehydrogenase, CoA-independent succinic semialdehyde dehydrogenase, succinyl-CoA synthetase, CoA-dependent succinic semialdehyde dehydrogenase, glutamate: succinic semialdehyde transaminase, .alpha.-ketoglutarate decarboxylase, or glutamate decarboxylase under substantially anaerobic conditions for a sufficient period of time to produce monomeric 4-hydroxybutanoic acid (4-HB), the process comprising fed-batch fermentation and batch separation; fed-batch fermentation and continuous separation, or continuous fermentation and continuous separation.
[0236] The culture and chemical hydrogenations described above also can be scaled up and grown continuously for manufacturing of 4-HB, 4-HBal, 4-HBCoA, GBL, BDO and/or THF or putrescine. Exemplary growth procedures include, for example, fed-batch fermentation and batch separation; fed-batch fermentation and continuous separation, or continuous fermentation and continuous separation. All of these processes are well known in the art. Employing the 4-HB producers allows for simultaneous 4-HB biosynthesis and chemical conversion to GBL, BDO and/or THF by employing the above hydrogenation procedures simultaneous with continuous cultures methods such as fermentation. Other hydrogenation procedures also are well known in the art and can be equally applied to the methods of the invention.
[0237] Fermentation procedures are particularly useful for the biosynthetic production of commercial quantities of 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine. Generally, and as with non-continuous culture procedures, the continuous and/or near-continuous production of 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine will include culturing a non-naturally occurring 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine producing organism of the invention in sufficient nutrients and medium to sustain and/or nearly sustain growth in an exponential phase. Continuous culture under such conditions can be include, for example, 1 day, 2, 3, 4, 5, 6 or 7 days or more. Additionally, continuous culture can include 1 week, 2, 3, 4 or 5 or more weeks and up to several months. Alternatively, organisms of the invention can be cultured for hours, if suitable for a particular application. It is to be understood that the continuous and/or near-continuous culture conditions also can include all time intervals in between these exemplary periods. It is further understood that the time of culturing the microbial organism of the invention is for a sufficient period of time to produce a sufficient amount of product for a desired purpose.
[0238] Fermentation procedures are well known in the art. Briefly, fermentation for the biosynthetic production of 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine or other 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine derived products, including intermediates, of the invention can be utilized in, for example, fed-batch fermentation and batch separation; fed-batch fermentation and continuous separation, or continuous fermentation and continuous separation. Examples of batch and continuous fermentation procedures well known in the art are exemplified further below in the Examples.
[0239] In addition, to the above fermentation procedures using the 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine producers of the invention for continuous production of substantial quantities of 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine, including monomeric 4-HB, respectively, the 4-HB producers also can be, for example, simultaneously subjected to chemical synthesis procedures as described previously for the chemical conversion of monomeric 4-HB to, for example, GBL, BDO and/or THF. The BDO producers can similarly be, for example, simultaneously subjected to chemical synthesis procedures as described previously for the chemical conversion of BDO to, for example, THF, GBL, pyrrolidones and/or other BDO family compounds. In addition, the products of the 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine producers can be separated from the fermentation culture and sequentially subjected to chemical conversion, as disclosed herein.
[0240] Briefly, hydrogenation of GBL in the fermentation broth can be performed as described by Frost et al., Biotechnology Progress 18: 201-211 (2002). Another procedure for hydrogenation during fermentation include, for example, the methods described in, for example, U.S. Pat. No. 5,478,952. This method is further exemplified in the Examples below.
[0241] Therefore, the invention additionally provides a method of manufacturing .gamma.-butyrolactone (GBL), tetrahydrofuran (THF) or 1,4-butanediol (BDO). The method includes fermenting a non-naturally occurring microbial organism having 4-hydroxybutanoic acid (4-HB) and/or 1,4-butanediol (BDO) biosynthetic pathways, the pathways comprise at least one exogenous nucleic acid encoding 4-hydroxybutanoate dehydrogenase, CoA-independent succinic semialdehyde dehydrogenase, succinyl-CoA synthetase, CoA-dependent succinic semialdehyde dehydrogenase, 4-hydroxybutyrate:CoA transferase, glutamate: succinic semialdehyde transaminase, .alpha.-ketoglutarate decarboxylase, glutamate decarboxylase, 4-hydroxybutanoate kinase, phosphotransbutyrylase, CoA-independent 1,4-butanediol semialdehyde dehydrogenase, CoA-dependent 1,4-butanediol semialdehyde dehydrogenase, CoA-independent 1,4-butanediol alcohol dehydrogenase or CoA-dependent 1,4-butanediol alcohol dehydrogenase, under substantially anaerobic conditions for a sufficient period of time to produce 1,4-butanediol (BDO), GBL or THF, the fermenting comprising fed-batch fermentation and batch separation; fed-batch fermentation and continuous separation, or continuous fermentation and continuous separation.
[0242] In addition to the biosynthesis of 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine and other products of the invention as described herein, the non-naturally occurring microbial organisms and methods of the invention also can be utilized in various combinations with each other and with other microbial organisms and methods well known in the art to achieve product biosynthesis by other routes. For example, one alternative to produce BDO other than use of the 4-HB producers and chemical steps or other than use of the BDO producer directly is through addition of another microbial organism capable of converting 4-HB or a 4-HB product exemplified herein to BDO.
[0243] One such procedure includes, for example, the fermentation of a 4-HB producing microbial organism of the invention to produce 4-HB, as described above and below. The 4-HB can then be used as a substrate for a second microbial organism that converts 4-HB to, for example, BDO, GBL and/or THF. The 4-HB can be added directly to another culture of the second organism or the original culture of 4-HB producers can be depleted of these microbial organisms by, for example, cell separation, and then subsequent addition of the second organism to the fermentation broth can utilized to produce the final product without intermediate purification steps. One exemplary second organism having the capacity to biochemically utilize 4-HB as a substrate for conversion to BDO, for example, is Clostridium acetobutylicum (see, for example, Jewell et al., Current Microbiology, 13:215-19 (1986)).
[0244] Thus, such a procedure includes, for example, the fermentation of a microbial organism that produces a 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway intermediate. The 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway intermediate can then be used as a substrate for a second microbial organism that converts the 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway intermediate to 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine. The 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway intermediate can be added directly to another culture of the second organism or the original culture of the 4-HB, 4-HBal, 4-HBCoA BDO or putrescine pathway intermediate producers can be depleted of these microbial organisms by, for example, cell separation, and then subsequent addition of the second organism to the fermentation broth can be utilized to produce the final product without intermediate purification steps.
[0245] In other embodiments, the non-naturally occurring microbial organisms and methods of the invention can be assembled in a wide variety of subpathways to achieve biosynthesis of, for example, 4-HB and/or BDO as described. In these embodiments, biosynthetic pathways for a desired product of the invention can be segregated into different microbial organisms and the different microbial organisms can be co-cultured to produce the final product. In such a biosynthetic scheme, the product of one microbial organism is the substrate for a second microbial organism until the final product is synthesized. For example, the biosynthesis of BDO can be accomplished as described previously by constructing a microbial organism that contains biosynthetic pathways for conversion of one pathway intermediate to another pathway intermediate or the product, for example, a substrate such as endogenous succinate through 4-HB to the final product BDO. Alternatively, BDO also can be biosynthetically produced from microbial organisms through co-culture or co-fermentation using two organisms in the same vessel. A first microbial organism being a 4-HB producer with genes to produce 4-HB from succinic acid, and a second microbial organism being a BDO producer with genes to convert 4-HB to BDO. For example, the biosynthesis of 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine can be accomplished by constructing a microbial organism that contains biosynthetic pathways for conversion of one pathway intermediate to another pathway intermediate or the product. Alternatively, 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine also can be biosynthetically produced from microbial organisms through co-culture or co-fermentation using two organisms in the same vessel, where the first microbial organism produces a 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine intermediate and the second microbial organism converts the intermediate to 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine.
[0246] Given the teachings and guidance provided herein, those skilled in the art will understand that a wide variety of combinations and permutations exist for the non-naturally occurring microbial organisms and methods of the invention together with other microbial organisms, with the co-culture of other non-naturally occurring microbial organisms having subpathways and with combinations of other chemical and/or biochemical procedures well known in the art to produce 4-HB, BDO, GBL and THF products of the invention.
[0247] It is understood that, in methods of the invention, any of the one or more exogenous nucleic acids can be introduced into a microbial organism to produce a non-naturally occurring microbial organism of the invention. The nucleic acids can be introduced so as to confer, for example, a 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine biosynthetic pathway onto the microbial organism. Alternatively, encoding nucleic acids can be introduced to produce an intermediate microbial organism having the biosynthetic capability to catalyze some of the required reactions to confer 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine biosynthetic capability. For example, a non-naturally occurring microbial organism having a 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine biosynthetic pathway can comprise at least two exogenous nucleic acids encoding desired enzymes or proteins, such as the combination of enzymes as disclosed herein (see Examples and FIGS. 1, 8-13, 58, 62 and 63), and the like. Thus, it is understood that any combination of two or more enzymes or proteins of a biosynthetic pathway can be included in a non-naturally occurring microbial organism of the invention. Similarly, it is understood that any combination of three or more enzymes or proteins of a biosynthetic pathway can be included in a non-naturally occurring microbial organism of the invention, for example,], and so forth, as desired and disclosed herein, so long as the combination of enzymes and/or proteins of the desired biosynthetic pathway results in production of the corresponding desired product. Similarly, any combination of four or more enzymes or proteins of a biosynthetic pathway as disclosed herein can be included in a non-naturally occurring microbial organism of the invention, as desired, so long as the combination of enzymes and/or proteins of the desired biosynthetic pathway results in production of the corresponding desired product.
[0248] To generate better producers, metabolic modeling can be utilized to optimize growth conditions. Modeling can also be used to design gene knockouts that additionally optimize utilization of the pathway (see, for example, U.S. patent publications US 2002/0012939, US 2003/0224363, US 2004/0029149, US 2004/0072723, US 2003/0059792, US 2002/0168654 and US 2004/0009466, and U.S. Pat. No. 7,127,379). Modeling analysis allows reliable predictions of the effects on cell growth of shifting the metabolism towards more efficient production of 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine.
[0249] One computational method for identifying and designing metabolic alterations favoring biosynthesis of a desired product is the OptKnock computational framework (Burgard et al., Biotechnol. Bioeng. 84:647-657 (2003)). OptKnock is a metabolic modeling and simulation program that suggests gene deletion or disruption strategies that result in genetically stable microorganisms which overproduce the target product. Specifically, the framework examines the complete metabolic and/or biochemical network of a microorganism in order to suggest genetic manipulations that force the desired biochemical to become an obligatory byproduct of cell growth. By coupling biochemical production with cell growth through strategically placed gene deletions or other functional gene disruption, the growth selection pressures imposed on the engineered strains after long periods of time in a bioreactor lead to improvements in performance as a result of the compulsory growth-coupled biochemical production. Lastly, when gene deletions are constructed there is a negligible possibility of the designed strains reverting to their wild-type states because the genes selected by OptKnock are to be completely removed from the genome. Therefore, this computational methodology can be used to either identify alternative pathways that lead to biosynthesis of a desired product or used in connection with the non-naturally occurring microbial organisms for further optimization of biosynthesis of a desired product.
[0250] Briefly, OptKnock is a term used herein to refer to a computational method and system for modeling cellular metabolism. The OptKnock program relates to a framework of models and methods that incorporate particular constraints into flux balance analysis (FBA) models. These constraints include, for example, qualitative kinetic information, qualitative regulatory information, and/or DNA microarray experimental data. OptKnock also computes solutions to various metabolic problems by, for example, tightening the flux boundaries derived through flux balance models and subsequently probing the performance limits of metabolic networks in the presence of gene additions or deletions. OptKnock computational framework allows the construction of model formulations that allow an effective query of the performance limits of metabolic networks and provides methods for solving the resulting mixed-integer linear programming problems. The metabolic modeling and simulation methods referred to herein as OptKnock are described in, for example, U.S. publication 2002/0168654, filed Jan. 10, 2002, in International Patent No. PCT/US02/00660, filed Jan. 10, 2002, and U.S. publication 2009/0047719, filed Aug. 10, 2007.
[0251] Another computational method for identifying and designing metabolic alterations favoring biosynthetic production of a product is a metabolic modeling and simulation system termed SimPheny.RTM.. This computational method and system is described in, for example, U.S. publication 2003/0233218, filed Jun. 14, 2002, and in International Patent Application No. PCT/US03/18838, filed Jun. 13, 2003. SimPheny.RTM. is a computational system that can be used to produce a network model in silico and to simulate the flux of mass, energy or charge through the chemical reactions of a biological system to define a solution space that contains any and all possible functionalities of the chemical reactions in the system, thereby determining a range of allowed activities for the biological system. This approach is referred to as constraints-based modeling because the solution space is defined by constraints such as the known stoichiometry of the included reactions as well as reaction thermodynamic and capacity constraints associated with maximum fluxes through reactions. The space defined by these constraints can be interrogated to determine the phenotypic capabilities and behavior of the biological system or of its biochemical components.
[0252] These computational approaches are consistent with biological realities because biological systems are flexible and can reach the same result in many different ways. Biological systems are designed through evolutionary mechanisms that have been restricted by fundamental constraints that all living systems must face. Therefore, constraints-based modeling strategy embraces these general realities. Further, the ability to continuously impose further restrictions on a network model via the tightening of constraints results in a reduction in the size of the solution space, thereby enhancing the precision with which physiological performance or phenotype can be predicted.
[0253] Given the teachings and guidance provided herein, those skilled in the art will be able to apply various computational frameworks for metabolic modeling and simulation to design and implement biosynthesis of a desired compound in host microbial organisms. Such metabolic modeling and simulation methods include, for example, the computational systems exemplified above as SimPheny.RTM. and OptKnock. For illustration of the invention, some methods are described herein with reference to the OptKnock computation framework for modeling and simulation. Those skilled in the art will know how to apply the identification, design and implementation of the metabolic alterations using OptKnock to any of such other metabolic modeling and simulation computational frameworks and methods well known in the art.
[0254] The methods described above will provide one set of metabolic reactions to disrupt. Elimination of each reaction within the set or metabolic modification can result in a desired product as an obligatory product during the growth phase of the organism. Because the reactions are known, a solution to the bilevel OptKnock problem also will provide the associated gene or genes encoding one or more enzymes that catalyze each reaction within the set of reactions. Identification of a set of reactions and their corresponding genes encoding the enzymes participating in each reaction is generally an automated process, accomplished through correlation of the reactions with a reaction database having a relationship between enzymes and encoding genes.
[0255] Once identified, the set of reactions that are to be disrupted in order to achieve production of a desired product are implemented in the target cell or organism by functional disruption of at least one gene encoding each metabolic reaction within the set. One particularly useful means to achieve functional disruption of the reaction set is by deletion of each encoding gene. However, in some instances, it can be beneficial to disrupt the reaction by other genetic aberrations including, for example, mutation, deletion of regulatory regions such as promoters or cis binding sites for regulatory factors, or by truncation of the coding sequence at any of a number of locations. These latter aberrations, resulting in less than total deletion of the gene set can be useful, for example, when rapid assessments of the coupling of a product are desired or when genetic reversion is less likely to occur.
[0256] To identify additional productive solutions to the above described bilevel OptKnock problem which lead to further sets of reactions to disrupt or metabolic modifications that can result in the biosynthesis, including growth-coupled biosynthesis of a desired product, an optimization method, termed integer cuts, can be implemented. This method proceeds by iteratively solving the OptKnock problem exemplified above with the incorporation of an additional constraint referred to as an integer cut at each iteration. Integer cut constraints effectively prevent the solution procedure from choosing the exact same set of reactions identified in any previous iteration that obligatorily couples product biosynthesis to growth. For example, if a previously identified growth-coupled metabolic modification specifies reactions 1, 2, and 3 for disruption, then the following constraint prevents the same reactions from being simultaneously considered in subsequent solutions. The integer cut method is well known in the art and can be found described in, for example, Burgard et al., Biotechnol. Prog. 17:791-797 (2001). As with all methods described herein with reference to their use in combination with the OptKnock computational framework for metabolic modeling and simulation, the integer cut method of reducing redundancy in iterative computational analysis also can be applied with other computational frameworks well known in the art including, for example, SimPheny.RTM..
[0257] The methods exemplified herein allow the construction of cells and organisms that biosynthetically produce a desired product, including the obligatory coupling of production of a target biochemical product to growth of the cell or organism engineered to harbor the identified genetic alterations. Therefore, the computational methods described herein allow the identification and implementation of metabolic modifications that are identified by an in silico method selected from OptKnock or SimPheny.RTM.. The set of metabolic modifications can include, for example, addition of one or more biosynthetic pathway enzymes and/or functional disruption of one or more metabolic reactions including, for example, disruption by gene deletion.
[0258] As discussed above, the OptKnock methodology was developed on the premise that mutant microbial networks can be evolved towards their computationally predicted maximum-growth phenotypes when subjected to long periods of growth selection. In other words, the approach leverages an organism's ability to self-optimize under selective pressures. The OptKnock framework allows for the exhaustive enumeration of gene deletion combinations that force a coupling between biochemical production and cell growth based on network stoichiometry. The identification of optimal gene/reaction knockouts requires the solution of a bilevel optimization problem that chooses the set of active reactions such that an optimal growth solution for the resulting network overproduces the biochemical of interest (Burgard et al., Biotechnol. Bioeng. 84:647-657 (2003)).
[0259] An in silico stoichiometric model of E. coli metabolism can be employed to identify essential genes for metabolic pathways as exemplified previously and described in, for example, U.S. patent publications US 2002/0012939, US 2003/0224363, US 2004/0029149, US 2004/0072723, US 2003/0059792, US 2002/0168654 and US 2004/0009466, and in U.S. Pat. No. 7,127,379. As disclosed herein, the OptKnock mathematical framework can be applied to pinpoint gene deletions leading to the growth-coupled production of a desired product. Further, the solution of the bilevel OptKnock problem provides only one set of deletions. To enumerate all meaningful solutions, that is, all sets of knockouts leading to growth-coupled production formation, an optimization technique, termed integer cuts, can be implemented. This entails iteratively solving the OptKnock problem with the incorporation of an additional constraint referred to as an integer cut at each iteration, as discussed above.
[0260] The methods exemplified above and further illustrated in the Examples below allow the construction of cells and organisms that biosynthetically produce, including obligatory couple production of a target biochemical product to growth of the cell or organism engineered to harbor the identified genetic alterations. In this regard, metabolic alterations have been identified that result in the biosynthesis of 4-HB and 1,4-butanediol. Microorganism strains constructed with the identified metabolic alterations produce elevated levels of 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine compared to unmodified microbial organisms. These strains can be beneficially used for the commercial production of 4-HB, BDO, THF, GBL, 4-HBal, 4-HBCoA or putrescine, for example, in continuous fermentation process without being subjected to the negative selective pressures.
[0261] Therefore, the computational methods described herein allow the identification and implementation of metabolic modifications that are identified by an in silico method selected from OptKnock or SimPheny.RTM.. The set of metabolic modifications can include, for example, addition of one or more biosynthetic pathway enzymes and/or functional disruption of one or more metabolic reactions including, for example, disruption by gene deletion.
[0262] It is understood that modifications which do not substantially affect the activity of the various embodiments of this invention are also included within the definition of the invention provided herein. Accordingly, the following examples are intended to illustrate but not limit the present invention.
[0263] Any of the non-naturally occurring microbial organisms described herein can be cultured to produce and/or secrete the biosynthetic products of the invention. For example, the 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine producers can be cultured for the biosynthetic production of 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine.
[0264] For the production of 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine, the recombinant strains are cultured in a medium with carbon source and other essential nutrients. It is highly desirable to maintain anaerobic conditions in the fermenter to reduce the cost of the overall process. Such conditions can be obtained, for example, by first sparging the medium with nitrogen and then sealing the flasks with a septum and crimp-cap. For strains where growth is not observed anaerobically, microaerobic conditions can be applied by perforating the septum with a small hole for limited aeration. Exemplary anaerobic conditions have been described previously and are well-known in the art. Exemplary aerobic and anaerobic conditions are described, for example, in U.S. publication 2009/0047719, filed Aug. 10, 2007. Fermentations can be performed in a batch, fed-batch or continuous manner, as disclosed herein.
[0265] If desired, the pH of the medium can be maintained at a desired pH, in particular neutral pH, such as a pH of around 7 by addition of a base, such as NaOH or other bases, or acid, as needed to maintain the culture medium at a desirable pH. The growth rate can be determined by measuring optical density using a spectrophotometer (600 nm), and the glucose uptake rate by monitoring carbon source depletion over time.
[0266] In addition to renewable feedstocks such as those exemplified above, the 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine producing microbial organisms of the invention also can be modified for growth on syngas as its source of carbon. In this specific embodiment, one or more proteins or enzymes are expressed in the 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine producing organisms to provide a metabolic pathway for utilization of syngas or other gaseous carbon source.
[0267] Synthesis gas, also known as syngas or producer gas, is the major product of gasification of coal and of carbonaceous materials such as biomass materials, including agricultural crops and residues. Syngas is a mixture primarily of H.sub.2 and CO and can be obtained from the gasification of any organic feedstock, including but not limited to coal, coal oil, natural gas, biomass, and waste organic matter. Gasification is generally carried out under a high fuel to oxygen ratio. Although largely H.sub.2 and CO, syngas can also include CO.sub.2 and other gases in smaller quantities. Thus, synthesis gas provides a cost effective source of gaseous carbon such as CO and, additionally, CO.sub.2.
[0268] The Wood-Ljungdahl pathway catalyzes the conversion of CO and H.sub.2 to acetyl-CoA and other products such as acetate. Organisms capable of utilizing CO and syngas also generally have the capability of utilizing CO.sub.2 and CO.sub.2/H.sub.2 mixtures through the same basic set of enzymes and transformations encompassed by the Wood-Ljungdahl pathway. H.sub.2-dependent conversion of CO.sub.2 to acetate by microorganisms was recognized long before it was revealed that CO also could be used by the same organisms and that the same pathways were involved. Many acetogens have been shown to grow in the presence of CO.sub.2 and produce compounds such as acetate as long as hydrogen is present to supply the necessary reducing equivalents (see for example, Drake, Acetogenesis, pp. 3-60 Chapman and Hall, New York, (1994)). This can be summarized by the following equation:
2CO.sub.2+4H.sub.2+nADP+nPi.fwdarw.CH.sub.3COOH+2H.sub.2O+nATP
Hence, non-naturally occurring microorganisms possessing the Wood-Ljungdahl pathway can utilize CO.sub.2 and H.sub.2 mixtures as well for the production of acetyl-CoA and other desired products.
[0269] The Wood-Ljungdahl pathway is well known in the art and consists of 12 reactions which can be separated into two branches: (1) methyl branch and (2) carbonyl branch. The methyl branch converts syngas to methyl-tetrahydrofolate (methyl-THF) whereas the carbonyl branch converts methyl-THF to acetyl-CoA. The reactions in the methyl branch are catalyzed in order by the following enzymes or proteins: ferredoxin oxidoreductase, formate dehydrogenase, formyltetrahydrofolate synthetase, methenyltetrahydrofolate cyclodehydratase, methylenetetrahydrofolate dehydrogenase and methylenetetrahydrofolate reductase. The reactions in the carbonyl branch are catalyzed in order by the following enzymes or proteins: methyltetrahydrofolate:corrinoid protein methyltransferase (for example, AcsE), corrinoid iron-sulfur protein, nickel-protein assembly protein (for example, AcsF), ferredoxin, acetyl-CoA synthase, carbon monoxide dehydrogenase and nickel-protein assembly protein (for example, CooC). Following the teachings and guidance provided herein for introducing a sufficient number of encoding nucleic acids to generate a 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway, those skilled in the art will understand that the same engineering design also can be performed with respect to introducing at least the nucleic acids encoding the Wood-Ljungdahl enzymes or proteins absent in the host organism. Therefore, introduction of one or more encoding nucleic acids into the microbial organisms of the invention such that the modified organism contains the complete Wood-Ljungdahl pathway will confer syngas utilization ability.
[0270] Accordingly, given the teachings and guidance provided herein, those skilled in the art will understand that a non-naturally occurring microbial organism can be produced that secretes the biosynthesized compounds of the invention when grown on a carbon source such as a carbohydrate. Such compounds include, for example, 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine and any of the intermediate metabolites in the 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway. All that is required is to engineer in one or more of the required enzyme or protein activities to achieve biosynthesis of the desired compound or intermediate including, for example, inclusion of some or all of the 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine biosynthetic pathways. Accordingly, the invention provides a non-naturally occurring microbial organism that produces and/or secretes 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine when grown on a carbohydrate or other carbon source and produces and/or secretes any of the intermediate metabolites shown in the 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway when grown on a carbohydrate or other carbon source. The 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine producing microbial organisms of the invention can initiate synthesis from an intermediate in a 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway, as disclosed herein.
[0271] To generate better producers, metabolic modeling can be utilized to optimize growth conditions. Modeling can also be used to design gene knockouts that additionally optimize utilization of the pathway (see, for example, U.S. patent publications US 2002/0012939, US 2003/0224363, US 2004/0029149, US 2004/0072723, US 2003/0059792, US 2002/0168654 and US 2004/0009466, and U.S. Pat. No. 7,127,379). Modeling analysis allows reliable predictions of the effects on cell growth of shifting the metabolism towards more efficient production of 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine.
[0272] One computational method for identifying and designing metabolic alterations favoring biosynthesis of a desired product is the OptKnock computational framework (Burgard et al., Biotechnol. Bioeng. 84:647-657 (2003)). OptKnock is a metabolic modeling and simulation program that suggests gene deletion strategies that result in genetically stable microorganisms which overproduce the target product. Specifically, the framework examines the complete metabolic and/or biochemical network of a microorganism in order to suggest genetic manipulations that force the desired biochemical to become an obligatory byproduct of cell growth. By coupling biochemical production with cell growth through strategically placed gene deletions or other functional gene disruption, the growth selection pressures imposed on the engineered strains after long periods of time in a bioreactor lead to improvements in performance as a result of the compulsory growth-coupled biochemical production. Lastly, when gene deletions are constructed there is a negligible possibility of the designed strains reverting to their wild-type states because the genes selected by OptKnock are to be completely removed from the genome. Therefore, this computational methodology can be used to either identify alternative pathways that lead to biosynthesis of a desired product or used in connection with the non-naturally occurring microbial organisms for further optimization of biosynthesis of a desired product.
[0273] Briefly, OptKnock is a term used herein to refer to a computational method and system for modeling cellular metabolism. The OptKnock program relates to a framework of models and methods that incorporate particular constraints into flux balance analysis (FBA) models. These constraints include, for example, qualitative kinetic information, qualitative regulatory information, and/or DNA microarray experimental data. OptKnock also computes solutions to various metabolic problems by, for example, tightening the flux boundaries derived through flux balance models and subsequently probing the performance limits of metabolic networks in the presence of gene additions or deletions. OptKnock computational framework allows the construction of model formulations that allow an effective query of the performance limits of metabolic networks and provides methods for solving the resulting mixed-integer linear programming problems. The metabolic modeling and simulation methods referred to herein as OptKnock are described in, for example, U.S. publication 2002/0168654, filed Jan. 10, 2002, in International Patent No. PCT/US02/00660, filed Jan. 10, 2002, and U.S. publication 2009/0047719, filed Aug. 10, 2007.
[0274] Another computational method for identifying and designing metabolic alterations favoring biosynthetic production of a product is a metabolic modeling and simulation system termed SimPheny.RTM.. This computational method and system is described in, for example, U.S. publication 2003/0233218, filed Jun. 14, 2002, and in International Patent Application No. PCT/US03/18838, filed Jun. 13, 2003. SimPheny.RTM. is a computational system that can be used to produce a network model in silico and to simulate the flux of mass, energy or charge through the chemical reactions of a biological system to define a solution space that contains any and all possible functionalities of the chemical reactions in the system, thereby determining a range of allowed activities for the biological system. This approach is referred to as constraints-based modeling because the solution space is defined by constraints such as the known stoichiometry of the included reactions as well as reaction thermodynamic and capacity constraints associated with maximum fluxes through reactions. The space defined by these constraints can be interrogated to determine the phenotypic capabilities and behavior of the biological system or of its biochemical components.
[0275] These computational approaches are consistent with biological realities because biological systems are flexible and can reach the same result in many different ways. Biological systems are designed through evolutionary mechanisms that have been restricted by fundamental constraints that all living systems must face. Therefore, constraints-based modeling strategy embraces these general realities. Further, the ability to continuously impose further restrictions on a network model via the tightening of constraints results in a reduction in the size of the solution space, thereby enhancing the precision with which physiological performance or phenotype can be predicted.
[0276] Given the teachings and guidance provided herein, those skilled in the art will be able to apply various computational frameworks for metabolic modeling and simulation to design and implement biosynthesis of a desired compound in host microbial organisms. Such metabolic modeling and simulation methods include, for example, the computational systems exemplified above as SimPheny.RTM. and OptKnock. For illustration of the invention, some methods are described herein with reference to the OptKnock computation framework for modeling and simulation. Those skilled in the art will know how to apply the identification, design and implementation of the metabolic alterations using OptKnock to any of such other metabolic modeling and simulation computational frameworks and methods well known in the art.
[0277] The methods described above will provide one set of metabolic reactions to disrupt. Elimination of each reaction within the set or metabolic modification can result in a desired product as an obligatory product during the growth phase of the organism. Because the reactions are known, a solution to the bilevel OptKnock problem also will provide the associated gene or genes encoding one or more enzymes that catalyze each reaction within the set of reactions. Identification of a set of reactions and their corresponding genes encoding the enzymes participating in each reaction is generally an automated process, accomplished through correlation of the reactions with a reaction database having a relationship between enzymes and encoding genes.
[0278] Once identified, the set of reactions that are to be disrupted in order to achieve production of a desired product are implemented in the target cell or organism by functional disruption of at least one gene encoding each metabolic reaction within the set. One particularly useful means to achieve functional disruption of the reaction set is by deletion of each encoding gene. However, in some instances, it can be beneficial to disrupt the reaction by other genetic aberrations including, for example, mutation, deletion of regulatory regions such as promoters or cis binding sites for regulatory factors, or by truncation of the coding sequence at any of a number of locations. These latter aberrations, resulting in less than total deletion of the gene set can be useful, for example, when rapid assessments of the coupling of a product are desired or when genetic reversion is less likely to occur.
[0279] To identify additional productive solutions to the above described bilevel OptKnock problem which lead to further sets of reactions to disrupt or metabolic modifications that can result in the biosynthesis, including growth-coupled biosynthesis of a desired product, an optimization method, termed integer cuts, can be implemented. This method proceeds by iteratively solving the OptKnock problem exemplified above with the incorporation of an additional constraint referred to as an integer cut at each iteration. Integer cut constraints effectively prevent the solution procedure from choosing the exact same set of reactions identified in any previous iteration that obligatorily couples product biosynthesis to growth. For example, if a previously identified growth-coupled metabolic modification specifies reactions 1, 2, and 3 for disruption, then the following constraint prevents the same reactions from being simultaneously considered in subsequent solutions. The integer cut method is well known in the art and can be found described in, for example, Burgard et al., Biotechnol. Prog. 17:791-797 (2001). As with all methods described herein with reference to their use in combination with the OptKnock computational framework for metabolic modeling and simulation, the integer cut method of reducing redundancy in iterative computational analysis also can be applied with other computational frameworks well known in the art including, for example, SimPheny.RTM..
[0280] The methods exemplified herein allow the construction of cells and organisms that biosynthetically produce a desired product, including the obligatory coupling of production of a target biochemical product to growth of the cell or organism engineered to harbor the identified genetic alterations. Therefore, the computational methods described herein allow the identification and implementation of metabolic modifications that are identified by an in silico method selected from OptKnock or SimPheny.RTM.. The set of metabolic modifications can include, for example, addition of one or more biosynthetic pathway enzymes and/or functional disruption of one or more metabolic reactions including, for example, disruption by gene deletion.
[0281] As discussed above, the OptKnock methodology was developed on the premise that mutant microbial networks can be evolved towards their computationally predicted maximum-growth phenotypes when subjected to long periods of growth selection. In other words, the approach leverages an organism's ability to self-optimize under selective pressures. The OptKnock framework allows for the exhaustive enumeration of gene deletion combinations that force a coupling between biochemical production and cell growth based on network stoichiometry. The identification of optimal gene/reaction knockouts requires the solution of a bilevel optimization problem that chooses the set of active reactions such that an optimal growth solution for the resulting network overproduces the biochemical of interest (Burgard et al., Biotechnol. Bioeng. 84:647-657 (2003)).
[0282] An in silico stoichiometric model of E. coli metabolism can be employed to identify essential genes for metabolic pathways as exemplified previously and described in, for example, U.S. patent publications US 2002/0012939, US 2003/0224363, US 2004/0029149, US 2004/0072723, US 2003/0059792, US 2002/0168654 and US 2004/0009466, and in U.S. Pat. No. 7,127,379. As disclosed herein, the OptKnock mathematical framework can be applied to pinpoint gene deletions leading to the growth-coupled production of a desired product. Further, the solution of the bilevel OptKnock problem provides only one set of deletions. To enumerate all meaningful solutions, that is, all sets of knockouts leading to growth-coupled production formation, an optimization technique, termed integer cuts, can be implemented. This entails iteratively solving the OptKnock problem with the incorporation of an additional constraint referred to as an integer cut at each iteration, as discussed above.
[0283] As disclosed herein, a nucleic acid encoding a desired activity of a 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway can be introduced into a host organism. In some cases, it can be desirable to modify an activity of a 4-HB, 4-HBal, 4-HBCoA BDO or putrescine pathway enzyme or protein to increase production of 4-HB, 4-HBal, 4-HBCoA BDO or putrescine. For example, known mutations that increase the activity of a protein or enzyme can be introduced into an encoding nucleic acid molecule. Additionally, optimization methods can be applied to increase the activity of an enzyme or protein and/or decrease an inhibitory activity, for example, decrease the activity of a negative regulator.
[0284] One such optimization method is directed evolution. Directed evolution is a powerful approach that involves the introduction of mutations targeted to a specific gene in order to improve and/or alter the properties of an enzyme. Improved and/or altered enzymes can be identified through the development and implementation of sensitive high-throughput screening assays that allow the automated screening of many enzyme variants (for example, >10.sup.4). Iterative rounds of mutagenesis and screening typically are performed to afford an enzyme with optimized properties. Computational algorithms that can help to identify areas of the gene for mutagenesis also have been developed and can significantly reduce the number of enzyme variants that need to be generated and screened. Numerous directed evolution technologies have been developed (for reviews, see Hibbert et al., Biomol. Eng 22:11-19 (2005); Huisman and Lalonde, In Biocatalysis in the pharmaceutical and biotechnology industries pgs. 717-742 (2007), Patel (ed.), CRC Press; Otten and Quax. Biomol. Eng 22:1-9 (2005); and Sen et al., Appl Biochem. Biotechnol 143:212-223 (2007)) to be effective at creating diverse variant libraries, and these methods have been successfully applied to the improvement of a wide range of properties across many enzyme classes. Enzyme characteristics that have been improved and/or altered by directed evolution technologies include, for example: selectivity/specificity, for conversion of non-natural substrates; temperature stability, for robust high temperature processing; pH stability, for bioprocessing under lower or higher pH conditions; substrate or product tolerance, so that high product titers can be achieved; binding (K.sub.m), including broadening substrate binding to include non-natural substrates; inhibition (K.sub.i), to remove inhibition by products, substrates, or key intermediates; activity (kcat), to increases enzymatic reaction rates to achieve desired flux; expression levels, to increase protein yields and overall pathway flux; oxygen stability, for operation of air sensitive enzymes under aerobic conditions; and anaerobic activity, for operation of an aerobic enzyme in the absence of oxygen.
[0285] A number of exemplary methods have been developed for the mutagenesis and diversification of genes to target desired properties of specific enzymes. Such methods are well known to those skilled in the art. Any of these can be used to alter and/or optimize the activity of a 4-HB, 4-HBal, 4-HBCoA, BDO or putrescine pathway enzyme or protein. Such methods include, but are not limited to EpPCR, which introduces random point mutations by reducing the fidelity of DNA polymerase in PCR reactions (Pritchard et al., J Theor. Biol. 234:497-509 (2005)); Error-prone Rolling Circle Amplification (epRCA), which is similar to epPCR except a whole circular plasmid is used as the template and random 6-mers with exonuclease resistant thiophosphate linkages on the last 2 nucleotides are used to amplify the plasmid followed by transformation into cells in which the plasmid is re-circularized at tandem repeats (Fujii et al., Nucleic Acids Res. 32:e145 (2004); and Fujii et al., Nat. Protoc. 1:2493-2497 (2006)); DNA or Family Shuffling, which typically involves digestion of two or more variant genes with nucleases such as Dnase I or EndoV to generate a pool of random fragments that are reassembled by cycles of annealing and extension in the presence of DNA polymerase to create a library of chimeric genes (Stemmer, Proc Natl Acad Sci USA 91:10747-10751 (1994); and Stemmer, Nature 370:389-391 (1994)); Staggered Extension (StEP), which entails template priming followed by repeated cycles of 2 step PCR with denaturation and very short duration of annealing/extension (as short as 5 sec) (Zhao et al., Nat. Biotechnol. 16:258-261 (1998)); Random Priming Recombination (RPR), in which random sequence primers are used to generate many short DNA fragments complementary to different segments of the template (Shao et al., Nucleic Acids Res 26:681-683 (1998)).
[0286] Additional methods include Heteroduplex Recombination, in which linearized plasmid DNA is used to form heteroduplexes that are repaired by mismatch repair (Volkov et al, Nucleic Acids Res. 27:e18 (1999); and Volkov et al., Methods Enzymol. 328:456-463 (2000)); Random Chimeragenesis on Transient Templates (RACHITT), which employs Dnase I fragmentation and size fractionation of single stranded DNA (ssDNA) (Coco et al., Nat. Biotechnol. 19:354-359 (2001)); Recombined Extension on Truncated templates (RETT), which entails template switching of unidirectionally growing strands from primers in the presence of unidirectional ssDNA fragments used as a pool of templates (Lee et al., J. Molec. Catalysis 26:119-129 (2003)); Degenerate Oligonucleotide Gene Shuffling (DOGS), in which degenerate primers are used to control recombination between molecules; (Bergquist and Gibbs, Methods Mol. Biol 352:191-204 (2007); Bergquist et al., Biomol. Eng 22:63-72 (2005); Gibbs et al., Gene 271:13-20 (2001)); Incremental Truncation for the Creation of Hybrid Enzymes (ITCHY), which creates a combinatorial library with 1 base pair deletions of a gene or gene fragment of interest (Ostermeier et al., Proc. Natl. Acad. Sci. USA 96:3562-3567 (1999); and Ostermeier et al., Nat. Biotechnol. 17:1205-1209 (1999)); Thio-Incremental Truncation for the Creation of Hybrid Enzymes (THIO-ITCHY), which is similar to ITCHY except that phosphothioate dNTPs are used to generate truncations (Lutz et al., Nucleic Acids Res 29:E16 (2001)); SCRATCHY, which combines two methods for recombining genes, ITCHY and DNA shuffling (Lutz et al., Proc. Natl. Acad. Sci. USA 98:11248-11253 (2001)); Random Drift Mutagenesis (RNDM), in which mutations made via epPCR are followed by screening/selection for those retaining usable activity (Bergquist et al., Biomol. Eng. 22:63-72 (2005)); Sequence Saturation Mutagenesis (SeSaM), a random mutagenesis method that generates a pool of random length fragments using random incorporation of a phosphothioate nucleotide and cleavage, which is used as a template to extend in the presence of "universal" bases such as inosine, and replication of an inosine-containing complement gives random base incorporation and, consequently, mutagenesis (Wong et al., Biotechnol. J. 3:74-82 (2008); Wong et al., Nucleic Acids Res. 32:e26 (2004); and Wong et al., Anal. Biochem. 341:187-189 (2005)); Synthetic Shuffling, which uses overlapping oligonucleotides designed to encode "all genetic diversity in targets" and allows a very high diversity for the shuffled progeny (Ness et al., Nat. Biotechnol. 20:1251-1255 (2002)); Nucleotide Exchange and Excision Technology NexT, which exploits a combination of dUTP incorporation followed by treatment with uracil DNA glycosylase and then piperidine to perform endpoint DNA fragmentation (Muller et al., Nucleic Acids Res. 33:e117 (2005)).
[0287] Further methods include Sequence Homology-Independent Protein Recombination (SHIPREC), in which a linker is used to facilitate fusion between two distantly related or unrelated genes, and a range of chimeras is generated between the two genes, resulting in libraries of single-crossover hybrids (Sieber et al., Nat. Biotechnol. 19:456-460 (2001)); Gene Site Saturation Mutagenesis.TM. (GSSM.TM.), in which the starting materials include a supercoiled double stranded DNA (dsDNA) plasmid containing an insert and two primers which are degenerate at the desired site of mutations (Kretz et al., Methods Enzymol. 388:3-11 (2004)); Combinatorial Cassette Mutagenesis (CCM), which involves the use of short oligonucleotide cassettes to replace limited regions with a large number of possible amino acid sequence alterations (Reidhaar-Olson et al. Methods Enzymol. 208:564-586 (1991); and Reidhaar-Olson et al. Science 241:53-57 (1988)); Combinatorial Multiple Cassette Mutagenesis (CMCM), which is essentially similar to CCM and uses epPCR at high mutation rate to identify hot spots and hot regions and then extension by CMCM to cover a defined region of protein sequence space (Reetz et al., Angew. Chem. Int. Ed Engl. 40:3589-3591 (2001)); the Mutator Strains technique, in which conditional ts mutator plasmids, utilizing the mutD5 gene, which encodes a mutant subunit of DNA polymerase III, to allow increases of 20 to 4000-X in random and natural mutation frequency during selection and block accumulation of deleterious mutations when selection is not required (Selifonova et al., Appl. Environ. Microbiol. 67:3645-3649 (2001)); Low et al., J. Mol. Biol. 260:359-3680 (1996)).
[0288] Additional exemplary methods include Look-Through Mutagenesis (LTM), which is a multidimensional mutagenesis method that assesses and optimizes combinatorial mutations of selected amino acids (Rajpal et al., Proc. Natl. Acad. Sci. USA 102:8466-8471 (2005)); Gene Reassembly, which is a DNA shuffling method that can be applied to multiple genes at one time or to create a large library of chimeras (multiple mutations) of a single gene (Tunable GeneReassembly.TM. (TGR.TM.) Technology supplied by Verenium Corporation), in Silico Protein Design Automation (PDA), which is an optimization algorithm that anchors the structurally defined protein backbone possessing a particular fold, and searches sequence space for amino acid substitutions that can stabilize the fold and overall protein energetics, and generally works most effectively on proteins with known three-dimensional structures (Hayes et al., Proc. Natl. Acad. Sci. USA 99:15926-15931 (2002)); and Iterative Saturation Mutagenesis (ISM), which involves using knowledge of structure/function to choose a likely site for enzyme improvement, performing saturation mutagenesis at chosen site using a mutagenesis method such as Stratagene QuikChange (Stratagene; San Diego Calif.), screening/selecting for desired properties, and, using improved clone(s), starting over at another site and continue repeating until a desired activity is achieved (Reetz et al., Nat. Protoc. 2:891-903 (2007); and Reetz et al., Angew. Chem. Int. Ed Engl. 45:7745-7751 (2006)).
[0289] Any of the aforementioned methods for mutagenesis can be used alone or in any combination. Additionally, any one or combination of the directed evolution methods can be used in conjunction with adaptive evolution techniques, as described herein.
[0290] It is understood that modifications which do not substantially affect the activity of the various embodiments of this invention are also provided within the definition of the invention provided herein. Accordingly, the following examples are intended to illustrate but not limit the present invention.
Example I
Biosynthesis of 4-Hydroxybutanoic Acid
[0291] This example describes exemplary biochemical pathways for 4-HB production.
[0292] Previous reports of 4-HB synthesis in microbes have focused on this compound as an intermediate in production of the biodegradable plastic poly-hydroxyalkanoate (PHA) (U.S. Pat. No. 6,117,658). The use of 4-HB/3-HB copolymers over poly-3-hydroxybutyrate polymer (PHB) can result in plastic that is less brittle (Saito and Doi, Intl. J. Biol. Macromol. 16:99-104 (1994)). The production of monomeric 4-HB described herein is a fundamentally distinct process for several reasons: (1) the product is secreted, as opposed to PHA which is produced intracellularly and remains in the cell; (2) for organisms that produce hydroxybutanoate polymers, free 4-HB is not produced, but rather the Coenzyme A derivative is used by the polyhydroxyalkanoate synthase; (3) in the case of the polymer, formation of the granular product changes thermodynamics; and (4) extracellular pH is not an issue for production of the polymer, whereas it will affect whether 4-HB is present in the free acid or conjugate base state, and also the equilibrium between 4-HB and GBL.
[0293] 4-HB can be produced in two enzymatic reduction steps from succinate, a central metabolite of the TCA cycle, with succinic semialdehyde as the intermediate (FIG. 1). The first of these enzymes, succinic semialdehyde dehydrogenase, is native to many organisms including E. coli, in which both NADH- and NADPH-dependent enzymes have been found (Donnelly and Cooper, Eur. J. Biochem. 113:555-561 (1981); Donnelly and Cooper, J. Bacteriol. 145:1425-1427 (1981); Marek and Henson, J. Bacteriol. 170:991-994 (1988)). There is also evidence supporting succinic semialdehyde dehydrogenase activity in S. cerevisiae (Ramos et al., Eur. J. Biochem. 149:401-404 (1985)), and a putative gene has been identified by sequence homology. However, most reports indicate that this enzyme proceeds in the direction of succinate synthesis, as shown in FIG. 1 (Donnelly and Cooper, supra; Lutke-Eversloh and Steinbuchel, FEMS Microbiol. Lett. 181:63-71 (1999)), participating in the degradation pathway of 4-HB and gamma-aminobutyrate. Succinic semialdehyde also is natively produced by certain microbial organisms such as E. coli through the TCA cycle intermediate .alpha.-ketogluterate via the action of two enzymes: glutamate:succinic semialdehyde transaminase and glutamate decarboxylase. An alternative pathway, used by the obligate anaerobe Clostridium kluyveri to degrade succinate, activates succinate to succinyl-CoA, then converts succinyl-CoA to succinic semialdehyde using an alternative succinic semialdehyde dehydrogenase which is known to function in this direction (Sohling and Gottschalk, Eur. J. Biochem. 212:121-127 (1993)). However, this route has the energetic cost of ATP required to convert succinate to succinyl-CoA.
[0294] The second enzyme of the pathway, 4-hydroxybutanoate dehydrogenase, is not native to E. coli or yeast but is found in various bacteria such as C. kluyveri and Ralstonia eutropha (Lutke-Eversloh and Steinbuchel, supra; Sohling and Gottschalk, J. Bacteriol. 178:871-880 (1996); Valentin et al., Eur. J. Biochem. 227:43-60 (1995); Wolff and Kenealy, Protein Expr. Purif. 6:206-212 (1995)). These enzymes are known to be NADH-dependent, though NADPH-dependent forms also exist. An additional pathway to 4-HB from alpha-ketoglutarate was demonstrated in E. coli resulting in the accumulation of poly(4-hydroxybutyric acid) (Song et al., Wei Sheng Wu Xue. Bao. 45:382-386 (2005)). The recombinant strain required the overexpression of three heterologous genes, PHA synthase (R. eutropha), 4-hydroxybutyrate dehydrogenase (R. eutropha) and 4-hydroxybutyrate:CoA transferase (C. kluyveri), along with two native E. coli genes: glutamate:succinic semialdehyde transaminase and glutamate decarboxylase. Steps 4 and 5 in FIG. 1 can alternatively be carried out by an alpha-ketoglutarate decarboxylase such as the one identified in Euglena gracilis (Shigeoka et al., Biochem. J. 282(Pt2):319-323 (1992); Shigeoka and Nakano, Arch. Biochem. Biophys. 288:22-28 (1991); Shigeoka and Nakano, Biochem J. 292(Pt 2):463-467 (1993)). However, this enzyme has not previously been applied to impact the production of 4-HB or related polymers in any organism.
[0295] The microbial production capabilities of 4-hydroxybutyrate were explored in two microbes, Escherichia coli and Saccharomyces cerevisiae, using in silico metabolic models of each organism. Potential pathways to 4-HB proceed via a succinate, succinyl-CoA, or alpha-ketoglutarate intermediate as shown in FIG. 1.
[0296] A first step in the 4-HB production pathway from succinate involves the conversion of succinate to succinic semialdehyde via an NADH- or NADPH-dependant succinic semialdehyde dehydrogenase. In E. coli, gabD is an NADP-dependant succinic semialdehyde dehydrogenase and is part of a gene cluster involved in 4-aminobutyrate uptake and degradation (Niegemann et al., Arch. Microbiol. 160:454-460 (1993); Schneider et al., J. Bacteriol. 184:6976-6986 (2002)). sad is believed to encode the enzyme for NAD-dependant succinic semialdehyde dehydrogenase activity (Marek and Henson, supra). S. cerevisiae contains only the NADPH-dependant succinic semialdehyde dehydrogenase, putatively assigned to UGA2, which localizes to the cytosol (Huh et al., Nature 425:686-691 (2003)). The maximum yield calculations assuming the succinate pathway to 4-HB in both E. coli and S. cerevisiae require only the assumption that a non-native 4-HB dehydrogenase has been added to their metabolic networks.
[0297] The pathway from succinyl-CoA to 4-hydroxybutyrate was described in U.S. Pat. No. 6,117,658 as part of a process for making polyhydroxyalkanoates comprising 4-hydroxybutyrate monomer units. Clostridium kluyveri is one example organism known to possess CoA-dependant succinic semialdehyde dehydrogenase activity (Sohling and Gottschalk, supra; Sohling and Gottschalk, supra). In this study, it is assumed that this enzyme, from C. kluyveri or another organism, is expressed in E. coli or S. cerevisiae along with a non-native or heterologous 4-HB dehydrogenase to complete the pathway from succinyl-CoA to 4-HB. The pathway from alpha-ketoglutarate to 4-HB was demonstrated in E. coli resulting in the accumulation of poly(4-hydroxybutyric acid) to 30% of dry cell weight (Song et al., supra). As E. coli and S. cerevisiae natively or endogenously possess both glutamate:succinic semialdehyde transaminase and glutamate decarboxylase (Coleman et al., J. Biol. Chem. 276:244-250 (2001)), the pathway from AKG to 4-HB can be completed in both organisms by assuming only that a non-native 4-HB dehydrogenase is present.
Example II
Biosynthesis of 1,4-Butanediol from Succinate and Alpha-Ketoglutarate
[0298] This example illustrates the construction and biosynthetic production of 4-HB and BDO from microbial organisms. Pathways for 4-HB and BDO are disclosed herein.
[0299] There are several alternative enzymes that can be utilized in the pathway described above. The native or endogenous enzyme for conversion of succinate to succinyl-CoA (Step 1 in FIG. 1) can be replaced by a CoA transferase such as that encoded by the cat1 gene C. kluyveri (Sohling and Gottschalk, Eur. J Biochem. 212:121-127 (1993)), which functions in a similar manner to Step 9. However, the production of acetate by this enzyme may not be optimal, as it might be secreted rather than being converted back to acetyl-CoA. In this respect, it also can be beneficial to eliminate acetate formation in Step 9. As one alternative to this CoA transferase, a mechanism can be employed in which the 4-HB is first phosphorylated by ATP and then converted to the CoA derivative, similar to the acetate kinase/phosphotransacetylase pathway in E. coli for the conversion of acetate to acetyl-CoA. The net cost of this route is one ATP, which is the same as is required to regenerate acetyl-CoA from acetate. The enzymes phosphotransbutyrylase (ptb) and butyrate kinase (bk) are known to carry out these steps on the non-hydroxylated molecules for butyrate production in C. acetobutylicum (Cary et al., Appl Environ Microbiol 56:1576-1583 (1990); Valentine, R. C. and R. S. Wolfe, J Biol Chem. 235:1948-1952 (1960)). These enzymes are reversible, allowing synthesis to proceed in the direction of 4-HB.
[0300] BDO also can be produced via .alpha.-ketoglutarate in addition to or instead of through succinate. A described previously, and exemplified further below, one pathway to accomplish product biosynthesis is with the production of succinic semialdehyde via .alpha.-ketoglutarate using the endogenous enzymes (FIG. 1, Steps 4-5). An alternative is to use an .alpha.-ketoglutarate decarboxylase that can perform this conversion in one step (FIG. 1, Step 8; Tian et al., Proc Natl Acad Sci US.A 102:10670-10675 (2005)).
[0301] For the construction of different strains of BDO-producing microbial organisms, a list of applicable genes was assembled for corroboration. Briefly, one or more genes within the 4-HB and/or BDO biosynthetic pathways were identified for each step of the complete BDO-producing pathway shown in FIG. 1, using available literature resources, the NCBI genetic database, and homology searches. The genes cloned and assessed in this study are presented below in Table 6, along with the appropriate references and URL citations to the polypeptide sequence. As discussed further below, some genes were synthesized for codon optimization while others were cloned via PCR from the genomic DNA of the native or wild-type organism. For some genes both approaches were used, and in this case the native genes are indicated by an "n" suffix to the gene identification number when used in an experiment. Note that only the DNA sequences differ; the proteins are identical.
TABLE-US-00006 TABLE 6 Genes expressed in host BDO-producting microbial organisms. Reaction Gene ID number Source number (FIG. 1) Gene name organism Enzyme name Link to protein sequence Reference 0001 9 Cat2 Clostridium 4-hydroxybutyrate ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=nuccore&id=1228100 1 kluyveri coenzyme A DSM 555 transferase 0002 12/13 adhE Clostridium Aldehyde/alcohol ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&val=15004739 2 acetobutylicum dehydrogenase ATCC 824 0003 12/13 adhE2 Clostridium Aldehyde/alcohol ncbi.nlm.nih.gov/entrez/viewer.fcgi?val=NP_149325.1 2 acetobutylicum dehydrogenase ATCC 824 0004 1 Cat1 Clostridium Succinate ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=nuccore&id=1228100 1 kluyveri coenzyme A DSM 555 transferase 0008 6 sucD Clostridium Succinic ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=nuccore&id=1228100 1 kluyveri semialdehyde DSM 555 dehydrogenase (CoA-dependent) 0009 7 4-HBd Ralstonia 4-hydroxybutyrate ncbi.nlm.nih.gov/entrez/viewer.fcgi?val=YP_726053.1 2 eutropha H16 dehydrogenase (NAD-dependent) 0010 7 4-HBd Clostridium 4-hydroxybutyrate ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=nuccore&id=1228100 1 kluyveri dehydrogenase DSM 555 (NAD-dependent) 0011 12/13 adhE E. coli Aldehyde/alcohol shigen.nig.ac.jp/ecoli/pec/genes.List.DetailAction.do?fromListFlag=true&f- eatureType=1&orfId=1219 dehydrogenase 0012 12/13 yqhD E. coli Aldehyde/alcohol shigen.nig.ac.jp/ecoli/pec/genes.List.DetailAction.do dehydrogenase 0013 13 bdhB Clostridium Butanol ncbi.nlm.nih.gov/entrez/viewer.fcgi?val=NP_349891.1 2 acetobutylicum dehydrogenase II ATCC 824 0020 11 ptb Clostridium Phospho- ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&id=15896327 2 acetobutylicum transbutyrylase ATCC 824 0021 10 buk1 Clostridium Butyrate kinase I ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&id=20137334 2 acetobutylicum ATCC 824 0022 10 buk2 Clostridium Butyrate kinase II ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&id=20137415 2 acetobutylicum ATCC 824 0023 13 adhEm isolated from Alcohol (37)d} metalibrary dehydrogenase of anaerobic sewage digester microbial consortia 0024 13 adhE Clostridium Alcohol genome.jp/dbget-bin/www_bget?cth:Cthe_0423 thermocellum dehydrogenase 0025 13 ald Clostridium Coenzyme A- ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&id=49036681 (31)d} beijerinckii acylating aldehyde dehydrogenase 0026 13 bdhA Clostridium Butanol ncbi.nlm.nih.gov/entrez/viewer.fcgi?val=NP_349892.1 2 acetobutylicum dehydrogenase ATCC 824 0027 12 bld Clostridium Butyraldehyde ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&id=31075383 4 saccharoperbutylacetonicum dehydrogenase 0028 13 bdh Clostridium Butanol ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&id=124221917 4 saccharoperbutylacetonicum dehydrogenase 0029 12/13 adhE Clostridium Aldehyde/alcohol genome.jp/dbget-bin/www_bget?ctc:CTC01366 tetani dehydrogenase 0030 12/13 adhE Clostridium Aldehyde/alcohol genome.jp/dbget-bin/www_bget?cpe:CPE2531 perfringens dehydrogenase 0031 12/13 adhE Clostridium Aldehyde/alcohol genome.jp/dbget-bin/www_bget?cdf:CD2966 difficile dehydrogenase 0032 8 sucA Mycobacterium .alpha.-ketoglutarate ncbi.nlm.nih.gov/entrez/viewer.fcgi?val=YP_977400.1 5 bovis decarboxylase BCG, Pasteur 0033 9 cat2 Clostridium 4-hydroxybutyrate ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&val=6249316 aminobutyricum coenzyme A transferase 0034 9 cat2 Porphyromonas 4-hydroxybutyrate ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&val=34541558 gingivalis coenzyme A W83 transferase 0035 6 sucD Porphyromonas Succinic ncbi.nlm.nih.gov/entrez/viewer.fcgi?val=NP_904963.1 gingivalis semialdehyde W83 dehydrogenase (CoA-dependent) 0036 7 4-HBd Porphyromonas NAD-dependent ncbi.nlm.nih.gov/entrez/viewer.fcgi?val=NP_904964.1 gingivalis 4-hydroxybutyrate W83 dehydrogenase 0037 7 gbd Uncultured 4-hydroxybutyrate ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=nuccore&id=5916168 6 bacterium dehydrogenase 0038 1 sucCD E. coli Succinyl-CoA shigen.nig.ac.jp/ecoli/pec/genes.List.DetailAction.do synthetase .sup.1Sohling and Gottschalk, Eur. J. Biochem. 212: 121-127 (1993); Sohling and Gottschalk, J. Bacteriol. 178: 871-880 (1996) .sup.2Nolling et al., J., J. Bacteriol. 183: 4823-4838 (2001) .sup.3Pohlmann et al., Nat. Biotechnol. 24: 1257-1262 (2006) .sup.4Kosaka et al., Biosci. Biotechnol. Biochem. 71: 58-68 (2007) .sup.5Brosch et al., Proc. Natl. Acad. Sci. U.S.A. 104: 5596-5601 (2007) .sup.6Henne et al., Appl. Environ. Microbiol. 65: 3901-3907 (1999)
[0302] Expression Vector Construction for BDO Pathway.
[0303] Vector backbones and some strains were obtained from Dr. Rolf Lutz of Expressys (expressys.de/). The vectors and strains are based on the pZ Expression System developed by Dr. Rolf Lutz and Prof. Hermann Bujard (Lutz, R. and H. Bujard, Nucleic Acids Res 25:1203-1210 (1997)). Vectors obtained were pZE13luc, pZA33luc, pZS*13luc and pZE22luc and contained the luciferase gene as a stuffer fragment. To replace the luciferase stuffer fragment with a lacZ-alpha fragment flanked by appropriate restriction enzyme sites, the luciferase stuffer fragment was first removed from each vector by digestion with EcoRI and XbaI. The lacZ-alpha fragment was PCR amplified from pUC19 with the following primers:
TABLE-US-00007 lacZalpha-RI (SEQ ID NO: 1) 5'GACGAATTCGCTAGCAAGAGGAGAAGTCGACATGTCCAATTCACTGGC CGTCGTTTTAC3' lacZalpha 3'BB (SEQ ID NO: 2) 5'-GACCCTAGGAAGCTTTCTAGAGTCGACCTATGCGGCATCAGAGCAG A-3'.
[0304] This generated a fragment with a 5' end of EcoRI site, NheI site, a Ribosomal Binding Site, a SalI site and the start codon. On the 3' end of the fragment contained the stop codon, XbaI, HindIII, and AvrII sites. The PCR product was digested with EcoRI and AvrII and ligated into the base vectors digested with EcoRI and XbaI (XbaI and AvrII have compatible ends and generate a non-site). Because NheI and XbaI restriction enzyme sites generate compatible ends that can be ligated together (but generate a NheI/XbaI non-site that is not digested by either enzyme), the genes cloned into the vectors could be "Biobricked" together (openwetware.org/wiki/Synthetic_Biology:BioBricks). Briefly, this method allows joining an unlimited number of genes into the vector using the same 2 restriction sites (as long as the sites do not appear internal to the genes), because the sites between the genes are destroyed after each addition.
[0305] All vectors have the pZ designation followed by letters and numbers indication the origin of replication, antibiotic resistance marker and promoter/regulatory unit. The origin of replication is the second letter and is denoted by E for ColE1, A for p15A and S for pSC101-based origins. The first number represents the antibiotic resistance marker (1 for Ampicillin, 2 for Kanamycin, 3 for Chloramphenicol, 4 for Spectinomycin and 5 for Tetracycline). The final number defines the promoter that regulated the gene of interest (1 for P.sub.LtetO-1, 2 for P.sub.LlacO-1, 3 for P.sub.A1lacO-1, and 4 for P.sub.lac/ara-1). The MCS and the gene of interest follows immediately after. For the work discussed here we employed two base vectors, pZA33 and pZE13, modified for the biobricks insertions as discussed above. Once the gene(s) of interest have been cloned into them, resulting plasmids are indicated using the four digit gene codes given in Table 6; e.g., pZA33-XXXX-YYYY- . . . .
[0306] Host Strain Construction.
[0307] The parent strain in all studies described here is E. coli K-12 strain MG1655. Markerless deletion strains in adhE, gabD, and aldA were constructed under service contract by a third party using the redET method (Datsenko, K. A. and B. L. Wanner, Proc Natl Acad Sci US.A 97:6640-6645 (2000)). Subsequent strains were constructed via bacteriophage P1 mediated transduction (Miller, J. Experiments in Molecular Genetics, Cold Spring Harbor Laboratories, New York (1973)). Strain C600Z1 (laci.sup.q, PN25-tetR, Sp.sup.R,lacY1, leuB6,mcrB+, supE44, thi-1, thr-1, tonA21) was obtained from Expressys and was used as a source of a lacI.sup.q allele for P1 transduction. Bacteriophage P1vir was grown on the C600Z1 E. coli strain, which has the spectinomycin resistance gene linked to the lacI.sup.q. The P1 lysate grown on C600Z1 was used to infect MG1655 with selection for spectinomycin resistance. The spectinomycin resistant colonies were then screened for the linked lacI.sup.q by determining the ability of the transductants to repress expression of a gene linked to a P.sub.A1lacO-1 promoter. The resulting strain was designated MG1655 lacI.sup.q. A similar procedure was used to introduce lacI.sup.Q into the deletion strains.
[0308] Production of 4-HB from Succinate.
[0309] For construction of a 4-HB producer from succinate, genes encoding steps from succinate to 4-HB and 4-HB-CoA (1, 6, 7, and 9 in FIG. 1) were assembled onto the pZA33 and pZE13 vectors as described below. Various combinations of genes were assessed, as well as constructs bearing incomplete pathways as controls (Tables 7 and 8). The plasmids were then transformed into host strains containing lacI.sup.Q, which allow inducible expression by addition of isopropyl .beta.-D-1-thiogalactopyranoside (IPTG). Both wild-type and hosts with deletions in genes encoding the native succinic semialdehyde dehydrogenase (step 2 in FIG. 1) were tested.
[0310] Activity of the heterologous enzymes were first tested in in vitro assays, using strain MG1655 lacI.sup.Q as the host for the plasmid constructs containing the pathway genes. Cells were grown aerobically in LB media (Difco) containing the appropriate antibiotics for each construct, and induced by addition of IPTG at 1 mM when the optical density (OD600) reached approximately 0.5. Cells were harvested after 6 hours, and enzyme assays conducted as discussed below.
[0311] In Vitro Enzyme Assays.
[0312] To obtain crude extracts for activity assays, cells were harvested by centrifugation at 4,500 rpm (Beckman-Coulter, Allegera X-15R) for 10 min. The pellets were resuspended in 0.3 mL BugBuster (Novagen) reagent with benzonase and lysozyme, and lysis proceeded for 15 minutes at room temperature with gentle shaking. Cell-free lysate was obtained by centrifugation at 14,000 rpm (Eppendorf centrifuge 5402) for 30 min at 4.degree. C. Cell protein in the sample was determined using the method of Bradford et al., Anal. Biochem. 72:248-254 (1976), and specific enzyme assays conducted as described below. Activities are reported in Units/mg protein, where a unit of activity is defined as the amount of enzyme required to convert 1 .mu.mol of substrate in 1 min. at room temperature. In general, reported values are averages of at least 3 replicate assays.
[0313] Succinyl-CoA transferase (Cat1) activity was determined by monitoring the formation of acetyl-CoA from succinyl-CoA and acetate, following a previously described procedure Sohling and Gottschalk, J. Bacteriol. 178:871-880 (1996). Succinyl-CoA synthetase (SucCD) activity was determined by following the formation of succinyl-CoA from succinate and CoA in the presence of ATP. The experiment followed a procedure described by Cha and Parks, J. Biol. Chem. 239:1961-1967 (1964). CoA-dependent succinate semialdehyde dehydrogenase (SucD) activity was determined by following the conversion of NAD to NADH at 340 nm in the presence of succinate semialdehyde and CoA (Sohling and Gottschalk, Eur. J. Biochem. 212:121-127 (1993)). 4-HB dehydrogenase (4-HBd) enzyme activity was determined by monitoring the oxidation of NADH to NAD at 340 nm in the presence of succinate semialdehyde. The experiment followed a published procedure Gerhardt et al. Arch. Microbiol. 174:189-199 (2000). 4-HB CoA transferase (Cat2) activity was determined using a modified procedure from Scherf and Buckel, Appl. Environ. Microbiol. 57:2699-2702 (1991). The formation of 4-HB-CoA or butyryl-CoA formation from acetyl-CoA and 4-HB or butyrate was determined using HPLC.
[0314] Alcohol (ADH) and aldehyde (ALD) dehydrogenase was assayed in the reductive direction using a procedure adapted from several literature sources (Durre et al., FEMS Microbiol. Rev. 17:251-262 (1995); Palosaari and Rogers, J. Bacteriol. 170:2971-2976 (1988) and Welch et al., Arch. Biochem. Biophys. 273:309-318 (1989). The oxidation of NADH is followed by reading absorbance at 340 nM every four seconds for a total of 240 seconds at room temperature. The reductive assays were performed in 100 mM MOPS (adjusted to pH 7.5 with KOH), 0.4 mM NADH, and from 1 to 50 .mu.l of cell extract. The reaction is started by adding the following reagents: 100 .mu.l of 100 mM acetaldehyde or butyraldehyde for ADH, or 100 .mu.l of 1 mM acetyl-CoA or butyryl-CoA for ALD. The Spectrophotometer is quickly blanked and then the kinetic read is started. The resulting slope of the reduction in absorbance at 340 nM per minute, along with the molar extinction coefficient of NAD(P)H at 340 nM (6000) and the protein concentration of the extract, can be used to determine the specific activity.
[0315] The enzyme activity of PTB is measured in the direction of butyryl-CoA to butyryl-phosphate as described in Cary et al. J. Bacteriol. 170:4613-4618 (1988). It provides inorganic phosphate for the conversion, and follows the increase in free CoA with the reagent 5,5'-dithiobis-(2-nitrobenzoic acid), or DTNB. DTNB rapidly reacts with thiol groups such as free CoA to release the yellow-colored 2-nitro-5-mercaptobenzoic acid (TNB), which absorbs at 412 nm with a molar extinction coefficient of 14,140 M cm.sup.-1. The assay buffer contained 150 mM potassium phosphate at pH 7.4, 0.1 mM DTNB, and 0.2 mM butyryl-CoA, and the reaction was started by addition of 2 to 50 .mu.L cell extract. The enzyme activity of BK is measured in the direction of butyrate to butyryl-phosphate formation at the expense of ATP. The procedure is similar to the assay for acetate kinase previously described Rose et al., J. Biol. Chem. 211:737-756 (1954). However we have found another acetate kinase enzyme assay protocol provided by Sigma to be more useful and sensitive. This assay links conversion of ATP to ADP by acetate kinase to the linked conversion of ADP and phosphoenol pyruvate (PEP) to ATP and pyruvate by pyruvate kinase, followed by the conversion of pyruvate and NADH to lactate and NAD+ by lactate dehydrogenase. Substituting butyrate for acetate is the only major modification to allow the assay to follow BK enzyme activity. The assay mixture contained 80 mM triethanolamine buffer at pH 7.6, 200 mM sodium butyrate, 10 mM MgCl2, 0.1 mM NADH, 6.6 mM ATP, 1.8 mM phosphoenolpyruvate. Pyruvate kinase, lactate dehydrogenase, and myokinase were added according to the manufacturer's instructions. The reaction was started by adding 2 to 50 .mu.L cell extract, and the reaction was monitored based on the decrease in absorbance at 340 nm indicating NADH oxidation.
[0316] Analysis of CoA Derivatives by HPLC.
[0317] An HPLC based assay was developed to monitor enzymatic reactions involving coenzyme A (CoA) transfer. The developed method allowed enzyme activity characterization by quantitative determination of CoA, acetyl CoA (AcCoA), butyryl CoA (BuCoA) and 4-hydroxybutyrate CoA (4-HBCoA) present in in-vitro reaction mixtures. Sensitivity down to low .mu.M was achieved, as well as excellent resolution of all the CoA derivatives of interest.
[0318] Chemical and sample preparation was performed as follows. Briefly, CoA, AcCoA, BuCoA and all other chemicals, were obtained from Sigma-Aldrich. The solvents, methanol and acetonitrile, were of HPLC grade. Standard calibration curves exhibited excellent linearity in the 0.01-1 mg/mL concentration range. Enzymatic reaction mixtures contained 100 mM Tris HCl buffer (pH 7), aliquots were taken at different time points, quenched with formic acid (0.04% final concentration) and directly analyzed by HPLC.
[0319] HPLC analysis was performed using an Agilent 1100 HPLC system equipped with a binary pump, degasser, thermostated autosampler and column compartment, and diode array detector (DAD), was used for the analysis. A reversed phase column, Kromasil 100 Sum C18, 4.6.times.150 mm (Peeke Scientific), was employed. 25 mM potassium phosphate (pH 7) and methanol or acetonitrile, were used as aqueous and organic solvents at 1 mL/min flow rate. Two methods were developed: a short one with a faster gradient for the analysis of well-resolved CoA, AcCoA and BuCoA, and a longer method for distinguishing between closely eluting AcCoA and 4-HBCoA. Short method employed acetonitrile gradient (0 min-5%, 6 min-30%, 6.5 min-5%, 10 min-5%) and resulted in the retention times 2.7, 4.1 and 5.5 min for CoA, AcCoA and BuCoA, respectively. In the long method methanol was used with the following linear gradient: 0 min-5%, 20 min-35%, 20.5 min-5%, 25 min-5%. The retention times for CoA, AcCoA, 4-HBCoA and BuCoA were 5.8, 8.4, 9.2 and 16.0 min, respectively. The injection volume was 5 .mu.L, column temperature 30.degree. C., and UV absorbance was monitored at 260 nm.
[0320] The results demonstrated activity of each of the four pathway steps (Table 7), though activity is clearly dependent on the gene source, position of the gene in the vector, and the context of other genes with which it is expressed. For example, gene 0035 encodes a succinic semialdehyde dehydrogenase that is more active than that encoded by 0008, and 0036 and 0010n are more active 4-HB dehydrogenase genes than 0009. There also seems to be better 4-HB dehydrogenase activity when there is another gene preceding it on the same operon.
TABLE-US-00008 TABLE 7 In vitro enzyme activities in cell extracts from MG1655 lacI.sup.Q containing the plasmids expressing genes in the 4-HB-CoA pathway. Activities are reported in Units/mg protein, where a unit of activity is defined as the amount of enzyme required to convert 1 .mu.mol of substrate in 1 min. at room temperature. Sample # pZE13 (a) pZA33 (b) OD600 Cell Prot (c) Cat1 SucD 4HBd Cat2 1 cat1 (0004) 2.71 6.43 1.232 0.00 2 cat1 (0004)-sucD (0035) 2.03 5.00 0.761 2.57 3 cat1 (0004)-sucD (0008) 1.04 3.01 0.783 0.01 4 sucD (0035) 2.31 6.94 2.32 5 sucD (0008) 1.10 4.16 0.05 6 4hbd (0009) 2.81 7.94 0.003 0.25 7 4hbd (0036) 2.63 7.84 3.31 8 4hbd (0010n) 2.00 5.08 2.57 9 cat1 (0004)-sucD (0035) 4hbd (0009) 2.07 5.04 0.600 1.85 0.01 10 cat1 (0004)-sucD (0035) 4hbd (0036) 2.08 5.40 0.694 1.73 0.41 11 cat1 (0004)-sucD (0035) 4hbd (0010n) 2.44 4.73 0.679 2.28 0.37 12 cat1 (0004)-sucD (0008) 4hbd (0009) 1.08 3.99 0.572 -0.01 0.02 13 cat1 (0004)-sucD (0008) 4hbd (0036) 0.77 2.60 0.898 -0.01 0.04 14 cat1 (0004)-sucD (0008) 4hbd (0010n) 0.63 2.47 0.776 0.00 0.00 15 cat2 (0034) 2.56 7.86 1.283 16 cat2(0034)-4hbd(0036) 3.13 8.04 24.86 0.993 17 cat2(0034)-4hbd(0010n) 2.38 7.03 7.45 0.675 18 4hbd(0036)-cat2(0034) 2.69 8.26 2.15 7.490 19 4hbd(0010n)-cat2(0034) 2.44 6.59 0.59 4.101 (a) Genes expressed from Plac on pZE13, a high-copy plasmid with colE1 origin and ampicillin resistance. Gene identification numbers are as given in Table 6 (b) Genes expressed from Plac on pZA33, a medium-copy plasmid with pACYC origin and chloramphenicol resistance. (c) Cell protein given as mg protein per mL extract.
[0321] Recombinant strains containing genes in the 4-HB pathway were then evaluated for the ability to produce 4-HB in vivo from central metabolic intermediates. Cells were grown anaerobically in LB medium to OD600 of approximately 0.4, then induced with 1 mM IPTG. One hour later, sodium succinate was added to 10 mM, and samples taken for analysis following an additional 24 and 48 hours. 4-HB in the culture broth was analyzed by GC-MS as described below. The results indicate that the recombinant strain can produce over 2 mM 4-HB after 24 hours, compared to essentially zero in the control strain (Table 8).
TABLE-US-00009 TABLE 8 Production of 4-HB from succinate in E. coli strains harboring plasmids expressing various combinations of 4-HB pathway genes. 24 Hours 48 Hours 4HB, 4HB 4HB, 4HB Sample # Host Strain pZE13 pZA33 OD600 .mu.M norm. (a) OD600 .mu.M norm. (a) 1 MG1655 laclq cat1 (0004)-sucD (0035) 4hbd (0009) 0.47 487 1036 1.04 1780 1711 2 MG1655 laclq cat1 (0004)-sucD (0035) 4hbd (0027) 0.41 111 270 0.99 214 217 3 MG1655 laclq cat1 (0004)-sucD (0035) 4hbd (0036) 0.47 863 1835 0.48 2152 4484 4 MG1655 laclq cat1 (0004)-sucD (0035) 4hbd (0010n) 0.46 956 2078 0.49 2221 4533 5 MG1655 laclq cat1 (0004)-sucD (0008) 4hbd (0009) 0.38 493 1296 0.37 1338 3616 6 MG1655 laclq cat1 (0004)-sucD (0008) 4hbd (0027) 0.32 26 81 0.27 87 323 7 MG1655 laclq cat1 (0004)-sucD (0008) 4hbd (0036) 0.24 506 2108 0.31 1448 4672 8 MG1655 laclq cat1 (0004)-sucD (0008) 4hbd (0010n) 0.24 78 324 0.56 233 416 9 MG1655 laclq gabD cat1 (0004)-sucD (0035) 4hbd (0009) 0.53 656 1237 1.03 1643 1595 10 MG1655 laclq gabD cat1 (0004)-sucD (0035) 4hbd (0027) 0.44 92 209 0.98 214 218 11 MG1655 laclq gabD cat1 (0004)-sucD (0035) 4hbd (0036) 0.51 1072 2102 0.97 2358 2431 12 MG1655 laclq gabD cat1 (0004)-sucD (0035) 4hbd (0010n) 0.51 981 1924 0.97 2121 2186 13 MG1655 laclq gabD cat1 (0004)-sucD (0008) 4hbd (0009) 0.35 407 1162 0.77 1178 1530 14 MG1655 laclq gabD cat1 (0004)-sucD (0008) 4hbd (0027) 0.51 19 36 1.07 50 47 15 MG1655 laclq gabD cat1 (0004)-sucD (0008) 4hbd (0036) 0.35 584 1669 0.78 1350 1731 16 MG1655 laclq gabD cat1 (0004)-sucD (0008) 4hbd (0010n) 0.32 74 232 0.82 232 283 17 MG1655 laclq vector only vector only 0.8 1 2 1.44 3 2 18 MG1655 laclq gabD vector only vector only 0.89 1 2 1.41 7 5 (a) Normalized 4-HB concentration, .mu.M/OD600 units
[0322] An alternate to using a CoA transferase (cat1) to produce succinyl-CoA from succinate is to use the native E. coli sucCD genes, encoding succinyl-CoA synthetase. This gene cluster was cloned onto pZE13 along with candidate genes for the remaining steps to 4-HB to create pZE13-0038-0035-0036.
[0323] Production of 4-HB from Glucose.
[0324] Although the above experiments demonstrate a functional pathway to 4-HB from a central metabolic intermediate (succinate), an industrial process would require the production of chemicals from low-cost carbohydrate feedstocks such as glucose or sucrose. Thus, the next set of experiments was aimed to determine whether endogenous succinate produced by the cells during growth on glucose could fuel the 4-HB pathway. Cells were grown anaerobically in M9 minimal medium (6.78 g/L Na.sub.2HPO.sub.4, 3.0 g/L KH.sub.2PO.sub.4, 0.5 g/L NaCl, 1.0 g/L NH.sub.4Cl, 1 mM MgSO.sub.4, 0.1 mM CaCl.sub.2) supplemented with 20 g/L glucose, 100 mM 3-(N-morpholino)propanesulfonic acid (MOPS) to improve the buffering capacity, 10 .mu.g/mL thiamine, and the appropriate antibiotics. 0.25 mM IPTG was added when OD600 reached approximately 0.2, and samples taken for 4-HB analysis every 24 hours following induction. In all cases 4-HB plateaued after 24 hours, with a maximum of about 1 mM in the best strains (FIG. 3a), while the succinate concentration continued to rise (FIG. 3b). This indicates that the supply of succinate to the pathway is likely not limiting, and that the bottleneck may be in the activity of the enzymes themselves or in NADH availability. 0035 and 0036 are clearly the best gene candidates for CoA-dependent succinic semialdehyde dehydrogenase and 4-HB dehydrogenase, respectively. The elimination of one or both of the genes encoding known (gabD) or putative (aldA) native succinic semialdehyde dehydrogenases had little effect on performance. Finally, it should be noted that the cells grew to a much lower OD in the 4-HB-producing strains than in the controls (FIG. 3c).
[0325] An alternate pathway for the production of 4-HB from glucose is via .alpha.-ketoglutarate. We explored the use of an .alpha.-ketoglutarate decarboxylase from Mycobacterium tuberculosis Tian et al., Proc. Natl. Acad. Sci. USA 102:10670-10675 (2005) to produce succinic semialdehyde directly from .alpha.-ketoglutarate (step 8 in FIG. 1). To demonstrate that this gene (0032) was functional in vivo, we expressed it on pZE13 in the same host as 4-HB dehydrogenase (gene 0036) on pZA33. This strain was capable of producing over 1.0 mM 4-HB within 24 hours following induction with 1 mM IPTG (FIG. 4). Since this strain does not express a CoA-dependent succinic semialdehyde dehydrogenase, the possibility of succinic semialdehyde production via succinyl-CoA is eliminated. It is also possible that the native genes responsible for producing succinic semialdehyde could function in this pathway (steps 4 and 5 in FIG. 1); however, the amount of 4-HB produced when the pZE13-0032 plasmid was left out of the host is the negligible.
[0326] Production of BDO from 4-HB.
[0327] The production of BDO from 4-HB required two reduction steps, catalyzed by dehydrogenases. Alcohol and aldehyde dehydrogenases (ADH and ALD, respectively) are NAD+/H and/or NADP+/H-dependent enzymes that together can reduce a carboxylic acid group on a molecule to an alcohol group, or in reverse, can perform the oxidation of an alcohol to a carboxylic acid. This biotransformation has been demonstrated in wild-type Clostridium acetobutylicum (Jewell et al., Current Microbiology, 13:215-19 (1986)), but neither the enzymes responsible nor the genes responsible were identified. In addition, it is not known whether activation to 4-HB-CoA is first required (step 9 in FIG. 1), or if the aldehyde dehydrogenase (step 12) can act directly on 4-HB. We developed a list of candidate enzymes from C. acetobutylicum and related organisms based on known activity with the non-hydroxylated analogues to 4-HB and pathway intermediates, or by similarity to these characterized genes (Table 6). Since some of the candidates are multifunctional dehydrogenases, they could potentially catalyze both the NAD(P)H-dependent reduction of the acid (or CoA-derivative) to the aldehyde, and of the aldehyde to the alcohol. Before beginning work with these genes in E. coli, we first validated the result referenced above using C. acetobutylicum ATCC 824. Cells were grown in Schaedler broth (Accumedia, Lansing, Mich.) supplemented with 10 mM 4-HB, in an anaerobic atmosphere of 10% CO.sub.2, 10% H.sub.2, and 80% N.sub.2 at 30.degree. C. Periodic culture samples were taken, centrifuged, and the broth analyzed for BDO by GC-MS as described below. BDO concentrations of 0.1 mM, 0.9 mM, and 1.5 mM were detected after 1 day, 2 days, and 7 days incubation, respectively. No BDO was detected in culture grown without 4-HB addition. To demonstrate that the BDO produced was derived from glucose, we grew the best BDO producing strain MG1655 lacI.sup.Q pZE13-0004-0035-0002 pZA33-0034-0036 in M9 minimal medium supplemented with 4 g/L uniformly labeled .sup.13C-glucose. Cells were induced at OD of 0.67 with 1 mM IPTG, and a sample taken after 24 hours. Analysis of the culture supernatant was performed by mass spectrometry.
[0328] Gene candidates for the 4-HB to BDO conversion pathway were next tested for activity when expressed in the E. coli host MG1655 lacI.sup.Q. Recombinant strains containing each gene candidate expressed on pZA33 were grown in the presence of 0.25 mM IPTG for four hours at 37.degree. C. to fully induce expression of the enzyme. Four hours after induction, cells were harvested and assayed for ADH and ALD activity as described above. Since 4-HB-CoA and 4-hydroxybutyraldehyde are not available commercially, assays were performed using the non-hydroxylated substrates (Table 9). The ratio in activity between 4-carbon and 2-carbon substrates for C. acetobutylicum adhE2 (0002) and E. coli adhE (0011) were similar to those previously reported in the literature a Atsumi et al., Biochim. Biophys. Acta. 1207:1-11 (1994).
TABLE-US-00010 TABLE 9 In vitro enzyme activities in cell extracts from MG1655 lacI.sup.Q containing pZA33 expressing gene candidates for aldehyde and alcohol dehydrogenases. Aldehyde dehydrogenase Alcohol dehydrogenase Substrate Gene Butyryl-CoA Acetyl-CoA Butyraldehyde Acetaldehyde 0002 0.0076 0.0046 0.0264 0.0247 0003n 0.0060 0.0072 0.0080 0.0075 0011 0.0069 0.0095 0.0265 0.0093 0013 N.D. N.D. 0.0130 0.0142 0023 0.0089 0.0137 0.0178 0.0235 0025 0 0.0001 N.D. N.D. 0026 0 0.0005 0.0024 0.0008 Activities are expressed in .mu.mol min.sup.-1 mg cell protein.sup.-1. N.D., not determined.
[0329] For the BDO production experiments, cat2 from Porphyromonas gingivalis W83 (gene 0034) was included on pZA33 for the conversion of 4-HB to 4-HB-CoA, while the candidate dehydrogenase genes were expressed on pZE13. The host strain was MG1655 lacI.sup.Q. Along with the alcohol and aldehyde dehydrogenase candidates, we also tested the ability of CoA-dependent succinic semialdehyde dehydrogenases (sucD) to function in this step, due to the similarity of the substrates. Cells were grown to an OD of about 0.5 in LB medium supplemented with 10 mM 4-HB, induced with 1 mM IPTG, and culture broth samples taken after 24 hours and analyzed for BDO as described below. The best BDO production occurred using adhE2 from C. acetobutylicum, sucD from C. kluyveri, or sucD from P. gingivalis (FIG. 5). Interestingly, the absolute amount of BDO produced was higher under aerobic conditions; however, this is primarily due to the lower cell density achieved in anaerobic cultures. When normalized to cell OD, the BDO production per unit biomass is higher in anaerobic conditions (Table 10).
TABLE-US-00011 TABLE 10 Absolute and normalized BDO concentrations from cultures of cells expressing adhE2 from C. acetobutylicum, sucD from C. kluyveri, or sucD from P. gingivalis (data from experiments 2, 9, and 10 in FIG. 3), as well as the negative control (experiment 1). Gene BDO OD expressed Conditions (.mu.M) (600 nm) BDO/OD none Aerobic 0 13.4 0 none Microaerobic 0.5 6.7 0.09 none Anaerobic 2.2 1.26 1.75 0002 Aerobic 138.3 9.12 15.2 0002 Microaerobic 48.2 5.52 8.73 0002 Anaerobic 54.7 1.35 40.5 0008n Aerobic 255.8 5.37 47.6 0008n Microaerobic 127.9 3.05 41.9 0008n Anaerobic 60.8 0.62 98.1 0035 Aerobic 21.3 14.0 1.52 0035 Microaerobic 13.1 4.14 3.16 0035 Anaerobic 21.3 1.06 20.1
[0330] As discussed above, it may be advantageous to use a route for converting 4-HB to 4-HB-CoA that does not generate acetate as a byproduct. To this aim, we tested the use of phosphotransbutyrylase (ptb) and butyrate kinase (bk) from C. acetobutylicum to carry out this conversion via steps 10 and 11 in FIG. 1. The native ptb/bk operon from C. acetobutylicum (genes 0020 and 0021) was cloned and expressed in pZA33. Extracts from cells containing the resulting construct were taken and assayed for the two enzyme activities as described herein. The specific activity of BK was approximately 65 U/mg, while the specific activity of PTB was approximately 5 U/mg. One unit (U) of activity is defined as conversion of 1 .mu.M substrate in 1 minute at room temperature. Finally, the construct was tested for participation in the conversion of 4-HB to BDO. Host strains were transformed with the pZA33-0020-0021 construct described and pZE13-0002, and compared to use of cat2 in BDO production using the aerobic procedure used above in FIG. 5. The BK/PTB strain produced 1 mM BDO, compared to 2 mM when using cat2 (Table 11). Interestingly, the results were dependent on whether the host strain contained a deletion in the native adhE gene.
TABLE-US-00012 TABLE 11 Absolute and normalized BDO concentrations from cultures of cells expressing adhE2 from C. acetobutylicum in pZE13 along with either cat2 from P. gingivalis (0034) or the PTB/BK genes from C. acetobutylicum on pZA33. Host strains were either MG1655 lacI.sup.Q or MG1655 .DELTA.adhE lacI.sup.Q. BDO OD Genes Host Strain (.mu.M) (600 nm) BDO/OD 0034 MG1655 lacI.sup.Q 0.827 19.9 0.042 0020 + 0021 MG1655 lacI.sup.Q 0.007 9.8 0.0007 0034 MG1655 .DELTA.adhE 2.084 12.5 0.166 lacI.sup.Q 0020 + 0021 MG1655 .DELTA.adhE 0.975 18.8 0.052 lacI.sup.Q
[0331] Production of BDO from Glucose.
[0332] The final step of pathway corroboration is to express both the 4-HB and BDO segments of the pathway in E. coli and demonstrate production of BDO in glucose minimal medium. New plasmids were constructed so that all the required genes fit on two plasmids. In general, cat1, adhE, and sucD genes were expressed from pZE13, and cat2 and 4-HBd were expressed from pZA33. Various combinations of gene source and gene order were tested in the MG1655 lacI.sup.Q background. Cells were grown anaerobically in M9 minimal medium (6.78 g/L Na.sub.2HPO.sub.4, 3.0 g/L KH.sub.2PO.sub.4, 0.5 g/L NaCl, 1.0 g/L NH.sub.4Cl, 1 mM MgSO.sub.4, 0.1 mM CaCl.sub.2) supplemented with 20 g/L glucose, 100 mM 3-(N-morpholino)propanesulfonic acid (MOPS) to improve the buffering capacity, 10 .mu.g/mL thiamine, and the appropriate antibiotics. 0.25 mM IPTG was added approximately 15 hours following inoculation, and culture supernatant samples taken for BDO, 4-HB, and succinate analysis 24 and 48 hours following induction. The production of BDO appeared to show a dependency on gene order (Table 12). The highest BDO production, over 0.5 mM, was obtained with cat2 expressed first, followed by 4-HBd on pZA33, and cat1 followed by P. gingivalis sucD on pZE13. The addition of C. acetobutylicum adhE2 in the last position on pZE13 resulted in slight improvement. 4-HB and succinate were also produced at higher concentrations.
TABLE-US-00013 TABLE 12 Production of BDO, 4-HB, and succinate in recombinant E. coli strains expressing combinations of BDO pathway genes, grown in minimal medium supplemented with 20 g/L glucose. Concentrations are given in mM. 24 Hours 48 Hours Sample pZE13 pZA33 Induction OD OD600nm Su 4HB BDO OD600nm Su 4HB BDO 1 cat1(0004)-sucD(0035) 4hbd (0036)-cat2(0034) 0.92 1.29 5.44 1.37 0.240 1.24 6.42 1.49 0.280 2 cat1(0004)-sucD(0008N) 4hbd (0036)-cat2(0034) 0.36 1.11 6.90 1.24 0.011 1.06 7.63 1.33 0.011 3 adhE(0002)-cat1(0004)- 4hbd (0036)-cat2(0034) 0.20 0.44 0.34 1.84 0.050 0.60 1.93 2.67 0.119 sucD(0035) 4 cat1(0004)-sucD(0035)- 4hbd (0036)-cat2(0034) 1.31 1.90 9.02 0.73 0.073 1.95 9.73 0.82 0.077 adhE(0002) 5 adhE(0002)-cat1(0004)- 4hbd (0036)-cat2(0034) 0.17 0.45 1.04 1.04 0.008 0.94 7.13 1.02 0.017 sucD(0008N) 6 cat1(0004)-sucD(0008N)- 4hbd (0036)-cat2(0034) 1.30 1.77 10.47 0.25 0.004 1.80 11.49 0.28 0.003 adhE(0002) 7 cat1(0004)-sucD(0035) cat2(0034)-4hbd(0036) 1.09 1.29 5.63 2.15 0.461 1.38 6.66 2.30 0.520 8 cat1(0004)-sucD(0008N) cat2(0034)-4hbd(0036) 1.81 2.01 11.28 0.02 0.000 2.24 11.13 0.02 0.000 9 adhE(0002)-cat1(0004)- cat2(0034)-4hbd(0036) 0.24 1.99 2.02 2.32 0.106 0.89 4.85 2.41 0.186 sucD(0035) 10 cat1(0004)-sucD(0035)- cat2(0034)-4hbd(0036) 0.98 1.17 5.30 2.08 0.569 1.33 6.15 2.14 0.640 adhE(0002) 11 adhE(0002)-cat1(0004)- cat2(0034)-4hbd(0036) 0.20 0.53 1.38 2.30 0.019 0.91 8.10 1.49 0.034 sucD(0008N) 12 cat1(0004)-sucD(0008N)- cat2(0034)-4hbd(0036) 2.14 2.73 12.07 0.16 0.000 3.10 11.79 0.17 0.002 adhE(0002) 13 vector only vector only 2.11 2.62 9.03 0.01 0.000 3.00 12.05 0.01 0.000
[0333] Analysis of BDO, 4-HB and Succinate by GCMS.
[0334] BDO, 4-HB and succinate in fermentation and cell culture samples were derivatized by silylation and quantitatively analyzed by GCMS using methods adapted from literature reports ((Simonov et al., J. Anal Chem. 59:965-971 (2004)). The developed method demonstrated good sensitivity down to 1 .mu.M, linearity up to at least 25 mM, as well as excellent selectivity and reproducibility.
[0335] Sample preparation was performed as follows: 1000 .mu.L filtered (0.2 .mu.m or 0.45 .mu.m syringe filters) samples, e.g. fermentation broth, cell culture or standard solutions, were dried down in a Speed Vac Concentrator (Savant SVC-100H) for approximately 1 hour at ambient temperature, followed by the addition of 20 .mu.L, 10 mM cyclohexanol solution, as an internal standard, in dimethylformamide. The mixtures were vortexed and sonicated in a water bath (Branson 3510) for 15 min to ensure homogeneity. 100 .mu.L silylation derivatization reagent, N,O-bis(trimethylsilyl)triflouro-acetimide (BSTFA) with 1% trimethylchlorosilane, was added, and the mixture was incubated at 70.degree. C. for 30 min. The derivatized samples were centrifuged for 5 min, and the clear solutions were directly injected into GCMS. All the chemicals and reagents were from Sigma-Aldrich, with the exception of BDO which was purchased from J.T. Baker.
[0336] GCMS was performed on an Agilent gas chromatograph 6890N, interfaced to a mass-selective detector (MSD) 5973N operated in electron impact ionization (EI) mode has been used for the analysis. A DB-5MS capillary column (J&W Scientific, Agilent Technologies), 30 m.times.0.25 mm i.d..times.0.25 .mu.m film thickness, was used. The GC was operated in a split injection mode introducing 1 .mu.L of sample at 20:1 split ratio. The injection port temperature was 250.degree. C. Helium was used as a carrier gas, and the flow rate was maintained at 1.0 mL/min. A temperature gradient program was optimized to ensure good resolution of the analytes of interest and minimum matrix interference. The oven was initially held at 80.degree. C. for 1 min, then ramped to 120.degree. C. at 2.degree. C./min, followed by fast ramping to 320.degree. C. at 100.degree. C./min and final hold for 6 min at 320.degree. C. The MS interface transfer line was maintained at 280.degree. C. The data were acquired using lowmass' MS tune settings and 30-400 m/z mass-range scan. The total analysis time was 29 min including 3 min solvent delay. The retention times corresponded to 5.2, 10.5, 14.0 and 18.2 min for BSTFA-derivatized cyclohexanol, BDO, 4-HB and succinate, respectively. For quantitative analysis, the following specific mass fragments were selected (extracted ion chromatograms): m/z 157 for internal standard cyclohexanol, 116 for BDO, and 147 for both 4-HB and succinate. Standard calibration curves were constructed using analyte solutions in the corresponding cell culture or fermentation medium to match sample matrix as close as possible. GCMS data were processed using Environmental Data Analysis ChemStation software (Agilent Technologies).
[0337] The results indicated that most of the 4-HB and BDO produced were labeled with .sup.13C (FIG. 6, right-hand sides). Mass spectra from a parallel culture grown in unlabeled glucose are shown for comparison (FIG. 6, left-hand sides). Note that the peaks seen are for fragments of the derivatized molecule containing different numbers of carbon atoms from the metabolite. The derivatization reagent also contributes some carbon and silicon atoms that naturally-occurring label distribution, so the results are not strictly quantitative.
[0338] Production of BDO from 4-HB Using Alternate Pathways.
[0339] The various alternate pathways were also tested for BDO production. This includes use of the native E. coli SucCD enzyme to convert succinate to succinyl-CoA (Table 13, rows 2-3), use of .alpha.-ketoglutarate decarboxylase in the .alpha.-ketoglutarate pathway (Table 13, row 4), and use of PTB/BK as an alternate means to generate the CoA-derivative of 4HB (Table 13, row 1). Strains were constructed containing plasmids expressing the genes indicated in Table 13, which encompass these variants. The results show that in all cases, production of 4-HB and BDO occurred (Table 13).
TABLE-US-00014 TABLE 13 Production of BDO, 4-HB, and succinate in recombinant E. coli strains genes for different BDO pathway variants, grown anaerobically in minimal medium supplemented with 20 g/L glucose, and harvested 24 hours after induction with 0.1 mM IPTG. Concentrations are given in mM. Genes on pZE13 Genes on pZA33 Succinate 4-HB BDO 0002 + 0004 + 0035 0020n - 0021n - 0036 0.336 2.91 0.230 0038 + 0035 0034 - 0036 0.814 2.81 0.126 0038 + 0035 0036 - 0034 0.741 2.57 0.114 0035 + 0032 0034 - 0036 5.01 0.538 0.154
Example III
Biosynthesis of 4-Hydroxybutanoic Acid, .gamma.-Butyrolactone and 1,4-Butanediol
[0340] This Example describes the biosynthetic production of 4-hydroxybutanoic acid, .gamma.-butyrolactone and 1,4-butanediol using fermentation and other bioprocesses.
[0341] Methods for the integration of the 4-HB fermentation step into a complete process for the production of purified GBL, 1,4-butanediol (BDO) and tetrahydrofuran (THF) are described below. Since 4-HB and GBL are in equilibrium, the fermentation broth will contain both compounds. At low pH this equilibrium is shifted to favor GBL. Therefore, the fermentation can operate at pH 7.5 or less, generally pH 5.5 or less. After removal of biomass, the product stream enters into a separation step in which GBL is removed and the remaining stream enriched in 4-HB is recycled. Finally, GBL is distilled to remove any impurities. The process operates in one of three ways: 1) fed-batch fermentation and batch separation; 2) fed-batch fermentation and continuous separation; 3) continuous fermentation and continuous separation. The first two of these modes are shown schematically in FIG. 7. The integrated fermentation procedures described below also are used for the BDO producing cells of the invention for biosynthesis of BDO and subsequent BDO family products.
[0342] Fermentation Protocol to Produce 4-HB/GBL (Batch):
[0343] The production organism is grown in a 10 L bioreactor sparged with an N.sub.2/CO.sub.2 mixture, using 5 L broth containing 5 g/L potassium phosphate, 2.5 g/L ammonium chloride, 0.5 g/L magnesium sulfate, and 30 g/L corn steep liquor, and an initial glucose concentration of 20 g/L. As the cells grow and utilize the glucose, additional 70% glucose is fed into the bioreactor at a rate approximately balancing glucose consumption. The temperature of the bioreactor is maintained at 30 degrees C. Growth continues for approximately 24 hours, until 4-HB reaches a concentration of between 20-200 g/L, with the cell density being between 5 and 10 g/L. The pH is not controlled, and will typically decrease to pH 3-6 by the end of the run. Upon completion of the cultivation period, the fermenter contents are passed through a cell separation unit (e.g., centrifuge) to remove cells and cell debris, and the fermentation broth is transferred to a product separations unit. Isolation of 4-HB and/or GBL would take place by standard separations procedures employed in the art to separate organic products from dilute aqueous solutions, such as liquid-liquid extraction using a water immiscible organic solvent (e.g., toluene) to provide an organic solution of 4-HB/GBL. The resulting solution is then subjected to standard distillation methods to remove and recycle the organic solvent and to provide GBL (boiling point 204-205.degree. C.) which is isolated as a purified liquid.
[0344] Fermentation Protocol to Produce 4-HB/GBL (Fully Continuous):
[0345] The production organism is first grown up in batch mode using the apparatus and medium composition described above, except that the initial glucose concentration is 30-50 g/L. When glucose is exhausted, feed medium of the same composition is supplied continuously at a rate between 0.5 L/hr and 1 L/hr, and liquid is withdrawn at the same rate. The 4-HB concentration in the bioreactor remains constant at 30-40 g/L, and the cell density remains constant between 3-5 g/L. Temperature is maintained at 30 degrees C., and the pH is maintained at 4.5 using concentrated NaOH and HCl, as required. The bioreactor is operated continuously for one month, with samples taken every day to assure consistency of 4-HB concentration. In continuous mode, fermenter contents are constantly removed as new feed medium is supplied. The exit stream, containing cells, medium, and products 4-HB and/or GBL, is then subjected to a continuous product separations procedure, with or without removing cells and cell debris, and would take place by standard continuous separations methods employed in the art to separate organic products from dilute aqueous solutions, such as continuous liquid-liquid extraction using a water immiscible organic solvent (e.g., toluene) to provide an organic solution of 4-HB/GBL. The resulting solution is subsequently subjected to standard continuous distillation methods to remove and recycle the organic solvent and to provide GBL (boiling point 204-205.degree. C.) which is isolated as a purified liquid.
[0346] GBL Reduction Protocol:
[0347] Once GBL is isolated and purified as described above, it will then be subjected to reduction protocols such as those well known in the art (references cited) to produce 1,4-butanediol or tetrahydrofuran (THF) or a mixture thereof. Heterogeneous or homogeneous hydrogenation catalysts combined with GBL under hydrogen pressure are well known to provide the products 1,4-butanediol or tetrahydrofuran (THF) or a mixture thereof. It is important to note that the 4-HB/GBL product mixture that is separated from the fermentation broth, as described above, may be subjected directly, prior to GBL isolation and purification, to these same reduction protocols to provide the products 1,4-butanediol or tetrahydrofuran or a mixture thereof. The resulting products, 1,4-butanediol and THF are then isolated and purified by procedures well known in the art.
[0348] Fermentation and Hydrogenation Protocol to Produce BDO or THF Directly (Batch):
[0349] Cells are grown in a 10 L bioreactor sparged with an N.sub.2/CO.sub.2 mixture, using 5 L broth containing 5 g/L potassium phosphate, 2.5 g/L ammonium chloride, 0.5 g/L magnesium sulfate, and 30 g/L corn steep liquor, and an initial glucose concentration of 20 g/L. As the cells grow and utilize the glucose, additional 70% glucose is fed into the bioreactor at a rate approximately balancing glucose consumption. The temperature of the bioreactor is maintained at 30 degrees C. Growth continues for approximately 24 hours, until 4-HB reaches a concentration of between 20-200 g/L, with the cell density being between 5 and 10 g/L. The pH is not controlled, and will typically decrease to pH 3-6 by the end of the run. Upon completion of the cultivation period, the fermenter contents are passed through a cell separation unit (e.g., centrifuge) to remove cells and cell debris, and the fermentation broth is transferred to a reduction unit (e.g., hydrogenation vessel), where the mixture 4-HB/GBL is directly reduced to either 1,4-butanediol or THF or a mixture thereof. Following completion of the reduction procedure, the reactor contents are transferred to a product separations unit. Isolation of 1,4-butanediol and/or THF would take place by standard separations procedures employed in the art to separate organic products from dilute aqueous solutions, such as liquid-liquid extraction using a water immiscible organic solvent (e.g., toluene) to provide an organic solution of 1,4-butanediol and/or THF. The resulting solution is then subjected to standard distillation methods to remove and recycle the organic solvent and to provide 1,4-butanediol and/or THF which are isolated as a purified liquids.
[0350] Fermentation and Hydrogenation Protocol to Produce BDO or THF Directly (Fully Continuous):
[0351] The cells are first grown up in batch mode using the apparatus and medium composition described above, except that the initial glucose concentration is 30-50 g/L. When glucose is exhausted, feed medium of the same composition is supplied continuously at a rate between 0.5 L/hr and 1 L/hr, and liquid is withdrawn at the same rate. The 4-HB concentration in the bioreactor remains constant at 30-40 g/L, and the cell density remains constant between 3-5 g/L. Temperature is maintained at 30 degrees C., and the pH is maintained at 4.5 using concentrated NaOH and HCl, as required. The bioreactor is operated continuously for one month, with samples taken every day to assure consistency of 4-HB concentration. In continuous mode, fermenter contents are constantly removed as new feed medium is supplied. The exit stream, containing cells, medium, and products 4-HB and/or GBL, is then passed through a cell separation unit (e.g., centrifuge) to remove cells and cell debris, and the fermentation broth is transferred to a continuous reduction unit (e.g., hydrogenation vessel), where the mixture 4-HB/GBL is directly reduced to either 1,4-butanediol or THF or a mixture thereof. Following completion of the reduction procedure, the reactor contents are transferred to a continuous product separations unit. Isolation of 1,4-butanediol and/or THF would take place by standard continuous separations procedures employed in the art to separate organic products from dilute aqueous solutions, such as liquid-liquid extraction using a water immiscible organic solvent (e.g., toluene) to provide an organic solution of 1,4-butanediol and/or THF. The resulting solution is then subjected to standard continuous distillation methods to remove and recycle the organic solvent and to provide 1,4-butanediol and/or THF which are isolated as a purified liquids.
[0352] Fermentation Protocol to Produce BDO Directly (Batch):
[0353] The production organism is grown in a 10 L bioreactor sparged with an N.sub.2/CO.sub.2 mixture, using 5 L broth containing 5 g/L potassium phosphate, 2.5 g/L ammonium chloride, 0.5 g/L magnesium sulfate, and 30 g/L corn steep liquor, and an initial glucose concentration of 20 g/L. As the cells grow and utilize the glucose, additional 70% glucose is fed into the bioreactor at a rate approximately balancing glucose consumption. The temperature of the bioreactor is maintained at 30 degrees C. Growth continues for approximately 24 hours, until BDO reaches a concentration of between 20-200 g/L, with the cell density generally being between 5 and 10 g/L. Upon completion of the cultivation period, the fermenter contents are passed through a cell separation unit (e.g., centrifuge) to remove cells and cell debris, and the fermentation broth is transferred to a product separations unit. Isolation of BDO would take place by standard separations procedures employed in the art to separate organic products from dilute aqueous solutions, such as liquid-liquid extraction using a water immiscible organic solvent (e.g., toluene) to provide an organic solution of BDO. The resulting solution is then subjected to standard distillation methods to remove and recycle the organic solvent and to provide BDO (boiling point 228-229.degree. C.) which is isolated as a purified liquid.
[0354] Fermentation Protocol to Produce BDO Directly (Fully Continuous):
[0355] The production organism is first grown up in batch mode using the apparatus and medium composition described above, except that the initial glucose concentration is 30-50 g/L. When glucose is exhausted, feed medium of the same composition is supplied continuously at a rate between 0.5 L/hr and 1 L/hr, and liquid is withdrawn at the same rate. The BDO concentration in the bioreactor remains constant at 30-40 g/L, and the cell density remains constant between 3-5 g/L. Temperature is maintained at 30 degrees C., and the pH is maintained at 4.5 using concentrated NaOH and HCl, as required. The bioreactor is operated continuously for one month, with samples taken every day to assure consistency of BDO concentration. In continuous mode, fermenter contents are constantly removed as new feed medium is supplied. The exit stream, containing cells, medium, and the product BDO, is then subjected to a continuous product separations procedure, with or without removing cells and cell debris, and would take place by standard continuous separations methods employed in the art to separate organic products from dilute aqueous solutions, such as continuous liquid-liquid extraction using a water immiscible organic solvent (e.g., toluene) to provide an organic solution of BDO. The resulting solution is subsequently subjected to standard continuous distillation methods to remove and recycle the organic solvent and to provide BDO (boiling point 228-229.degree. C.) which is isolated as a purified liquid (mpt 20.degree. C.).
Example IV
Exemplary BDO Pathways
[0356] This example describes exemplary enzymes and corresponding genes for 1,4-butandiol (BDO) synthetic pathways.
[0357] Exemplary BDO synthetic pathways are shown in FIGS. 8-13. The pathways depicted in FIGS. 8-13 are from common central metabolic intermediates to 1,4-butanediol. All transformations depicted in FIGS. 8-13 fall into the 18 general categories of transformations shown in Table 14. Below is described a number of biochemically characterized candidate genes in each category. Specifically listed are genes that can be applied to catalyze the appropriate transformations in FIGS. 9-13 when cloned and expressed in a host organism. The top three exemplary genes for each of the key steps in FIGS. 9-13 are provided in Tables 15-23 (see below). Exemplary genes were provided for the pathways depicted in FIG. 8 are described herein.
TABLE-US-00015 TABLE 14 Enzyme types required to convert common central metabolic intermediates into 1,4-butanediol. The first three digits of each label correspond to the first three Enzyme Commission number digits which denote the general type of transformation independent of substrate specificity. Label Function 1.1.1.a Oxidoreductase (ketone to hydroxyl or aldehyde to alcohol) 1.1.1.c Oxidoreductase (2 step, acyl-CoA to alcohol) 1.2.1.b Oxidoreductase (acyl-CoA to aldehyde) 1.2.1.c Oxidoreductase (2-oxo acid to acyl-CoA, decarboxylation) 1.2.1.d Oxidoreductase (phosphorylating/dephosphorylating) 1.3.1.a Oxidoreductase operating on CH--CH donors 1.4.1.a Oxidoreductase operating on amino acids 2.3.1.a Acyltransferase (transferring phosphate group) 2.6.1.a Aminotransferase 2.7.2.a Phosphotransferase, carboxyl group acceptor 2.8.3.a Coenzyme-A transferase 3.1.2.a Thiolester hydrolase (CoA specific) 4.1.1.a Carboxy-lyase 4.2.1.a Hydro-lyase 4.3.1.a Ammonia-lyase 5.3.3.a Isomerase 5.4.3.a Aminomutase 6.2.1.a Acid-thiol ligase
1.1.1.a--Oxidoreductase (Aldehyde to Alcohol or Ketone to Hydroxyl)
[0358] Aldehyde to alcohol. Exemplary genes encoding enzymes that catalyze the conversion of an aldehyde to alcohol, that is, alcohol dehydrogenase or equivalently aldehyde reductase, include alrA encoding a medium-chain alcohol dehydrogenase for C2-C14 (Tani et al. Appl. Environ. Microbiol. 66:5231-5235 (2000)), ADH2 from Saccharomyces cerevisiae (Atsumi et al. Nature 451:86-89 (2008)), yqhD from E. coli which has preference for molecules longer than C(3) (Sulzenbacher et al. Journal of Molecular Biology 342:489-502 (2004)), and bdh I and bdh II from C. acetobutylicum which converts butyryaldehyde into butanol (Walter et al. Journal of Bacteriology 174:7149-7158 (1992)). The protein sequences for each of these exemplary gene products, if available, can be found using the following GenBank accession numbers:
TABLE-US-00016 Gene Accession No. GI No. Organism alrA BAB12273.1 9967138 Acinetobacter sp. Strain M-1 ADH2 NP_014032.1 6323961 Saccharymyces cerevisiae yqhD NP_417484.1 16130909 Escherichia coli bdh I NP_349892.1 15896543 Clostridium acetobutylicum bdh II NP_349891.1 15896542 Clostridium acetobutylicum
[0359] Enzymes exhibiting 4-hydroxybutyrate dehydrogenase activity (EC 1.1.1.61) also fall into this category. Such enzymes have been characterized in Ralstonia eutropha (Bravo et al. J. Forensic Sci. 49:379-387 (2004), Clostridium kluyveri (Wolff et al. Protein Expr. Purif. 6:206-212 (1995)) and Arabidopsis thaliana (Breitkreuz et al. J. Biol. Chem. 278:41552-41556 (2003)).
TABLE-US-00017 Gene Accession No. GI No. Organism 4hbd YP_726053.1 113867564 Ralstonia eutropha H16 4hbd L21902.1 146348486 Clostridium kluyveri DSM 555 4hbd Q94B07 75249805 Arabidopsis thaliana
[0360] Another exemplary enzyme is 3-hydroxyisobutyrate dehydrogenase which catalyzes the reversible oxidation of 3-hydroxyisobutyrate to methylmalonate semialdehyde. This enzyme participates in valine, leucine and isoleucine degradation and has been identified in bacteria, eukaryotes, and mammals. The enzyme encoded by P84067 from Thermus thermophilus HB8 has been structurally characterized (Lokanath et al. J Mol Biol 352:905-17 (2005)). The reversibility of the human 3-hydroxyisobutyrate dehydrogenase was demonstrated using isotopically-labeled substrate (Manning et al. Biochem J 231:481-484 (1985)). Additional genes encoding this enzyme include 3hidh in Homo sapiens (Hawes et al. Methods Enzymol. 324:218-228 (2000)) and Oryctolagus cuniculus (Chowdhury et al. Biosci. Biotechnol Biochem. 60:2043-2047 (1996); Hawes et al. Methods Enzymol. 324:218-228 (2000)), mmsb in Pseudomonas aeruginosa, and dhat in Pseudomonas putida (Aberhart et al. J Chem. Soc. [Perkin 1] 6:1404-1406 (1979); Chowdhury et al. Biosci. Biotechnol Biochem. 67:438-441 (2003); Chowdhury et al. Biosci. Biotechnol Biochem. 60:2043-2047 (1996)).
TABLE-US-00018 Gene Accession No. GI No. Organism P84067 P84067 75345323 Thermus thermophilus mmsb P28811.1 127211 Pseudomonas aeruginosa dhat Q59477.1 2842618 Pseudomonas putida 3hidh P31937.2 12643395 Homo sapiens 3hidh P32185.1 416872 Oryctolagus cuniculus
[0361] Several 3-hydroxyisobutyrate dehydrogenase enzymes have also been shown to convert malonic semialdehyde to 3-hydroxyproprionic acid (3-HP). Three gene candidates exhibiting this activity are mmsB from Pseudomonas aeruginosa PAO1(62), mmsB from Pseudomonas putida KT2440 (Liao et al., US Publication 2005/0221466) and mmsB from Pseudomonas putida E23 (Chowdhury et al., Biosci. Biotechnol. Biochem. 60:2043-2047 (1996)). An enzyme with 3-hydroxybutyrate dehydrogenase activity in Alcaligenes faecalis M3A has also been identified (Gokam et al., U.S. Pat. No. 7,393,676; Liao et al., US Publication No. 2005/0221466). Additional gene candidates from other organisms including Rhodobacter spaeroides can be inferred by sequence similarity.
TABLE-US-00019 Gene Accession No. GI No. Organism mmsB AAA25892.1 151363 Pseudomonas aeruginosa mmsB NP_252259.1 15598765 Pseudomonas aeruginosa PAO1 mmsB NP_746775.1 26991350 Pseudomonas putida KT2440 mmsB JC7926 60729613 Pseudomonas putida E23 orfB1 AAL26884 16588720 Rhodobacter spaeroides
[0362] The conversion of malonic semialdehyde to 3-HP can also be accomplished by two other enzymes: NADH-dependent 3-hydroxypropionate dehydrogenase and NADPH-dependent malonate semialdehyde reductase. An NADH-dependent 3-hydroxypropionate dehydrogenase is thought to participate in beta-alanine biosynthesis pathways from propionate in bacteria and plants (Rathinasabapathi, B. Journal of Plant Pathology 159:671-674 (2002); Stadtman, E. R. J. Am. Chem. Soc. 77:5765-5766 (1955)). This enzyme has not been associated with a gene in any organism to date. NADPH-dependent malonate semialdehyde reductase catalyzes the reverse reaction in autotrophic CO.sub.2-fixing bacteria. Although the enzyme activity has been detected in Metallosphaera sedula, the identity of the gene is not known (Alber et al. J. Bacteriol. 188:8551-8559 (2006)).
[0363] Ketone to Hydroxyl.
[0364] There exist several exemplary alcohol dehydrogenases that convert a ketone to a hydroxyl functional group. Two such enzymes from E. coli are encoded by malate dehydrogenase (mdh) and lactate dehydrogenase (ldhA). In addition, lactate dehydrogenase from Ralstonia eutropha has been shown to demonstrate high activities on substrates of various chain lengths such as lactate, 2-oxobutyrate, 2-oxopentanoate and 2-oxoglutarate (Steinbuchel and. Schlegel, Eur. J. Biochem. 130:329-334 (1983)). Conversion of alpha-ketoadipate into alpha-hydroxyadipate can be catalyzed by 2-ketoadipate reductase, an enzyme reported to be found in rat and in human placenta (Suda et al. Arch. Biochem. Biophys. 176:610-620 (1976); Suda et al. Biochem. Biophys. Res. Commun. 77:586-591 (1977)). An additional candidate for this step is the mitochondrial 3-hydroxybutyrate dehydrogenase (bdh) from the human heart which has been cloned and characterized (Marks et al. J. Biol. Chem. 267:15459-15463 (1992)). This enzyme is a dehydrogenase that operates on a 3-hydroxyacid. Another exemplary alcohol dehydrogenase converts acetone to isopropanol as was shown in C. beijerinckii (Ismaiel et al. J. Bacteriol. 175:5097-5105 (1993)) and T. brockii (Lamed et al. Biochem. J. 195:183-190 (1981); Peretz and Burstein Biochemistry 28:6549-6555 (1989)).
TABLE-US-00020 Gene Accession No. GI No. Organism mdh AAC76268.1 1789632 Escherichia coli ldhA NP_415898.1 16129341 Escherichia coli ldh YP_725182.1 113866693 Ralstonia eutropha bdh AAA58352.1 177198 Homo sapiens adh AAA23199.2 60592974 Clostridium beijerinckii NRRL B593 adh P14941.1 113443 Thermoanaerobacter brockii HTD4
[0365] Exemplary 3-hydroxyacyl dehydrogenases which convert acetoacetyl-CoA to 3-hydroxybutyryl-CoA include hbd from C. acetobutylicum (Boynton et al. Journal of Bacteriology 178:3015-3024 (1996)), hbd from C. beijerinckii (Colby et al. Appl Environ. Microbiol 58:3297-3302 (1992)), and a number of similar enzymes from Metallosphaera sedula (Berg et al. Archaea. Science 318:1782-1786 (2007)).
TABLE-US-00021 Gene Accession No. GI No. Organism hbd NP_349314.1 15895965 Clostridium acetobutylicum hbd AAM14586.1 20162442 Clostridium beijerinckii Msed_1423 YP_001191505 146304189 Metallosphaera sedula Msed_0399 YP_001190500 146303184 Metallosphaera sedula Msed_0389 YP_001190490 146303174 Metallosphaera sedula Msed_1993 YP_001192057 146304741 Metallosphaera sedula
1.1.1.c--Oxidoredutase (2 Step, Acyl-CoA to Alcohol)
[0366] Exemplary 2-step oxidoreductases that convert an acyl-CoA to alcohol include those that transform substrates such as acetyl-CoA to ethanol (for example, adhE from E. coli (Kessler et al. FEBS. Lett. 281:59-63 (1991)) and butyryl-CoA to butanol (for example, adhE2 from C. acetobutylicum (Fontaine et al. J. Bacteriol. 184:821-830 (2002)). In addition to reducing acetyl-CoA to ethanol, the enzyme encoded by adhE in Leuconostoc mesenteroides has been shown to oxide the branched chain compound isobutyraldehyde to isobutyryl-CoA (Kazahaya et al. J. Gen. Appl. Microbiol. 18:43-55 (1972); Koo et al. Biotechnol Lett. 27:505-510 (2005)).
TABLE-US-00022 Gene Accession No. GI No. Organism adhE NP_415757.1 16129202 Escherichia coli adhE2 AAK09379.1 12958626 Clostridium acetobutylicum adhE AAV66076.1 55818563 Leuconostoc mesenteroides
[0367] Another exemplary enzyme can convert malonyl-CoA to 3-HP. An NADPH-dependent enzyme with this activity has characterized in Chloroflexus aurantiacus where it participates in the 3-hydroxypropionate cycle (Hugler et al., J. Bacteriol. 184:2404-2410 (2002); Strauss and Fuchs, Eur. J. Biochem. 215:633-643 (1993)). This enzyme, with a mass of 300 kDa, is highly substrate-specific and shows little sequence similarity to other known oxidoreductases (Hugler et al., J. Bacteriol. 184:2404-2410 (2002)). No enzymes in other organisms have been shown to catalyze this specific reaction; however there is bioinformatic evidence that other organisms may have similar pathways (Klatt et al., Environ. Microbiol. 9:2067-2078 (2007)). Enzyme candidates in other organisms including Roseiflexus castenholzii, Erythrobacter sp. NAP1 and marine gamma proteobacterium HTCC2080 can be inferred by sequence similarity.
TABLE-US-00023 Gene Accession No. GI No. Organism mcr AAS20429.1 42561982 Chloroflexus aurantiacus Rcas_2929 YP_001433009.1 156742880 Roseiflexus castenholzii NAP1_02720 ZP_01039179.1 85708113 Erythrobacter sp. NAP1 MGP2080_00535 ZP_01626393.1 119504313 Marine gamma proteobacterium HTCC2080
[0368] Longer chain acyl-CoA molecules can be reduced by enzymes such as the jojoba (Simmondsia chinensis) FAR which encodes an alcohol-forming fatty acyl-CoA reductase. Its overexpression in E. coli resulted in FAR activity and the accumulation of fatty alcohol (Metz et al. Plant Physiology 122:635-644) 2000)).
TABLE-US-00024 Gene Accession No. GI No. Organism FAR AAD38039.1 5020215 Simmondsia chinensis
1.2.1.b--Oxidoreductase (Acyl-CoA to Aldehyde)
[0369] Several acyl-CoA dehydrogenases are capable of reducing an acyl-CoA to its corresponding aldehyde. Exemplary genes that encode such enzymes include the Acinetobacter calcoaceticus acr1 encoding a fatty acyl-CoA reductase (Reiser and Somerville, J. Bacteriology 179:2969-2975 (1997)), the Acinetobacter sp. M-1 fatty acyl-CoA reductase (Ishige et al. Appl. Environ. Microbiol. 68:1192-1195 (2002)), and a CoA- and NADP-dependent succinate semialdehyde dehydrogenase encoded by the sucD gene in Clostridium kluyveri (Sohling and Gottschalk J Bacteriol 178:871-80 (1996); Sohling and Gottschalk J Bacteriol. 178:871-880 (1996)). SucD of P. gingivalis is another succinate semialdehyde dehydrogenase (Takahashi et al. J. Bacteriol. 182:4704-4710 (2000)). The enzyme acylating acetaldehyde dehydrogenase in Pseudomonas sp, encoded by bphG, is yet another as it has been demonstrated to oxidize and acylate acetaldehyde, propionaldehyde, butyraldehyde, isobutyraldehyde and formaldehyde (Powlowski et al. J Bacteriol. 175:377-385 (1993)).
TABLE-US-00025 Gene Accession No. GI No. Organism acr1 YP_047869.1 50086359 Acinetobacter calcoaceticus acr1 AAC45217 1684886 Acinetobacter baylyi acr1 BAB85476.1 18857901 Acinetobacter sp. Strain M-1 sucD P38947.1 730847 Clostridium kluyveri sucD NP_904963.1 34540484 Porphyromonas gingivalis bphG BAA03892.1 425213 Pseudomonas sp
[0370] An additional enzyme type that converts an acyl-CoA to its corresponding aldehyde is malonyl-CoA reductase which transforms malonyl-CoA to malonic semialdehyde. Malonyl-CoA reductase is a key enzyme in autotrophic carbon fixation via the 3-hydroxypropionate cycle in thermoacidophilic archael bacteria (Berg et al. Science 318:1782-1786 (2007); Thauer, R. K. Science 318:1732-1733 (2007)). The enzyme utilizes NADPH as a cofactor and has been characterized in Metallosphaera and Sulfolobus spp (Alber et al. J. Bacteriol. 188:8551-8559 (2006); Hugler et al. J. Bacteriol. 184:2404-2410 (2002)). The enzyme is encoded by Msed_0709 in Metallosphaera sedula (Alber et al. J. Bacteriol. 188:8551-8559 (2006); Berg et al. Science 318:1782-1786 (2007)). A gene encoding a malonyl-CoA reductase from Sulfolobus tokodaii was cloned and heterologously expressed in E. coli (Alber et al. J. Bacteriol. 188:8551-8559 (2006)). Although the aldehyde dehydrogenase functionality of these enzymes is similar to the bifunctional dehydrogenase from Chloroflexus aurantiacus, there is little sequence similarity. Both malonyl-CoA reductase enzyme candidates have high sequence similarity to aspartate-semialdehyde dehydrogenase, an enzyme catalyzing the reduction and concurrent dephosphorylation of aspartyl-4-phosphate to aspartate semialdehyde. Additional gene candidates can be found by sequence homology to proteins in other organisms including Sulfolobus solfataricus and Sulfolobus acidocaldarius.
TABLE-US-00026 Gene Accession No. GI No. Organism Msed_0709 YP_001190808.1 146303492 Metallosphaera sedula mcr NP_378167.1 15922498 Sulfolobus tokodaii asd-2 NP_343563.1 15898958 Sulfolobus solfataricus Saci_2370 YP_256941.1 70608071 Sulfolobus acidocaldarius
1.2.1.c--Oxidoreductase (2-Oxo Acid to Acyl-CoA, Decarboxylation)
[0371] Enzymes in this family include 1) branched-chain 2-keto-acid dehydrogenase, 2) alpha-ketoglutarate dehydrogenase, and 3) the pyruvate dehydrogenase multienzyme complex (PDHC). These enzymes are multi-enzyme complexes that catalyze a series of partial reactions which result in acylating oxidative decarboxylation of 2-keto-acids. Each of the 2-keto-acid dehydrogenase complexes occupies key positions in intermediary metabolism, and enzyme activity is typically tightly regulated (Fries et al. Biochemistry 42:6996-7002 (2003)). The enzymes share a complex but common structure composed of multiple copies of three catalytic components: alpha-ketoacid decarboxylase (E1), dihydrolipoamide acyltransferase (E2) and dihydrolipoamide dehydrogenase (E3). The E3 component is shared among all 2-keto-acid dehydrogenase complexes in an organism, while the E1 and E2 components are encoded by different genes. The enzyme components are present in numerous copies in the complex and utilize multiple cofactors to catalyze a directed sequence of reactions via substrate channeling. The overall size of these dehydrogenase complexes is very large, with molecular masses between 4 and 10 million Da (that is, larger than a ribosome).
[0372] Activity of enzymes in the 2-keto-acid dehydrogenase family is normally low or limited under anaerobic conditions in E. coli. Increased production of NADH (or NADPH) could lead to a redox-imbalance, and NADH itself serves as an inhibitor to enzyme function. Engineering efforts have increased the anaerobic activity of the E. coli pyruvate dehydrogenase complex (Kim et al. Appl. Environ. Microbiol. 73:1766-1771 (2007); Kim et al. J. Bacteriol. 190:3851-3858) 2008); Zhou et al. Biotechnol. Lett. 30:335-342 (2008)). For example, the inhibitory effect of NADH can be overcome by engineering an H322Y mutation in the E3 component (Kim et al. J. Bacteriol. 190:3851-3858 (2008)). Structural studies of individual components and how they work together in complex provide insight into the catalytic mechanisms and architecture of enzymes in this family (Aevarsson et al. Nat. Struct. Biol. 6:785-792 (1999); Zhou et al. Proc. Natl. Acad. Sci. U.S.A. 98:14802-14807 (2001)). The substrate specificity of the dehydrogenase complexes varies in different organisms, but generally branched-chain keto-acid dehydrogenases have the broadest substrate range.
[0373] Alpha-ketoglutarate dehydrogenase (AKGD) converts alpha-ketoglutarate to succinyl-CoA and is the primary site of control of metabolic flux through the TCA cycle (Hansford, R. G. Curr. Top. Bioenerg. 10:217-278 (1980)). Encoded by genes sucA, sucB and lpd in E. coli, AKGD gene expression is downregulated under anaerobic conditions and during growth on glucose (Park et al. Mol. Microbiol. 15:473-482 (1995)). Although the substrate range of AKGD is narrow, structural studies of the catalytic core of the E2 component pinpoint specific residues responsible for substrate specificity (Knapp et al. J. Mol. Biol. 280:655-668 (1998)). The Bacillus subtilis AKGD, encoded by odhAB (E1 and E2) and pdhD (E3, shared domain), is regulated at the transcriptional level and is dependent on the carbon source and growth phase of the organism (Resnekov et al. Mol. Gen. Genet. 234:285-296 (1992)). In yeast, the LPD1 gene encoding the E3 component is regulated at the transcriptional level by glucose (Roy and Dawes J. Gen. Microbiol. 133:925-933 (1987)). The E1 component, encoded by KGD1, is also regulated by glucose and activated by the products of HAP2 and HAP3 (Repetto and Tzagoloff Mol. Cell Biol. 9:2695-2705 (1989)). The AKGD enzyme complex, inhibited by products NADH and succinyl-CoA, is well-studied in mammalian systems, as impaired function of has been linked to several neurological diseases (Tretter and dam-Vizi Philos. Trans. R. Soc. Lond B Biol. Sci. 360:2335-2345 (2005)).
TABLE-US-00027 Gene Accession No. GI No. Organism sucA NP_415254.1 16128701 Escherichia coli str. K12 substr. MG1655 sucB NP_415255.1 16128702 Escherichia coli str. K12 substr. MG1655 lpd NP_414658.1 16128109 Escherichia coli str. K12 substr. MG1655 odhA P23129.2 51704265 Bacillus subtilis odhB P16263.1 129041 Bacillus subtilis pdhD P21880.1 118672 Bacillus subtilis KGD1 NP_012141.1 6322066 Saccharomyces cerevisiae KGD2 NP_010432.1 6320352 Saccharomyces cerevisiae LPD1 NP_116635.1 14318501 Saccharomyces cerevisiae
[0374] Branched-chain 2-keto-acid dehydrogenase complex (BCKAD), also known as 2-oxoisovalerate dehydrogenase, participates in branched-chain amino acid degradation pathways, converting 2-keto acids derivatives of valine, leucine and isoleucine to their acyl-CoA derivatives and CO.sub.2. The complex has been studied in many organisms including Bacillus subtilis (Wang et al. Eur. J. Biochem. 213:1091-1099 (1993)), Rattus norvegicus (Namba et al. J. Biol. Chem. 244:4437-4447 (1969)) and Pseudomonas putida (Sokatch J. Bacteriol. 148:647-652 (1981)). In Bacillus subtilis the enzyme is encoded by genes pdhD (E3 component), bfmBB (E2 component), bfmBAA and bfmBAB (E1 component) (Wang et al. Eur. J. Biochem. 213:1091-1099 (1993)). In mammals, the complex is regulated by phosphorylation by specific phosphatases and protein kinases. The complex has been studied in rat hepatocites (Chicco et al. J. Biol. Chem. 269:19427-19434 (1994)) and is encoded by genes Bckdha (E1 alpha), Bckdhb (E1 beta), Dbt (E2), and Dld (E3). The E1 and E3 components of the Pseudomonas putida BCKAD complex have been crystallized (Aevarsson et al. Nat. Struct. Biol. 6:785-792 (1999); Mattevi Science 255:1544-1550 (1992)) and the enzyme complex has been studied (Sokatch et al. J. Bacteriol. 148:647-652 (1981)). Transcription of the P. putida BCKAD genes is activated by the gene product of bkdR (Hester et al. Eur. J. Biochem. 233:828-836 (1995)). In some organisms including Rattus norvegicus (Paxton et al. Biochem. J. 234:295-303 (1986)) and Saccharomyces cerevisiae (Sinclair et al. Biochem. Mol. Biol. Int. 31:911-922 (1993)), this complex has been shown to have a broad substrate range that includes linear oxo-acids such as 2-oxobutanoate and alpha-ketoglutarate, in addition to the branched-chain amino acid precursors. The active site of the bovine BCKAD was engineered to favor alternate substrate acetyl-CoA (Meng and Chuang, Biochemistry 33:12879-12885 (1994)).
TABLE-US-00028 Gene Accession No. GI No. Organism bfmBB NP_390283.1 16079459 Bacillus subtilis bfmBAA NP_390285.1 16079461 Bacillus subtilis bfmBAB NP_390284.1 16079460 Bacillus subtilis pdhD P21880.1 118672 Bacillus subtilis lpdV P09063.1 118677 Pseudomonas putida bkdB P09062.1 129044 Pseudomonas putida bkdA1 NP_746515.1 26991090 Pseudomonas putida bkdA2 NP_746516.1 26991091 Pseudomonas putida Bckdha NP_036914.1 77736548 Rattus norvegicus Bckdhb NP_062140.1 158749538 Rattus norvegicus Dbt NP_445764.1 158749632 Rattus norvegicus Dld NP_955417.1 40786469 Rattus norvegicus
[0375] The pyruvate dehydrogenase complex, catalyzing the conversion of pyruvate to acetyl-CoA, has also been extensively studied. In the E. coli enzyme, specific residues in the E1 component are responsible for substrate specificity (Bisswanger, H. J Biol Chem. 256:815-822 (1981); Bremer, J. Eur. J Biochem. 8:535-540 (1969); Gong et al. J Biol Chem. 275:13645-13653 (2000)). As mentioned previously, enzyme engineering efforts have improved the E. coli PDH enzyme activity under anaerobic conditions (Kim et al. Appl. Environ. Microbiol. 73:1766-1771 (2007); Kim J. Bacteriol. 190:3851-3858 (2008); Zhou et al. Biotechnol. Lett. 30:335-342 (2008)). In contrast to the E. coli PDH, the B. subtilis complex is active and required for growth under anaerobic conditions (Nakano J. Bacteriol. 179:6749-6755 (1997)). The Klebsiella pneumoniae PDH, characterized during growth on glycerol, is also active under anaerobic conditions (Menzel et al. J. Biotechnol. 56:135-142 (1997)). Crystal structures of the enzyme complex from bovine kidney (Zhou et al. Proc. Natl. Acad. Sci. U.S.A. 98:14802-14807 (2001)) and the E2 catalytic domain from Azotobacter vinelandii are available (Mattevi et al. Science 255:1544-1550 (1992)). Some mammalian PDH enzymes complexes can react on alternate substrates such as 2-oxobutanoate, although comparative kinetics of Rattus norvegicus PDH and BCKAD indicate that BCKAD has higher activity on 2-oxobutanoate as a substrate (Paxton et al. Biochem. J. 234:295-303 (1986)).
TABLE-US-00029 Gene Accession No. GI No. Organism aceE NP_414656.1 16128107 Escherichia coli str. K12 substr. MG1655 aceF NP_414657.1 16128108 Escherichia coli str. K12 substr. MG1655 lpd NP_414658.1 16128109 Escherichia coli str. K12 substr. MG1655 pdhA P21881.1 3123238 Bacillus subtilis pdhB P21882.1 129068 Bacillus subtilis pdhC P21883.2 129054 Bacillus subtilis pdhD P21880.1 118672 Bacillus subtilis aceE YP_001333808.1 152968699 Klebsiella pneumonia MGH78578 aceF YP_001333809.1 152968700 Klebsiella pneumonia MGH78578 lpdA YP_001333810.1 152968701 Klebsiella pneumonia MGH78578 Pdha1 NP_001004072.2 124430510 Rattus norvegicus Pdha2 NP_446446.1 16758900 Rattus norvegicus Dlat NP_112287.1 78365255 Rattus norvegicus Dld NP_955417.1 40786469 Rattus norvegicus
[0376] As an alternative to the large multienzyme 2-keto-acid dehydrogenase complexes described above, some anaerobic organisms utilize enzymes in the 2-ketoacid oxidoreductase family (OFOR) to catalyze acylating oxidative decarboxylation of 2-keto-acids. Unlike the dehydrogenase complexes, these enzymes contain iron-sulfur clusters, utilize different cofactors, and use ferredoxin or flavodixin as electron acceptors in lieu of NAD(P)H. While most enzymes in this family are specific to pyruvate as a substrate (POR) some 2-keto-acid:ferredoxin oxidoreductases have been shown to accept a broad range of 2-ketoacids as substrates including alpha-ketoglutarate and 2-oxobutanoate (Fukuda and Wakagi Biochim. Biophys. Acta 1597:74-80 (2002); Zhang et al. J. Biochem. 120:587-599 (1996)). One such enzyme is the OFOR from the thermoacidophilic archaeon Sulfolobus tokodaii 7, which contains an alpha and beta subunit encoded by gene ST2300 (Fukuda and Wakagi Biochim. Biophys. Acta 1597:74-80 (2002); Zhang et al. J. Biochem. 120:587-599 (1996)). A plasmid-based expression system has been developed for efficiently expressing this protein in E. coli (Fukuda et al. Eur. J. Biochem. 268:5639-5646 (2001)) and residues involved in substrate specificity were determined (Fukuda and Wakagi Biochim. Biophys. Acta 1597:74-80 (2002)). Two OFORs from Aeropyrum pernix str. K1 have also been recently cloned into E. coli, characterized, and found to react with a broad range of 2-oxoacids (Nishizawa et al. FEBS Lett. 579:2319-2322 (2005)). The gene sequences of these OFOR candidates are available, although they do not have GenBank identifiers assigned to date. There is bioinformatic evidence that similar enzymes are present in all archaea, some anaerobic bacteria and amitochondrial eukarya (Fukuda and Wakagi Biochim. Biophys. Acta 1597:74-80 (2005)). This class of enzyme is also interesting from an energetic standpoint, as reduced ferredoxin could be used to generate NADH by ferredoxin-NAD reductase (Petitdemange et al. Biochim. Biophys. Acta 421:334-337 (1976)). Also, since most of the enzymes are designed to operate under anaerobic conditions, less enzyme engineering may be required relative to enzymes in the 2-keto-acid dehydrogenase complex family for activity in an anaerobic environment.
TABLE-US-00030 Gene Accession No. GI No. Organism ST2300 NP_378302.1 15922633 Sulfolobus tokodaii 7
1.2.1.d--Oxidoreductase (Phosphorylating/Dephosphorylating)
[0377] Exemplary enzymes in this class include glyceraldehyde 3-phosphate dehydrogenase which converts glyceraldehyde-3-phosphate into D-glycerate 1,3-bisphosphate (for example, E. coli gapA (Branlant and Branlant Eur. J. Biochem. 150:61-66(1985)), aspartate-semialdehyde dehydrogenase which converts L-aspartate-4-semialdehyde into L-4-aspartyl-phosphate (for example, E. coli asd (Biellmann et al. Eur. J. Biochem. 104:53-58 (1980)), N-acetyl-gamma-glutamyl-phosphate reductase which converts N-acetyl-L-glutamate-5-semialdehyde into N-acetyl-L-glutamyl-5-phosphate (for example, E. coli argC (Parsot et al. Gene 68:275-283 (1988)), and glutamate-5-semialdehyde dehydrogenase which converts L-glutamate-5-semialdehyde into L-glutamyl-5-phosphate (for example, E. coli proA (Smith et al. J. Bacteriol. 157:545-551 (1984)).
TABLE-US-00031 Gene Accession No. GI No. Organism gapA P0A9B2.2 71159358 Escherichia coli asd NP_417891.1 16131307 Escherichia coli argC NP_418393.1 16131796 Escherichia coli proA NP_414778.1 16128229 Escherichia coli
1.3.1.a--Oxidoreductase Operating on CH--CH Donors
[0378] An exemplary enoyl-CoA reductase is the gene product of bcd from C. acetobutylicum (Atsumi et al. Metab Eng (2007); Boynton et al. Journal of Bacteriology 178:3015-3024 (1996), which naturally catalyzes the reduction of crotonyl-CoA to butyryl-CoA. Activity of this enzyme can be enhanced by expressing bcd in conjunction with expression of the C. acetobutylicum etfAB genes, which encode an electron transfer flavoprotein. An additional candidate for the enoyl-CoA reductase step is the mitochondrial enoyl-CoA reductase from E. gracilis (Hoffmeister et al. Journal of Biological Chemistry 280:4329-4338 (2005)). A construct derived from this sequence following the removal of its mitochondrial targeting leader sequence was cloned in E. coli resulting in an active enzyme (Hoffmeister et al., supra, (2005)). This approach is well known to those skilled in the art of expressing eukaryotic genes, particularly those with leader sequences that may target the gene product to a specific intracellular compartment, in prokaryotic organisms. A close homolog of this gene, TDE0597, from the prokaryote Treponema denticola represents a third enoyl-CoA reductase which has been cloned and expressed in E. coli (Tucci and Martin FEBS Letters 581:1561-1566 (2007)).
TABLE-US-00032 Gene Accession No. GI No. Organism bcd NP_349317.1 15895968 Clostridium acetobutylicum etfA NP_349315.1 15895966 Clostridium acetobutylicum etfB NP_349316.1 15895967 Clostridium acetobutylicum TER Q5EU90.1 62287512 Euglena gracilis TDE0597 NP_971211.1 42526113 Treponema denticola
[0379] Exemplary 2-enoate reductase (EC 1.3.1.31) enzymes are known to catalyze the NADH-dependent reduction of a wide variety of .alpha., .beta.-unsaturated carboxylic acids and aldehydes (Rohdich et al. J. Biol. Chem. 276:5779-5787 (2001)). 2-Enoate reductase is encoded by enr in several species of Clostridia (Giesel and Simon Arch Microbiol. 135(1): p. 51-57 (2001) including C. tyrobutyricum, and C. thermoaceticum (now called Moorella thermoaceticum) (Rohdich et al., supra, (2001)). In the recently published genome sequence of C. kluyveri, 9 coding sequences for enoate reductases have been reported, out of which one has been characterized (Seedorf et al. Proc Natl Acad Sci U.S.A 105(6):2128-33 (2008)). The enr genes from both C. tyrobutyricum and C. thermoaceticum have been cloned and sequenced and show 59% identity to each other. The former gene is also found to have approximately 75% similarity to the characterized gene in C. kluyveri (Giesel and Simon Arch Microbiol 135(1):51-57 (1983)). It has been reported based on these sequence results that enr is very similar to the dienoyl CoA reductase in E. coli (fadH) (163 Rohdich et al., supra (2001)). The C. thermoaceticum enr gene has also been expressed in an enzymatically active form in E. coli (163 Rohdich et al., supra (2001)).
TABLE-US-00033 Gene Accession No. GI No. Organism fadH NP_417552.1 16130976 Escherichia coli enr ACA54153.1 169405742 Clostridium botulinum A3 str enr CAA71086.1 2765041 Clostridium tyrobutyricum enr CAA76083.1 3402834 Clostridium kluyveri enr YP_430895.1 83590886 Moorella thermoacetica
1.4.1.a--Oxidoreductase Operating on Amino Acids
[0380] Most oxidoreductases operating on amino acids catalyze the oxidative deamination of alpha-amino acids with NAD+ or NADP+ as acceptor. Exemplary oxidoreductases operating on amino acids include glutamate dehydrogenase (deaminating), encoded by gdhA, leucine dehydrogenase (deaminating), encoded by ldh, and aspartate dehydrogenase (deaminating), encoded by nadX. The gdhA gene product from Escherichia coli (Korber et al. J. Mol. Biol. 234:1270-1273 (1993); McPherson and Wootton Nucleic. Acids Res. 11:5257-5266 (1983)), gdh from Thermotoga maritima (Kort et al. Extremophiles 1:52-60 (1997); Lebbink, et al. J. Mol. Biol. 280:287-296 (1998)); Lebbink et al. J. Mol. Biol. 289:357-369 (1999)), and gdhA1 from Halobacterium salinarum (Ingoldsby et al. Gene 349:237-244 (2005)) catalyze the reversible interconversion of glutamate to 2-oxoglutarate and ammonia, while favoring NADP(H), NAD(H), or both, respectively. The ldh gene of Bacillus cereus encodes the LeuDH protein that has a wide of range of substrates including leucine, isoleucine, valine, and 2-aminobutanoate (Ansorge and Kula Biotechnol Bioeng. 68:557-562 (2000); Stoyan et al. J. Biotechnol 54:77-80 (1997)). The nadX gene from Thermotoga maritime encoding for the aspartate dehydrogenase is involved in the biosynthesis of NAD (Yang et al. J. Biol. Chem. 278:8804-8808 (2003)).
TABLE-US-00034 Gene Accession No. GI No. Organism gdhA P00370 118547 Escherichia coli gdh P96110.4 6226595 Thermotoga maritima gdhA1 NP_279651.1 15789827 Halobacterium salinarum ldh P0A393 61222614 Bacillus cereus nadX NP_229443.1 15644391 Thermotoga maritima
[0381] The lysine 6-dehydrogenase (deaminating), encoded by lysDH gene, catalyze the oxidative deamination of the .epsilon.-amino group of L-lysine to form 2-aminoadipate-6-semialdehyde, which in turn nonenzymatically cyclizes to form .DELTA.1-piperideine-6-carboxylate (Misono and Nagasaki J. Bacteriol. 150:398-401 (1982)). The lysDH gene from Geobacillus stearothermophilus encodes a thermophilic NAD-dependent lysine 6-dehydrogenase (Heydari et al. Appl Environ. Microbiol 70:937-942 (2004)). In addition, the lysDH gene from Aeropyrum pernix K1 is identified through homology from genome projects.
TABLE-US-00035 Gene Accession No. GI No. Organism lysDH AB052732 13429872 Geobacillus stearothermophilus lysDH NP_147035.1 14602185 Aeropyrum pernix K1 ldh P0A393 61222614 Bacillus cereus
2.3.1.a--Acyltransferase (Transferring Phosphate Group)
[0382] Exemplary phosphate transferring acyltransferases include phosphotransacetylase, encoded by pta, and phosphotransbutyrylase, encoded by ptb. The pta gene from E. coli encodes an enzyme that can convert acetyl-CoA into acetyl-phosphate, and vice versa (Suzuki, T. Biochim. Biophys. Acta 191:559-569 (1969)). This enzyme can also utilize propionyl-CoA instead of acetyl-CoA forming propionate in the process (Hesslinger et al. Mol. Microbiol 27:477-492 (1998)). Similarly, the ptb gene from C. acetobutylicum encodes an enzyme that can convert butyryl-CoA into butyryl-phosphate (Walter et al. Gene 134(1): p. 107-11 (1993)); Huang et al. J Mol Microbiol Biotechnol 2(1): p. 33-38 (2000). Additional ptb genes can be found in butyrate-producing bacterium L2-50 (Louis et al. J. Bacteriol. 186:2099-2106 (2004)) and Bacillus megaterium (Vazquez et al. Curr. Microbiol 42:345-349 (2001)).
TABLE-US-00036 Gene Accession No. GI No. Organism pta NP_416800.1 16130232 Escherichia coli ptb NP_349676 15896327 Clostridium acetobutylicum ptb AAR19757.1 38425288 butyrate-producing bacterium L2-50 ptb CAC07932.1 10046659 Bacillus megaterium
2.6.1.a--Aminotransferase
[0383] Aspartate aminotransferase transfers an amino group from aspartate to alpha-ketoglutarate, forming glutamate and oxaloacetate. This conversion is catalyzed by, for example, the gene products of aspC from Escherichia coli (Yagi et al. FEBS Lett. 100:81-84 (1979); Yagi et al. Methods Enzymol. 113:83-89 (1985)), AAT2 from Saccharomyces cerevisiae (Yagi et al. J Biochem. 92:35-43 (1982)) and ASPS from Arabidopsis thaliana (48, 108, 225 48. de la et al. Plant J 46:414-425 (2006); Kwok and Hanson J Exp. Bot. 55:595-604 (2004); Wilkie and Warren Protein Expr. Purif. 12:381-389 (1998)). Valine aminotransferase catalyzes the conversion of valine and pyruvate to 2-ketoisovalerate and alanine. The E. coli gene, avtA, encodes one such enzyme (Whalen and Berg J. Bacteriol. 150:739-746 (1982)). This gene product also catalyzes the amination of .alpha.-ketobutyrate to generate .alpha.-aminobutyrate, although the amine donor in this reaction has not been identified (Whalen and Berg J. Bacteriol. 158:571-574 (1984)). The gene product of the E. coli serC catalyzes two reactions, phosphoserine aminotransferase and phosphohydroxythreonine aminotransferase (Lam and Winkler J. Bacteriol. 172:6518-6528 (1990)), and activity on non-phosphorylated substrates could not be detected (Drewke et al. FEBS. Lett. 390:179-182 (1996)).
TABLE-US-00037 Gene Accession No. GI No. Organism aspC NP_415448.1 16128895 Escherichia coli AAT2 P23542.3 1703040 Saccharomyces cerevisiae ASP5 P46248.2 20532373 Arabidopsis thaliana avtA YP_026231.1 49176374 Escherichia coli serC NP_415427.1 16128874 Escherichia coli
[0384] Cargill has developed a beta-alanine/alpha-ketoglutarate aminotransferase for producing 3-HP from beta-alanine via malonyl-semialdehyde (PCT/US2007/076252 (Jessen et al)). The gene product of SkPYD4 in Saccharomyces kluyveri was also shown to preferentially use beta-alanine as the amino group donor (Andersen et al. FEBS. J. 274:1804-1817 (2007)). SkUGA1 encodes a homologue of Saccharomyces cerevisiae GABA aminotransferase, UGA1 (Ramos et al. Eur. J. Biochem. 149:401-404 (1985)), whereas SkPYD4 encodes an enzyme involved in both .beta.-alanine and GABA transamination (Andersen et al. FEBS. J. 274:1804-1817 (2007)). 3-Amino-2-methylpropionate transaminase catalyzes the transformation from methylmalonate semialdehyde to 3-amino-2-methylpropionate. The enzyme has been characterized in Rattus norvegicus and Sus scrofa and is encoded by Abat (Kakimoto et al. Biochim. Biophys. Acta 156:374-380 (1968); Tamaki et al. Methods Enzymol. 324:376-389 (2000)). Enzyme candidates in other organisms with high sequence homology to 3-amino-2-methylpropionate transaminase include Gta-1 in C. elegans and gabT in Bacillus subtilus. Additionally, one of the native GABA aminotransferases in E. coli, encoded by gene gabT, has been shown to have broad substrate specificity (Liu et al. Biochemistry 43:10896-10905 (2004); Schulz et al. Appl Environ Microbiol 56:1-6 (1990)). The gene product of puuE catalyzes the other 4-aminobutyrate transaminase in E. coli (Kurihara et al. J. Biol. Chem. 280:4602-4608 (2005)).
TABLE-US-00038 Gene Accession No. GI No. Organism SkyPYD4 ABF58893.1 98626772 Saccharomyces kluyveri SkUGA1 ABF58894.1 98626792 Saccharomyces kluyveri UGA1 NP_011533.1 6321456 Saccharomyces cerevisiae Abat P50554.3 122065191 Rattus norvegicus Abat P80147.2 120968 Sus scrofa Gta-1 Q21217.1 6016091 Caenorhabditis elegans gabT P94427.1 6016090 Bacillus subtilus gabT P22256.1 120779 Escherichia coli K12 puuE NP_415818.1 16129263 Escherichia coli K12
[0385] The X-ray crystal structures of E. coli 4-aminobutyrate transaminase unbound and bound to the inhibitor were reported (Liu et al. Biochemistry 43:10896-10905 (2004)). The substrates binding and substrate specificities were studied and suggested. The roles of active site residues were studied by site-directed mutagenesis and X-ray crystallography (Liu et al. Biochemistry 44:2982-2992 (2005)). Based on the structural information, attempt was made to engineer E. coli 4-aminobutyrate transaminase with novel enzymatic activity. These studies provide a base for evolving transaminase activity for BDO pathways.
2.7.2.a--Phosphotransferase, Carboxyl Group Acceptor
[0386] Exemplary kinases include the E. coli acetate kinase, encoded by ackA (Skarstedt and Silverstein J. Biol. Chem. 251:6775-6783 (1976)), the C. acetobutylicum butyrate kinases, encoded by buk1 and buk2 (Walter et al. Gene 134(1):107-111 (1993) (Huang et al. J Mol Microbiol Biotechnol 2(1):33-38 (2000)], and the E. coli gamma-glutamyl kinase, encoded by proB (Smith et al. J. Bacteriol. 157:545-551 (1984)). These enzymes phosphorylate acetate, butyrate, and glutamate, respectively. The ackA gene product from E. coli also phosphorylates propionate (Hesslinger et al. Mol. Microbiol 27:477-492 (1998)).
TABLE-US-00039 Gene Accession No. GI No. Organism ackA NP_416799.1 16130231 Escherichia coli buk1 NP_349675 15896326 Clostridium acetobutylicum buk2 Q97II1 20137415 Clostridium acetobutylicum proB NP_414777.1 16128228 Escherichia coli
2.8.3.a--Coenzyme-A Transferase
[0387] In the CoA-transferase family, E. coli enzyme acyl-CoA:acetate-CoA transferase, also known as acetate-CoA transferase (EC 2.8.3.8), has been shown to transfer the CoA moiety to acetate from a variety of branched and linear acyl-CoA substrates, including isobutyrate (Matthies and Schink Appl Environ Microbiol 58:1435-1439 (1992)), valerate (Vanderwinkel et al. Biochem. Biophys. Res Commun. 33:902-908 (1968)) and butanoate (Vanderwinkel, supra (1968)). This enzyme is encoded by atoA (alpha subunit) and atoD (beta subunit) in E. coli sp. K12 (Korolev et al. Acta Crystallogr. D Biol Crystallogr. 58:2116-2121 (2002); Vanderwinkel, supra (1968)) and actA and cg0592 in Corynebacterium glutamicum ATCC 13032 (Duncan et al. Appl Environ Microbiol 68:5186-5190 (2002)). Additional genes found by sequence homology include atoD and atoA in Escherichia coli UT 189.
TABLE-US-00040 Gene Accession No. GI No. Organism atoA P76459.1 2492994 Escherichia coli K12 atoD P76458.1 2492990 Escherichia coli K12 actA YP_226809.1 62391407 Corynebacterium glutamicum ATCC 13032 cg0592 YP_224801.1 62389399 Corynebacterium glutamicum ATCC 13032 atoA ABE07971.1 91073090 Escherichia coli UT189 atoD ABE07970.1 91073089 Escherichia coli UT189
[0388] Similar transformations are catalyzed by the gene products of cat1, cat2, and cat3 of Clostridium kluyveri which have been shown to exhibit succinyl-CoA, 4-hydroxybutyryl-CoA, and butyryl-CoA acetyltransferase activity, respectively (Seedorf et al. Proc Natl Acad Sci U.S.A. 105(6):2128-2133 (2008); Sohling and Gottschalk J Bacteriol 178(3):871-880 (1996)].
TABLE-US-00041 Gene Accession No. GI No. Organism cat1 P38946.1 729048 Clostridium kluyveri cat2 P38942.2 1705614 Clostridium kluyveri cat3 EDK35586.1 146349050 Clostridium kluyveri
[0389] The glutaconate-CoA-transferase (EC 2.8.3.12) enzyme from anaerobic bacterium Acidaminococcus fermentans reacts with diacid glutaconyl-CoA and 3-butenoyl-CoA (Mack and Buckel FEBS Lett. 405:209-212 (1997)). The genes encoding this enzyme are gctA and gctB. This enzyme has reduced but detectable activity with other CoA derivatives including glutaryl-CoA, 2-hydroxyglutaryl-CoA, adipyl-CoA and acrylyl-CoA (Buckel et al. Eur. J. Biochem. 118:315-321 (1981)). The enzyme has been cloned and expressed in E. coli (Mac et al. Eur. J. Biochem. 226:41-51 (1994)).
TABLE-US-00042 Gene Accession No. GI No. Organism gctA CAA57199.1 559392 Acidaminococcus fermentans gctB CAA57200.1 559393 Acidaminococcus fermentans
3.1.2.a--Thiolester Hydrolase (CoA Specific)
[0390] In the CoA hydrolase family, the enzyme 3-hydroxyisobutyryl-CoA hydrolase is specific for 3-HIBCoA and has been described to efficiently catalyze the desired transformation during valine degradation (Shimomura et al. J Biol Chem 269:14248-14253 (1994)). Genes encoding this enzyme include hibch of Rattus norvegicus (Shimomura et al., supra (1994); Shimomura et al. Methods Enzymol. 324:229-240 (2000) and Homo sapiens (Shimomura et al., supra, 2000). Candidate genes by sequence homology include hibch of Saccharomyces cerevisiae and BC_2292 of Bacillus cereus.
TABLE-US-00043 Gene Accession No. GI No. Organism hibch Q5XIE6.2 146324906 Rattus norvegicus hibch Q6NVY1.2 146324905 Homo sapiens hibch P28817.2 2506374 Saccharomyces cerevisiae BC_2292 Q81DR3 81434808 Bacillus cereus
[0391] The conversion of adipyl-CoA to adipate can be carried out by an acyl-CoA hydrolase or equivalently a thioesterase. The top E. coli gene candidate is tesB (Naggert et al. J Biol Chem. 266(17):11044-11050 (1991)] which shows high similarity to the human acot8 which is a dicarboxylic acid acetyltransferase with activity on adipyl-CoA (Westin et al. J Biol Chem 280(46): 38125-38132 (2005). This activity has also been characterized in the rat liver (Deana, Biochem Int. 26(4): p. 767-773 (1992)).
TABLE-US-00044 Gene Accession No. GI No. Organism tesB NP_414986 16128437 Escherichia coli acot8 CAA15502 3191970 Homo sapiens acot8 NP_570112 51036669 Rattus norvegicus
[0392] Other potential E. coli thiolester hydrolases include the gene products of tesA (Bonner and Bloch, J Biol Chem. 247(10):3123-3133 (1972)), ybgC (Kuznetsova et al., FEMS Microbiol Rev. 29(2):263-279 (2005); Zhuang et al., FEBS Lett. 516(1-3):161-163 (2002)) paaI (Song et al., J Biol Chem. 281(16):11028-11038 (2006)), and ybdB (Leduc et al., J Bacteriol. 189(19):7112-7126 (2007)).
TABLE-US-00045 Gene Accession No. GI No. Organism tesA NP_415027 16128478 Escherichia coli ybgC NP_415264 16128711 Escherichia coli paaI NP_415914 16129357 Escherichia coli ybdB NP_415129 16128580 Escherichia coli
[0393] Several eukaryotic acetyl-CoA hydrolases (EC 3.1.2.1) have broad substrate specificity. The enzyme from Rattus norvegicus brain (Robinson et al. Biochem. Biophys. Res. Commun. 71:959-965 (1976)) can react with butyryl-CoA, hexanoyl-CoA and malonyl-CoA.
TABLE-US-00046 Gene Accession No. GI No. Organism acot12 NP_570103.1 18543355 Rattus norvegicus
4.1.1.a--Carboxy-Lyase
[0394] An exemplary carboxy-lyase is acetolactate decarboxylase which participates in citrate catabolism and branched-chain amino acid biosynthesis, converting 2-acetolactate to acetoin. In Lactococcus lactis the enzyme is composed of six subunits, encoded by gene aldB, and is activated by valine, leucine and isoleucine (Goupil et al. Appl. Environ. Microbiol. 62:2636-2640 (1996); Goupil-Feuillerat et al. J. Bacteriol. 182:5399-5408 (2000)). This enzyme has been overexpressed and characterized in E. coli (Phalip et al. FEBS Lett. 351:95-99 (1994)). In other organisms the enzyme is a dimer, encoded by aldC in Streptococcus thermophilus (Monnet et al. Lett. Appl. Microbiol. 36:399-405 (2003)), aldB in Bacillus brevis (Diderichsen et al. J. Bacteriol. 172:4315-4321 (1990); Najmudin et al. Acta Crystallogr. D. Biol. Crystallogr. 59:1073-1075 (2003)) and budA from Enterobacter aerogenes (Diderichsen et al. J. Bacteriol. 172:4315-4321 (1990)). The enzyme from Bacillus brevis was cloned and overexpressed in Bacillus subtilis and characterized crystallographically (Najmudin et al. Acta Crystallogr. D. Biol. Crystallogr. 59:1073-1075 (2003)). Additionally, the enzyme from Leuconostoc lactis has been purified and characterized but the gene has not been isolated (O'Sullivan et al. FEMS Microbiol. Lett. 194:245-249 (2001)).
TABLE-US-00047 Gene Accession No. GI No. Organism aldB NP_267384.1 15673210 Lactococcus lactis aldC Q8L208 75401480 Streptococcus thermophilus aldB P23616.1 113592 Bacillus brevis budA P05361.1 113593 Enterobacter aerogenes
[0395] Aconitate decarboxylase catalyzes the final step in itaconate biosynthesis in a strain of Candida and also in the filamentous fungus Aspergillus terreus (Bonnarme et al. J Bacteriol. 177:3573-3578 (1995); Willke and Vorlop Appl Microbiol Biotechnol 56:289-295 (2001)). Although itaconate is a compound of biotechnological interest, the aconitate decarboxylase gene or protein sequence has not been reported to date.
[0396] 4-oxalocronate decarboxylase has been isolated from numerous organisms and characterized. Genes encoding this enzyme include dmpH and dmpE in Pseudomonas sp. (strain 600) (Shingler et al. J Bacteriol. 174:711-724 (1992)), xylII and xylIII from Pseudomonas putida (Kato and Asano Arch. Microbiol 168:457-463 (1997); Lian and Whitman J. Am. Chem. Soc. 116:10403-10411 (1994); Stanley et al. Biochemistry 39:3514 (2000)) and Reut_B5691 and Reut_B5692 from Ralstonia eutropha JMP134 (Hughes et al. J Bacteriol. 158:79-83 (1984)). The genes encoding the enzyme from Pseudomonas sp. (strain 600) have been cloned and expressed in E. coli (Shingler et al. J Bacteriol. 174:711-724 (1992)).
TABLE-US-00048 Gene Accession No. GI No. Organism dmpH CAA43228.1 45685 Pseudomonas sp. CF600 dmpE CAA43225.1 45682 Pseudomonas sp. CF600 xylII YP_709328.1 111116444 Pseudomonas putida xylIII YP_709353.1 111116469 Pseudomonas putida Reut_B5691 YP_299880.1 73539513 Ralstonia eutropha JMP134 Reut_B5692 YP_299881.1 73539514 Ralstonia eutropha JMP134
[0397] An additional class of decarboxylases has been characterized that catalyze the conversion of cinnamate (phenylacrylate) and substituted cinnamate derivatives to the corresponding styrene derivatives. These enzymes are common in a variety of organisms and specific genes encoding these enzymes that have been cloned and expressed in E. coli are: pad 1 from Saccharomyces cerevisae (Clausen et al. Gene 142:107-112 (1994)), pdc from Lactobacillus plantarum (Barthelmebs et al. Appl Environ Microbiol 67:1063-1069 (2001); Qi et al. Metab Eng 9:268-276 (2007); Rodriguez et al. J. Agric. Food Chem. 56:3068-3072 (2008)), pofK (pad) from Klebsiella oxytoca (Hashidoko et al. Biosci. Biotech. Biochem. 58:217-218 (1994); Uchiyama et al. Biosci. Biotechnol. Biochem. 72:116-123 (2008)), Pedicoccus pentosaceus (Barthelmebs et al. Appl Environ Microbiol 67:1063-1069 (2001)), and padC from Bacillus subtilis and Bacillus pumilus (Lingen et al. Protein Eng 15:585-593 (2002)). A ferulic acid decarboxylase from Pseudomonas fluorescens also has been purified and characterized (Huang et al. J. Bacteriol. 176:5912-5918 (1994)). Importantly, this class of enzymes have been shown to be stable and do not require either exogenous or internally bound co-factors, thus making these enzymes ideally suitable for biotransformations (Sariaslani, Annu. Rev. Microbiol. 61:51-69 (2007)).
TABLE-US-00049 Accession Gene No. GI No. Organism pad1 AB368798 188496948 Saccharomyces cerevisae BAG32372.1 188496949 pdc U63827 1762615, 1762616 Lactobacillus plantarum AAC45282.1 pofK AB330293, 149941607, 149941608 Klebsiella oxytoca (pad) BAF65031.1 padC AF017117 2394281, 2394282 Bacillus subtilis AAC46254.1 pad AJ276891 11322456, 11322458 Pedicoccus pentosaceus CAC16794.1 pad AJ278683 11691809, 11691810 Bacillus pumilus CAC18719.1
[0398] Additional decarboxylase enzymes can form succinic semialdehyde from alpha-ketoglutarate. These include the alpha-ketoglutarate decarboxylase enzymes from Euglena gracilis (Shigeoka et al. Biochem. J. 282(Pt 2):319-323 (1992); Shigeoka and Nakano Arch. Biochem. Biophys. 288:22-28 (1991); Shigeoka and Nakano Biochem. J. 292 (Pt 2):463-467 (1993)), whose corresponding gene sequence has yet to be determined, and from Mycobacterium tuberculosis (Tian et al. Proc Natl Acad Sci U.S.A. 102:10670-10675 (2005)). In addition, glutamate decarboxylase enzymes can convert glutamate into 4-aminobutyrate such as the products of the E. coli gadA and gadB genes (De Biase et al. Protein. Expr. Purif. 8:430-438 (1993)).
TABLE-US-00050 Gene Accession No. GI No. Organism kgd O50463.4 160395583 Mycobacterium tuberculosis gadA NP_417974 16131389 Escherichia coli gadB NP_416010 16129452 Escherichia coli
Keto-Acid Decarboxylases
[0399] Pyruvate decarboxylase (PDC, EC 4.1.1.1), also termed keto-acid decarboxylase, is a key enzyme in alcoholic fermentation, catalyzing the decarboxylation of pyruvate to acetaldehyde. This enzyme has a broad substrate range for aliphatic 2-keto acids including 2-ketobutyrate, 2-ketovalerate, 3-hydroxypyruvate and 2-phenylpyruvate (Berg et al. Science 318:1782-1786 (2007)). The PDC from Zymomonas mobilus, encoded by pdc, has been a subject of directed engineering studies that altered the affinity for different substrates (Siegert et al. Protein Eng Des Sel 18:345-357 (2005)). The PDC from Saccharomyces cerevisiae has also been extensively studied, engineered for altered activity, and functionally expressed in E. coli (Killenberg-Jabs et al. Eur. J. Biochem. 268:1698-1704 (2001); Li and Jordan Biochemistry 38:10004-10012 (1999); ter Schure et al. Appl. Environ. Microbiol. 64:1303-1307 (1998)). The crystal structure of this enzyme is available (Killenberg-Jabs Eur. J. Biochem. 268:1698-1704 (2001)). Other well-characterized PDC candidates include the enzymes from Acetobacter pasteurians (Chandra et al. Arch. Microbiol. 176:443-451 (2001)) and Kluyveromyces lactis (Krieger et al. Eur. J. Biochem. 269:3256-3263 (2002)).
TABLE-US-00051 Gene Accession No. GI No. Organism pdc P06672.1 118391 Zymomonas mobilus pdc1 P06169 30923172 Saccharomyces cerevisiae pdc Q8L388 75401616 Acetobacter pasteurians pdc1 Q12629 52788279 Kluyveromyces lactis
[0400] Like PDC, benzoylformate decarboxylase (EC 4.1.1.7) has a broad substrate range and has been the target of enzyme engineering studies. The enzyme from Pseudomonas putida has been extensively studied and crystal structures of this enzyme are available (Hasson et al. Biochemistry 37:9918-9930 (1998); Polovnikova et al. Biochemistry 42:1820-1830 (2003)). Site-directed mutagenesis of two residues in the active site of the Pseudomonas putida enzyme altered the affinity (Km) of naturally and non-naturally occurring substrates (Siegert Protein Eng Des Sel 18:345-357 (2005)). The properties of this enzyme have been further modified by directed engineering (Lingen et al. Protein Eng 15:585-593 (2002)); Lingen Chembiochem 4:721-726 (2003)). The enzyme from Pseudomonas aeruginosa, encoded by mdlC, has also been characterized experimentally (Barrowman et al. FEMS Microbiology Letters 34:57-60 (1986)). Additional gene candidates from Pseudomonas stutzeri, Pseudomonas fluorescens and other organisms can be inferred by sequence homology or identified using a growth selection system developed in Pseudomonas putida (Henning et al. Appl. Environ. Microbiol. 72:7510-7517 (2006)).
TABLE-US-00052 Gene Accession No. GI No. Organism mdlC P20906.2 3915757 Pseudomonas putida mdlC Q9HUR2.1 81539678 Pseudomonas aeruginosa dpgB ABN80423.1 126202187 Pseudomonas stutzeri ilvB-1 YP_260581.1 70730840 Pseudomonas fluorescens
4.2.1.a--Hydro-Lyase
[0401] The 2-(hydroxymethyl)glutarate dehydratase of Eubacterium barkeri is an exemplary hydro-lyase. This enzyme has been studied in the context of nicotinate catabolism and is encoded by hmd (Alhapel et al. Proc Natl Acad Sci USA 103:12341-12346 (2006)). Similar enzymes with high sequence homology are found in Bacteroides capillosus, Anaerotruncus colihominis, and Natranaerobius thermophilius.
TABLE-US-00053 Gene Accession No. GI No. Organism hmd ABC88407.1 86278275 Eubacterium barkeri BACCAP_02294 ZP_02036683.1 154498305 Bacteroides capillosus ATCC 29799 ANACOL_02527 ZP_02443222.1 167771169 Anaerotruncus colihominis DSM 17241 NtherDRAFT_2368 ZP_02852366.1 169192667 Natranaerobius thermophilus JW/NM-WN- LF
[0402] A second exemplary hydro-lyase is fumarate hydratase, an enzyme catalyzing the dehydration of malate to fumarate. A wealth of structural information is available for this enzyme and researchers have successfully engineered the enzyme to alter activity, inhibition and localization (Weaver, T. Acta Crystallogr. D Biol Crystallogr. 61:1395-1401 (2005)). Additional fumarate hydratases include those encoded by fumC from Escherichia coli (Estevez et al. Protein Sci. 11:1552-1557 (2002); Hong and Lee Biotechnol. Bioprocess Eng. 9:252-255 (2004); Rose and Weaver Proc Natl Acad Sci USA 101:3393-3397 (2004)), Campylobacter jejuni (Smith et al. Int. J Biochem. Cell Biol 31:961-975 (1999)) and Thermus thermophilus (Mizobata et al. Arch. Biochem. Biophys. 355:49-55 (1998)), and fumH from Rattus norvegicus (Kobayashi et al. J Biochem. 89:1923-1931(1981)). Similar enzymes with high sequence homology include fum1 from Arabidopsis thaliana and fumC from Corynebacterium glutamicum.
TABLE-US-00054 Gene Accession No. GI No. Organism fumC P05042.1 120601 Escherichia coli K12 fumC O69294.1 9789756 Campylobacter jejuni fumC P84127 75427690 Thermus thermophilus fumH P14408.1 120605 Rattus norvegicus fum1 P93033.2 39931311 Arabidopsis thaliana fumC Q8NRN8.1 39931596 Corynebacterium glutamicum
[0403] Citramalate hydrolyase, also called 2-methylmalate dehydratase, converts 2-methylmalate to mesaconate. 2-Methylmalate dehydratase activity was detected in Clostridium tetanomorphum, Morganella morganii, Citrobacter amalonaticus in the context of the glutamate degradation VI pathway (Kato and Asano Arch. Microbiol 168:457-463 (1997)); however the genes encoding this enzyme have not been sequenced to date.
[0404] The gene product of crt from C. acetobutylicum catalyzes the dehydration of 3-hydroxybutyryl-CoA to crotonyl-CoA (Atsumi et al. Metab Eng.; 29 (2007)); Boynton et al. Journal of Bacteriology 178:3015-3024 (1996)). The enoyl-CoA hydratases, phaA and phaB, of P. putida are believed to carry out the hydroxylation of double bonds during phenylacetate catabolism; (Olivera et al. Proc Natl Acad Sci USA 95(11):6419-6424 (1998)). The paaA and paaB from P. fluorescens catalyze analogous transformations (14 Olivera et al., supra, 1998). Lastly, a number of Escherichia coli genes have been shown to demonstrate enoyl-CoA hydratase functionality including maoC (Park and Lee J Bacteriol 185(18):5391-5397 (2003)), paaF (Park and Lee Biotechnol Bioeng. 86(6):681-686 (2004a)); Park and Lee Appl Biochem Biotechnol. 113-116: 335-346 (2004b)); Ismail et al. Eur J Biochem 270(14):p. 3047-3054 (2003), and paaG (Park and Lee, supra, 2004; Park and Lee supra, 2004b; Ismail et al., supra, 2003).
TABLE-US-00055 Gene Accession No. GI No. Organism maoC NP_415905.1 16129348 Escherichia coli paaF NP_415911.1 16129354 Escherichia coli paaG NP_415912.1 16129355 Escherichia coli crt NP_349318.1 15895969 Clostridium acetobutylicum paaA NP_745427.1 26990002 Pseudomonas putida paaB NP_745426.1 26990001 Pseudomonas putida phaA ABF82233.1 106636093 Pseudomonas fluorescens phaB ABF82234.1 106636094 Pseudomonas fluorescens
[0405] The E. coli genes fadA and fadB encode a multienzyme complex that exhibits ketoacyl-CoA thiolase, 3-hydroxyacyl-CoA dehydrogenase, and enoyl-CoA hydratase activities (Yang et al. Biochemistry 30(27): p. 6788-6795 (1991); Yang et al. J Biol Chem 265(18): p. 10424-10429 (1990); Yang et al. J Biol Chem 266(24): p. 16255 (1991); Nakahigashi and Inokuchi Nucleic Acids Res 18(16): p. 4937 (1990)). The fadI and fadJ genes encode similar functions and are naturally expressed only anaerobically (Campbell et al. Mol Microbiol 47(3): p. 793-805 (2003). A method for producing poly[(R)-3-hydroxybutyrate] in E. coli that involves activating fadB (by knocking out a negative regulator, fadR) and co-expressing a non-native ketothiolase (phaA from Ralstonia eutropha) has been described previously (Sato et al. J Biosci Bioeng 103(1): 38-44 (2007)). This work clearly demonstrates that a .beta.-oxidation enzyme, in particular the gene product of fadB which encodes both 3-hydroxyacyl-CoA dehydrogenase and enoyl-CoA hydratase activities, can function as part of a pathway to produce longer chain molecules from acetyl-CoA precursors.
TABLE-US-00056 Gene Accession No. GI No. Organism fadA YP_026272.1 49176430 Escherichia coli fadB NP_418288.1 16131692 Escherichia coli fadI NP_416844.1 16130275 Escherichia coli fadJ NP_416843.1 16130274 Escherichia coli fadR NP_415705.1 16129150 Escherichia coli
4.3.1.a--Ammonia-Lyase
[0406] Aspartase (EC 4.3.1.1), catalyzing the deamination of aspartate to fumarate, is a widespread enzyme in microorganisms, and has been characterized extensively (Viola, R. E. Adv. Enzymol. Relat Areas Mol. Biol 74:295-341 (2000)). The crystal structure of the E. coli aspartase, encoded by aspA, has been solved (Shi et al. Biochemistry 36:9136-9144 (1997)). The E. coli enzyme has also been shown to react with alternate substrates aspartatephenylmethylester, asparagine, benzyl-aspartate and malate (Ma et al. Ann N.Y. Acad Sci 672:60-65 (1992)). In a separate study, directed evolution was been employed on this enzyme to alter substrate specificity (Asano et al. Biomol. Eng 22:95-101 (2005)). Enzymes with aspartase functionality have also been characterized in Haemophilus influenzae (Sjostrom et al. Biochim. Biophys. Acta 1324:182-190 (1997)), Pseudomonas fluorescens (Takagi et al. J. Biochem. 96:545-552 (1984)), Bacillus subtilus (Sjostrom et al. Biochim. Biophys. Acta 1324:182-190 (1997)) and Serratia marcescens (Takagi and Kisumi J Bacteriol. 161:1-6 (1985)).
TABLE-US-00057 Gene Accession No. GI No. Organism aspA NP_418562 90111690 Escherichia coli K12 subsp. MG1655 aspA P44324.1 1168534 Haemophilus influenzae aspA P07346.1 114273 Pseudomonas fluorescens ansB P26899.1 114271 Bacillus subtilus aspA P33109.1 416661 Serratia marcescens
[0407] 3-methylaspartase (EC 4.3.1.2), also known as beta-methylaspartase or 3-methylaspartate ammonia-lyase, catalyzes the deamination of threo-3-methylasparatate to mesaconate. The 3-methylaspartase from Clostridium tetanomorphum has been cloned, functionally expressed in E. coli, and crystallized (Asuncion et al. Acta Crystallogr. D Biol Crystallogr. 57:731-733 (2001); Asuncion et al. J Biol Chem. 277:8306-8311 (2002); Botting et al. Biochemistry 27:2953-2955 (1988); Goda et al. Biochemistry 31:10747-10756 (1992). In Citrobacter amalonaticus, this enzyme is encoded by BAA28709 (Kato and Asano Arch. Microbiol 168:457-463 (1997)). 3-Methylaspartase has also been crystallized from E. coli YG1002 (Asano and Kato FEMS Microbiol Lett. 118:255-258 (1994)) although the protein sequence is not listed in public databases such as GenBank. Sequence homology can be used to identify additional candidate genes, including CTC_02563 in C. tetani and ECs0761 in Escherichia coli O157:H7.
TABLE-US-00058 Gene Accession No. GI No. Organism MAL AAB24070.1 259429 Clostridium tetanomorphum BAA28709 BAA28709.1 3184397 Citrobacter amalonaticus CTC_02563 NP_783085.1 28212141 Clostridium tetani ECs0761 BAB34184.1 13360220 Escherichia coli O157:H7 str. Sakai
[0408] Ammonia-lyase enzyme candidates that form enoyl-CoA products include beta-alanyl-CoA ammonia-lyase (EC 4.3.1.6), which deaminates beta-alanyl-CoA, and 3-aminobutyryl-CoA ammonia-lyase (EC 4.3.1.14). Two beta-alanyl-CoA ammonia lyases have been identified and characterized in Clostridium propionicum (Herrmann et al. FEBS J. 272:813-821 (2005)). No other beta-alanyl-CoA ammonia lyases have been studied to date, but gene candidates can be identified by sequence similarity. One such candidate is MXAN_4385 in Myxococcus xanthus.
TABLE-US-00059 Gene Accession No. GI No. Organism ac12 CAG29275.1 47496504 Clostridium propionicum acl1 CAG29274.1 47496502 Clostridium propionicum MXAN_4385 YP_632558.1 108756898 Myxococcus xanthus
5.3.3.a--Isomerase
[0409] The 4-hydroxybutyryl-CoA dehydratases from both Clostridium aminobutyrium and C. kluyveri catalyze the reversible conversion of 4-hydroxybutyryl-CoA to crotonyl-CoA and posses an intrinsic vinylacetyl-CoA .DELTA.-isomerase activity (Scherf and Buckel Eur. J Biochem. 215:421-429 (1993); Scherf et al. Arch. Microbiol 161:239-245 (1994)). Both native enzymes were purified and characterized, including the N-terminal amino acid sequences (Scherf and Buckel, supra, 1993; Scherf et al., supra, 1994). The abfD genes from C. aminobutyrium and C. kluyveri match exactly with these N-terminal amino acid sequences, thus are encoding the 4-hydroxybutyryl-CoA dehydratases/vinylacetyl-CoA .DELTA.-isomerase. In addition, the abfD gene from Porphyromonas gingivalis ATCC 33277 is identified through homology from genome projects.
TABLE-US-00060 Gene Accession No. GI No. Organism abfD YP_001396399.1 153955634 Clostridium kluyveri DSM 555 abfD P55792 84028213 Clostridium aminobutyricum abfD YP_001928843 188994591 Porphyromonas gingivalis ATCC 33277
5.4.3.a--Aminomutase
[0410] Lysine 2,3-aminomutase (EC 5.4.3.2) is an exemplary aminomutase that converts lysine to (3S)-3,6-diaminohexanoate, shifting an amine group from the 2- to the 3-position. The enzyme is found in bacteria that ferment lysine to acetate and butyrate, including as Fusobacterium nuleatum (kamA) (Barker et al. J. Bacteriol. 152:201-207 (1982)) and Clostridium subterminale (kamA) (Chirpich et al. J. Biol. Chem. 245:1778-1789 (1970)). The enzyme from Clostridium subterminale has been crystallized (Lepore et al. Proc. Natl. Acad. Sci. U.S.A 102:13819-13824 (2005)). An enzyme encoding this function is also encoded by yodO in Bacillus subtilus (Chen et al. Biochem. J. 348 Pt 3:539-549 (2000)). The enzyme utilizes pyridoxal 5'-phosphate as a cofactor, requires activation by S-Adenosylmethoionine, and is stereoselective, reacting with the only with L-lysine. The enzyme has not been shown to react with alternate substrates.
TABLE-US-00061 Gene Accession No. GI No. Organism yodO O34676.1 4033499 Bacillus subtilus kamA Q9XBQ8.1 75423266 Clostridium subterminale kamA Q8RHX4 81485301 Fusobacterium nuleatum subsp. nuleatum
[0411] A second aminomutase, beta-lysine 5,6-aminomutase (EC 5.4.3.3), catalyzes the next step of lysine fermentation to acetate and butyrate, which transforms (3S)-3,6-diaminohexanoate to (3S,5S)-3,5-diaminohexanoate, shifting a terminal amine group from the 6- to the 5-position. This enzyme also catalyzes the conversion of lysine to 2,5-diaminohexanoate and is also called lysine-5,6-aminomutase (EC 5.4.3.4). The enzyme has been crystallized in Clostridium sticklandii (kamD, kamE) (Berkovitch et al. Proc. Natl. Acad. Sci. U.S.A 101:15870-15875 (2004)). The enzyme from Porphyromonas gingivalis has also been characterized (Tang et al. Biochemistry 41:8767-8776 (2002)).
TABLE-US-00062 Gene Accession No. GI No. Organism kamD AAC79717.1 3928904 Clostridium sticklandii kamE AAC79718.1 3928905 Clostridium sticklandii kamD NC_002950.2 34539880, 34540809 Porphyromonas gingivalis W83 kamE NC_002950.2 34539880, 34540810 Porphyromonas gingivalis W83
[0412] Ornithine 4,5-aminomutase (EC 5.4.3.5) converts D-ornithine to 2,4-diaminopentanoate, also shifting a terminal amine to the adjacent carbon. The enzyme from Clostridium sticklandii is encoded by two genes, oraE and oraS, and has been cloned, sequenced and expressed in E. coli (Chen et al. J. Biol. Chem. 276:44744-44750 (2001)). This enzyme has not been characterized in other organisms to date.
TABLE-US-00063 Gene Accession No. GI No. Organism oraE AAK72502 17223685 Clostridium sticklandii oraS AAK72501 17223684 Clostridium sticklandii
[0413] Tyrosine 2,3-aminomutase (EC 5.4.3.6) participates in tyrosine biosynthesis, reversibly converting tyrosine to 3-amino-3-(4-hydroxyphenyl)propanoate by shifting an amine from the 2- to the 3-position. In Streptomyces globisporus the enzyme has also been shown to react with tyrosine derivatives (Christenson et al. Biochemistry 42:12708-12718 (2003)). Sequence information is not available.
[0414] Leucine 2,3-aminomutase (EC 5.4.3.7) converts L-leucine to beta-leucine during leucine degradation and biosynthesis. An assay for leucine 2,3-aminomutase detected activity in many organisms (Poston, J. M. Methods Enzymol. 166:130-135 (1988)) but genes encoding the enzyme have not been identified to date.
[0415] Cargill has developed a novel 2,3-aminomutase enzyme to convert L-alanine to .beta.-alanine, thus creating a pathway from pyruvate to 3-HP in four biochemical steps (Liao et al., U.S. Publication No. 2005-0221466).
6.2.1.a--Acid-Thiol Ligase
[0416] An exemplary acid-thiol ligase is the gene products of sucCD of E. coli which together catalyze the formation of succinyl-CoA from succinate with the concaminant consumption of one ATP, a reaction which is reversible in vivo (Buck et al. Biochemistry 24(22): p. 6245-6252 (1985)). Additional exemplary CoA-ligases include the rat dicarboxylate-CoA ligase for which the sequence is yet uncharacterized (Vamecq et al. Biochem J. 230(3): p. 683-693 (1985)), either of the two characterized phenylacetate-CoA ligases from P. chrysogenum (Lamas-Maceiras et al. Biochem J 395(1):147-155 (2006); Wang et al. Biochem Biophys Res Commun, 360(2):453-458 (2007)), the phenylacetate-CoA ligase from Pseudomonas putida (Martinez-Blanco et al. J Biol Chem. 265(12):7084-7090 (1990)), and the 6-carboxyhexanoate-CoA ligase from Bacillus subtilis (Bower et al. J Bacteriol 178(14):4122-4130 (1996)).
TABLE-US-00064 Gene Accession No. GI No. Organism sucC NP_415256.1 16128703 Escherichia coli sucD AAC73823.1 1786949 Escherichia coli phl CAJ15517.1 77019264 Penicillium chrysogenum phlB ABS19624.1 152002983 Penicillium chrysogenum paaF AAC24333.2 22711873 Pseudomonas putida bioW NP_390902.2 50812281 Bacillus subtilis
Example V
Exemplary BDO Pathway from Succinyl-CoA
[0417] This example describes exemplary BDO pathways from succinyl-CoA.
[0418] BDO pathways from succinyl-CoA are described herein and have been described previously (see U.S. application Ser. No. 12/049,256, filed Mar. 14, 2008, and PCT application serial No. US08/57168, filed Mar. 14, 2008, each of which is incorporated herein by reference). Additional pathways are shown in FIG. 8A. Enzymes of such exemplary BDO pathways are listed in Table 15, along with exemplary genes encoding these enzymes.
[0419] Briefly, succinyl-CoA can be converted to succinic semialdehyde by succinyl-CoA reductase (or succinate semialdehyde dehydrogenase) (EC 1.2.1.b). Succinate semialdehyde can be converted to 4-hydroxybutyrate by 4-hydroxybutyrate dehydrogenase (EC 1.1.1.a), as previously described. Alternatively, succinyl-CoA can be converted to 4-hydroxybutyrate by succinyl-CoA reductase (alcohol forming) (EC 1.1.1.c). 4-Hydroxybutyrate can be converted to 4-hydroxybutyryl-CoA by 4-hydroxybutyryl-CoA transferase (EC 2.8.3.a), as previously described, or by 4-hydroxybutyryl-CoA hydrolase (EC 3.1.2.a) or 4-hydroxybutyryl-CoA ligase (or 4-hydroxybutyryl-CoA synthetase) (EC 6.2.1.a). Alternatively, 4-hydroxybutyrate can be converted to 4-hydroxybutyryl-phosphate by 4-hydroxybutyrate kinase (EC 2.7.2.a), as previously described. 4-Hydroxybutyryl-phosphate can be converted to 4-hydroxybutyryl-CoA by phosphotrans-4-hydroxybutyrylase (EC 2.3.1.a), as previously described. Alternatively, 4-hydroxybutyryl-phosphate can be converted to 4-hydroxybutanal by 4-hydroxybutanal dehydrogenase (phosphorylating) (EC 1.2.1.d). 4-Hydroxybutyryl-CoA can be converted to 4-hydroxybutanal by 4-hydroxybutyryl-CoA reductase (or 4-hydroxybutanal dehydrogenase) (EC 1.2.1.b). Alternatively, 4-hydroxybutyryl-CoA can be converted to 1,4-butanediol by 4-hydroxybutyryl-CoA reductase (alcohol forming) (EC 1.1.1.c). 4-Hydroxybutanal can be converted to 1,4-butanediol by 1,4-butanediol dehydrogenase (EC 1.1.1.a), as previously described.
TABLE-US-00065 TABLE 15 BDO pathway from succinyl-CoA. EC Desired GenBank ID FIG. class substrate Desired product Enzyme name Gene name (if available) Organism Known Substrates 8A 1.2.1.b succinyl-CoA succinic succinyl-CoA sucD P38947.1 Clostridium kluyveri succinyl-CoA semialdehyde reductase (or succinate semialdehyde dehydrogenase) sucD NP_904963.1 Porphyromonas succinyl-CoA gingivalis Msed_0709 YP_001190808.1 Metallosphaera sedula malonyl-CoA 8A 1.1.1.a succinate 4- 4-hydroxy- 4hbd YP_726053.1 Ralstonia eutropha 4-hydroxybutyrate semialdehyde hydroxybutyrate butyrate H16 dehydrogenase 4hbd L21902.1 Clostridium kluyveri 4-hydroxybutyrate DSM 555 4hbd Q94B07 Arabidopsis thaliana 4-hydroxybutyrate 8A 1.1.1.c succinyl-CoA 4- succinyl-CoA adhE2 AAK09379.1 Clostridium butanoyl-CoA hydroxybutyrate reductase acetobutylicum (alcohol forming) mcr AAS20429.1 Chloroflexus malonyl-CoA aurantiacus FAR AAD38039.1 Simmondsia chinensis long chain acyl- CoA 8A 2.8.3.a 4- 4-hydroxybutyryl- 4-hydroxy- cat1, cat2, P38946.1, Clostridium kluyveri succinate, 4- hydroxybutyrate CoA butyryl-CoA cat3 P38942.2, hydroxybutyrate, transferase EDK35586.1 butyrate gctA, gctB CAA57199.1, Acidaminococcus glutarate CAA57200.1 fermentans atoA, atoD P76459.1, Escherichia coli butanoate P76458.1 8A 3.1.2.a 4- 4- 4-hydroxy- tesB NP_414986 Escherichia coli adipyl-CoA hydroxybutyrate hydroxybutyryl- butyryl-CoA CoA hydrolase acot12 NP_570103.1 Rattus norvegicus butyryl-CoA hibch Q6NVY1.2 Homo sapiens 3- hydroxypropanoyl- CoA 8A 6.2.1.a 4- 4- 4-hydroxy- sucCD NP_415256.1, Escherichia coli succinate hydroxybutyrate hydroxybutyryl- butyryl-CoA AAC73823.1 CoA ligase (or 4- hydroxy- butyryl-CoA synthetase) phl CAJ15517.1 Penicillium phenylacetate chrysogenum bioW NP_390902.2 Bacillus subtilis 6-carboxyhexanoate 8A 2.7.2.a 4- 4- 4-hydroxy- ackA NP_416799.1 Escherichia coli acetate, propionate hydroxybutyrate hydroxybutyryl- butyrate phosphate kinase buk1 NP_349675 Clostridium butyrate acetobutylicum buk2 Q97II1 Clostridium butyrate acetobutylicum 8A 2.3.1.a 4- 4- phospho- ptb NP_349676 Clostridium butyryl-phosphate hydroxybutyryl- hydroxybutyryl- trans-4- acetobutylicum phosphate CoA hydroxy- butyrylase ptb AAR19757.1 butyrate-producing butyryl-phosphate bacterium L2-50 ptb CAC07932.1 Bacillus megaterium butyryl-phosphate 8A 1.2.1.d 4- 4-hydroxybutanal 4-hydroxy- asd NP_417891.1 Escherichia coli L-4-aspartyl- hydroxybutyryl- butanal phosphate phosphate dehydrogenase (phosphoryl- ating) proA NP_414778.1 Escherichia coli L-glutamyl-5- phospate gapA P0A9B2.2 Escherichia coli Glyceraldehyde-3- phosphate 8A 1.2.1.b 4- 4-hydroxybutanal 4-hydroxy- sucD P38947.1 Clostridium kluyveri succinyl-CoA hydroxybutyryl- butyryl-CoA CoA reductase (or 4- hydroxybutanal dehydrogenase) sucD NP_904963.1 Porphyromonas succinyl-CoA gingivalis Msed_0709 YP_001190808.1 Metallosphaera sedula malonyl-CoA 8A 1.1.1.c 4- 1,4-butanediol 4-hydroxy- adhE2 AAK09379.1 Clostridium butanoyl-CoA hydroxybutyryl- butyryl-CoA acetobutylicum CoA reductase (alcohol forming) mcr AAS20429.1 Chloroflexus malonyl-CoA aurantiacus FAR AAD38039.1 Simmondsia chinensis long chain acyl- CoA 8A 1.1.1.a 4- 1,4-butanediol 1,4-butanediol ADH2 NP_014032.1 Saccharymyces general hydroxybutanal dehydrogenase cerevisiae yqhD NP_417484.1 Escherichia coli >C3 4hbd L21902.1 Clostridium kluyveri Succinate DSM 555 semialdehyde
Example VI
Additional Exemplary BDO Pathways from Alpha-Ketoglutarate
[0420] This example describes exemplary BDO pathways from alpha-ketoglutarate.
[0421] BDO pathways from succinyl-CoA are described herein and have been described previously (see U.S. application Ser. No. 12/049,256, filed Mar. 14, 2008, and PCT application serial No. US08/57168, filed Mar. 14, 2008, each of which is incorporated herein by reference). Additional pathways are shown in FIG. 8B. Enzymes of such exemplary BDO pathways are listed in Table 16, along with exemplary genes encoding these enzymes.
[0422] Briefly, alpha-ketoglutarate can be converted to succinic semialdehyde by alpha-ketoglutarate decarboxylase (EC 4.1.1.a), as previously described. Alternatively, alpha-ketoglutarate can be converted to glutamate by glutamate dehydrogenase (EC 1.4.1.a). 4-Aminobutyrate can be converted to succinic semialdehyde by 4-aminobutyrate oxidoreductase (deaminating) (EC 1.4.1.a) or 4-aminobutyrate transaminase (EC 2.6.1.a). Glutamate can be converted to 4-aminobutyrate by glutamate decarboxylase (EC 4.1.1.a). Succinate semialdehyde can be converted to 4-hydroxybutyrate by 4-hydroxybutyrate dehydrogenase (EC 1.1.1.a), as previously described. 4-Hydroxybutyrate can be converted to 4-hydroxybutyryl-CoA by 4-hydroxybutyryl-CoA transferase (EC 2.8.3.a), as previously described, or by 4-hydroxybutyryl-CoA hydrolase (EC 3.1.2.a), or 4-hydroxybutyryl-CoA ligase (or 4-hydroxybutyryl-CoA synthetase) (EC 6.2.1.a). 4-Hydroxybutyrate can be converted to 4-hydroxybutyryl-phosphate by 4-hydroxybutyrate kinase (EC 2.7.2.a). 4-Hydroxybutyryl-phosphate can be converted to 4-hydroxybutyryl-CoA by phosphotrans-4-hydroxybutyrylase (EC 2.3.1.a), as previously described. Alternatively, 4-hydroxybutyryl-phosphate can be converted to 4-hydroxybutanal by 4-hydroxybutanal dehydrogenase (phosphorylating) (EC 1.2.1.d). 4-Hydroxybutyryl-CoA can be converted to 4-hydroxybutanal by 4-hydroxybutyryl-CoA reductase (or 4-hydroxybutanal dehydrogenase) (EC 1.2.1.b), as previously described. 4-Hydroxybutyryl-CoA can be converted to 1,4-butanediol by 4-hydroxybutyryl-CoA reductase (alcohol forming) (EC 1.1.1.c). 4-Hydroxybutanal can be converted to 1,4-butanediol by 1,4-butanediol dehydrogenase (EC 1.1.1.a), as previously described.
TABLE-US-00066 TABLE 16 BDO pathway from alpha-ketoglutarate. Desired Desired GenBank ID Known FIG. EC class substrate product Enzyme name Gene name (if available) Organism Substrates 8B 4.1.1.a alpha- succinic alpha-ketoglutarate kgd O50463.4 Mycobacterium alpha- ketoglutarate semialdehyde decarboxylase tuberculosis ketoglutarate gadA NP_417974 Escherichia coli glutamate gadB NP_416010 Escherichia coli glutamate 8B 1.4.1.a alpha- glutamate glutamate gdhA P00370 Escherichia coli glutamate ketoglutarate dehydrogenase gdh P96110.4 Thermotoga glutamate maritima gdhA1 NP_279651.1 Halobacterium glutamate salinarum 8B 1.4.1.a 4-aminobutyrate succinic 4-aminobutyrate lysDH AB052732 Geobacillus lysine semialdehyde oxidoreductase stearothermo- (deaminating) philus lysDH NP_147035.1 Aeropyrum lysine pernix K1 ldh P0A393 Bacillus cereus leucine, isoleucine, valine, 2- aminobutanoate 8B 2.6.1.a 4-aminobutyrate succinic 4-aminobutyrate gabT P22256.1 Escherichia coli 4- semialdehyde transaminase aminobutyryate puuE NP_415818.1 Escherichia coli 4- aminobutyryate UGA1 NP_011533.1 Saccharomyces 4- cerevisiae aminobutyryate 8B 4.1.1.a glutamate 4-aminobutyrate glutamate gadA NP_417974 Escherichia coli glutamate decarboxylase gadB NP_416010 Escherichia coli glutamate kgd O50463.4 Mycobacterium alpha- tuberculosis ketoglutarate 8B 1.1.1.a succinate 4- 4-hydroxybutyrate 4hbd YP_726053.1 Ralstonia 4- semialdehyde hydroxybutyrate dehydrogenase eutropha H16 hydroxybutyrate 4hbd L21902.1 Clostridium 4- kluyveri hydroxybutyrate DSM 555 4hbd Q94B07 Arabidopsis 4- thaliana hydroxybutyrate 8B 2.8.3.a 4- 4- 4-hydroxybutyryl- cat1, cat2, P38946.1, Clostridium succinate, 4- hydroxybutyrate hydroxybutyryl- CoA transferase cat3 P38942.2, kluyveri hydroxybutyrate, CoA EDK35586.1 butyrate gctA, gctB CAA57199.1, Acidaminococcus glutarate CAA57200.1 fermentans atoA, atoD P76459.1, Escherichia coli butanoate P76458.1 8B 3.1.2.a 4- 4- 4-hydroxybutyryl- tesB NP_414986 Escherichia coli adipyl-CoA hydroxybutyrate hydroxybutyryl- CoA hydrolase CoA acot12 NP_570103.1 Rattus norvegicus butyryl-CoA hibch Q6NVY1.2 Homo sapiens 3- hydroxypro- panoyl-CoA 8B 6.2.1.a 4- 4- 4-hydroxybutyryl- sucCD NP_415256.1, Escherichia coli succinate hydroxybutyrate hydroxybutyryl- CoA ligase (or 4- AAC73823.1 CoA hydroxybutyryl- CoA synthetase) phl CAJ15517.1 Penicillium phenylacetate chrysogenum bioW NP_390902.2 Bacillus subtilis 6- carboxyhexanoate 8B 2.7.2.a 4- 4- 4-hydroxybutyrate ackA NP_416799.1 Escherichia coli acetate, hydroxybutyrate hydroxybutyryl- kinase propionate phosphate buk1 NP_349675 Clostridium butyrate acetobutylicum buk2 Q97II1 Clostridium butyrate acetobutylicum 8B 2.3.1.a 4- 4- phosphotrans-4- ptb NP_349676 Clostridium butyryl- hydroxybutyryl- hydroxybutyryl- hydroxybutyrylase acetobutylicum phosphate phosphate CoA ptb AAR19757.1 butyrate- butyryl- producing phosphate bacterium L2-50 ptb CAC07932.1 Bacillus butyryl- megaterium phosphate 8B 1.2.1.d 4- 4- 4-hydroxybutanal asd NP_417891.1 Escherichia coli L-4-aspartyl- hydroxybutyryl- hydroxybutanal dehydrogenase phosphate phosphate (phosphorylating) proA NP_414778.1 Escherichia coli L-glutamyl-5- phospate gapA P0A9B2.2 Escherichia coli Glyceraldehyde- 3-phosphate 8B 1.2.1.b 4- 4- 4-hydroxybutyryl- sucD P38947.1 Clostridium succinyl-CoA hydroxybutyryl- hydroxybutanal CoA reductase (or kluyveri CoA 4-hydroxybutanal dehydrogenase) sucD NP_904963.1 Porphyromonas succinyl-CoA gingivalis Msed_0709 YP_001190808.1 Metallosphaera malonyl-CoA sedula 8B 1.1.1.c 4- 1,4-butanediol 4-hydroxybutyryl- adhE2 AAK09379.1 Clostridium butanoyl-CoA hydroxybutyryl- CoA reductase acetobutylicum CoA (alcohol forming) mcr AAS20429.1 Chloroflexus malonyl-CoA aurantiacus FAR AAD38039.1 Simmondsia long chain acyl- chinensis CoA 8B 1.1.1.a 4- 1,4-butanediol 1,4-butanediol ADH2 NP_014032.1 Saccharymyces general hydroxybutanal dehydrogenase cerevisiae yqhD NP_417484.1 Escherichia coli >C3 4hbd L21902.1 Clostridium Succinate kluyveri semialdehyde DSM 555
Example VII
BDO Pathways from 4-Aminobutyrate
[0423] This example describes exemplary BDO pathways from 4-aminobutyrate.
[0424] FIG. 9A depicts exemplary BDO pathways in which 4-aminobutyrate is converted to BDO. Enzymes of such an exemplary BDO pathway are listed in Table 17, along with exemplary genes encoding these enzymes.
[0425] Briefly, 4-aminobutyrate can be converted to 4-aminobutyryl-CoA by 4-aminobutyrate CoA transferase (EC 2.8.3.a), 4-aminobutyryl-CoA hydrolase (EC 3.1.2.a), or 4-aminobutyrate-CoA ligase (or 4-aminobutyryl-CoA synthetase) (EC 6.2.1.a). 4-aminobutyryl-CoA can be converted to 4-oxobutyryl-CoA by 4-aminobutyryl-CoA oxidoreductase (deaminating) (EC 1.4.1.a) or 4-aminobutyryl-CoA transaminase (EC 2.6.1.a). 4-oxobutyryl-CoA can be converted to 4-hydroxybutyryl-CoA by 4-hydroxybutyryl-CoA dehydrogenase (EC 1.1.1.a). 4-hydroxybutyryl-CoA can be converted to 1,4-butanediol by 4-hydroxybutyryl-CoA reductase (alcohol forming) (EC 1.1.1.c). Alternatively, 4-hydroxybutyryl-CoA can be converted to 4-hydroxybutanal by 4-hydroxybutyryl-CoA reductase (or 4-hydroxybutanal dehydrogenase) (EC 1.2.1.b). 4-hydroxybutanal can be converted to 1,4-butanediol by 1,4-butanediol dehydrogenase (EC 1.1.1.a).
TABLE-US-00067 TABLE 17 BDO pathway from 4-aminobutyrate. Desired Desired GenBank ID Known FIG. EC class substrate product Enzyme name Gene name (if available) Organism Substrates 9A 2.8.3.a 4- 4- 4-aminobutyrate cat1, cat2, P38946.1, Clostridium succinate, 4- aminobutyrate aminobutyryl- CoA transferase cat3 P38942.2, kluyveri hydroxybutyrate, CoA EDK35586.1 butyrate gctA, gctB CAA57199.1, Acidaminococcus glutarate CAA57200.1 fermentans atoA, atoD P76459.1, Escherichia coli butanoate P76458.1 9A 3.1.2.a 4- 4- 4-aminobutyryl- tesB NP_414986 Escherichia coli adipyl-CoA aminobutyrate aminobutyryl- CoA hydrolase CoA acot12 NP_570103.1 Rattus norvegicus butyryl-CoA hibch Q6NVY1.2 Homo sapiens 3- hydroxypropanoyl- CoA 9A 6.2.1.a 4- 4- 4-aminobutyrate- sucCD NP_415256.1, Escherichia coli succinate aminobutyrate aminobutyryl- CoA ligase (or 4- AAC73823.1 CoA aminobutyryl- CoA synthetase) phl CAJ15517.1 Penicillium phenylacetate chrysogenum bioW NP_390902.2 Bacillus subtilis 6- carboxyhexanoate 9A 1.4.1.a 4- 4-oxobutyryl- 4-aminobutyryl- lysDH AB052732 Geobacillus lysine aminobutyryl- CoA CoA stearothermophilus CoA oxidoreductase (deaminating) lysDH NP_147035.1 Aeropyrum lysine pernix K1 ldh P0A393 Bacillus cereus leucine, isoleucine, valine, 2- aminobutanoate 9A 2.6.1.a 4- 4-oxobutyryl- 4-aminobutyryl- gabT P22256.1 Escherichia coli 4-aminobutyryate aminobutyryl- CoA CoA transaminase CoA abat P50554.3 Rattus norvegicus 3-amino-2- methylpropionate SkyPYD4 ABF58893.1 Saccharomyces beta-alanine 9A 1.1.1.a 4-oxobutyryl- 4- 4-hydroxybutyryl- ADH2 NP_014032.1 kluyveri general CoA hydroxy- CoA Saccharymyces butyryl- dehydrogenase cerevisiae CoA yqhD NP_417484.1 Escherichia coli >C3 4hbd L21902.1 Clostridium Succinate kluyveri semialdehyde DSM 555 8 1.1.1.c 4- 1,4-butanediol 4-hydroxybutyryl- adhE2 AAK09379.1 Clostridium butanoyl-CoA hydroxybutyryl- CoA reductase acetobutylicum CoA (alcohol forming) mcr AAS20429.1 Chloroflexus malonyl-CoA aurantiacus FAR AAD38039.1 Simmondsia long chain acyl- chinensis CoA 8 1.2.1.b 4- 4- 4-hydroxybutyryl- sucD P38947.1 Clostridium Succinyl-CoA hydroxybutyryl- hydroxybutanal CoA reductase (or kluyveri CoA 4-hydroxybutanal dehydrogenase) sucD NP_904963.1 Porphyromonas Succinyl-CoA gingivalis Msed_0709 YP_001190808.1 Metallosphaera Malonyl-CoA sedula 8 1.1.1.a 4- 1,4-butanediol 1,4-butanediol ADH2 NP_014032.1 Saccharymyces general hydroxybutanal dehydrogenase cerevisiae yqhD NP_417484.1 Escherichia coli >C3 4hbd L21902.1 Clostridium Succinate kluyveri semialdehyde DSM 555
[0426] Enzymes for another exemplary BDO pathway converting 4-aminobutyrate to BDO is shown in FIG. 9A. Enzymes of such an exemplary BDO pathway are listed in Table 18, along with exemplary genes encoding these enzymes.
[0427] Briefly, 4-aminobutyrate can be converted to 4-aminobutyryl-CoA by 4-aminobutyrate CoA transferase (EC 2.8.3.a), 4-aminobutyryl-CoA hydrolase (EC 3.1.2.a) or 4-aminobutyrate-CoA ligase (or 4-aminobutyryl-CoA synthetase) (EC 6.2.1.a). 4-aminobutyryl-CoA can be converted to 4-aminobutan-1-ol by 4-aminobutyryl-CoA reductase (alcohol forming) (EC 1.1.1.c). Alternatively, 4-aminobutyryl-CoA can be converted to 4-aminobutanal by 4-aminobutyryl-CoA reductase (or 4-aminobutanal dehydrogenase) (EC 1.2.1.b), and 4-aminobutanal converted to 4-aminobutan-1-ol by 4-aminobutan-1-ol dehydrogenase (EC 1.1.1.a). 4-aminobutan-1-ol can be converted to 4-hydroxybutanal by 4-aminobutan-1-ol oxidoreductase (deaminating) (EC 1.4.1.a) or 4-aminobutan-1-ol transaminase (EC 2.6.1.a). 4-hydroxybutanal can be converted to 1,4-butanediol by 1,4-butanediol dehydrogenase (EC 1.1.1.a).
TABLE-US-00068 TABLE 18 BDO pathway from 4-aminobutyrate. Desired Desired GenBank ID (if Known FIG. EC class substrate product Enzyme name Gene name available) Organism Substrate 9A 2.8.3.a 4- 4- 4-aminobutyrate CoA cat1, cat2, P38946.1, Clostridium kluyveri succinate, 4- aminobutyrate aminobutyryl- transferase cat3 P38942.2, hydroxybutyrate, CoA EDK35586.1 butyrate gctA, gctB CAA57199.1, Acidaminococcus glutarate CAA57200.1 fermentans atoA, atoD P76459.1, Escherichia coli butanoate P76458.1 9A 3.1.2.a 4- 4- 4-aminobutyryl-CoA tesB NP_414986 Escherichia coli adipyl-CoA aminobutyrate aminobutyryl- hydrolase CoA acot12 NP_570103.1 Rattus norvegicus butyryl-CoA hibch Q6NVY1.2 Homo sapiens 3-hydroxy- propanoyl- CoA 9A 6.2.1.a 4- 4- 4-aminobutyrate-CoA sucCD NP_415256.1, Escherichia coli succinate aminobutyrate aminobutyryl- ligase (or 4- AAC73823.1 CoA aminobutyryl-CoA synthetase) phl CAJ15517.1 Penicillium phenylacetate chrysogenum bioW NP_390902.2 Bacillus subtilis 6- carboxyhexanoate 9A 1.1.1.c 4- 4-aminobutan- 4-aminobutyryl-CoA adhE2 AAK09379.1 Clostridium butanoyl-CoA aminobutyryl- 1-ol reductase (alcohol acetobutylicum CoA forming) mcr AAS20429.1 Chloroflexus malonyl-CoA aurantiacus FAR AAD38039.1 Simmondsia long chain chinensis acyl-CoA 9A 1.2.1.b 4- 4-aminobutanal 4-aminobutyryl-CoA sucD P38947.1 Clostridium kluyveri Succinyl-CoA aminobutyryl- reductase (or 4- CoA aminobutanal dehydrogenase) sucD NP_904963.1 Porphyromonas Succinyl-CoA gingivalis Msed_0709 YP_001190808.1 Metallosphaera Malonyl-CoA sedula 9A 1.1.1.a 4-aminobutanal 4-aminobutan- 4-aminobutan-1-ol ADH2 NP_014032.1 Saccharymyces general 1-ol dehydrogenase cerevisiae yqhD NP_417484.1 Escherichia coli >C3 4hbd L21902.1 Clostridium kluyveri Succinate DSM 555 semialdehyde 9A 1.4.1.a 4-aminobutan- 4- 4-aminobutan-1-ol lysDH AB052732 Geobacillus lysine 1-ol hydroxybutanal oxidoreductase stearothermophilus (deaminating) lysDH NP_147035.1 Aeropyrum pernix lysine K1 ldh P0A393 Bacillus cereus leucine, isoleucine, valine, 2- aminobutanoate 9A 2.6.1.a 4-aminobutan- 4- 4-aminobutan-1-ol gabT P22256.1 Escherichia coli 4- 1-ol hydroxybutanal transaminase aminobutyryate abat P50554.3 Rattus norvegicus 3-amino-2- methylpropionate SkyPYD4 ABF58893.1 Saccharomyces beta-alanine kluyveri 9A 1.1.1.a 4- 1,4-butanediol 1,4-butanediol ADH2 NP_014032.1 Saccharymyces general hydroxybutanal dehydrogenase cerevisiae yqhD NP_417484.1 Escherichia coli >C3 4hbd L21902.1 Clostridium kluyveri Succinate DSM 555 semialdehyde
[0428] FIG. 9B depicts exemplary BDO pathway in which 4-aminobutyrate is converted to BDO. Enzymes of such an exemplary BDO pathway are listed in Table 19, along with exemplary genes encoding these enzymes.
[0429] Briefly, 4-aminobutyrate can be converted to [(4-aminobutanolyl)oxy] phosphonic acid by 4-aminobutyrate kinase (EC 2.7.2.a). [(4-aminobutanolyl)oxy] phosphonic acid can be converted to 4-aminobutanal by 4-aminobutyraldehyde dehydrogenase (phosphorylating) (EC 1.2.1.d). 4-aminobutanal can be converted to 4-aminobutan-1-ol by 4-aminobutan-1-ol dehydrogenase (EC 1.1.1.a). 4-aminobutan-1-ol can be converted to 4-hydroxybutanal by 4-aminobutan-1-ol oxidoreductase (deaminating) (EC 1.4.1.a) or 4-aminobutan-1-ol transaminase (EC 2.6.1.a). Alternatively, [(4-aminobutanolyl)oxy] phosphonic acid can be converted to [(4-oxobutanoyl)oxy] phosphonic acid by [(4-aminobutanolyl)oxy]phosphonic acid oxidoreductase (deaminating) (EC 1.4.1.a) or [(4-aminobutanolyl)oxy]phosphonic acid transaminase (EC 2.6.1.a). [(4-oxobutanoyl)oxy] phosphonic acid can be converted to 4-hydroxybutyryl-phosphate by 4-hydroxybutyryl-phosphate dehydrogenase (EC 1.1.1.a). 4-hydroxybutyryl-phosphate can be converted to 4-hydroxybutanal by 4-hydroxybutyraldehyde dehydrogenase (phosphorylating) (EC 1.2.1.d). 4-hydroxybutanal can be converted to 1,4-butanediol by 1,4-butanediol dehydrogenase (EC 1.1.1.a).
TABLE-US-00069 TABLE 19 BDO pathway from 4-aminobutyrate. Desired Desired Gene GenBank ID Known FIG. EC class substrate product Enzyme name name (if available) Organism Substrate 9B 2.7.2.a 4- [(4- 4- ackA NP_416799.1 Escherichia coli acetate, aminobutyrate aminobutanolyl) aminobutyrate propionate oxy] kinase phosphonic acid buk1 NP_349675 Clostridium butyrate acetobutylicum proB NP_414777.1 Escherichia coli glutamate 9B 1.2.1.d [(4- 4- 4- asd NP_417891.1 Escherichia coli L-4-aspartyl- aminobutanolyl) aminobutanal aminobutyraldehyde phosphate oxy] dehydrogenase phosphonic (phosphorylating) acid proA NP_414778.1 Escherichia coli L-glutamyl-5- phospate gapA P0A9B2.2 Escherichia coli Glyceraldehyde-3- phosphate 9B 1.1.1.a 4-aminobutanal 4-aminobutan- 4-aminobutan- ADH2 NP_014032.1 Saccharymyces general 1-ol 1-ol cerevisiae dehydrogenase yqhD NP_417484.1 Escherichia coli >C3 4hbd L21902.1 Clostridium kluyveri Succinate DSM 555 semialdehyde 9B 1.4.1.a 4-aminobutan- 4- 4-aminobutan- lysDH AB052732 Geobacillus lysine 1-ol hydroxybutanal 1-ol stearothermophilus oxidoreductase (deaminating) lysDH NP_147035.1 Aeropyrum pernix lysine K1 ldh P0A393 Bacillus cereus leucine, isoleucine, valine, 2-aminobutanoate 9B 2.6.1.a 4-aminobutan- 4- 4-aminobutan- gabT P22256.1 Escherichia coli 4-aminobutyryate 1-ol hydroxybutanal 1-ol transaminase abat P50554.3 Rattus norvegicus 3-amino-2- methylpropionate SkyPYD4 ABF58893.1 Saccharomyces beta-alanine kluyveri 9B 1.4.1.a [(4- [(4- [(4- lysDH AB052732 Geobacillus lysine aminobutanolyl) oxobutanolyl) aminobutanolyl) stearothermophilus oxy] oxy] oxy]phosphonic phosphonic phosphonic acid acid acid oxidoreductase (deaminating) lysDH NP_147035.1 Aeropyrum pernix lysine K1 ldh P0A393 Bacillus cereus leucine, isoleucine, valine, 2-aminobutanoate 9B 2.6.1.a [(4- [(4- [(4- gabT P22256.1 Escherichia coli 4-aminobutyryate aminobutanolyl) oxobutanolyl) aminobutanolyl) oxy] oxy] oxy]phosphonic phosphonic phosphonic acid acid acid transaminase SkyPYD4 ABF58893.1 Saccharomyces beta-alanine kluyveri serC NP_415427.1 Escherichia coli phosphoserine, phospho- hydroxythreonine 9B 1.1.1.a [(4- 4- 4- ADH2 NP_014032.1 Saccharymyces general oxobutanolyl) hydroxybutyryl- hydroxybutyryl- cerevisiae oxy] phosphate phosphate phosphonic dehydrogenase acid yqhD NP_417484.1 Escherichia coli >C3 4hbd L21902.1 Clostridium kluyveri Succinate DSM 555 semialdehyde 9B 1.2.1.d 4- 4- 4- asd NP_417891.1 Escherichia coli L-4-aspartyl- hydroxybutyryl- hydroxybutanal hydroxy- phosphate phosphate butyraldehyde dehydrogenase (phosphorylating) proA NP_414778.1 Escherichia coli L-glutamyl-5- phospate gapA P0A9B2.2 Escherichia coli Glyceraldehyde- 3-phosphate 9B 1.1.1.a 4- 1,4-butanediol 1,4-butanediol ADH2 NP_014032.1 Saccharymyces general hydroxybutanal dehydrogenase cerevisiae yqhD NP_417484.1 Escherichia coli >C3 4hbd L21902.1 Clostridium kluyveri Succinate DSM 555 semialdehyde
[0430] FIG. 9C shows an exemplary pathway through acetoacetate.
Example VIII
Exemplary BDO Pathways from Alpha-Ketoglutarate
[0431] This example describes exemplary BDO pathways from alpha-ketoglutarate.
[0432] FIG. 10 depicts exemplary BDO pathways in which alpha-ketoglutarate is converted to BDO. Enzymes of such an exemplary BDO pathway are listed in Table 20, along with exemplary genes encoding these enzymes.
[0433] Briefly, alpha-ketoglutarate can be converted to alpha-ketoglutaryl-phosphate by alpha-ketoglutarate 5-kinase (EC 2.7.2.a). Alpha-ketoglutaryl-phosphate can be converted to 2,5-dioxopentanoic acid by 2,5-dioxopentanoic semialdehyde dehydrogenase (phosphorylating) (EC 1.2.1.d). 2,5-dioxopentanoic acid can be converted to 5-hydroxy-2-oxopentanoic acid by 2,5-dioxopentanoic acid reductase (EC 1.1.1.a). Alternatively, alpha-ketoglutarate can be converted to alpha-ketoglutaryl-CoA by alpha-ketoglutarate CoA transferase (EC 2.8.3.a), alpha-ketoglutaryl-CoA hydrolase (EC 3.1.2.a) or alpha-ketoglutaryl-CoA ligase (or alpha-ketoglutaryl-CoA synthetase) (EC 6.2.1.a). Alpha-ketoglutaryl-CoA can be converted to 2,5-dioxopentanoic acid by alpha-ketoglutaryl-CoA reductase (or 2,5-dioxopentanoic acid dehydrogenase) (EC 1.2.1.b). 2,5-Dioxopentanoic acid can be converted to 5-hydroxy-2-oxopentanoic acid by 5-hydroxy-2-oxopentanoic acid dehydrogenase. Alternatively, alpha-ketoglutaryl-CoA can be converted to 5-hydroxy-2-oxopentanoic acid by alpha-ketoglutaryl-CoA reductase (alcohol forming) (EC 1.1.1.c). 5-hydroxy-2-oxopentanoic acid can be converted to 4-hydroxybutanal by 5-hydroxy-2-oxopentanoic acid decarboxylase (EC 4.1.1.a). 4-hydroxybutanal can be converted to 1,4-butanediol by 1,4-butanediol dehydrogenase (EC 1.1.1.a). 5-hydroxy-2-oxopentanoic acid can be converted to 4-hydroxybutyryl-CoA by 5-hydroxy-2-oxopentanoic acid dehydrogenase (decarboxylation) (EC 1.2.1.c).
TABLE-US-00070 TABLE 20 BDO pathway from alpha-ketoglutarate. Desired Desired Gene GenBank ID FIG. EC class substrate product Enzyme name name (if available) Organism Known Substrate 10 2.7.2.a alpha- alpha- alpha- ackA NP_416799.1 Escherichia coli acetate, propionate ketoglutarate ketoglutaryl- ketoglutarate 5- phosphate kinase buk1 NP_349675 Clostridium butyrate acetobutylicum proB NP_414777.1 Escherichia coli glutamate 10 1.2.1.d alpha- 2,5- 2,5- proA NP_414778.1 Escherichia coli L-glutamyl- ketoglutaryl- dioxopentanoic dioxopentanoic 5-phospate phosphate acid semialdehyde dehydrogenase (phosphorylating) asd NP_417891.1 Escherichia coli L-4-aspartyl- phosphate gapA P0A9B2.2 Escherichia coli Glyceraldehyde- 3-phosphate 10 1.1.1.a 2,5- 5-hydroxy-2- 2,5- ADH2 NP_014032.1 Saccharymyces general dioxopentanoic oxopentanoic dioxopentanoic cerevisiae acid acid acid reductase yqhD NP_417484.1 Escherichia coli >C3 4hbd L21902.1 Clostridium Succinate kluyveri semialdehyde DSM 555 10 2.8.3.a alpha- alpha- alpha- cat1, cat2, P38946.1, Clostridium succinate, ketoglutarate ketoglutaryl- ketoglutarate cat3 P38942.2, kluyveri 4-hydroxybutyrate, CoA CoA EDK35586.1 butyrate transferase gctA, gctB CAA57199.1, Acidaminococcus glutarate CAA57200.1 fermentans atoA, atoD P76459.1, Escherichia coli butanoate P76458.1 10 3.1.2.a alpha- alpha- alpha- tesB NP_414986 Escherichia coli adipyl-CoA ketoglutarate ketoglutaryl- ketoglutaryl- CoA CoA hydrolase acot12 NP_570103.1 Rattus norvegicus butyryl-CoA hibch Q6NVY1.2 Homo sapiens 3-hydroxy- propanoyl-CoA 10 6.2.1.a alpha- alpha- alpha- sucCD NP_415256.1, Escherichia coli succinate ketoglutarate ketoglutaryl- ketoglutaryl- AAC73823.1 CoA CoA ligase (or alpha- ketoglutaryl- CoA synthetase) phl CAJ15517.1 Penicillium phenylacetate chrysogenum bioW NP_390902.2 Bacillus 6-carboxy- subtilis hexanoate 10 1.2.1.b alpha- 2,5- alpha- sucD P38947.1 Clostridium Succinyl-CoA ketoglutaryl- dioxopentanoic ketoglutaryl- kluyveri CoA acid CoA reductase (or 2,5- dioxopentanoic acid dehydrogenase) Msed_0709 YP_001190808.1 Metallosphaera Malonyl-CoA sedula bphG BAA03892.1 Pseudomonas sp Acetaldehyde, Propionaldehyde, Butyraldehyde, Isobutyraldehyde and Formaldehyde 10 1.1.1.a 2,5- 5-hydroxy-2- 5-hydroxy-2- ADH2 NP_014032.1 Saccharymyces general dioxopentanoic oxopentanoic oxopentanoic yqhD NP_417484.1 cerevisiae >C3 acid acid acid 4hbd L21902.1 Escherichia coli Succinate dehydrogenase Clostridium semialdehyde kluyveri DSM 555 10 1.1.1.c alpha- 5-hydroxy-2- alpha- adhE2 AAK09379.1 Clostridium butanoyl- ketoglutaryl- oxopentanoic ketoglutaryl- acetobutylicum CoA CoA acid CoA reductase (alcohol forming) mcr AAS20429.1 Chloroflexus malonyl-CoA aurantiacus FAR AAD38039.1 Simmondsia long chain chinensis acyl-CoA 10 4.1.1.a 5-hydroxy-2- 4- 5-hydroxy-2- pdc P06672.1 Zymomonas mobilus 2-oxopentanoic oxopentanoic hydroxybutanal oxopentanoic acid acid acid decarboxylase mdlC P20906.2 Pseudomonas 2-oxopentanoic putida acid pdc1 P06169 Saccharomyces pyruvate cerevisiae 10 1.1.1.a 4- 1,4-butanediol 1,4-butanediol ADH2 NP_014032.1 Saccharomyces general hydroxybutanal dehydrogenase cerevisiae yqhD NP_417484.1 Escherichia coli >C3 4hbd L21902.1 Clostridium Succinate kluyveri semialdehyde DSM 555 10 1.2.1.c 5-hydroxy-2- 4- 5-hydroxy-2- sucA, sucB, NP_415254.1, Escherichia coli Alpha- oxopentanoic hydroxybutyryl- oxopentanoic lpd NP_415255.1, ketoglutarate acid CoA acid NP_414658.1 dehydrogenase (decarboxylation) bfmBB, NP_390283.1, Bacillus subtilis 2-keto acids bfmBAA, NP_390285.1, derivatives of bfmBAB, NP_390284.1, valine, leucine bfmBAB, P21880.1 and isoleucine pdhD Bckdha, NP_036914.1, Rattus norvegicus 2-keto acids Bckdhb, NP_062140.1, derivatives of Dbt, Dld NP_445764.1, valine, leucine NP_955417.1 and isoleucine
Example IX
Exemplary BDO Pathways from Glutamate
[0434] This example describes exemplary BDO pathways from glutamate.
[0435] FIG. 11 depicts exemplary BDO pathways in which glutamate is converted to BDO. Enzymes of such an exemplary BDO pathway are listed in Table 21, along with exemplary genes encoding these enzymes.
[0436] Briefly, glutamate can be converted to glutamyl-CoA by glutamate CoA transferase (EC 2.8.3.a), glutamyl-CoA hydrolase (EC 3.1.2.a) or glutamyl-CoA ligase (or glutamyl-CoA synthetase) (EC 6.2.1.a). Alternatively, glutamate can be converted to glutamate-5-phosphate by glutamate 5-kinase (EC 2.7.2.a). Glutamate-5-phosphate can be converted to glutamate-5-semialdehyde by glutamate-5-semialdehyde dehydrogenase (phosphorylating) (EC 1.2.1.d). Glutamyl-CoA can be converted to glutamate-5-semialdehyde by glutamyl-CoA reductase (or glutamate-5-semialdehyde dehydrogenase) (EC 1.2.1.b). Glutamate-5-semialdehyde can be converted to 2-amino-5-hydroxypentanoic acid by glutamate-5-semialdehyde reductase (EC 1.1.1.a). Alternatively, glutamyl-CoA can be converted to 2-amino-5-hydroxypentanoic acid by glutamyl-CoA reductase (alcohol forming) (EC 1.1.1.c). 2-Amino-5-hydroxypentanoic acid can be converted to 5-hydroxy-2-oxopentanoic acid by 2-amino-5-hydroxypentanoic acid oxidoreductase (deaminating) (EC 1.4.1.a) or 2-amino-5-hydroxypentanoic acid transaminase (EC 2.6.1.a). 5-Hydroxy-2-oxopentanoic acid can be converted to 4-hydroxybutanal by 5-hydroxy-2-oxopentanoic acid decarboxylase (EC 4.1.1.a). 4-Hydroxybutanal can be converted to 1,4-butanediol by 1,4-butanediol dehydrogenase (EC 1.1.1.a). Alternatively, 5-hydroxy-2-oxopentanoic acid can be converted to 4-hydroxybutyryl-CoA by 5-hydroxy-2-oxopentanoic acid dehydrogenase (decarboxylation) (EC 1.2.1.c).
TABLE-US-00071 TABLE 21 BDO pathway from glutamate. Desired Desired GenBank ID (if FIG. EC class substrate product Enzyme name Gene name available) Organism Known Substrate 11 2.8.3.a glutamate glutamyl-CoA glutamate CoA cat1, cat2, P38946.1, Clostridium succinate, 4- transferase cat3 P38942.2, kluyveri hydroxybutyrate, EDK35586.1 butyrate gctA, gctB CAA57199.1, Acidaminococcus glutarate CAA57200.1 fermentans atoA, atoD P76459.1, Escherichia coli butanoate P76458.1 11 3.1.2.a glutamate glutamyl-CoA glutamyl-CoA tesB NP_414986 Escherichia coli adipyl-CoA hydrolase acot12 NP_570103.1 Rattus norvegicus butyryl-CoA hibch Q6NVY1.2 Homo sapiens 3-hydrox- ypropanoyl- CoA 11 6.2.1.a glutamate glutamyl-CoA glutamyl-CoA sucCD NP_415256.1, Escherichia coli succinate ligase (or AAC73823.1 glutamyl- CoA synthetase) phl CAJ15517.1 Penicillium phenylacetate chrysogenum bioW NP_390902.2 Bacillus subtilis 6-carboxy- hexanoate 11 2.7.2.a glutamate glutamate-5- glutamate ackA NP_416799.1 Escherichia coli acetate, phosphate 5-kinase propionate buk1 NP_349675 Clostridium butyrate acetobutylicum proB NP_414777.1 Escherichia coli glutamate 11 1.2.1.d glutamate-5- glutamate-5- glutamate-5- proA NP_414778.1 Escherichia coli L-glutamyl-5- phosphate semialdehyde semialdehyde phospate dehydrogenase (phosphorylating) asd NP_417891.1 Escherichia coli L-4-aspartyl- phosphate gapA P0A9B2.2 Escherichia coli Glyceraldehyde- 3-phosphate 11 1.2.1.b glutamyl-CoA glutamate-5- glutamyl-CoA sucD P38947.1 Clostridium Succinyl-CoA semialdehyde reductase (or kluyveri glutamate-5- semialdehyde dehydrogenase) Msed_0709 YP_001190808.1 Metallosphaera Malonyl-CoA sedula bphG BAA03892.1 Pseudomonas sp Acetaldehyde, Propionaldehyde, Butyraldehyde, Isobutyraldehyde and Formaldehyde 11 1.1.1.a glutamate-5- 2-amino-5- glutamate-5- ADH2 NP_014032.1 Saccharymyces general semialdehyde hydroxypentanoic semialdehyde cerevisiae acid reductase yqhD NP_417484.1 Escherichia coli >C3 4hbd L21902.1 Clostridium Succinate kluyveri semialdehyde DSM 555 11 1.1.1.c glutamyl-CoA 2-amino-5- glutamyl-CoA adhE2 AAK09379.1 Clostridium butanoyl-CoA hydroxypentanoic reductase (alcohol acetobutylicum acid forming) mcr AAS20429.1 Chloroflexus malonyl-CoA aurantiacus FAR AAD38039.1 Simmondsia long chain chinensis acyl-CoA 11 1.4.1.a 2-amino-5- 5-hydroxy-2- 2-amino-5- gdhA P00370 Escherichia coli glutamate hydroxypentanoic oxopentanoic hydroxypentanoic acid acid acid oxidoreductase (deaminating) ldh P0A393 Bacillus cereus leucine, isoleucine, valine, 2- aminobutanoate nadX NP_229443.1 Thermotoga aspartate maritima 11 2.6.1.a 2-amino-5- 5-hydroxy-2- 2-amino-5- aspC NP_415448.1 Escherichia coli aspartate hydroxypentanoic oxopentanoic hydroxypentanoic acid acid acid transaminase AAT2 P23542.3 Saccharomyces aspartate cerevisiae avtA YP_026231.1 Escherichia coli valine, alpha- aminobutyrate 11 4.1.1.a 5-hydroxy-2- 4- 5-hydroxy-2- pdc P06672.1 Zymomonas 2-oxopentanoic oxopentanoic hydroxybutanal oxopentanoic acid mobilus acid acid decarboxylase mdlC P20906.2 Pseudomonas 2-oxopentanoic putida acid pdc1 P06169 Saccharomyces pyruvate cerevisiae 11 1.1.1.a 4- 1,4-butanediol 1,4-butanediol ADH2 NP_014032.1 Saccharymyces general hydroxybutanal dehydrogenase cerevisiae yqhD NP_417484.1 Escherichia coli >C3 4hbd L21902.1 Clostridium Succinate kluyveri semialdehyde DSM 555 11 1.2.1.c 5-hydroxy-2- 4- 5-hydroxy-2- sucA, sucB, NP_415254.1, Escherichia coli Alpha- oxopentanoic hydroxybutyryl- oxopentanoic acid lpd NP_415255.1, ketoglutarate acid CoA dehydrogenase NP_414658.1 (decarboxylation) bfmBB, NP_390283.1, Bacillus subtilis 2-keto acids bfmBAA, NP_390285.1, derivatives bfmBAB, NP_390284.1, of valine, bfmBAB, P21880.1 leucine and pdhD isoleucine Bckdha, NP_036914.1, Rattus norvegicus 2-keto acids Bckdhb, NP_062140.1, derivatives Dbt, Dld NP_445764.1, of valine, NP_955417.1 leucine and isoleucine
Example X
Exemplary BDO from Acetoacetyl-CoA
[0437] This example describes an exemplary BDO pathway from acetoacetyl-CoA.
[0438] FIG. 12 depicts exemplary BDO pathways in which acetoacetyl-CoA is converted to BDO. Enzymes of such an exemplary BDO pathway are listed in Table 22, along with exemplary genes encoding these enzymes.
[0439] Briefly, acetoacetyl-CoA can be converted to 3-hydroxybutyryl-CoA by 3-hydroxybutyryl-CoA dehydrogenase (EC 1.1.1.a). 3-Hydroxybutyryl-CoA can be converted to crotonoyl-CoA by 3-hydroxybutyryl-CoA dehydratase (EC 4.2.1.a). Crotonoyl-CoA can be converted to vinylacetyl-CoA by vinylacetyl-CoA .DELTA.-isomerase (EC 5.3.3.3). Vinylacetyl-CoA can be converted to 4-hydroxybutyryl-CoA by 4-hydroxybutyryl-CoA dehydratase (EC 4.2.1.a). 4-Hydroxybutyryl-CoA can be converted to 1,4-butanediol by 4-hydroxybutyryl-CoA reductase (alcohol forming) (EC 1.1.1.c). Alternatively, 4-hydroxybutyryl-CoA can be converted to 4-hydroxybutanal by 4-hydroxybutyryl-CoA reductase (or 4-hydroxybutanal dehydrogenase) (EC 1.2.1.b). 4-Hydroxybutanal can be converted to 1,4-butanediol by 1,4-butanediol dehydrogenase (EC 1.1.1.a).
TABLE-US-00072 TABLE 22 BDO pathway from acetoacetyl-CoA. Desired Desired GenBank ID FIG. EC class substrate product Enzyme name Gene name (if available) Organism Known Substrate 12 1.1.1.a acetoacetyl- 3- 3-hydroxybutyryl- hbd NP_349314.1 Clostridium 3-hydroxybutyryl- CoA hydroxybutyryl- CoA dehydrogenase acetobutylicum CoA CoA hbd AAM14586.1 Clostridium 3-hydroxybutyryl- beijerinckii CoA Msed_1423 YP_001191505 Metallosphaera presumed 3- sedula hydroxybutyryl- CoA 12 4.2.1.a 3- crotonoyl- 3-hydroxybutyryl- crt NP_349318.1 Clostridium 3-hydroxybutyryl- hydroxybutyryl- CoA CoA dehydratase acetobutylicum CoA CoA maoC NP_415905.1 Escherichia coli 3-hydroxybutyryl- CoA paaF NP_415911.1 Escherichia coli 3-hydroxyadipyl- CoA 12 5.3.3.3 crotonoyl-CoA vinylacetyl- vinylacetyl-CoA .DELTA.- abfD YP_001396399.1 Clostridium 4-hydroxybutyryl- CoA isomerase kluyveri CoA DSM 555 abfD P55792 Clostridium 4-hydroxybutyryl- aminobutyricum CoA abfD YP_001928843 Porphyromonas 4-hydroxybutyryl- gingivalis CoA ATCC 33277 12 4.2.1.a vinylacetyl- 4- 4-hydroxybutyryl- abfD YP_001396399.1 Clostridium 4-hydroxybutyryl- CoA hydroxybutyryl- CoA dehydratase kluyveri CoA CoA DSM 555 abfD P55792 Clostridium 4-hydroxybutyryl- aminobutyricum CoA abfD YP_001928843 Porphyromonas 4-hydroxybutyryl- gingivalis ATCC 33277 CoA 12 1.1.1.c 4- 1,4- 4-hydroxybutyryl- adhE2 AAK09379.1 Clostridium butanoyl-CoA hydroxybutyryl- butanediol CoA reductase acetobutylicum CoA (alcohol forming) mcr AAS20429.1 Chloroflexus malonyl-CoA aurantiacus FAR AAD38039.1 Simmondsia long chain chinensis acyl-CoA 12 1.2.1.b 4- 4- 4-hydroxybutyryl- sucD P38947.1 Clostridium Succinyl-CoA hydroxybutyryl- hydroxybutanal CoA reductase (or kluyveri CoA 4-hydroxybutanal dehydrogenase) sucD NP_904963.1 Porphyromonas Succinyl-CoA gingivalis Msed_0709 YP_001190808.1 Metallosphaera Malonyl-CoA sedula 12 1.1.1.a 4- 1,4- 1,4-butanediol ADH2 NP_014032.1 Saccharymyces general hydroxybutanal butanediol dehydrogenase cerevisiae yqhD NP_417484.1 Escherichia coli >C3 4hbd L21902.1 Clostridium Succinate kluyveri semialdehyde DSM 555
Example XI
Exemplary BDO Pathway from Homoserine
[0440] This example describes an exemplary BDO pathway from homoserine.
[0441] FIG. 13 depicts exemplary BDO pathways in which homoserine is converted to BDO. Enzymes of such an exemplary BDO pathway are listed in Table 23, along with exemplary genes encoding these enzymes.
[0442] Briefly, homoserine can be converted to 4-hydroxybut-2-enoate by homoserine deaminase (EC 4.3.1.a). Alternatively, homoserine can be converted to homoserine-CoA by homoserine CoA transferase (EC 2.8.3.a), homoserine-CoA hydrolase (EC 3.1.2.a) or homoserine-CoA ligase (or homoserine-CoA synthetase) (EC 6.2.1.a). Homoserine-CoA can be converted to 4-hydroxybut-2-enoyl-CoA by homoserine-CoA deaminase (EC 4.3.1.a). 4-Hydroxybut-2-enoate can be converted to 4-hydroxybut-2-enoyl-CoA by 4-hydroxybut-2-enoyl-CoA transferase (EC 2.8.3.a), 4-hydroxybut-2-enoyl-CoA hydrolase (EC 3.1.2.a), or 4-hydroxybut-2-enoyl-CoA ligase (or 4-hydroxybut-2-enoyl-CoA synthetase) (EC 6.2.1.a). Alternatively, 4-hydroxybut-2-enoate can be converted to 4-hydroxybutyrate by 4-hydroxybut-2-enoate reductase (EC 1.3.1.a). 4-Hydroxybutyrate can be converted to 4-hydroxybutyryl-coA by 4-hydroxybutyryl-CoA transferase (EC 2.8.3.a), 4-hydroxybutyryl-CoA hydrolase (EC 3.1.2.a), or 4-hydroxybutyryl-CoA ligase (or 4-hydroxybutyryl-CoA synthetase) (EC 6.2.1.a). 4-Hydroxybut-2-enoyl-CoA can be converted to 4-hydroxybutyryl-CoA by 4-hydroxybut-2-enoyl-CoA reductase (EC 1.3.1.a). 4-Hydroxybutyryl-CoA can be converted to 1,4-butanediol by 4-hydroxybutyryl-CoA reductase (alcohol forming) (EC 1.1.1.c). Alternatively, 4-hydroxybutyryl-CoA can be converted to 4-hydroxybutanal by 4-hydroxybutyryl-CoA reductase (or 4-hydroxybutanal dehydrogenase) (EC 1.2.1.b). 4-Hydroxybutanal can be converted to 1,4-butanediol by 1,4-butanediol dehydrogenase (EC 1.1.1.a).
TABLE-US-00073 TABLE 23 BDO pathway from homoserine. Desired Desired GenBank ID Known FIG. EC class substrate product Enzyme name Gene name (if available) Organism Substrate 13 4.3.1.a homoserine 4-hydroxybut-2- homoserine aspA NP_418562 Escherichia coli aspartate enoate deaminase aspA P44324.1 Haemophilus aspartate influenzae aspA P07346 Pseudomonas aspartate fluorescens 13 2.8.3.a homoserine homoserine- homoserine CoA cat1, cat2, P38946.1, Clostridium succinate, 4- CoA transferase cat3 P38942.2, kluyveri hydroxybutyrate, EDK35586.1 butyrate gctA, gctB CAA57199.1, Acidaminococcus glutarate CAA57200.1 fermentans atoA, atoD P76459.1, Escherichia coli butanoate P76458.1 13 3.1.2.a homoserine homoserine- homoserine-CoA tesB NP_414986 Escherichia coli adipyl-CoA CoA hydrolase acot12 NP_570103.1 Rattus norvegicus butyryl-CoA hibch Q6NVY1.2 Homo sapiens 3- hydroxy- propanoyl- CoA 13 6.2.1.a homoserine homoserine- homoserine-CoA sucCD NP_415256.1, Escherichia coli succinate CoA ligase (or AAC73823.1 homoserine-CoA synthetase) phl CAJ15517.1 Penicillium phenylacetate chrysogenum bioW NP_390902.2 Bacillus subtilis 6- carboxyhexanoate 13 4.3.1.a homoserine- 4-hydroxybut-2- homoserine-CoA acl1 CAG29274.1 Clostridium beta-alanyl-CoA CoA enoyl-CoA deaminase propionicum acl2 CAG29275.1 Clostridium beta-alanyl-CoA propionicum MXAN_4385 YP_632558.1 Myxococcus beta-alanyl-CoA xanthus 13 2.8.3.a 4-hydroxybut- 4-hydroxybut-2- 4-hydroxybut-2- cat1, cat2, P38946.1, Clostridium succinate, 4- 2-enoate enoyl-CoA enoyl-CoA cat3 P38942.2, kluyveri hydroxybutyrate, transferase EDK35586.1 butyrate gctA, gctB CAA57199.1, Acidaminococcus glutarate CAA57200.1 fermentans atoA, atoD P76459.1, Escherichia coli butanoate P76458.1 13 3.1.2.a 4-hydroxybut- 4-hydroxybut-2- 4-hydroxybut-2- tesB NP_414986 Escherichia coli adipyl-CoA 2-enoate enoyl-CoA enoyl-CoA hydrolase acot12 NP_570103.1 Rattus norvegicus butyryl-CoA hibch Q6NVY1.2 Homo sapiens 3- hydroxy- propanoyl- CoA 13 6.2.1.a 4-hydroxybut- 4-hydroxybut-2- 4-hydroxybut-2- sucCD NP_415256.1, Escherichia coli succinate 2-enoate enoyl-CoA enoyl-CoA AAC73823.1 ligase (or 4-hydroxybut-2- enoyl-CoA synthetase) phl CAJ15517.1 Penicillium phenylacetate chrysogenum bioW NP_390902.2 Bacillus subtilis 6- carboxyhexanoate 13 1.3.1.a 4-hydroxybut- 4- 4-hydroxybut-2- enr CAA71086.1 Clostridium 2-enoate hydroxybutyrate enoate reductase tyrobutyricum enr CAA76083.1 Clostridium kluyveri enr YP_430895.1 Moorella thermoacetica 13 2.8.3.a 4- 4- 4-hydroxybutyryl- cat1, cat2, P38946.1, Clostridium succinate, 4- hydroxybutyrate hydroxybutyryl- CoA transferase cat3 P38942.2, kluyveri hydroxybutyrate, coA EDK35586.1 butyrate gctA, gctB CAA57199.1, Acidaminococcus glutarate CAA57200.1 fermentans atoA, atoD P76459.1, Escherichia coli butanoate P76458.1 13 3.1.2.a 4- 4- 4-hydroxybutyryl- tesB NP_414986 Escherichia coli adipyl-CoA hydroxybutyrate hydroxybutyryl- CoA hydrolase coA acot12 NP_570103.1 Rattus norvegicus butyryl-CoA hibch Q6NVY1.2 Homo sapiens 3- hydroxy- propanoyl- CoA 13 6.2.1.a 4- 4- 4-hydroxybutyryl- sucCD NP_415256.1, Escherichia coli succinate hydroxybutyrate hydroxybutyryl- CoA ligase (or 4- AAC73823.1 coA hydroxybutyryl- CoA synthetase) phl CAJ15517.1 Penicillium phenylacetate chrysogenum bioW NP_390902.2 Bacillus subtilis 6- carboxyhexanoate 13 1.3.1.a 4-hydroxybut-2- 4- 4-hydroxybut-2- bcd, etfA, NP_349317.1, Clostridium enoyl-CoA hydroxybutyryl- enoyl-CoA etfB NP_349315.1, acetobutylicum CoA reductase NP_349316.1 TER Q5EU90.1 Euglena gracilis TDE0597 NP_971211.1 Treponema denticola 8 1.1.1.c 4- 1,4-butanediol 4-hydroxybutyryl- adhE2 AAK09379.1 Clostridium butanoyl-CoA hydroxybutyryl- CoA reductase acetobutylicum CoA (alcohol forming) mcr AAS20429.1 Chloroflexus malonyl-CoA aurantiacus FAR AAD38039.1 Simmondsia long chain acyl- chinensis CoA 8 1.2.1.b 4- 4- 4-hydroxybutyryl- sucD P38947.1 Clostridium Succinyl-CoA hydroxybutyryl- hydroxybutanal CoA reductase kluyveri CoA (or 4- hydroxybutanal dehydrogenase) sucD NP_904963.1 Porphyromonas Succinyl-CoA gingivalis Msed_0709 YP_001190808.1 Metallosphaera Malonyl-CoA sedula 8 1.1.1.a 4-hydroxybutanal 1,4-butanediol 1,4-butanediol ADH2 NP_014032.1 Saccharymyces general dehydrogenase cerevisiae yqhD NP_417484.1 Escherichia coli >C3 4hbd L21902.1 Clostridium Succinate kluyveri semialdehyde DSM 555
Example XII
BDO Producing Strains Expressing Succinyl-CoA Synthetase
[0443] This example describes increased production of BDO in BDO producing strains expressing succinyl-CoA synthetase.
[0444] As discussed above, succinate can be a precursor for production of BDO by conversion to succinyl-CoA (see also WO2008/115840, WO 2009/023493, U.S. publication 2009/0047719, U.S. publication 2009/0075351). Therefore, the host strain was genetically modified to overexpress the E. coli sucCD genes, which encode succinyl-CoA synthetase. The nucleotide sequence of the E. coli sucCD operon is shown in FIG. 14A, and the amino acid sequences for the encoded succinyl-CoA synthetase subunits are shown in FIGS. 14B and 14C. Briefly, the E. coli sucCD genes were cloned by PCR from E. coli chromosomal DNA and introduced into multicopy plasmids pZS*13, pZA13, and pZE33 behind the PA1lacO-1 promoter (Lutz and Bujard, Nucleic Acids Res. 25:1203-1210 (1997)) using standard molecular biology procedures.
[0445] The E. coli sucCD genes, which encode the succinyl-CoA synthetase, were overexpressed. The results showed that introducing into the strains sucCD to express succinyl-CoA synthetase improved BDO production in various strains compared to either native levels of expression or expression of cat1, which is a succinyl-CoA/acetyl-CoA transferase. Thus, BDO production was improved by overexpressing the native E. coli sucCD genes encoding succinyl-CoA synthetase.
Example XIII
Expression of Heterologous Genes Encoding BDO Pathway Enzymes
[0446] This example describes the expression of various non-native pathway enzymes to provide improved production of BDO.
[0447] Alpha-Ketoglutarate Decarboxylase.
[0448] The Mycobacterium bovis sucA gene encoding alpha-ketoglutarate decarboxylase was expressed in host strains. Overexpression of M. bovis sucA improved BDO production (see also WO2008/115840, WO 2009/023493, U.S. publication 2009/0047719, U.S. publication 2009/0075351). The nucleotide and amino acid sequences of M. bovis sucA and the encoded alpha-ketoglutarate decarboxylase are shown in FIG. 15.
[0449] To construct the M. bovis sucA expressing strains, fragments of the sucA gene encoding the alpha-ketoglutarate decarboxylase were amplified from the genomic DNA of Mycobacterium bovis BCG (ATCC 19015; American Type Culture Collection, Manassas Va.) using primers shown below. The full-length gene was assembled by ligation reaction of the four amplified DNA fragments, and cloned into expression vectors pZS*13 and pZE23 behind the P.sub.A1lacO-1 promoter (Lutz and Bujard, Nucleic Acids Res. 25:1203-1210 (1997)). The nucleotide sequence of the assembled gene was verified by DNA sequencing.
TABLE-US-00074 Primers for fragment 1: (SEQ ID NO: 3) 5'-ATGTACCGCAAGTTCCGC-3' (SEQ ID NO: 4) 5'-CAATTTGCCGATGCCCAG-3' Primers for fragment 2: (SEQ ID NO: 5) 5'-GCTGACCACTGAAGACTTTG-3' (SEQ ID NO: 6) 5'-GATCAGGGCTTCGGTGTAG-3' Primers for fragment 3: (SEQ ID NO: 7) 5'-TTGGTGCGGGCCAAGCAGGATCTGCTC-3' (SEQ ID NO: 8) 5'-TCAGCCGAACGCCTCGTCGAGGATCTCCTG-3' Primers for fragment 4: (SEQ ID NO: 9) 5'-TGGCCAACATAAGTTCACCATTCGGGCAAAAC-3' (SEQ ID NO: 10) 5'-TCTCTTCAACCAGCCATTCGTTTTGCCCG-3'
[0450] Functional expression of the alpha-ketoglutarate decarboxylase was demonstrated using both in vitro and in vivo assays. The SucA enzyme activity was measured by following a previously reported method (Tian et al., Proc. Natl. Acad. Sci. USA 102:10670-10675 (2005)). The reaction mixture contained 50 mM potassium phosphate buffer, pH 7.0, 0.2 mM thiamine pyrophosphate, 1 mM MgCl.sub.2, 0.8 mM ferricyanide, 1 mM alpha-ketoglutarate and cell crude lysate. The enzyme activity was monitored by the reduction of ferricyanide at 430 nm. The in vivo function of the SucA enzyme was verified using E. coli whole-cell culture. Single colonies of E. coli MG1655 lacI.sup.q transformed with plasmids encoding the SucA enzyme and the 4-hydroxybutyrate dehydrogenase (4Hbd) was inoculated into 5 mL of LB medium containing appropriate antibiotics. The cells were cultured at 37.degree. C. overnight aerobically. A 200 uL of this overnight culture was introduced into 8 mL of M9 minimal medium (6.78 g/L Na.sub.2HPO.sub.4, 3.0 g/L KH.sub.2PO.sub.4, 0.5 g/L NaCl, 1.0 g/L NH.sub.4Cl, 1 mM MgSO.sub.4, 0.1 mM CaCl.sub.2) supplemented with 20 g/L glucose, 100 mM 3-(N-morpholino)propanesulfonic acid (MOPS) to improve the buffering capacity, 10 .mu.g/mL thiamine, and the appropriate antibiotics. Microaerobic conditions were established by initially flushing capped anaerobic bottles with nitrogen for 5 minutes, then piercing the septum with a 23G needle following inoculation. The needle was kept in the bottle during growth to allow a small amount of air to enter the bottles. The protein expression was induced with 0.2 mM isopropyl .beta.-D-1-thiogalactopyranoside (IPTG) when the culture reached mid-log growth phase. As controls, E. coli MG1655 lacI.sup.q strains transformed with only the plasmid encoding the 4-hydroxybutyrate dehydrogenase and only the empty vectors were cultured under the same condition (see Table 23). The accumulation of 4-hydroxybutyrate (4HB) in the culture medium was monitored using LCMS method. Only the E. coli strain expressing the Mycobacterium alpha-ketoglutarate decarboxylase produced significant amount of 4HB (see FIG. 16).
TABLE-US-00075 TABLE 24 Three strains containing various plasmid controls and encoding sucA and 4-hydroxybutyrate dehydrogenase. Host pZE13 pZA33 1 MG1655 lacIq vector vector 2 MG1655 lacIq vector 4hbd 3 MG1655 lacIq sucA 4hbd
[0451] A separate experiment demonstrated that the alpha-ketoglutarate decarboxylase pathway functions independently of the reductive TCA cycle. E. coli strain ECKh-401 (.DELTA.adhE .DELTA.ldhA .DELTA.pflB .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh .DELTA.arcA) was used as the host strain (see Table 25). All the three constructs contained the gene encoding 4HB dehydrogenase (4Hbd). Construct 1 also contained the gene encoding the alpha-ketoglutarate decarboxylase (sucA). Construct 2 contained the genes encoding the succinyl-CoA synthetase (sucCD) and the CoA-dependent succinate semialdehyde dehydrogenase (sucD), which are required for the synthesis of 4HB via the reductive TCA cycle. Construct 3 contains all the genes from 1 and 2. The three E. coli strains were cultured under the same conditions as described above except the second culture was under the micro-aerobic condition. By expressing the SucA enzyme, construct 3 produced more 4HB than construct 2, which relies on the reductive TCA cycle for 4HB synthesis (see FIG. 17).
[0452] Further support for the contribution of alpha-ketoglutarate decarboxylase to production of 4HB and BDO was provided by flux analysis experiments. Cultures of ECKh-432, which contains both sucCD-sucD and sucA on the chromosome, were grown in M9 minimal medium containing a mixture of 1-13C-glucose (60%) and U-13C-glucose (40%). The biomass was harvested, the protein isolated and hydrolyzed to amino acids, and the label distribution of the amino acids analyzed by gas chromatography-mass spectrometry (GCMS) as described previously (Fischer and Sauer, Eur. J. Biochem. 270:880-891 (2003)). In addition, the label distribution of the secreted 4HB and BDO was analyzed by GCMS as described in WO2008115840 A2. This data was used to calculate the intracellular flux distribution using established methods (Suthers et al., Metab. Eng. 9:387-405 (2007)). The results indicated that between 56% and 84% of the alpha-ketoglutarate was channeled through alpha-ketoglutarate decarboxylase into the BDO pathway. The remainder was oxidized by alpha-ketoglutarate dehydrogenase, which then entered BDO via the succinyl-CoA route.
[0453] These results demonstrate 4-hydroxybutyrate producing strains that contain the sucA gene from Mycobacterium bovis BCG expressed on a plasmid. When the plasmid encoding this gene is not present, 4-hydroxybutyrate production is negligible when sucD (CoA-dependant succinate semialdehyde dehydrogenase) is not expressed. The M. bovis gene is a close homolog of the Mycobacterium tuberculosis gene whose enzyme product has been previously characterized (Tian et al., supra, 2005).
[0454] Succinate Semialdehyde Dehydrogenase (CoA-Dependent), 4-Hydroxybutyrate Dehydrogenase, and 4-Hydroxybutyryl-CoA/Acetyl-CoA Transferase.
[0455] The genes from Porphyromonas gingivalis W83 can be effective components of the pathway for 1,4-butanediol production (see also WO2008/115840, WO 2009/023493, U.S. publication 2009/0047719, U.S. publication 2009/0075351). The nucleotide sequence of CoA-dependent succinate semialdehyde dehydrogenase (sucD) from Porphyromonas gingivalis is shown in FIG. 18A, and the encoded amino acid sequence is shown in FIG. 18B. The nucleotide sequence of 4-hydroxybutyrate dehydrogenase (4hbd) from Porphymonas gingivalis is shown in FIG. 19A, and the encoded amino acid sequence is shown in FIG. 19B. The nucleotide sequence of 4-hydroxybutyrate CoA transferase (cat2) from Porphyromonas gingivalis is shown in FIG. 20A, and the encoded amino acid sequence is shown in FIG. 20B.
[0456] Briefly, the genes from Porphyromonas gingivalis W83 encoding succinate semialdehyde dehydrogenase (CoA-dependent) and 4-hydroxybutyrate dehydrogenase, and in some cases additionally 4-hydroxybutyryl-CoA/acetyl-CoA, were cloned by PCR from P. gingivalis chromosomal DNA and introduced into multicopy plasmids pZS*13, pZA13, and pZE33 behind the PA1lacO-1 promoter (Lutz and Bujard, Nucleic Acids Res. 25:1203-1210 (1997)) using standard molecular biology procedures. These plasmids were then introduced into host strains.
[0457] The Porphyromonas gingivalis W83 genes were introduced into production strains as described above. Some strains included only succinate semialdehyde dehydrogenase (CoA-dependant) and 4-hydroxybutyrate dehydrogenase without 4-hydroxybutyryl-CoA/acetyl-CoA transferase.
[0458] Butyrate Kinase and Phosphotransbutyrylase.
[0459] Butyrate kinase (BK) and phosphotransbutyrylase (PTB) enzymes can be utilized to produce 4-hydroxybutyryl-CoA (see also WO2008/115840, WO 2009/023493, U.S. publication 2009/0047719, U.S. publication 2009/0075351). In particular, the Clostridium acetobutylicum genes, buk1 and ptb, can be utilized as part of a functional BDO pathway.
[0460] Initial experiments involved the cloning and expression of the native C. acetobutylicum PTB (020) and BK (021) genes in E. coli. Where required, the start codon and stop codon for each gene were modified to "ATG" and "TAA," respectively, for more optimal expression in E. coli. The C. acetobutylicum gene sequences (020N and 021N) and their corresponding translated peptide sequences are shown in FIGS. 21 and 22.
[0461] The PTB and BK genes exist in C. acetobutylicum as an operon, with the PTB (020) gene expressed first. The two genes are connected by the sequence "atta aagttaagtg gaggaatgtt aac" (SEQ ID NO:11) that includes a re-initiation ribosomal binding site for the downstream BK (021) gene. The two genes in this context were fused to lac-controlled promoters in expression vectors for expression in E. coli (Lutz and Bujard, Nucleic Acids Res. 25:1203-1210 (1997)).
[0462] Expression of the two proteins from these vector constructs was found to be low in comparison with other exogenously expressed genes due to the high incidence of codons in the C. acetobutylicum genes that occur only rarely in E. coli. Therefore new 020 and 021 genes were predicted that changed rare codons for alternates that are more highly represented in E. coli gene sequences. This method of codon optimization followed algorithms described previously (Sivaraman et al., Nucleic Acids Res. 36:e16(2008)). This method predicts codon replacements in context with their frequency of occurrence when flanked by certain codons on either side. Alternative gene sequences for 020 (FIG. 23) and 021 (FIG. 24) were determined in which increasing numbers of rare codons were replaced by more prevalent codons (A<B<C<D) based on their incidence in the neighboring codon context. No changes in actual peptide sequence compared to the native 020 and 021 peptide sequences were introduced in these predicted sequences.
[0463] The improvement in expression of the BK and PTB proteins resulting from codon optimization is shown in FIG. 25A. Expression of the native gene sequences is shown in lane 2, while expression of the 020B-021B and 020C-021C is shown in lanes 3 and 4, respectively. Higher levels of protein expression in the codon-optimized operons 020B-021B (2021B) and 020C-021C (2021C) also resulted in increased activity compared to the native operon (2021n) in equivalently-expressed E. coli crude extracts (FIG. 25B).
[0464] The codon optimized operons were expressed on a plasmid in strain ECKh-432 (.DELTA.adhE .DELTA.ldhA .DELTA.pflB .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh .DELTA.arcA gltAR163L fimD:: E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd fimD:: M. bovis sucA, C. kluyveri 4hbd) along with the C. acetobutylicum aldehyde dehydrogenase to provide a complete BDO pathway. Cells were cultured in M9 minimal medium containing 20 g/L glucose, using a 23G needle to maintain microaerobic conditions as described above. The resulting conversion of glucose to the final product BDO was measured. Also measured was the accumulation of gamma-butyrylactone (GBL), which is a spontaneously rearranged molecule derived from 4Hb-CoA, the immediate product of the PTB-BK enzyme pair. FIG. 26 shows that expression of the native 2021n operon resulted in comparable BDO levels to an alternative enzyme function, Cat2 (034), that is capable of converting 4HB and free CoA to 4HB-CoA. GBL levels of 034 were significantly higher than 2021n, suggesting that the former enzyme has more activity than PTB-BK expressed from the native genes. However levels of both BDO and GBL were higher than either 034 or 2021n when the codon-optimized variants 2021B and 2021C were expressed, indicating that codon optimization of the genes for PTB and BK significantly increases their contributions to BDO synthesis in E. coli.
[0465] These results demonstrate that butyrate kinase (BK) and phosphotransbutyrylase (PTB) enzymes can be employed to convert 4-hydroxybutyrate to 4-hydroxybutyryl-CoA. This eliminates the need for a transferase enzyme such as 4-hydroxybutyryl-CoA/Acetyl-CoA transferase, which would generate one mole of acetate per mol of 4-hydroxybutyryl-CoA produced. The enzymes from Clostridium acetobutylicum are present in a number of engineered strains for BDO production.
[0466] 4-Hydroxybutyryl-CoA Reductase.
[0467] The Clostridium beijerinckii ald gene can be utilized as part of a functional BDO pathway (see also WO2008/115840, WO 2009/023493, U.S. publication 2009/0047719, U.S. publication 2009/0075351). The Clostridium beijerinckii ald can also be utilized to lower ethanol production in BDO producing strains. Additionally, a specific codon-optimized ald variant (GNM0025B) was found to improve BDO production.
[0468] The native C. beijerinckii ald gene (025n) and the predicted protein sequence of the enzyme are shown in FIG. 27. As was seen for the Clostridium acetobutylicum PTB and BK genes, expression of the native C. beijerinckii ald gene was very low in E. coli. Therefore, four codon-optimized variants for this gene were predicted. FIGS. 28A-28D show alternative gene sequences for 025, in which increasing numbers of rare codons are replaced by more prevalent codons (A<B<C<D) based on their incidence in the neighboring codon context (25A, P=0.05; 25B, P=0.1; 25C, P=0.15; 25D, P=1). No changes in actual peptide sequence compared to the native 025 peptide sequence were introduced in these predictions. Codon optimization significantly increased expression of the C. beijerinckii ald (see FIG. 29), which resulted in significantly higher conversion of glucose to BDO in cells expressing the entire BDO pathway (FIG. 30A).
[0469] The native and codon-optimized genes were expressed on a plasmid along with P. gingivalis Cat2, in the host strain ECKh-432 (.DELTA.adhE .DELTA.ldhA .DELTA.pflB .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh .DELTA.arcA gltAR163L .DELTA.ackA fimD:: E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd fimD:: M. bovis sucA, C. kluyveri 4hbd), thus containing a complete BDO pathway. Cells were cultured microaerobically in M9 minimal medium containing 20 g/L glucose as described above. The relative production of BDO and ethanol by the C. beijerinckii Ald enzyme (expressed from codon-optimized variant gene 025B) was compared with the C. acetobutylicum AdhE2 enzyme (see FIG. 30B). The C. acetobutylicum AdhE2 enzyme (002C) produced nearly 4 times more ethanol than BDO. In comparison, the C. beijerinckii Ald (025B) (in conjunction with an endogenous ADH activity) produced equivalent amounts of BDO, yet the ratio of BDO to ethanol production was reversed for this enzyme compared to 002C. This suggests that the C. beijerinckii Ald is more specific for 4HB-CoA over acetyl-coA than the C. acetobutylicum AdhE2, and therefore the former is the preferred enzyme for inclusion in the BDO pathway.
[0470] The Clostridium beijerinckii ald gene (Toth et al., Appl. Environ. Microbiol. 65:4973-4980 (1999)) was tested as a candidate for catalyzing the conversion of 4-hydroxybutyryl-CoA to 4-hydroxybutanal. Over fifty aldehyde dehydrogenases were screened for their ability to catalyze the conversion of 4-hydroxybutyryl-CoA to 4-hydroxybutyraldehyde. The C. beijerinckii ald gene was chosen for implementation into BDO-producing strains due to the preference of this enzyme for 4-hydroxybutyryl-CoA as a substrate as opposed to acetyl-CoA. This is important because most other enzymes with aldehyde dehydrogenase functionality (for example, adhE2 from C. acetobutylicum (Fontaine et al., J Bacteriol. 184:821-830 (2002)) preferentially convert acetyl-CoA to acetaldehyde, which in turn is converted to ethanol. Utilization of the C. beijerinckii gene lowers the amount of ethanol produced as a byproduct in BDO-producing organisms. Also, a codon-optimized version of this gene expresses very well in E. coli (Sivaraman et al., Nucleic Acids Res. 36:e16 (2008)).
[0471] 4-Hydroxybutanal Reductase.
[0472] 4-hydroxybutanal reductase activity of adh1 from Geobacillus thermoglucosidasius (M10EXG) was utilized. This led to improved BDO production by increasing 4-hydroxybutanal reductase activity over endogenous levels.
[0473] Multiple alcohol dehydrogenases were screened for their ability to catalyze the reduction of 4-hydroxybutanal to BDO. Most alcohol dehydrogenases with high activity on butyraldehyde exhibited far lower activity on 4-hydroxybutyraldehyde. One notable exception is the adh1 gene from Geobacillus thermoglucosidasius M10EXG (Jeon et al., J Biotechnol. 135:127-133 (2008)) (GNM0084), which exhibits high activity on both 4-hydroxybutanal and butanal.
[0474] The native gene sequence and encoded protein sequence if the adh1 gene from Geobacillus thermoglucosidasius are shown in FIG. 31. The G. thermoglucosidasius ald1 gene was expressed in E. coli.
[0475] The Adh1 enzyme (084) expressed very well from its native gene in E. coli (see FIG. 32A). In ADH enzyme assays, the E. coli expressed enzyme showed very high reductive activity when butyraldehyde or 4HB-aldehyde were used as the substrates (see FIG. 32B). The Km values determined for these substrates were 1.2 mM and 4.0 mM, respectively. These activity values showed that the Adh1 enzyme was the most active on reduction of 4HB-aldehyde of all the candidates tested.
[0476] The 084 enzyme was tested for its ability to boost BDO production when coupled with the C. beijerinckii ald. The 084 gene was inserted behind the C. beijerinckii ald variant 025B gene to create a synthetic operon that results in coupled expression of both genes. Similar constructs linked 025B with other ADH candidate genes, and the effect of including each ADH with 025B on BDO production was tested. The host strain used was ECKh-459 (.DELTA.adhE ldhA .DELTA.pflB .DELTA.lpdA::fnr-pflB6-K.p.lpdA322 .DELTA.mdh .DELTA.arcA gltAR163L fimD:: E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd fimD:: M. bovis sucA, C. kluyveri 4hbd fimD:: C. acetobutylicum buk1, C. acetobutylicum ptb), which contains the remainder of the BDO pathway on the chromosome. The 084 ADH expressed in conjunction with 025B showed the highest amount of BDO (right arrow in FIG. 33) when compared with 025B only (left arrow in FIG. 33) and in conjunction with endogenous ADH functions. It also produced more BDO than did other ADH enzymes when paired with 025B, indicated as follows: 026A-C, codon-optimized variants of Clostridium acetobutylicum butanol dehydrogenase; 050, Zymomonas mobilis alcohol dehydrogenase I; 052, Citrobacter freundii 1,3-propanediol dehydrogenase; 053, Lactobacillus brevis 1,3-propanediol dehydrogenase; 057, Bacteroides fragilis lactaldehyde reductase; 058, E. coli 1,3-propanediol dehydrogenase; 071, Bacillus subtilis 168 alpha-ketoglutarate semialdehyde dehydrogenase. The constructs labeled "PT5lacO" are those in which the genes are driven by the PT5lacO promoter. In all other cases, the PA1lacO-1 promoter was used. This shows that inclusion of the 084 ADH in the BDO pathway increased BDO production.
Example XIV
BDO Producing Strains Expressing Pyruvate Dehydrogenase
[0477] This example describes the utilization of pyruvate dehydrogenase (PDH) to enhance BDO production. Heterologous expression of the Klebsiella pneumonia lpdA gene was used to enhance BDO production.
[0478] Computationally, the NADH-generating conversion of pyruvate to acetyl-CoA is required to reach the maximum theoretical yield of 1,4-butanediol (see also WO2008/115840, WO 2009/023493, U.S. publication 2009/0047719, U.S. publication 2009/0075351; WO 2008/018930; Kim et al., Appl. Environ. Microbiol. 73:1766-1771 (2007); Kim et al., J. Bacteriol. 190:3851-3858 (2008); Menzel et al., J. Biotechnol. 56:135-142 (1997)). Lack of PDH activity was shown to reduce the maximum anaerobic theoretical yield of BDO by 11% if phosphoenolpyruvate carboxykinase (PEPCK) activity cannot be attained and by 3% if PEPCK activity can be attained. More importantly, however, absence of PDH activity in the OptKnock strain #439, described in WO 2009/023493 and U.S. publication 2009/0047719, which has the knockout of ADHEr, ASPT, LDH_D, MDH and PFLi, would reduce the maximum anaerobic yield of BDO by 54% or by 43% if PEPCK activity is absent or present, respectively. In the presence of an external electron acceptor, lack of PDH activity would reduce the maximum yield of the knockout strain by 10% or by 3% assuming that PEPCK activity is absent or present, respectively.
[0479] PDH is one of the most complicated enzymes of central metabolism and is comprised of 24 copies of pyruvate decarboxylase (E1) and 12 molecules of dihydrolipoyl dehydrogenase (E3), which bind to the outside of the dihydrolipoyl transacetylase (E2) core. PDH is inhibited by high NADH/NAD, ATP/ADP, and Acetyl-CoA/CoA ratios. The enzyme naturally exhibits very low activity under oxygen-limited or anaerobic conditions in organisms such as E. coli due in large part to the NADH sensitivity of E3, encoded by lpdA. To this end, an NADH-insensitive version of the lpdA gene from Klebsiella pneumonia was cloned and expressed to increase the activity of PDH under conditions where the NADH/NAD ratio is expected to be high.
[0480] Replacement of the Native lpdA.
[0481] The pyruvate dehydrogenase operon of Klebsiella pneumoniae is between 78 and 95% identical at the nucleotide level to the equivalent operon of E. coli. It was shown previously that K. pneumoniae has the ability to grow anaerobically in presence of glycerol (Menzel et al., J. Biotechnol. 56:135-142 (1997); Menzel et al., Biotechnol. Bioeng. 60:617-626 (1998)). It has also been shown that two mutations in the lpdA gene of the operon of E. coli would increase its ability to grow anaerobically (Kim et al. Appl. Environ. Microbiol. 73:1766-1771 (2007); Kim et al., J. Bacteriol. 190:3851-3858 (2008)). The lpdA gene of K. pneumonia was amplified by PCR using genomic DNA (ATCC700721D) as template and the primers KP-lpdA-Bam (5'-acacgcggatccaacgtcccgg-3')(SEQ ID NO:12) and KP-lpdA-Nhe (5'-agcggctccgctagccgcttatg-3')(SEQ ID NO:13). The resulting fragment was cloned into the vector pCR-BluntII-TOPO (Invitrogen; Carlsbad Calif.), leading to plasmid pCR-KP-lpdA.
[0482] The chromosomal gene replacement was performed using a non-replicative plasmid and the sacB gene from Bacillus subtilis as a means of counterselection (Gay et al., J. Bacteriol. 153:1424-1431 (1983)). The vector used is pRE118 (ATCC87693) deleted of the oriT and IS sequences, which is 3.6 kb in size and carrying the kanamycin resistance gene. The sequence was confirmed, and the vector was called pRE118-V2 (see FIG. 34).
[0483] The E. coli fragments flanking the lpdA gene were amplified by PCR using the combination of primers: EC-aceF-Pst (5'-aagccgttgctgcagctcttgagc-3')(SEQ ID NO:14)+EC-aceF-Bam2 (5'-atctccggcggtcggatccgtcg-3')(SEQ ID NO:15) and EC-yacH-Nhe (5'-aaagcggctagccacgccgc-3')(SEQ ID NO:16)+EC-yacH-Kpn (5'-attacacgaggtacccaacg-3')(SEQ ID NO:17). A BamHI-XbaI fragment containing the lpdA gene of K. pneumonia was isolated from plasmid pCR-KP-lpdA and was then ligated to the above E. coli fragments digested with PstI+BamHI and NheI-KpnI respectively, and the pRE118-V2 plasmid digested with KpnI and PstI. The resulting plasmid (called pRE118-M2.1 lpdA yac) was subjected to Site Directed Mutagenesis (SDM) using the combination of primers KP-lpdA-HisTyr-F (5'-atgctggcgtacaaaggtgtcc-3')(SEQ ID NO:18) and (5'-ggacacctttgtacgccagcat-3')(SEQ ID NO:19) for the mutation of the His 322 residue to a Tyr residue or primers KP-lpdA-GluLys-F (5'-atcgcctacactaaaccagaagtgg-3')(SEQ ID NO:20) and KP-lpdA-GluLys-R (5'-ccacttctggtttagtgtaggcgat-3')(SEQ ID NO:21) for the mutation of the residue Glu 354 to Lys residue. PCR was performed with the Polymerase Pfu Turbo (Stratagene; San Diego Calif.). The sequence of the entire fragment as well as the presence of only the desired mutations was verified. The resulting plasmid was introduced into electro competent cells of E. coli .DELTA.adhE::Frt-.DELTA.ldhA::Frt by transformation. The first integration event in the chromosome was selected on LB agar plates containing Kanamycin (25 or 50 mg/L). Correct insertions were verified by PCR using 2 primers, one located outside the region of insertion and one in the kanamycin gene (5'-aggcagttccataggatggc-3')(SEQ ID NO:22). Clones with the correct insertion were selected for resolution. They were sub-cultured twice in plain liquid LB at the desired temperature and serial dilutions were plated on LB-no salt-sucrose 10% plates. Clones that grew on sucrose containing plates were screened for the loss of the kanamycin resistance gene on LB-low salt agar medium and the lpdA gene replacement was verified by PCR and sequencing of the encompassing region. Sequence of the insertion region was verified, and is as described below. One clone (named 4-4-P1) with mutation Glu354Lys was selected. This clone was then transduced with P1 lysate of E. coli .DELTA.PflB::Frt leading to strain ECKh-138 (.DELTA.adhE .DELTA.ldhA .DELTA.pflB .DELTA.lpdA::K.p.lpdA322).
[0484] The sequence of the ECKh-138 region encompassing the aceF and lpdA genes is shown in FIG. 35. The K. pneumonia lpdA gene is underlined, and the codon changed in the Glu354Lys mutant shaded. The protein sequence comparison of the native E. coli lpdA and the mutant K. pneumonia lpdA is shown in FIG. 36.
[0485] To evaluate the benefit of using K. pneumoniae lpdA in a BDO production strain, the host strains AB3 and ECKh-138 were transformed with plasmids expressing the entire BDO pathway from strong, inducible promoters. Specifically, E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd were expressed on the medium copy plasmid pZA33, and P. gingivalis Cat2 and C. acetobutylicum AdhE2 were expressed on the high copy plasmid pZE13. These plasmids have been described in the literature (Lutz and H. Bujard, Nucleic Acids Res 25:1203-1210 (1997)), and their use for BDO pathway expression is described in Example XIII and WO2008/115840.
[0486] Cells were grown anaerobically at 37.degree. C. in M9 minimal medium (6.78 g/L Na.sub.2HPO.sub.4, 3.0 g/L KH.sub.2PO.sub.4, 0.5 g/L NaCl, 1.0 g/L NH.sub.4Cl, 1 mM MgSO.sub.4, 0.1 mM CaCl.sub.2) supplemented with 20 g/L glucose, 100 mM 3-(N-morpholino)propanesulfonic acid (MOPS) to improve the buffering capacity, 10 .mu.g/mL thiamine, and the appropriate antibiotics. Microaerobic conditions were established by initially flushing capped anaerobic bottles with nitrogen for 5 minutes, then piercing the septum with a 23G needle following inoculation. The needle was kept in the bottle during growth to allow a small amount of air to enter the bottles. 0.25 mM IPTG was added when OD600 reached approximately 0.2 to induce the pathway genes, and samples taken for analysis every 24 hours following induction. The culture supernatants were analyzed for BDO, 4HB, and other by-products as described in Example II and in WO2008/115840. BDO and 4HB production in ECKh-138 was significantly higher after 48 hours than in AB3 or the host used in previous work, MG1655 .DELTA.ldhA (FIG. 37).
[0487] PDH Promoter Replacement.
[0488] It was previously shown that the replacement of the pdhR repressor by a transcriptional fusion containing the Fnr binding site, one of the pflB promoters, and its ribosome binding site (RBS), thus leading to expression of the aceEF-lpd operon by an anaerobic promoter, should increase pdh activity anaerobically (Zhou et al., Biotechnol. Lett. 30:335-342 (2008)). A fusion containing the Fnr binding site, the pflB-p6 promoter and an RBS binding site were constructed by overlapping PCR. Two fragments were amplified, one using the primers aceE-upstream-RC (5'-tgacatgtaacacctaccttctgtgcctgtgccagtggttgctgtgatatagaag-3')(SEQ ID NO:23) and pflBp6-Up-Nde (5'-ataataatacatatgaaccatgcgagttacgggcctataagccaggcg-3')(SEQ ID NO:24) and the other using primers aceE-EcoRV-EC (5'-agtttttcgatatctgcatcagacaccggcacattgaaacgg-3')(SEQ ID NO:25) and aceE-upstream (5'-ctggcacaggcacagaaggtaggtgttacatgtcagaacgtttacacaatgacgtggatc-3')(SEQ ID NO:26). The tw fragments were assembled by overlapping PCR, and the final DNA fragment was digested with the restriction enzymes NdeI and BamHI. This fragment was subsequently introduced upstream of the aceE gene of the E. coli operon using pRE118-V2 as described above. The replacement was done in strains ECKh-138 and ECKh-422. The nucleotide sequence encompassing the 5' region of the aceE gene was verified and is shown in FIG. 37. FIG. 37 shows the nucleotide sequence of 5' end of the aceE gene fused to the pflB-p6 promoter and ribosome binding site (RBS). The 5' italicized sequence shows the start of the aroP gene, which is transcribed in the opposite direction from the pdh operon. The 3' italicized sequence shows the start of the aceE gene. In upper case: pflB RBS. Underlined: FNR binding site. In bold: pflB-p6 promoter sequence.
[0489] lpdA Promoter Replacement.
[0490] The promoter region containing the fnr binding site, the pflB-p6 promoter and the RBS of the pflB gene was amplified by PCR using chromosomal DNA template and primers aceF-pflBp6-fwd (5'-agacaaatcggttgccgtttgttaagccaggcgagatatgatctatatc-3')(SEQ ID NO:27) and lpdA-RB S-B-rev (5'-gagttttgatttcagtactcatcatgtaacacctaccttcttgctgtgatatag-3')(SEQ ID NO:28). Plasmid 2-4a was amplified by PCR using primers B-RBS-lpdA fwd (5'-ctatatcacagcaagaaggtaggtgttacatgatgagtactgaaatcaaaactc-3')(SEQ ID NO:29) and pflBp6-aceF-rev (5'-gatatagatcatatctcgcctggcttaacaaacggcaaccgatttgtct-3')(SEQ ID NO:30). The two resulting fragments were assembled using the BPS cloning kit (BPS Bioscience; San Diego Calif.). The resulting construct was sequenced verified and introduced into strain ECKh-439 using the pRE118-V2 method described above. The nucleotide sequence encompassing the aceF-lpdA region in the resulting strain ECKh-456 is shown in FIG. 39.
[0491] The host strain ECKh-439 (.DELTA.adhE .DELTA.ldhA .DELTA.pflB .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh .DELTA.arcA gltAR163L ackA fimD:: E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd fimD:: M. bovis sucA, C. kluyveri 4hbd), the construction of which is described below, and the pdhR and lpdA promoter replacement derivatives ECKh-455 and ECKh-456, were tested for BDO production. The strains were transformed with pZS*13 containing P. gingivalis Cat2 and C. beijerinckii Ald to provide a complete BDO pathway. Cells were cultured in M9 minimal medium supplemented with 20 g/L glucose as described above. 48 hours after induction with 0.2 mM IPTG, the concentrations of BDO, 4HB, and pyruvate were as shown in FIG. 40. The promoter replacement strains produce slightly more BDO than the isogenic parent.
[0492] These results demonstrated that expression of pyruvate dehydrogenase increased production of BDO in BDO producing strains.
Example XV
BDO Producing Strains Expressing Citrate Synthase and Aconitase
[0493] This example describes increasing activity of citrate synthase and aconitase to increase production of BDO. An R163L mutation into gltA was found to improve BDO production. Additionally, an arcA knockout was used to improve BDO production.
[0494] Computationally, it was determined that flux through citrate synthase (CS) and aconitase (ACONT) is required to reach the maximum theoretical yield of 1,4-butanediol (see also WO2008/115840, WO 2009/023493, U.S. publication 2009/0047719, U.S. publication 2009/0075351). Lack of CS or ACONT activity would reduce the maximum theoretical yield by 14% under anaerobic conditions. In the presence of an external electron acceptor, the maximum yield is reduced by 9% or by 6% without flux through CS or ACONT assuming the absence or presence of PEPCK activity, respectively. As with pyruvate dehydrogenase (PDH), the importance of CS and ACONT is greatly amplified in the knockout strain background in which ADHEr, ASPT, LDH_D, MDH and PFLi are knocked out (design #439)(see WO 2009/023493 and U.S. publication 2009/0047719, which is incorporated herein by reference).
[0495] The minimal OptKnock strain design described in WO 2009/023493 and U.S. publication 2009/0047719 had one additional deletion beyond ECKh-138, the mdh gene, encoding malate dehydrogenase. Deletion of this gene is intended to prevent flux to succinate via the reductive TCA cycle. The mdh deletion was performed using the .lamda. red homologeous recombination method (Datsenko and Wanner, Proc. Natl. Acad. Sci. USA 97:6640-6645 (2000)). The following oligonucleotides were used to PCR amplify the chloramphenicol resistance gene (CAT) flanked by FRT sites from pKD3:
TABLE-US-00076 S-mdh-Kan (SEQ ID NO: 31) 5'-TAT TGT GCA TAC AGA TGA ATT TTT ATG CAA ACA GTC AGC CCT GAA GAA GGG TGT AGG CTG GAG CTG CTT C-3' AS-mdh-Kan (SEQ ID NO: 32) 5'-CAA AAA ACC GGA GTC TGT GCT CCG GTT TTT TAT TAT CCG CTA ATC AAT TAC ATA TGA ATA TCC TCC TTA G-3'.
Underlined regions indicate homology to pKD3 plasmid and bold sequence refers to sequence homology upstream and downstream of the mdh ORF. After purification, the PCR product was electroporated into ECKh-138 electrocompetent cells that had been transformed with pRedET (tet) and prepared according to the manufacturer's instructions (genebridges.com/gb/pdf/K001%20Q%20E%20BAC%20Modification%20Kit-version2.- 6-2007-screen.pdf). The PCR product was designed so that it integrated into the ECKh-138 genome at a region upstream of the mdh gene, as shown in FIG. 41.
[0496] Recombinants were selected for chloramphenicol resistance and streak purified. Loss of the mdh gene and insertion of CAT was verified by diagnostic PCR. To remove the CAT gene, a temperature sensitive plasmid pCP20 containing a FLP recombinase (Datsenko and Wanner, Proc. Natl. Acad. Sci. USA 97:6640-6645 (2000)) was transformed into the cell at 30.degree. C. and selected for ampicillin resistance (AMP). Transformants were grown nonselectively at 42.degree. C. overnight to thermally induce FLP synthesis and to cause lose of the plasmid. The culture was then streak purified, and individual colonies were tested for loss of all antibiotic resistances. The majority lost the FRT-flanked resistance gene and the FLP helper plasmid simultaneously. There was also a "FRT" scar leftover. The resulting strain was named ECKh-172.
[0497] CS and ACONT are not highly active or highly expressed under anaerobic conditions. To this end, the arcA gene, which encodes for a global regulator of the TCA cycle, was deleted. ArcA works during microaerobic conditions to induce the expression of gene products that allow the activity of central metabolism enzymes that are sensitive to low oxygen levels, aceE, pflB and adhE. It was shown that microaerobically, a deletion in arcA/arcB increases the specific activities of ldh, icd, gltA, mdh, and gdh genes (Salmon et al., J. Biol. Chem. 280:15084-15096 (2005); Shalel-Levanon et al., Biotechnol. Bioeng. 92(2):147-159 (2005). The upstream and downstream regions of the arcA gene of E. coli MG1655 were amplified by PCR using primers ArcA-up-EcoRI (5'-ataataatagaattcgtttgctacctaaattgccaactaaatcgaaacagg-3')(SEQ ID NO:33) with ArcA-up-KpnI (5'-tattattatggtaccaatatcatgcagcaaacggtgcaacattgccg-3')(SEQ ID NO:34) and ArcA-down-EcoRI (5'-tgatctggaagaattcatcggattaccaccgtcaaaaaaaacggcg-3')(SEQ ID NO:35) with ArcA-down-PstI (5'-ataaaaccctgcagcggaaacgaagttttatccatttttggttacctg-3')(SEQ ID NO:36), respectively. These fragments were subsequently digested with the restriction enzymes EcoRI and KpnI (upstream fragment) and EcoRI and PstI (downstream). They were then ligated into the pRE118-V2 plasmid digested with PstI and KpnI, leading to plasmid pRE118-.DELTA.arcA. The sequence of plasmid pRE118-.DELTA.arcA was verified. pRE118-.DELTA.arcA was introduced into electro-competent cells of E. coli strain ECKh-172 (.DELTA.adhE .DELTA.ldhA .DELTA.pflB .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh). After integration and resolution on LB-no salt-sucrose plates as described above, the deletion of the arcA gene in the chromosome of the resulting strain ECKh-401 was verified by sequencing and is shown in FIG. 42.
[0498] The gltA gene of E. coli encodes for a citrate synthase. It was previously shown that this gene is inhibited allosterically by NADH, and the amino acids involved in this inhibition have been identified (Pereira et al., J. Biol. Chem. 269(1):412-417 (1994); Stokell et al., J. Biol. Chem. 278(37):35435-35443 (2003)). The gltA gene of E. coli MG1655 was amplified by PCR using primers gltA-up (5'-ggaagagaggctggtacccagaagccacagcagga-3')(SEQ ID NO:37) and gltA-PstI (5'-gtaatcactgcgtaagcgccatgccccggcgttaattc-3')(SEQ ID NO:38). The amplified fragment was cloned into pRE118-V2 after digestion with KpnI and PstI. The resulting plasmid was called pRE118-gltA. This plasmid was then subjected to site directed mutagensis (SDM) using primers R163L-f (5'-attgccgcgttcctcctgctgtcga-3')(SEQ ID NO:39) and R163L-r (5'-cgacagcaggaggaacgcggcaat-3')(SEQ ID NO:40) to change the residue Arg 163 to a Lys residue. The sequence of the entire fragment was verified by sequencing. A variation of the .lamda. red homologeous recombination method (Datsenko and Wanner, Proc. Natl. Acad. Sci. USA 97:6640-6645 (2000)) was used to replace the native gltA gene with the R163L mutant allele without leaving a Frt scar. The general recombination procedure is the same as used to make the mdh deletion described above. First, the strain ECKh-172 was made streptomycin resistant by introducing an rpsL null mutation using the X red homologeous recombination method. Next, a recombination was done to replace the entire wild-type gltA coding region in this strain with a cassette comprised of a kanamycin resistance gene (kanR) and a wild-type copy of the E. coli rpsL gene. When introduced into an E. coli strain harboring an rpsL null mutation, the cassette causes the cells to change from resistance to the drug streptomycin to streptomycin sensitivity. DNA fragments were then introduced that included each of the mutant versions of the gltA gene along with appropriate homologous ends, and resulting colony growth was tested in the presence of streptomycin. This selected for strains in which the kanR/rpsL cassette had been replaced by the mutant gltA gene. Insertion of the mutant gene in the correct locus was confirmed by PCR and DNA sequencing analyses. The resulting strain was called ECKh-422, and has the genotype .DELTA.adhE .DELTA.ldhA .DELTA.pflB .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh .DELTA.arcA gltAR163L. The region encompassing the mutated gltA gene of strain ECKh-422 was verified by sequencing, as shown in FIG. 43.
[0499] Crude extracts of the strains ECKh-401 and the gltAR163L mutant ECKh-422 were then evaluated for citrate synthase activity. Cells were harvested by centrifugation at 4,500 rpm (Beckman-Coulter, Allegera X-15R; Fullerton Calif.) for 10 min. The pellets were resuspended in 0.3 mL BugBuster (Novagen/EMD; San Diego Calif.) reagent with benzonase and lysozyme, and lysis proceeded for 15 minutes at room temperature with gentle shaking. Cell-free lysate was obtained by centrifugation at 14,000 rpm (Eppendorf centrifuge 5402; Hamburg Germany) for 30 min at 4.degree. C. Cell protein in the sample was determined using the method of Bradford (Bradford, Anal. Biochem. 72:248-254 (1976)).
[0500] Citrate synthase activity was determined by following the formation of free coenzyme A (HS-CoA), which is released from the reaction of acetyl-CoA with oxaloacetate. The free thiol group of HS-CoA reacts with 5,5'-dithiobis-(2-nitrobenzoic acid) (DTNB) to form 5-thio-2-nitrobenzoic acid (TNB). The concentration of TNB is then monitored spectrophotometrically by measuring the absorbance at 410 nm (maximum at 412 nm). The assay mixture contained 100 mM Tris/HCl buffer (pH 7.5), 20 mM acetyl-CoA, 10 mM DTNB, and 20 mM oxaloacetate. For the evaluation of NADH inhibition, 0.4 mM NADH was also added to the reaction. The assay was started by adding 5 microliters of the cell extract, and the rate of reaction was measured by following the absorbance change over time. A unit of specific activity is defined as the .mu.mol of product converted per minute per mg protein.
[0501] FIG. 44 shows the citrate synthase activity of wild type gltA gene product and the R163L mutant. The assay was performed in the absence or presence of 0.4 mM NADH.
[0502] Strains ECKh-401 and ECKh-422 were transformed with plasmids expressing the entire BDO pathway. E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd, and M. bovis sucA were expressed on the low copy plasmid pZS*13, and P. gingivalis Cat2 and C. acetobutylicum AdhE2 were expressed on the medium copy plasmid pZE23. Cultures of these strains were grown microaerobically in M9 minimal medium supplemented with 20 g/L glucose and the appropriate antibiotics as described above. The 4HB and BDO concentrations at 48 hours post-induction averaged from duplicate cultures are shown in FIG. 45. Both are higher in ECKh-422 than in ECKh-401, demonstrating that the enhanced citrate synthase activity due to the gltA mutation results in increased flux to the BDO pathway.
[0503] The host strain modifications described in this section were intended to redirect carbon flux through the oxidative TCA cycle, which is consistent with the OptKnock strain design described in WO 2009/023493 and U.S. publication 2009/0047719. To demonstrate that flux was indeed routed through this pathway, .sup.13C flux analysis was performed using the strain ECKh-432, which is a version of ECKh-422 in which the upstream pathway is integrated into the chromosome (as described in Example XVII). To complete the BDO pathway, P. gingivalis Cat2 and C. beijerinckii Ald were expressed from pZS*13. Four parallel cultures were grown in M9 minimal medium (6.78 g/L Na.sub.2HPO.sub.4, 3.0 g/L KH.sub.2PO.sub.4, 0.5 g/L NaCl, 1.0 g/L NH.sub.4Cl, 1 mM MgSO.sub.4, 0.1 mM CaCl.sub.2) containing 4 g/L total glucose of four different labeling ratios (.sup.1-13C, only the first carbon atom in the glucose molecule is labeled with .sup.13C; uniform-.sup.13C, all carbon atoms are .sup.13C): [0504] 1. 80 mol % unlabeled, 20 mol % uniform-.sup.13C [0505] 2. 10 mol % unlabeled, 90 mol % uniform-.sup.13C [0506] 3. 90 mol % .sup.1-13C, 10 mol % uniform-.sup.13C [0507] 4. 40 mol % .sup.1-13C, 60 mol % uniform-.sup.13C
[0508] Parallel unlabeled cultures were grown in duplicate, from which frequent samples were taken to evaluate growth rate, glucose uptake rate, and product formation rates. In late exponential phase, the labeled cultures were harvested, the protein isolated and hydrolyzed to amino acids, and the label distribution of the amino acids analyzed by gas chromatography-mass spectrometry (GCMS) as described previously (Fischer and Sauer, Eur. J. Biochem. 270:880-891 (2003)). In addition, the label distribution of the secreted 4HB and BDO in the broth from the labeled cultures was analyzed by GCMS as described in WO2008115840. This data was collectively used to calculate the intracellular flux distribution using established methods (Suthers et al., Metab. Eng. 9:387-405 (2007)). The resulting central metabolic fluxes and associated 95% confidence intervals are shown in FIG. 46. Values are molar fluxes normalized to a glucose uptake rate of 1 mmol/hr. The result indicates that carbon flux is routed through citrate synthase in the oxidative direction, and that most of the carbon enters the BDO pathway rather than completing the TCA cycle. Furthermore, it confirms there is essentially no flux between malate and oxaloacetate due to the mdh deletion in this strain.
[0509] The advantage of using a knockout strain such as strains designed using OptKnock for BDO production (see WO 2009/023493 and U.S. publication 2009/0047719) can be observed by comparing typical fermentation profiles of ECKh-422 with that of the original strain ECKh-138, in which BDO is produced from succinate via the reductive TCA cycle (see FIG. 47). Fermentations were performed with 1 L initial culture volume in 2 L Biostat B+ bioreactors (Sartorius; Cedex France) using M9 minimal medium supplemented with 20 g/L glucose. The temperature was controlled at 37.degree. C., and the pH was controlled at 7.0 using 2 M NH.sub.4OH or Na.sub.2CO.sub.3. Cells were grown aerobically to an OD600 of approximately 10, at which time the cultures were induced with 0.2 mM IPTG. One hour following induction, the air flow rate was reduced to 0.02 standard liters per minute for microaerobic conditions. The agitation rate was set at 700 rpm. Concentrated glucose was fed to maintain glucose concentration in the vessel between 0.5 and 10 g/L. Both strains were transformed with plasmids bearing the entire BDO pathway, as in the examples above. In ECKh-138, acetate, pyruvate, and 4HB dominate the fermentation, while with ECKh-422 BDO is the major product.
Example XVI
BDO Strains Expression Phosphoenolpyruvate Carboxykinase
[0510] This example describes the utilization of phosphoenolpyruvate carboxykinase (PEPCK) to enhance BDO production. The Haemophilus influenza PEPCK gene was used for heterologous expression.
[0511] Computationally, it was demonstrated that the ATP-generating conversion of oxaloacetate to phosphoenolpyruvate is required to reach the maximum theoretical yield of 1,4-butanediol (see also WO2008/115840, WO 2009/023493, U.S. publication 2009/0047719, U.S. publication 2009/0075351). Lack of PEPCK activity was shown to reduce the maximum theoretical yield of BDO by 12% assuming anaerobic conditions and by 3% assuming an external electron acceptor such as nitrate or oxygen is present.
[0512] In organisms such as E. coli, PEPCK operates in the gluconeogenic and ATP-consuming direction from oxaloacetate towards phosphoenolpyruvate. It has been hypothesized that kinetic limitations of PEPCK of E. coli prevent it from effectively catalyzing the formation of oxaloacetate from PEP. PEP carboxylase (PPC), which does not generate ATP but is required for efficient growth, is naturally utilized by E. coli to form oxaloacetate from phosphoenolpyruvate. Therefore, three non native PEPCK enzymes (Table 26) were tested for their ability to complement growth of a PPC mutant strain of E. coli in glucose minimal media.
TABLE-US-00077 TABLE 26 Sources of phosphoenolpyruvate carboxykinase sequences. Accession Number, PEPCK Source Strain GenBank Reference Sequence Haemophilus influenza NC_000907.1 Actinobacillus succinogenes YP_001343536.1 Mannheimia succiniciproducens YP_089485.1
[0513] Growth complementation studies involved plasmid based expression of the candidate genes in .DELTA.ppc mutant E. coli JW3978 obtained from the Keio collection (Baba et al., Molecular Systems Biology 2:2006.0008 (2006)). The genes were cloned behind the PA1lacO-1 promoter in the expression vectors pZA23 (medium copy) and pZE13 (high copy). These plasmids have been described previously (Lutz and Bujard, Nucleic Acids Res. 25:1203-1210 (1997)), and their use in expression BDO pathway genes has been described previously in WO2008115840.
[0514] Pre-cultures were grown aerobically in M9 minimal media with 4 g/L glucose. All pre-cultures were supplemented with aspartate (2 mM) to provide the .DELTA.ppc mutants with a source for generating TCA cycle intermediates independent of PEPCK expression. M9 minimal media was also used in the test conditions with 4 g/L glucose, but no aspartate was added and IPTG was added to 0.5 mM. Table 27 shows the results of the growth complementation studies.
TABLE-US-00078 TABLE 27 Complementation of .DELTA.ppc mutants with PEPCK from H. influenzae, A. succinogenes and M. succinoproducens when expressed from vectors pZA23 or pZE13. PEPCK Source Strain Vector Time (h) OD.sub.600 H. influenzae pZA23BB 40 0.950 .DELTA.ppc Control pZA23BB 40 0.038 A. succinogenes pZA23BB 40 0.055 M. succinoproducens pZA23BB 40 0.214 A. succinogenes pZE13BB 40 0.041 M. succinoproducens pZE13BB 40 0.024 .DELTA.ppc Control pZE13BB 40 0.042
[0515] Haemophilus influenza PEPCK was found to complement growth in .DELTA.ppc mutant E. coli best among the genes that were tested in the plasmid based screening. This gene was then integrated into the PPC locus of wild-type E. coli (MG1655) using the SacB counter selection method with pRE118-V2 discussed above (Gay et al., J. Bacteriol. 153:1424-1431 (1983)). PEPCK was integrated retaining the E. coli native PPC promoter, but utilizing the non-native PEPCK terminator. The sequence of this region following replacement of ppc by H. influenzae pepck is shown in FIG. 48. The pepck coding region is underlined.
[0516] Techniques for adaptive evolution were applied to improve the growth rate of the E. coli mutant (.DELTA.ppc::H. inf pepCK). M9 minimal media with 4 g/L glucose and 50 mM sodium bicarbonate was used to culture and evolve this strain in an anaerobic environment. The high sodium bicarbonate concentration was used to drive the equilibrium of the PEPCK reaction toward oxaloacetate formation. To maintain exponential growth, the culture was diluted 2-fold whenever an OD600 of 0.5 was achieved. After about 100 generations over 3 weeks of adaptive evolution, anaerobic growth rates improved from about 8 h to that of wild type, about 2 h. Following evolution, individual colonies were isolated, and growth in anaerobic bottles was compared to that of the initial mutant and wild-type strain (see FIG. 49). M9 medium with 4 g/L glucose and 50 mM sodium bicarbonate was used.
[0517] The ppc/pepck gene replacement procedure described above was then repeated, this time using the BDO-producing strains ECKh-432 (.DELTA.adhE .DELTA.ldhA .DELTA.pflB .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh .DELTA.arcA gltAR163L .DELTA.ackA fimD:: E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd fimD:: M. bovis sucA, C. kluyveri 4hbd) and ECKh-439 as the hosts. These strains contain the TCA cycle enhancements discussed above as well as the upstream pathway integrated in the chromosome. ECKh-439 is a derivative of ECKh-432 that has the ackA gene deleted, which encodes acetate kinase. This deletion was performed using the sacB counterselection method described above.
[0518] The .DELTA.ppc::H. inf pepCK derivative of ECKh-439, called ECKh-453, was run in a fermentation. The downstream BDO pathway was supplied by pZS*13 containing P. gingivalis Cat2 and C. beijerinckii Ald. This was performed with 1L initial culture volume in 2L Biostat B+ bioreactors (Sartorius) using M9 minimal medium supplemented with 20 g/L glucose and 50 mM NaHCO.sub.3. The temperature was controlled at 37.degree. C., and the pH was controlled at 7.0 using 2 M NH.sub.4OH or Na.sub.2CO.sub.3. Cells were grown aerobically to an OD600 of approximately 2, at which time the cultures were induced with 0.2 mM IPTG. One hour following induction, the air flow rate was reduced to 0.01 standard liters per minute for microaerobic conditions. The agitation rate was initially set at 700 rpm. The aeration rate was gradually increased throughout the fermentation as the culture density increased. Concentrated glucose solution was fed to maintain glucose concentration in the vessel between 0.5 and 10 g/L. The product profile is shown in FIG. 50. The observed phenotype, in which BDO and acetate are produced in approximately a one-to-one molar ratio, is highly similar to that predicted in WO 2009/023493 for design #439 (ADHEr, ASPT, LDH_D, MDH, PFLi). The deletion targeting the ASPT reaction was deemed unnecessary as the natural flux through aspartate ammonia-lyase is low.
[0519] A key feature of OptKnock strains is that production of the metabolite of interest is generally coupled to growth, and further, that, production should occur during exponential growth as well as in stationary phase. The growth coupling potential of ECKh-432 and ECKh-453 was evaluated by growth in microaerobic bottles with frequent sampling during the exponential phase. M9 medium containing 4 g/L glucose and either 10 mM NaHCO.sub.3 (for ECKh-432) or 50 mM NaHCO.sub.3 (for ECKh-453) was used, and 0.2 mM IPTG was included from inoculation. 18G needles were used for microaerobic growth of ECKh-432, while both 18G and 27G needles were tested for ECKh-453. The higher gauge needles result in less aeration. As shown in FIG. 51, ECKh-432 does not begin producing BDO until 5 g/L glucose has been consumed, corresponding to the onset of stationary phase. ECKh-453 produces BDO more evenly throughout the experiment. In addition, growth coupling improves as the aeration of the culture is reduced.
Example XVII
Integration of BDO Pathway Encoding Genes at Specific Integration Sites
[0520] This example describes integration of various BDO pathway genes into the fimD locus to provide more efficient expression and stability.
[0521] The entire upstream BDO pathway, leading to 4HB, has been integrated into the E. coli chromosome at the fimD locus. The succinate branch of the upstream pathway was integrated into the E. coli chromosome using the .lamda. red homologeous recombination method (Datsenko and Wanner, Proc. Natl. Acad. Sci. USA 97:6640-6645 (2000)). The recipient E. coli strain was ECKh-422 (.DELTA.adhE .DELTA.ldhA .DELTA.pflB .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh .DELTA.arcA gltAR163L). A polycistronic DNA fragment containing a promoter, the sucCD gene, the sucD gene and the 4hbd gene and a terminator sequence was inserted into the AflIII site of the pKD3 plasmid. The following primers were used to amplify the operon together with the chloramphenicol marker from the plasmid. The underlined sequences are homologeous to the target insertion site.
TABLE-US-00079 (SEQ ID NO: 41) 5'-GTTTGCACGCTATAGCTGAGGTTGTTGTCTTCCAGCAACGTACCGTA TACAATAGGCGTATCACGAGGCCCTTTC-3' (SEQ ID NO: 42) 5'-GCTACAGCATGTCACACGATCTCAACGGTCGGATGACCAATCTGGCT GGTATGGGAATTAGCCATGGTCC-3'
[0522] Following DpnI treatment and DNA electrophoresis, the purified PCR product was used to transform E. coli strain harboring plasmid pKD46. The candidate strain was selected on plates containing chloramphenicol. Genomic DNA of the candidate strain was purified. The insertion sequence was amplified and confirmed by DNA sequencing. The chloramphenicol-resistant marker was removed from chromosome by flipase. The nucleotide sequence of the region after insertion and marker removal is shown in FIG. 52.
[0523] The alpha-ketoglutarate branch of the upstream pathway was integrated into the chromosome by homologeous recombination. The plasmid used in this modification was derived from vector pRE118-V2, as referenced in Example XIV, which contains a kanamycin-resistant gene, a gene encoding the levansucrase (sacB) and a R6K conditional replication ori. The integration plasmid also contained a polycistronic sequence with a promoter, the sucA gene, the C. kluyveri 4hbd gene, and a terminator being inserted between two 1.5-kb DNA fragments that are homologeous to the flanking regions of the target insertion site. The resulting plasmid was used to transform E. coli strain. The integration candidate was selected on plates containing kanamycin. The correct integration site was verified by PCR. To resolve the antibiotic marker from the chromosome, the cells were selected for growth on medium containing sucrose. The final strain was verified by PCR and DNA sequencing. The nucleotide sequence of the chromosomal region after insertion and marker removal is shown in FIG. 53.
[0524] The resulting upstream pathway integration strain ECKh-432 was transformed with a plasmid harboring the downstream pathway genes. The construct was able to produce BDO from glucose in minimal medium (see FIG. 54).
Example XVIII
Use of a Non-Phosphotransferase Sucrose Uptake System to Reduce Pyruvate Byproduct Formation
[0525] This example describes the utilization of a non-phosphotransferase (PTS) sucrose uptake system to reduce pyruvate as a byproduct in the conversion of sucrose to BDO.
[0526] Strains engineered for the utilization of sucrose via a phosphotransferase (PTS) system produce significant amounts of pyruvate as a byproduct. Therefore, the use of a non-PTS sucrose system can be used to decrease pyruvate formation because the import of sucrose would not be accompanied by the conversion of phosphoenolpyruvate (PEP) to pyruvate. This will increase the PEP pool and the flux to oxaloacetate through PPC or PEPCK.
[0527] Insertion of a non-PTS sucrose operon into the rrnC region was performed. To generate a PCR product containing the non-PTS sucrose genes flanked by regions of homology to the rrnC region, two oligos were used to PCR amplify the csc genes from Mach1.TM. (Invitrogen, Carlsbad, Calif.). This strain is a descendent of W strain which is an E. coli strain known to be able to catabolize sucrose (Orencio-Trejo et al., Biotechnology Biofuels 1:8 (2008)). The sequence was derived from E. coli W strain KO11 (accession AY314757) (Shukla et al., Biotechnol. Lett. 26:689-693 (2004)) and includes genes encoding a sucrose permease (cscB), D-fructokinase (cscK), sucrose hydrolase (cscA), and a LacI-related sucrose-specific repressor (cscR). The first 53 amino acids of cscR was effectively removed by the placement of the AS primer. The sequences of the oligos were: rrnC 23S del S-CSC 5'-TGT GAG TGA AAG TCA CCT GCC TTA ATA TCT CAA AAC TCA TCT TCG GGT GAC GAA ATA TGG CGT GAC TCG ATA C-3' (SEQ ID NO:43) and rrnC 23S del AS-CSC 5'-TCT GTA TCA GGC TGA AAA TCT TCT CTC ATC CGC CAA AAC AGC TTC GGC GTT AAG ATG CGC GCT CAA GGA C-3' (SEQ ID NO:44). Underlined regions indicate homology to the csc operon, and bold sequence refers to sequence homology upstream and downstream of the rrnC region. The sequence of the entire PCR product is shown in FIG. 55.
[0528] After purification, the PCR product was electroporated into MG1655 electrocompetent cells which had been transformed with pRedET (tet) and prepared according to manufacturer's instructions (genebridges.com/gb/pdf/K001%20Q%20E%20BAC%20Modification%20Kit-version2.- 6-2007-screen.pdf). The PCR product was designed so that it integrated into genome into the rrnC region of the chromosome. It effectively deleted 191 nucleotides upstream of rrlC (23S rRNA), all of the rrlC rRNA gene and 3 nucleotides downstream of rrlC and replaced it with the sucrose operon, as shown in FIG. 56.
[0529] Transformants were grown on M9 minimal salts medium with 0.4% sucrose and individual colonies tested for presence of the sucrose operon by diagnostic PCR. The entire rrnC::crcAKB region was transferred into the BDO host strain ECKh-432 by P1 transduction (Sambrook et al., Molecular Cloning: A Laboratory Manual, Third Ed., Cold Spring Harbor Laboratory, New York (2001), resulting in ECKh-463 (.DELTA.adhE .DELTA.ldhA .DELTA.pflB .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh .DELTA.arcA gltAR163L fimD:: E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd fimD:: M. bovis sucA, C. kluyveri 4hbd rrnC::cscAKB). Recombinants were selected by growth on sucrose and verified by diagnostic PCR.
[0530] ECKh-463 was transformed with pZS*13 containing P. gingivalis Cat2 and C. beijerinckii Ald to provide a complete BDO pathway. Cells were cultured in M9 minimal medium (6.78 g/L Na.sub.2HPO.sub.4, 3.0 g/L KH.sub.2PO.sub.4, 0.5 g/L NaCl, 1.0 g/L NH.sub.4Cl, 1 mM MgSO.sub.4, 0.1 mM CaCl.sub.2) supplemented with 10 g/L sucrose. 0.2 mM IPTG was present in the culture from the start. Anaerobic conditions were maintained using a bottle with 23G needle. As a control, ECKh-432 containing the same plasmid was cultured on the same medium, except with 10 g/L glucose instead of sucrose. FIG. 57 shows average product concentration, normalized to culture OD600, after 48 hours of growth. The data is for 6 replicate cultures of each strain. This demonstrates that BDO production from ECKh-463 on sucrose is similar to that of the parent strain on sucrose.
Example XIX
Summary of BDO Producing Strains
[0531] This example describes various BDO producing strains.
[0532] Table 28 summarizes various BDO producing strains disclosed above in Examples XII-XVIII.
TABLE-US-00080 TABLE 28 Summary of various BDO production strains. Host Host Host Strain # Strain # chromosome Description Plasmid-based 1 .DELTA.ldhA Single deletion E. coli sucCD, P. gingivalis derivative of E. coli sucD, P. gingivalis 4hbd, MG1655 P. gingivalis Cat2, C. acetobutylicum AdhE2 2 AB3 .DELTA.adhE .DELTA.ldhA .DELTA.pflB Succinate producing E. coli sucCD, P. gingivalis strain; derivative of sucD, P. gingivalis 4hbd, E. coli MG1655 P. gingivalis Cat2, C. acetobutylicum AdhE2 3 ECKh-138 .DELTA.adhE .DELTA.ldhA .DELTA.pflB Improvement of E. coli sucCD, P. gingivalis .DELTA.lpdA::K.p.lpdA322 lpdA to increase sucD, P. gingivalis 4hbd, pyruvate P. gingivalis Cat2, dehydrogenase flux C. acetobutylicum AdhE2 4 ECKh-138 .DELTA.adhE .DELTA.ldhA .DELTA.pflB E. coli sucCD, P. gingivalis .DELTA.lpdA::K.p.lpdA322 sucD, P. gingivalis 4hbd, C. acetobutylicum buk1, C. acetobutylicum ptb, C. acetobutylicum AdhE2 5 ECKh-401 .DELTA.adhE .DELTA.ldhA .DELTA.pflB Deletions in mdh and E. coli sucCD, P. gingivalis .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh .DELTA.arcA arcA to direct flux sucD, P. gingivalis 4hbd, through oxidative P. gingivalis Cat2, TCA cycle C. acetobutylicum AdhE2 6 ECKh-401 .DELTA.adhE .DELTA.ldhA .DELTA.pflB M. bovis sucA, E. coli sucCD, .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh .DELTA.arcA P. gingivalis sucD, P. gingivalis 4hbd, P. gingivalis Cat2, C. acetobutylicum AdhE2 7 ECKh-422 .DELTA.adhE .DELTA.ldhA .DELTA.pflB Mutation in citrate E. coli sucCD, P. gingivalis .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh .DELTA.arcA synthase to improve sucD, P. gingivalis 4hbd, gltAR163L anaerobic activity P. gingivalis Cat2, C. acetobutylicum AdhE2 8 ECKh-422 .DELTA.adhE .DELTA.ldhA .DELTA.pflB M. bovis sucA, E. coli sucCD, .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh .DELTA.arcA P. gingivalis sucD, gltAR163L P. gingivalis 4hbd, P. gingivalis Cat2, C. acetobutylicum AdhE2 9 ECKh-422 .DELTA.adhE .DELTA.ldhA .DELTA.pflB M. bovis sucA, E. coli sucCD, .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh .DELTA.arcA P. gingivalis sucD, gltAR163L P. gingivalis 4hbd, P. gingivalis Cat2, C. beijerinckii Ald 10 ECKh-426 .DELTA.adhE .DELTA.ldhA .DELTA.pflB Succinate branch of P. gingivalis Cat2, .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh .DELTA.arcA upstream pathway C. beijerinckii Ald gltAR163L fimD:: E. coli sucCD, integrated into P. gingivalis sucD, P. gingivalis 4hbd ECKh-422 11 ECKh-432 .DELTA.adhE .DELTA.ldhA .DELTA.pflB Succinate and alpha- P. gingivalis Cat2, .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh .DELTA.arcA ketoglutarate C. beijerinckii Ald gltAR163L fimD:: E. coli sucCD, upstream pathway P. gingivalis sucD, P. gingivalis 4hbd branches integrated fimD:: M. bovis sucA, C. kluyveri into ECKh-422 4hbd 12 ECKh-432 .DELTA.adhE .DELTA.ldhA .DELTA.pflB C. acetobutylicum buk1, .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh .DELTA.arcA C. acetobutylicum ptb, gltAR163L fimD:: E. coli sucCD, C. beijerinckii Ald P. gingivalis sucD, P. gingivalis 4hbd fimD:: M. bovis sucA, C. kluyveri 4hbd 13 ECKh-439 .DELTA.adhE .DELTA.ldhA .DELTA.pflB Acetate kinase P. gingivalis Cat2, .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh .DELTA.arcA deletion of ECKh- C. beijerinckii Ald gltAR163L .DELTA.ackA fimD:: E. coli 432 sucCD, P. gingivalis sucD, P. gingivalis 4hbd fimD:: M. bovis sucA, C. kluyveri 4hbd 14 ECKh-453 .DELTA.adhE .DELTA.ldhA .DELTA.pflB Acetate kinase P. gingivalis Cat2, .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh .DELTA.arcA deletion and C. beijerinckii Ald gltAR163L .DELTA.ackA .DELTA.ppc::H.i.ppck PPC/PEPCK fimD:: E. coli sucCD, P. gingivalis replacement of sucD, P. gingivalis 4hbd fimD:: ECKh-432 M. bovis sucA, C. kluyveri 4hbd 15 ECKh-456 .DELTA.adhE .DELTA.ldhA .DELTA.pflB .DELTA.lpdA::fnr-pflB6- Replacement of lpdA P. gingivalis Cat2, K.p.lpdA322 .DELTA.mdh .DELTA.arcA gltAR163L promoter with C. beijerinckii Ald fimD:: E. coli sucCD, P. gingivalis anaerobic promoter sucD, P. gingivalis 4hbd fimD:: in ECKh-432 M. bovis sucA, C. kluyveri 4hbd 16 ECKh-455 .DELTA.adhE .DELTA.ldhA .DELTA.pflB .DELTA.lpdA:: Replacement of P. gingivalis Cat2, K.p.lpdA322 .DELTA.pdhR:: fnr-pflB6 .DELTA.mdh pdhR and aceEF C. beijerinckii Ald .DELTA.arcA gltAR163L fimD:: E. coli promoter with sucCD, P. gingivalis sucD, anaerobic promoter P. gingivalis 4hbd fimD:: M. bovis sucA, in ECKh-432 C. kluyveri 4hbd 17 ECKh-459 .DELTA.adhE .DELTA.ldhA .DELTA.pflB .DELTA.lpdA:: Integration of C. beijerinckii Ald K.p.lpdA322 .DELTA.mdh .DELTA.arcA gltAR163L BK/PTB into ECKh- fimD:: E. coli sucCD, P. gingivalis 432 sucD, P. gingivalis 4hbd fimD:: M. bovis sucA, C. kluyveri 4hbd fimD:: C. acetobutylicum buk1, C. acetobutylicum ptb 18 ECKh-459 .DELTA.adhE .DELTA.ldhA .DELTA.pflB .DELTA.lpdA:: C. beijerinckii Ald, K.p.lpdA322 .DELTA.mdh .DELTA.arcA gltAR163L G. thermoglucosidasius adh1 fimD:: E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd fimD:: M. bovis sucA, C. kluyveri 4hbd fimD:: C. acetobutylicum buk1, C. acetobutylicum ptb 19 ECKh-463 .DELTA.adhE .DELTA.ldhA .DELTA.pflB Non-PTS sucrose P. gingivalis Cat2, .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh genes inserted into C. beijerinckii Ald .DELTA.arcA gltAR163L fimD:: E. coli ECKh-432 sucCD, P. gingivalis sucD, P. gingivalis 4hbd fimD:: M. bovis sucA, C. kluyveri 4hbd rrnC::cscAKB 20 ECKh-463 .DELTA.adhE .DELTA.ldhA .DELTA.pflB C. acetobutylicum buk1, .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh C. acetobutylicum ptb, .DELTA.arcA gltAR163L fimD:: E. coli C. beijerinckii Ald sucCD, P. gingivalis sucD, P. gingivalis 4hbd fimD:: M. bovis sucA, C. kluyveri 4hbd rrnC::cscAKB
[0533] The strains summarized in Table 28 are as follows. Strain 1: Single deletion derivative of E. coli MG1655, with deletion of endogenous ldhA; plasmid expression of E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd, P. gingivalis Cat2, C. acetobutylicum AdhE2. Strain 2: Host strain AB3, a succinate producing strain, derivative of E. coli MG1655, with deletions of endogenous adhE ldhA pflB; plasmid expression of E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd, P. gingivalis Cat2, C. acetobutylicum AdhE2.
[0534] Strain 3: Host strain ECKh-138, deletion of endogenous adhE, ldhA, pflB, deletion of endogenous lpdA and chromosomal insertion of Klebsiella pneumoniae lpdA with a Glu354Lys mutation at the lpdA locus; plasmid expression of E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd, P. gingivalis Cat2, C. acetobutylicum AdhE2; strain provides improvement of lpdA to increase pyruvate dehydrogenase flux. Strain 4: Host strain ECKh-138, deletion of endogenous adhE, ldhA, pflB, and lpdA, chromosomal insertion of Klebsiella pneumoniae lpdA with a Glu354Lys mutation; plasmid expression E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd, C. acetobutylicum buk1, C. acetobutylicum ptb, C. acetobutylicum AdhE2.
[0535] Strain 5: Host strain ECKh-401, deletion of endogenous adhE, ldhA, pflB, deletion of endogenous lpdA and chromosomal insertion of Klebsiella pneumoniae lpdA with a Glu354Lys mutation at the lpdA locus, deletion of endogenous mdh and arcA; plasmid expression of E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd, P. gingivalis Cat2, C. acetobutylicum AdhE2; strain has deletions in mdh and arcA to direct flux through oxidative TCA cycle. Strain 6: host strain ECKh-401, deletion of endogenous adhE, ldhA, pflB, deletion of endogenous lpdA and chromosomal insertion of Klebsiella pneumoniae lpdA with a Glu354Lys mutation at the lpdA locus, deletion of endogenous mdh and arcA; plasmid expression of M. bovis sucA, E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd, P. gingivalis Cat2, C. acetobutylicum AdhE2.
[0536] Strain 7: Host strain ECKh-422, deletion of endogenous adhE, ldhA, pflB, deletion of endogenous lpdA and chromosomal insertion of Klebsiella pneumoniae lpdA with a Glu354Lys mutation at the lpdA locus, deletion of endogenous mdh and arcA, chromosomal replacement of gltA with gltA Arg163Leu mutant; plasmid expression of E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd, P. gingivalis Cat2, C. acetobutylicum AdhE2; strain has mutation in citrate synthase to improve anaerobic activity. Strain 8: strain ECKh-422, deletion of endogenous adhE, ldhA, pflB, deletion of endogenous lpdA and chromosomal insertion of Klebsiella pneumoniae lpdA with a Glu354Lys mutation at the lpdA locus, deletion of endogenous mdh and arcA, chromosomal replacement of gltA with gltA Arg163Leu mutant; plasmid expression of M. bovis sucA, E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd, P. gingivalis Cat2, C. acetobutylicum AdhE2. Strain 9: host strain ECKh-422, deletion of endogenous adhE, ldhA, pflB, deletion of endogenous lpdA and chromosomal insertion of Klebsiella pneumoniae lpdA with a Glu354Lys mutation at the lpdA locus, deletion of endogenous mdh and arcA, chromosomal replacement of gltA with gltA Arg163Leu mutant; plasmid expression of M. bovis sucA, E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd, P. gingivalis Cat2, C. beijerinckii Ald.
[0537] Strain 10: host strain ECKh-426, deletion of endogenous adhE, ldhA, pflB, deletion of endogenous lpdA and chromosomal insertion of Klebsiella pneumoniae lpdA with a Glu354Lys mutation at the lpdA locus, deletion of endogenous mdh and arcA, chromosomal replacement of gltA with gltA Arg163Leu mutant, chromosomal insertion at the fimD locus of E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd; plasmid expression of P. gingivalis Cat2, C. beijerinckii Ald; strain has succinate branch of upstream pathway integrated into strain ECKh-422 at the fimD locus. Strain 11: host strain ECKh-432, deletion of endogenous adhE, ldhA, pflB, deletion of endogenous lpdA and chromosomal insertion of Klebsiella pneumoniae lpdA with a Glu354Lys mutation at the lpdA locus, deletion of endogenous mdh and arcA, chromosomal replacement of gltA with gltA Arg163Leu mutant, chromosomal insertion at the fimD locus of E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd, chromosomal insertion at the fimD locus of M. bovis sucA, C. kluyveri 4hbd; plasmid expression of P. gingivalis Cat2, C. beijerinckii Ald; strain has succinate and alpha-ketoglutarate upstream pathway branches integrated into ECKh-422. Strain 12: host strain ECKh-432, deletion of endogenous adhE, ldhA, pflB, deletion of endogenous lpdA and chromosomal insertion of Klebsiella pneumoniae lpdA with a Glu354Lys mutation at the lpdA locus, deletion of endogenous mdh and arcA, chromosomal replacement of gltA with gltA Arg163Leu mutant, chromosomal insertion at the fimD locus of E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd, chromosomal insertion at the fimD locus of M. bovis sucA, C. kluyveri 4hbd; plasmid expression of C. acetobutylicum buk1, C. acetobutylicum ptb, C. beijerinckii Ald.
[0538] Strain 13: host strain ECKh-439, deletion of endogenous adhE, ldhA, pflB, deletion of endogenous lpdA and chromosomal insertion of Klebsiella pneumoniae lpdA with a Glu354Lys mutation at the lpdA locus, deletion of endogenous mdh and arcA, chromosomal replacement of gltA with gltA Arg163Leu mutant, deletion of endogenous ackA, chromosomal insertion at the fimD locus of E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd, chromosomal insertion at the fimD locus of M. bovis sucA, C. kluyveri 4hbd; plasmid expression of P. gingivalis Cat2, C. beijerinckii Ald; strain has acetate kinase deletion in strain ECKh-432. Strain 14: host strain ECKh-453, deletion of endogenous adhE, ldhA, pflB, deletion of endogenous lpdA and chromosomal insertion of Klebsiella pneumoniae lpdA with a Glu354Lys mutation at the lpdA locus, deletion of endogenous mdh and arcA, chromosomal replacement of gltA with gltA Arg163Leu mutant, deletion of endogenous ackA, deletion of endogenous ppc and insertion of Haemophilus influenza ppck at the ppc locus, chromosomal insertion at the fimD locus of E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd, chromosomal insertion at the fimD locus of M. bovis sucA, C. kluyveri 4hbd; plasmid expression of P. gingivalis Cat2, C. beijerinckii Ald; strain has acetate kinase deletion and PPC/PEPCK replacement in strain ECKh-432.
[0539] Strain 15: host strain ECKh-456, deletion of endogenous adhE, ldhA, pflB, deletion of endogenous lpdA and chromosomal insertion of Klebsiella pneumoniae lpdA with a Glu354Lys mutation at the lpdA locus, deletion of endogenous mdh and arcA, chromosomal replacement of gltA with gltA Arg163Leu mutant, chromosomal insertion at the fimD locus of E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd, chromosomal insertion at the fimD locus of M. bovis sucA, C. kluyveri 4hbd, replacement of lpdA promoter with fnr binding site, pflB-p6 promoter and RBS of pflB; plasmid expression of P. gingivalis Cat2, C. beijerinckii Ald; strain has replacement of lpdA promoter with anaerobic promoter in strain ECKh-432. Strain 16: host strain ECKh-455, deletion of endogenous adhE, ldhA, pflB, deletion of endogenous lpdA and chromosomal insertion of Klebsiella pneumoniae lpdA with a Glu354Lys mutation at the lpdA locus, deletion of endogenous mdh and arcA, chromosomal replacement of gltA with gltA Arg163Leu mutant, chromosomal insertion at the fimD locus of E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd, chromosomal insertion at the fimD locus of M. bovis sucA, C. kluyveri 4hbdI, replacement of pdhR and aceEF promoter with fnr binding site, pflB-p6 promoter and RBS of pflB; plasmid expression of P. gingivalis Cat2, C. beijerinckii Ald; strain has replacement of pdhR and aceEF promoter with anaerobic promoter in ECKh-432.
[0540] Strain 17: host strain ECKh-459, deletion of endogenous adhE, ldhA, pflB, deletion of endogenous lpdA and chromosomal insertion of Klebsiella pneumoniae lpdA with a Glu354Lys mutation at the lpdA locus, deletion of endogenous mdh and arcA, chromosomal replacement of gltA with gltA Arg163Leu mutant, chromosomal insertion at the fimD locus of E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd, chromosomal insertion at the fimD locus of M. bovis sucA, C. kluyveri 4hbd, chromosomal insertion at the fimD locus of C. acetobutylicum buk1, C. acetobutylicum ptb; plasmid expression of C. beijerinckii Ald; strain has integration of BK/PTB into strain ECKh-432. Strain 18: host strain ECKh-459, deletion of endogenous adhE, ldhA, pflB, deletion of endogenous lpdA and chromosomal insertion of Klebsiella pneumoniae lpdA with a Glu354Lys mutation at the lpdA locus, deletion of endogenous mdh and arcA, chromosomal replacement of gltA with gltA Arg163Leu mutant, chromosomal insertion at the fimD locus of E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd, chromosomal insertion at the fimD locus of M. bovis sucA, C. kluyveri 4hbd, chromosomal insertion at the fimD locus of C. acetobutylicum buk1, C. acetobutylicum ptb; plasmid expression of C. beijerinckii Ald, G. thermoglucosidasius adh1.
[0541] Strain 19: host strain ECKh-463, deletion of endogenous adhE, ldhA, pflB, deletion of endogenous lpdA and chromosomal insertion of Klebsiella pneumoniae lpdA with a Glu354Lys mutation at the lpdA locus, deletion of endogenous mdh and arcA, chromosomal replacement of gltA with gltA Arg163Leu mutant, chromosomal insertion at the fimD locus of E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd, chromosomal insertion at the fimD locus of M. bovis sucA, C. kluyveri 4hbd, insertion at the rrnC locus of non-PTS sucrose operon genes sucrose permease (cscB), D-fructokinase (cscK), sucrose hydrolase (cscA), and a LacI-related sucrose-specific repressor (cscR); plasmid expression of P. gingivalis Cat2, C. beijerinckii Ald; strain has non-PTS sucrose genes inserted into strain ECKh-432. Strain 20: host strain ECKh-463 deletion of endogenous adhE, ldhA, pflB, deletion of endogenous lpdA and chromosomal insertion of Klebsiella pneumoniae lpdA with a Glu354Lys mutation at the lpdA locus, deletion of endogenous mdh and arcA, chromosomal replacement of gltA with gltA Arg163Leu mutant, chromosomal insertion at the fimD locus of E. coli sucCD, P. gingivalis sucD, P. gingivalis 4hbd, chromosomal insertion at the fimD locus of M. bovis sucA, C. kluyveri 4hbd, insertion at the rrnC locus of non-PTS sucrose operon; plasmid expression of C. acetobutylicum buk1, C. acetobutylicum ptb, C. beijerinckii Ald.
[0542] In addition to the BDO producing strains disclosed herein, including those disclosed in Table 28, it is understood that additional modifications can be incorporated that further increase production of BDO and/or decrease undesirable byproducts. For example, a BDO producing strain, or a strain of Table 28, can incorporate additional knockouts to further increase the production of BDO or decrease an undesirable byproduct. Exemplary knockouts have been described previously (see U.S. publication 2009/0047719). Such knockout strains include, but are not limited to, ADHEr, NADH6; ADHEr, PPCK; ADHEr, SUCD4; ADHEr, ATPS4r; ADHEr, FUM; ADHEr, MDH; ADHEr, PFLi, PPCK; ADHEr, PFLi, SUCD4; ADHEr, ACKr, NADH6; ADHEr, NADH6, PFLi; ADHEr, ASPT, MDH; ADHEr, NADH6, PPCK; ADHEr, PPCK, THD2; ADHEr, ATPS4r, PPCK; ADHEr, MDH, THD2; ADHEr, FUM, PFLi; ADHEr, PPCK, SUCD4; ADHEr, GLCpts, PPCK; ADHEr, GLUDy, MDH; ADHEr, GLUDy, PPCK; ADHEr, FUM, PPCK; ADHEr, MDH, PPCK; ADHEr, FUM, GLUDy; ADHEr, FUM, HEX1; ADHEr, HEX1, PFLi; ADHEr, HEX1, THD2; ADHEr, FRD2, LDH_D, MDH; ADHEr, FRD2, LDH_D, ME2; ADHEr, MDH, PGL, THD2; ADHEr, G6PDHy, MDH, THD2; ADHEr, PFLi, PPCK, THD2; ADHEr, ACKr, AKGD, ATPS4r; ADHEr, GLCpts, PFLi, PPCK; ADHEr, ACKr, ATPS4r, SUCOAS; ADHEr, GLUDy, PFLi, PPCK; ADHEr, ME2, PFLi, SUCD4; ADHEr, GLUDy, PFLi, SUCD4; ADHEr, ATPS4r, LDH_D, SUCD4; ADHEr, FUM, HEX1, PFLi; ADHEr, MDH, NADH6, THD2; ADHEr, ATPS4r, MDH, NADH6; ADHEr, ATPS4r, FUM, NADH6; ADHEr, ASPT, MDH, NADH6; ADHEr, ASPT, MDH, THD2; ADHEr, ATPS4r, GLCpts, SUCD4; ADHEr, ATPS4r, GLUDy, MDH; ADHEr, ATPS4r, MDH, PPCK; ADHEr, ATPS4r, FUM, PPCK; ADHEr, ASPT, GLCpts, MDH; ADHEr, ASPT, GLUDy, MDH; ADHEr, ME2, SUCD4, THD2; ADHEr, FUM, PPCK, THD2; ADHEr, MDH, PPCK, THD2; ADHEr, GLUDy, MDH, THD2; ADHEr, HEX1, PFLi, THD2; ADHEr, ATPS4r, G6PDHy, MDH; ADHEr, ATPS4r, MDH, PGL; ADHEr, ACKr, FRD2, LDH_D; ADHEr, ACKr, LDH_D, SUCD4; ADHEr, ATPS4r, FUM, GLUDy; ADHEr, ATPS4r, FUM, HEX1; ADHEr, ATPS4r, MDH, THD2; ADHEr, ATPS4r, FRD2, LDH_D; ADHEr, ATPS4r, MDH, PGDH; ADHEr, GLCpts, PPCK, THD2; ADHEr, GLUDy, PPCK, THD2; ADHEr, FUM, HEX1, THD2; ADHEr, ATPS4r, ME2, THD2; ADHEr, FUM, ME2, THD2; ADHEr, GLCpts, GLUDy, PPCK; ADHEr, ME2, PGL, THD2; ADHEr, G6PDHy, ME2, THD2; ADHEr, ATPS4r, FRD2, LDH_D, ME2; ADHEr, ATPS4r, FRD2, LDH_D, MDH; ADHEr, ASPT, LDH_D, MDH, PFLi; ADHEr, ATPS4r, GLCpts, NADH6, PFLi; ADHEr, ATPS4r, MDH, NADH6, PGL; ADHEr, ATPS4r, G6PDHy, MDH, NADH6; ADHEr, ACKr, FUM, GLUDy, LDH_D; ADHEr, ACKr, GLUDy, LDH_D, SUCD4; ADHEr, ATPS4r, G6PDHy, MDH, THD2; ADHEr, ATPS4r, MDH, PGL, THD2; ADHEr, ASPT, G6PDHy, MDH, PYK; ADHEr, ASPT, MDH, PGL, PYK; ADHEr, ASPT, LDH_D, MDH, SUCOAS; ADHEr, ASPT, FUM, LDH_D, MDH; ADHEr, ASPT, LDH_D, MALS, MDH; ADHEr, ASPT, ICL, LDH_D, MDH; ADHEr, FRD2, GLUDy, LDH_D, PPCK; ADHEr, FRD2, LDH_D, PPCK, THD2; ADHEr, ACKr, ATPS4r, LDH_D, SUCD4; ADHEr, ACKr, ACS, PPC, PPCK; ADHEr, GLUDy, LDH_D, PPC, PPCK; ADHEr, LDH_D, PPC, PPCK, THD2; ADHEr, ASPT, ATPS4r, GLCpts, MDH; ADHEr, G6PDHy, MDH, NADH6, THD2; ADHEr, MDH, NADH6, PGL, THD2; ADHEr, ATPS4r, G6PDHy, GLCpts, MDH; ADHEr, ATPS4r, GLCpts, MDH, PGL; ADHEr, ACKr, LDH_D, MDH, SUCD4.
[0543] Table 29 shows the reactions of corresponding genes to be knocked out of a host organism such as E. coli. The corresponding metabolite corresponding to abbreviations in Table 29 are shown in Table 30.
TABLE-US-00081 TABLE 29 Corresponding genes to be knocked out to prevent a particular reaction from occurring in E. coli. Reaction Reaction Genes Encoding the Enzyme(s) Abbreviation Stoichiometry* Catalyzing Each Reaction& ACKr [c]: ac + atp <==> actp + adp (b3115 or b2296 or b1849) ACS [c]: ac + atp + coa --> accoa + amp + ppi b4069 ACt6 ac[p] + h[p] <==> ac[c] + h[c] Non-gene associated ADHEr [c]: etoh + nad <==> acald + h + nadh (b0356 or b1478 or b1241) [c]: acald + coa + nad <==> accoa + h + nadh (b1241 or b0351) AKGD [c]: akg + coa + nad --> co2 + nadh + succoa (b0116 and b0726 and b0727) ASNS2 [c]: asp-L + atp + nh4 --> amp + asn-L + h + ppi b3744 ASPT [c]: asp-L --> fum + nh4 b4139 ATPS4r adp[c] + (4) h[p] + pi[c] <==> atp[c] + (3) h[c] + h2o[c] (((b3736 and b3737 and b3738) and (b3731 and b3732 and b3733 and b3734 and b3735)) or ((b3736 and b3737 and b3738) and (b3731 and b3732 and b3733 and b3734 and b3735) and b3739)) CBMK2 [c]: atp + co2 + nh4 <==> adp + cbp + (2) h (b0521 or b0323 or b2874) EDA [c]: 2ddg6p --> g3p + pyr b1850 ENO [c]: 2pg <==> h2o + pep b2779 FBA [c]: fdp <==> dhap + g3p (b2097 or b2925 or b1773) FBP [c]: fdp + h2o --> f6p + pi (b4232 or b3925) FDH2 for[p] + (2) h[c] + q8[c] --> co2[c] + h[p] + q8h2[c] ((b3892 and b3893 and b3894) for[p] + (2) h[c] + mqn8[c] --> co2[c] + h[p] + mql8[c] or (b1474 and b1475 and b1476)) FRD2 [c]: fum + mql8 --> mqn8 + succ (b4151 and b4152 and b4153 [c]: 2dmmql8 + fum --> 2dmmq8 + succ and b4154) FTHFD [c]: 10fthf + h2o --> for + h + thf b1232 FUM [c]: fum + h2o <==> mal-L (b1612 or b4122 or b1611) G5SD [c]: glu5p + h + nadph --> glu5sa + nadp + pi b0243 G6PDHy [c]: g6p + nadp <==> 6pgl + h + nadph b1852 GLCpts glc-D[p] + pep[c] --> g6p[c] + pyr[c] ((b2417 and b1101 and b2415 and b2416) or (b1817 and b1818 and b1819 and b2415 and b2416) or (b2417 and b1621 and b2415 and b2416)) GLU5K [c]: atp + glu-L --> adp + glu5p b0242 GLUDy [c]: glu-L + h2o + nadp <==> akg + h + nadph + nh4 b1761 GLYCL [c]: gly + nad + thf --> co2 + mlthf + nadh + nh4 (b2904 and b2903 and b2905 and b0116) HEX1 [c]: atp + glc-D --> adp + g6p + h b2388 ICL [c]: icit --> glx + succ b4015 LDH_D [c]: lac-D + nad <==> h + nadh + pyr (b2133 or b1380) MALS [c]: accoa + glx + h2o --> coa + h + mal-L (b4014 or b2976) MDH [c]: mal-L + nad <==> h + nadh + oaa b3236 ME2 [c]: mal-L + nadp --> co2 + nadph + pyr b2463 MTHFC [c]: h2o + methf <==> 10fthf + h b0529 NADH12 [c]: h + mqn8 + nadh --> mql8 + nad b1109 [c]: h + nadh + q8 --> nad + q8h2 [c]: 2dmmq8 + h + nadh --> 2dmmql8 + nad NADH6 (4) h[c] + nadh[c] + q8[c] --> (3) h[p] + nad[c] + q8h2[c] (b2276 and b2277 and b2278 (4) h[c] + mqn8[c] + nadh[c] --> (3) h[p] + mql8[c] + and b2279 and b2280 and nad[c] b2281 and b2282 and b2283 2dmmq8[c] + (4) h[c] + nadh[c] --> 2dmmql8[c] + (3) and b2284 and b2285 and h[p] + nad[c] b2286 and b2287 and b2288) PFK [c]: atp + f6p --> adp + fdp + h (b3916 or b1723) PFLi [c]: coa + pyr --> accoa + for (((b0902 and b0903) and b2579) or (b0902 and b0903) or (b0902 and b3114) or (b3951 and b3952)) PGDH [c]: 6pgc + nadp --> co2 + nadph + ru5p-D b2029 PGI [c]: g6p <==> f6p b4025 PGL [c]: 6pgl + h2o --> 6pgc + h b0767 PGM [c]: 2pg <==> 3pg (b3612 or b4395 or b0755) PPC [c]: co2 + h2o + pep --> h + oaa + pi b3956 PPCK [c]: atp + oaa --> adp + co2 + pep b3403 PRO1z [c]: fad + pro-L --> 1pyr5c + fadh2 + h b1014 PYK [c]: adp + h + pep --> atp + pyr b1854 or b1676) PYRt2 h[p] + pyr[p] <==> h[c] + pyr[c] Non-gene associated RPE [c]: ru5p-D <==> xu5p-D (b4301 or b3386) SO4t2 so4[e] <==> so4[p] (b0241 or b0929 or b1377 or b2215) SUCD4 [c]: q8 + succ --> fum + q8h2 (b0721 and b0722 and b0723 and b0724) SUCOAS [c]: atp + coa + succ <==> adp + pi + succoa (b0728 and b0729) SULabc atp[c] + h2o[c] + so4[p] --> adp[c] + h[c] + pi[c] + ((b2422 and b2425 and so4[c] b2424 and b2423) or (b0763 and b0764 and b0765) or (b2422 and b2424 and b2423 and b3917)) TAL [c]: g3p + s7p <==> e4p + f6p (b2464 or b0008) THD2 (2) h[p] + nadh[c] + nadp[c] --> (2) h[c] + nad[c] + (b1602 and b1603) nadph[c] THD5 [c]: nad + nadph --> nadh + nadp (b3962 or (b1602 and b1603)) TPI [c]: dhap <==> g3p b3919
TABLE-US-00082 TABLE 30 Metabolite names corresponding to abbreviations used in Table 29. Metabolite Metabolite Abbreviation Name 10fthf 10-Formyltetrahydrofolate 1pyr5c 1-Pyrroline-5-carboxylate 2ddg6p 2-Dehydro-3-deoxy-D-gluconate 6-phosphate 2dmmq8 2-Demethylmenaquinone 8 2dmmql8 2-Demethylmenaquinol 8 2pg D-Glycerate 2-phosphate 3pg 3-Phospho-D-glycerate 6pgc 6-Phospho-D-gluconate 6pgl 6-phospho-D-glucono-1,5-lactone ac Acetate acald Acetaldehyde accoa Acetyl-CoA actp Acetyl phosphate adp ADP akg 2-Oxoglutarate amp AMP asn-L L-Asparagine asp-L L-Aspartate atp ATP cbp Carbamoyl phosphate co2 CO2 coa Coenzyme A dhap Dihydroxyacetone phosphate e4p D-Erythrose 4-phosphate etoh Ethanol f6p D-Fructose 6-phosphate fad Flavin adenine dinucleotide oxidized fadh2 Flavin adenine dinucleotide reduced fdp D-Fructose 1,6-bisphosphate for Formate fum Fumarate g3p Glyceraldehyde 3-phosphate g6p D-Glucose 6-phosphate glc-D D-Glucose glu5p L-Glutamate 5-phosphate glu5sa L-Glutamate 5-semialdehyde glu-L L-Glutamate glx Glyoxylate gly Glycine h H+ h2o H2O icit Isocitrate lac-D D-Lactate mal-L L-Malate methf 5,10-Methenyltetrahydrofolate mlthf 5,10-Methylenetetrahydrofolate mql8 Menaquinol 8 mqn8 Menaquinone 8 nad Nicotinamide adenine dinucleotide nadh Nicotinamide adenine dinucleotide - reduced nadp Nicotinamide adenine dinucleotide phosphate nadph Nicotinamide adenine dinucleotide phosphate - reduced nh4 Ammonium oaa Oxaloacetate pep Phosphoenolpyruvate pi Phosphate ppi Diphosphate pro-L L-Proline pyr Pyruvate q8 Ubiquinone-8 q8h2 Ubiquinol-8 ru5p-D D-Ribulose 5-phosphate s7p Sedoheptulose 7-phosphate so4 Sulfate succ Succinate succoa Succinyl-CoA thf 5,6,7,8-Tetrahydrofolate xu5p-D D-Xylulose 5-phosphate
Example XX
Exemplary Pathways for Producing BDO
[0544] This example describes exemplary pathways to produce 4-hydroxybutanal (4-HBal) and/or BDO using a carboxylic acid reductase as a BDO pathway enzyme.
[0545] An exemplary pathway for production of BDO includes use of an NAD+ or NADP+ aryl-aldehyde dehydrogenase (E.C.: 1.2.1.29 and 1.2.1.30) to convert 4-hydroxybutyrate to 4-hydroxybutanal and an alcohol dehydrogenase to convert 4-hydroxybutanal to 1,4-butanediol. 4-Hydroxybutyrate can be derived from the tricarboxylic acid cycle intermediates succinyl-CoA and/or alpha-ketoglutarate as shown in FIG. 58.
[0546] Aryl-Aldehyde Dehydrogenase (or Carboxylic Acid Reductase).
[0547] An aryl-aldehyde dehydrogenase, or equivalently a carboxylic acid reductase, can be found in Nocardia iowensis. Carboxylic acid reductase catalyzes the magnesium, ATP and NADPH-dependent reduction of carboxylic acids to their corresponding aldehydes (Venkitasubramanian et al., J. Biol. Chem. 282:478-485 (2007)) and is capable of catalyzing the conversion of 4-hydroxybutyrate to 4-hydroxybutanal. This enzyme, encoded by car, was cloned and functionally expressed in E. coli (Venkitasubramanian et al., J. Biol. Chem. 282:478-485 (2007)). Expression of the npt gene product improved activity of the enzyme via post-transcriptional modification. The npt gene encodes a specific phosphopantetheine transferase (PPTase) that converts the inactive apo-enzyme to the active holo-enzyme. The natural substrate of this enzyme is vanillic acid, and the enzyme exhibits broad acceptance of aromatic and aliphatic substrates (Venkitasubramanian et al., in Biocatalysis in the Pharmaceutical and Biotechnology Industires, ed. R. N. Patel, Chapter 15, pp. 425-440, CRC Press LLC, Boca Raton, Fla. (2006)).
TABLE-US-00083 Gene GenBank name GI No. Accession No. Organism car 40796035 AAR91681.1 Nocardia iowensis (sp. NRRL 5646) npt 114848891 ABI83656.1 Nocardia iowensis (sp. NRRL 5646)
[0548] Additional car and npt genes can be identified based on sequence homology.
TABLE-US-00084 Gene GenBank name GI No. Accession No. Organism fadD9 121638475 YP_978699.1 Mycobacterium bovis BCG BCG_2812c 121638674 YP_978898.1 Mycobacterium bovis BCG nfa20150 54023983 YP_118225.1 Nocardia farcinica IFM 10152 nfa40540 54026024 YP_120266.1 Nocardia farcinica IFM 10152 SGR_6790 182440583 YP_001828302.1 Streptomyces griseus subsp. griseus NBRC 13350 SGR_665 182434458 YP_001822177.1 Streptomyces griseus subsp. griseus NBRC 13350 MSMEG_2956 YP_887275.1 YP_887275.1 Mycobacterium smegmatis MC2 155 MSMEG_5739 YP_889972.1 118469671 Mycobacterium smegmatis MC2 155 MSMEG_2648 YP_886985.1 118471293 Mycobacterium smegmatis MC2 155 MAP1040c NP_959974.1 41407138 Mycobacterium avium subsp. paratuberculosis K-10 MAP2899c NP_961833.1 41408997 Mycobacterium avium subsp. paratuberculosis K-10 MMAR_2117 YP_001850422.1 183982131 Mycobacterium marinum M MMAR_2936 YP_001851230.1 183982939 Mycobacterium marinum M MMAR_1916 YP_001850220.1 183981929 Mycobacterium marinum M TpauDRAFT_33060 ZP_04027864.1 227980601 Tsukamurella paurometabola DSM 20162 TpauDRAFT_20920 ZP_04026660.1 ZP_04026660.1 Tsukamurella paurometabola DSM 20162 CPCC7001_1320 ZP_05045132.1 254431429 Cyanobium PCC7001 DDBDRAFT_0187729 XP_636931.1 66806417 Dictyostelium discoideum AX4
[0549] An additional enzyme candidate found in Streptomyces griseus is encoded by the griC and griD genes. This enzyme is believed to convert 3-amino-4-hydroxybenzoic acid to 3-amino-4-hydroxybenzaldehyde as deletion of either griC or griD led to accumulation of extracellular 3-acetylamino-4-hydroxybenzoic acid, a shunt product of 3-amino-4-hydroxybenzoic acid metabolism (Suzuki, et al., J. Antibiot. 60(6):380-387 (2007)). Co-expression of griC and griD with SGR_665, an enzyme similar in sequence to the Nocardia iowensis npt, can be beneficial.
TABLE-US-00085 Gene GenBank name GI No. Accession No. Organism griC 182438036 YP_001825755.1 Streptomyces griseus subsp. griseus NBRC 13350 griD 182438037 YP_001825756.1 Streptomyces griseus subsp. griseus NBRC 13350
[0550] An enzyme with similar characteristics, alpha-aminoadipate reductase (AAR, EC 1.2.1.31), participates in lysine biosynthesis pathways in some fungal species. This enzyme naturally reduces alpha-aminoadipate to alpha-aminoadipate semialdehyde. The carboxyl group is first activated through the ATP-dependent formation of an adenylate that is then reduced by NAD(P)H to yield the aldehyde and AMP. Like CAR, this enzyme utilizes magnesium and requires activation by a PPTase. Enzyme candidates for AAR and its corresponding PPTase are found in Saccharomyces cerevisiae (Morris et al., Gene 98:141-145 (1991)), Candida albicans (Guo et al., Mol. Genet. Genomics 269:271-279 (2003)), and Schizosaccharomyces pombe (Ford et al., Curr. Genet. 28:131-137 (1995)). The AAR from S. pombe exhibited significant activity when expressed in E. coli (Guo et al., Yeast 21:1279-1288 (2004)). The AAR from Penicillium chrysogenum accepts S-carboxymethyl-L-cysteine as an alternate substrate, but did not react with adipate, L-glutamate or diaminopimelate (Hijarrubia et al., J. Biol. Chem. 278:8250-8256 (2003)). The gene encoding the P. chrysogenum PPTase has not been identified to date.
TABLE-US-00086 Gene GenBank name GI No. Accession No. Organism LYS2 171867 AAA34747.1 Saccharomyces cerevisiae LYS5 1708896 P50113.1 Saccharomyces cerevisiae LYS2 2853226 AAC02241.1 Candida albicans LYS5 28136195 AAO26020.1 Candida albicans Lys1p 13124791 P40976.3 Schizosaccharomyces pombe Lys7p 1723561 Q10474.1 Schizosaccharomyces pombe Lys2 3282044 CAA74300.1 Penicillium chrysogenum
[0551] There are several advantages of using carboxylic acid reductase for BDO production. There are at least two advantages of forming 4-hydroxybutanal from 4-hydroxybutyrate via a carboxylic acid reductase compared to forming 4-hydroxybutanal from an activated version of 4-hydroxybutyrate (for example, 4-hydroxybutyryl-CoA, 4-hydroxybutyryl-Pi) via an acyl-CoA or acyl-phosphate reductase. First, the formation of gamma-butyrolactone (GBL) as a byproduct is greatly reduced. It is believed that the activated versions of 4-hydroxybutyrate cyclize to GBL more readily than unactivated 4-hydroxybutyrate. The use of carboxylic acid reductase eliminates the need to pass through a free activated 4-hydroxybutyrate intermediate, thus reducing the formation of GBL as a byproduct accompanying BDO production. Second, the formation of ethanol as a byproduct is greatly reduced. Ethanol is often formed in varying amounts when an aldehyde- or an alcohol-forming 4-hydroxybutyryl-CoA reductase is used to convert 4-hydroxybutyryl-CoA to 4-hydroxybutanal or 1,4-butanediol, respectively. This is because most, if not all, aldehyde- or alcohol-forming 4-hydroxybutyryl-CoA reductases can accept acetyl-CoA as a substrate in addition to 4-hydroxybutyryl-CoA. Aldehyde-forming enzymes, for example, often catalyze the conversion of acetyl-CoA to acetaldehyde, which is subsequently reduced to ethanol by native or non-native alcohol dehydrogenases. Alcohol-forming 4-hydroxybutyryl-CoA reductases that accept acetyl-CoA as a substrate will convert acetyl-CoA directly to ethanol. It appears that carboxylic acid reductase enzymes have far less activity on acetyl-CoA than aldehyde- or alcohol-forming acyl-CoA reductase enzymes, and thus their application for BDO production results in minimal ethanol byproduct formation (see below).
Example XXI
Biosynthesis of 1,4-Butanediol Using a Carboxylic Acid Reductase Enzyme
[0552] This example describes the generation of a microbial organism that produces 1,4-butanediol using a carboxylic acid reductase enzyme.
[0553] Escherichia coli is used as a target organism to engineer the pathway for 1,4-butanediol synthesis described in FIG. 58. E. coli provides a good host for generating a non-naturally occurring microorganism capable of producing 1,4-butanediol. E. coli is amenable to genetic manipulation and is known to be capable of producing various products, like ethanol, acetic acid, formic acid, lactic acid, and succinic acid, effectively under various oxygenation conditions.
[0554] Integration of 4-Hydroxybutyrate Pathway Genes into Chromosome: Construction of ECKh-432.
[0555] The carboxylic acid reductase enzyme was expressed in a strain of E. coli designated ECKh-432 whose construction is described in Example XVII. This strain contained the components of the BDO pathway, leading to 4HB, integrated into the chromosome of E. coli at the fimD locus.
[0556] As described in Example XVII, the succinate branch of the upstream pathway was integrated into the E. coli chromosome using the .lamda. red homologeous recombination method (Datsenko and Wanner, Proc. Natl. Acad. Sci. USA 97:6640-6645 (2000)). A polycistronic DNA fragment containing a promoter, the sucCD gene of Escherichia coli encoding succinyl-CoA ligase, the sucD gene of Porphyromonas gingivalis encoding succinyl-CoA reductase (aldehyde forming) (step A of FIG. 58), the 4hbd gene of Porphyromonas gingivalis encoding 4-hydroxybutyrate dehydrogenase (step C of FIG. 58), and a terminator sequence was inserted into the AflIII site of the pKD3 plasmid.
[0557] As described in Example XVII, the alpha-ketoglutarate branch of the upstream pathway was integrated into the chromosome by homologeous recombination. The plasmid used in this modification was pRE118-V2 (pRE118 (ATCC87693) deleted of the oriT and IS sequences), which contains a kanamycin-resistant gene, a gene encoding the levansucrase (sacB) and a R6K conditional replication ori. The integration plasmid also contained a polycistronic sequence with a promoter, the sucA gene from Mycobacterium bovis encoding alpha-ketoglutarate decarboxylase (step B of FIG. 58), the Clostridium kluyveri 4hbd gene encoding 4-hydroxybutyrate dehydrogenase (step C of FIG. 58), and a terminator being inserted between two 1.5-kb DNA fragments that are homologous to the flanking regions of the target insertion site. The resulting plasmid was used to transform E. coli strain. The integration candidate was selected on plates containing kanamycin. The correct integration site was verified by PCR. To resolve the antibiotic marker from the chromosome, the cells were selected for growth on medium containing sucrose. The final strain was verified by PCR and DNA sequencing.
[0558] The recipient E. coli strain was ECKh-422 (.DELTA.adhE .DELTA.ldhA .DELTA.pflB .DELTA.lpdA::K.p.lpdA322 .DELTA.mdh .DELTA.arcA gltAR163L) whose construction is described in Example XV. ECKh-422 contains a mutation gltAR163L leading to NADH-insensitivity of citrate synthase encoded by gltA. It further contains an NADH-insensitive version of the lpdA gene from Klebsiella pneumonia integrated into the chromosome as described below.
[0559] Replacement of the native lpdA was replaced with a NADH-insensitive lpdA from Klebsiella pneumonia, as described in Example XIV. The resulting vector was designated pRE118-V2 (see FIG. 34).]
[0560] Cloning and Expression of Carboxylic Acid Reductase and PPTase.
[0561] To generate an E. coli strain engineered to produce 1,4-butanediol, nucleic acids encoding a carboxylic acid reductase and phosphopantetheine transferase are expressed in E. coli using well known molecular biology techniques (see, for example, Sambrook, supra, 2001; Ausubel supra, 1999). In particular, the car (AAR91681.1) and npt (ABI83656.1) genes were cloned into the pZS*13 vector (Expressys, Ruelzheim, Germany) under the PA1/lacO promoter. The car gene (GNM_720) was cloned by PCR from Nocardia genomic DNA. Its nucleic acid and protein sequences are shown in FIGS. 59A and 59B, respectively.
[0562] A codon-optimized version of the npt gene (GNM_721) was synthesized by GeneArt (Regensburg, Germany). Its nucleic acid and protein sequences are shown in FIGS. 60A and 60B, respectively. The resulting vector from cloning GNM_720 and GNM_721 into pZS*13 is shown in FIG. 61.
[0563] The plasmid was transformed into ECKh-432 to express the proteins and enzymes required for 1,4-butanediol production. Alternate versions of the plasmid containing only GNM_720 and only GNM_721 were also constructed.
[0564] Demonstration of 1,4-BDO Production Using Carboxylic Acid Reductase.
[0565] Functional expression of the 1,4-butanediol pathway was demonstrated using E. coli whole-cell culture. A single colony of E. coli ECKh-432 transformed with the pZS*13 plasmid containing both GNM_720 and GNM_721 was inoculated into 5 mL of LB medium containing appropriate antibiotics. Similarly, single colonies of E. coli ECKh-432 transformed with the pZS*13 plasmids containing either GNM_720 or GNM_721 were inoculated into additional 5 mL aliquots of LB medium containing appropriate antibiotics. Ten mL micro-aerobic cultures were started by inoculating fresh minimal in vivo conversion medium (see below) containing the appropriate antibiotics with 1% of the first cultures.
[0566] Recipe of the minimal in vivo conversion medium (for 1000 mL) is as follows:
TABLE-US-00087 final concentration 1M MOPS/KOH buffer 40 mM Glucose (40%) 1% 10XM9 salts solution 1X MgSO4 (1M) 1 mM trace minerals (x1000) 1X 1M NaHCO3 10 mM
Microaerobic conditions were established by initially flushing capped anaerobic bottles with nitrogen for 5 minutes, then piercing the septum with an 18G needle following inoculation. The needle was kept in the bottle during growth to allow a small amount of air to enter the bottles. Protein expression was induced with 0.2 mM IPTG when the culture reached mid-log growth phase. This is considered: time=0 hr. The culture supernatants were analyzed for BDO, 4HB, and other by-products as described above and in WO2008115840 (see Table 31).
TABLE-US-00088 TABLE 31 Production of BDO, 4-HB and other products in various strains. mM Strain pZS*13S OD600 OD600 PA SA LA 4HB BDO GBL ETOH.sub.Enz ECKh-432 720 0.420 2.221 6.36 0.00 0.10 7.71 3.03 0.07 >LLOQ ECKh-432 721 0.323 2.574 1.69 0.00 0.00 12.60 0.00 0.00 >LLOQ ECKh-432 720/721 0.378 2.469 1.70 0.00 0.01 4.23 9.16 0.24 1.52 PA = pyruvate, SA = succinate, LA = lactate, 4HB = 4-hydroxybutyrate, BDO = 1,4-butanediol, GBL = gamma-butyrolactone, Etoh = ethanol, LLOQ = lower limit of quantification
[0567] These results demonstrate that the carboxylic acid reductase gene, GNM_720, is required for BDO formation in ECKh-432 and its effectiveness is increased when co-expressed with the PPTase, GNM_721. GBL and ethanol were produced in far smaller quantities than BDO in the strains expressing GNM_720 by itself or in combination with GNM_721.
[0568] Additional Pathways to BDO Employing Carboxylic Acid Reductase.
[0569] It is expected that carboxylic acid reductase can function as a component of many pathways to 1,4-butanediol from the TCA cycle metabolites: succinate, succinyl-CoA, and alpha-ketoglutarate. Several of these pathways are disclosed in FIG. 62. All routes can lead to theoretical BDO yields greater than or equal to 1 mol/mol assuming glucose as the carbon source. Similar high theoretical yields can be obtained from additional substrates including sucrose, xylose, arabinose, synthesis gas, among many others. It is expected that the expression of carboxylic acid reductase alone or in combination with PPTase (that is, to catalyze steps F and D of FIG. 62) is sufficient for 1,4-butanediol production from succinate provided that sufficient endogenous alcohol dehydrogenase activity is present to catalyze steps C and E of FIG. 62. Candidate enzymes for steps A through Z of FIG. 62 are described in section XXIII
Example XXII
Pathways to Putrescine that Employ Carboxylic Acid Reductase
[0570] This example describes exemplary putrescine pathways utilizing carboxylic acid reductase.
[0571] Putrescine, also known as 1,4-diaminobutane or butanediamine, is an organic chemical compound of the formula NH.sub.2(CH.sub.2).sub.4NH.sub.2. It can be reacted with adipic acid to yield the polyamide Nylon-4,6, which is marketed by DSM (Heerlen, Netherlands) under the trade name Stanyl.TM.. Putrescine is naturally produced, for example, by the natural breakdown of amino acids in living and dead organisms. E. coli has been engineered to produce putrescine by overexpressing the native ornithine biosynthetic machinery as well as an ornithine decarboxylase (Qian, et al., Biotechnol. Bioeng. 104(4):651-662 (2009)).
[0572] FIG. 63 describes a number of additional biosynthetic pathways leading to the production of putrescine from succinate, succinyl-CoA, or alpha-ketoglutarate and employing a carboxylic acid reductase. Note that none of these pathways require formation of an activated version of 4-aminobutyrate such as 4-aminobutyryl-CoA, which can be reduced by an acyl-CoA reductase to 4-aminobutanal but also can readily cyclize to its lactam, 2-pyrrolidinone (Ohsugi, et al., J. Biol. Chem. 256:7642-7651 (1981)). All routes can lead to theoretical putrescine yields greater than or equal to 1 mol/mol assuming glucose as the carbon source. Similar high theoretical yields can be obtained from additional substrates including sucrose, xylose, arabinose, synthesis gas, among many others. Candidate enzymes for steps A through U of FIG. 63 are described in Example XXIII
Example XXIII
Exemplary Enzymes for Production of C4 Compounds
[0573] This example describes exemplary enzymes for production of C4 compounds such as 1,4-butanediol, 4-hydroxybutanal and putrescine.
[0574] Enzyme Classes.
[0575] All transformations depicted in FIGS. 58, 62 and 63 fall into the general categories of transformations shown in Table 32. This example describes a number of biochemically characterized genes in each category. Specifically listed are genes that can be applied to catalyze the appropriate transformations in FIGS. 58, 62 and 63 when cloned and expressed. The first three digits of each label correspond to the first three Enzyme Commission number digits which denote the general type of transformation independent of substrate specificity.
TABLE-US-00089 TABLE 32 Classes of Enzyme Transformations Depicted in FIGS. 58, 62 and 63. LABEL FUNCTION 1.1.1.a Oxidoreductase (oxo to alcohol) 1.1.1.c Oxidoreductase (2 step, acyl-CoA to alcohol) 1.2.1.b Oxidoreductase (acyl-CoA to aldehyde) 1.2.1.c Oxidoreductase (2-oxo acid to acyl-CoA, decarboxylation) 1.2.1.d Oxidoreductase (phosphonate reductase) 1.2.1.e Acid reductase 1.4.1.a Oxidoreductase (aminating) 2.3.1.a Acyltransferase (transferring phosphate group to CoA) 2.6.1.a Aminotransferase 2.7.2.a Phosphotransferase (carboxy acceptor) 2.8.3.a CoA transferase 3.1.2.a CoA hydrolase 4.1.1.a Carboxy-lyase 6.2.1.a CoA synthetase
[0576] 1. 1. 1a Oxidoreductase (oxo to alcohol)
[0577] Aldehyde to Alcohol.
[0578] Three transformations described in FIGS. 58, 62 and 63 involve the conversion of an aldehyde to alcohol. These are 4-hydroxybutyrate dehydrogenase (step C, FIGS. 58 and 62), 1,4-butanediol dehydrogenase (step E, FIGS. 58 and 62), and 5-hydroxy-2-pentanoic acid (step Y, FIG. 62). Exemplary genes encoding enzymes that catalyze the conversion of an aldehyde to alcohol, that is, alcohol dehydrogenase or equivalently aldehyde reductase, include alrA encoding a medium-chain alcohol dehydrogenase for C2-C14 (Tani et al. Appl. Environ. Microbiol. 66:5231-5235 (2000)), ADH2 from Saccharomyces cerevisiae (Atsumi et al. Nature 451:86-89 (2008)), yqhD from E. coli, which has preference for molecules longer than C(3) (Sulzenbacher et al. J. Mol. Biol. 342:489-502 (2004)), and bdh I and bdh II from C. acetobutylicum, which converts butyryaldehyde into butanol (Walter et al. J. Bacteriol. 174:7149-7158 (1992)). The protein sequences for each of exemplary gene products can be found using the following GenBank accession numbers:
TABLE-US-00090 Gene Accession No. GI No. Organism alrA BAB12273.1 9967138 Acinetobacter sp. Strain M-1 ADH2 NP_014032.1 6323961 Saccharymyces cerevisiae yqhD NP_417484.1 16130909 Escherichia coli bdh I NP_349892.1 15896543 Clostridium acetobutylicum bdh II NP_349891.1 15896542 Clostridium acetobutylicum
[0579] Enzymes exhibiting 4-hydroxybutyrate dehydrogenase activity (EC 1.1.1.61) also fall into this category. Such enzymes have been characterized in Ralstonia eutropha (Bravo et al. J. Forensic Sci. 49:379-387 (2004)), Clostridium kluyveri (Wolff et al., Protein Expr. Purif. 6:206-212 (1995)) and Arabidopsis thaliana (Breitkreuz et al. J. Biol. Chem. 278:41552-41556 (2003)).
TABLE-US-00091 Gene Accession No. GI No. Organism 4hbd YP_726053.1 113867564 Ralstonia eutropha H16 4hbd EDK35022.1 146348486 Clostridium kluyveri DSM 555 4hbd Q94B07 75249805 Arabidopsis thaliana
[0580] The adh1 gene from Geobacillus thermoglucosidasius M10EXG (Jeon et al., J. Biotechnol. 135:127-133 (2008)) was shown to exhibit high activity on both 4-hydroxybutanal and butanal (see above). Thus this enzyme exhibits 1,4-butanediol dehydrogenase activity.
TABLE-US-00092 Gene Accession No. GI No. Organism adh1 AAR91477.1 40795502 Geobacillus thermoglucosidasius M10EXG
[0581] Another exemplary enzyme is 3-hydroxyisobutyrate dehydrogenase, which catalyzes the reversible oxidation of 3-hydroxyisobutyrate to methylmalonate semialdehyde. This enzyme participates in valine, leucine and isoleucine degradation and has been identified in bacteria, eukaryotes, and mammals. The enzyme encoded by P84067 from Thermus thermophilus HB8 has been structurally characterized (Lokanath et al. J. Mol. Biol. 352:905-17 (2005)). The reversibility of the human 3-hydroxyisobutyrate dehydrogenase was demonstrated using isotopically-labeled substrate (Manning et al., Biochem. J. 231:481-484 (1985)). Additional genes encoding this enzyme include 3hidh in Homo sapiens (Hawes et al., Methods Enzymol. 324:218-228 (2000)) and Oryctolagus cuniculus (Chowdhury et al., Biosci. Biotechnol. Biochem. 60:2043-2047 (1996); Hawes et al. Methods Enzymol. 324:218-228 (2000)), mmsb in Pseudomonas aeruginosa, and dhat in Pseudomonas putida (Aberhart et al., J. Chem. Soc. [Perkin 1] 6:1404-1406 (1979); Chowdhury et al., Biosci. Biotechnol Biochem. 67:438-441 (2003); Chowdhury et al., Biosci. Biotechnol. Biochem. 60:2043-2047 (1996)).
TABLE-US-00093 Gene Accession No. GI No. Organism P84067 P84067 75345323 Thermus thermophilus mmsb P28811.1 127211 Pseudomonas aeruginosa dhat Q59477.1 2842618 Pseudomonas putida 3hidh P31937.2 12643395 Homo sapiens 3hidh P32185.1 416872 Oryctolagus cuniculus
[0582] Several 3-hydroxyisobutyrate dehydrogenase enzymes have also been shown to convert malonic semialdehyde to 3-hydroxyproprionic acid (3-HP). Three gene candidates exhibiting this activity are mmsB from Pseudomonas aeruginosa PAO1(62), mmsB from Pseudomonas putida KT2440 (Liao et al., US Publication 2005/0221466) and mmsB from Pseudomonas putida E23 (Chowdhury et al., Biosci. Biotechnol. Biochem. 60:2043-2047 (1996)). An enzyme with 3-hydroxybutyrate dehydrogenase activity in Alcaligenes faecalis M3A has also been identified (Gokam et al., U.S. Pat. No. 7,393,676; Liao et al., US Publication No. 2005/0221466). Additional gene candidates from other organisms including Rhodobacter spaeroides can be inferred by sequence similarity.
TABLE-US-00094 Gene Accession No. GI No. Organism mmsB AAA25892.1 151363 Pseudomonas aeruginosa mmsB NP_252259.1 15598765 Pseudomonas aeruginosa PAO1 mmsB NP_746775.1 26991350 Pseudomonas putida KT2440 mmsB JC7926 60729613 Pseudomonas putida E23 orfB1 AAL26884 16588720 Rhodobacter spaeroides
[0583] The conversion of malonic semialdehyde to 3-HP can also be accomplished by two other enzymes, NADH-dependent 3-hydroxypropionate dehydrogenase and NADPH-dependent malonate semialdehyde reductase. An NADH-dependent 3-hydroxypropionate dehydrogenase is thought to participate in beta-alanine biosynthesis pathways from propionate in bacteria and plants (Rathinasabapathi, J. Plant Pathol. 159:671-674 (2002); Stadtman, J. Am. Chem. Soc. 77:5765-5766 (1955)). This enzyme has not been associated with a gene in any organism to date. NADPH-dependent malonate semialdehyde reductase catalyzes the reverse reaction in autotrophic CO.sub.2-fixing bacteria. Although the enzyme activity has been detected in Metallosphaera sedula, the identity of the gene is not known (Alber et al. J. Bacteriol. 188:8551-8559 (2006)).
1.1.1.c Oxidoreductase (2 Step, Acyl-CoA to Alcohol).
[0584] Steps S and W of FIG. 62 depict bifunctional reductase enzymes that can form 4-hydroxybutyrate and 1,4-butanediol, respectively. Exemplary 2-step oxidoreductases that convert an acyl-CoA to alcohol include those that transform substrates such as acetyl-CoA to ethanol (for example, adhE from E. coli (Kessler et al., FEBS. Lett. 281:59-63 (1991)) and butyryl-CoA to butanol (for example, adhE2 from C. acetobutylicum (Fontaine et al., J. Bacteriol. 184:821-830 (2002)). The C. acetobutylicum adhE2 gene was shown to convert 4-hydroxybutyryl-CoA to 1,4-butanediol (see above). In addition to reducing acetyl-CoA to ethanol, the enzyme encoded by adhE in Leuconostoc mesenteroides has been shown to oxidize the branched chain compound isobutyraldehyde to isobutyryl-CoA (Kazahaya et al. J., Gen. Appl. Microbiol. 18:43-55 (1972); Koo et al., Biotechnol. Lett. 27:505-510 (2005)).
TABLE-US-00095 Gene Accession No. GI No. Organism adhE NP_415757.1 16129202 Escherichia coli adhE2 AAK09379.1 12958626 Clostridium acetobutylicum adhE AAV66076.1 55818563 Leuconostoc mesenteroides
[0585] Another exemplary enzyme can convert malonyl-CoA to 3-HP. An NADPH-dependent enzyme with this activity has characterized in Chloroflexus aurantiacus, where it participates in the 3-hydroxypropionate cycle (Hugler et al., J. Bacteriol. 184:2404-2410 (2002); Strauss and Fuchs, Eur. J. Biochem. 215:633-643 (1993)). This enzyme, with a mass of 300 kDa, is highly substrate-specific and shows little sequence similarity to other known oxidoreductases (Hugler et al., J. Bacteriol. 184:2404-2410 (2002)). No enzymes in other organisms have been shown to catalyze this specific reaction; however there is bioinformatic evidence that other organisms may have similar pathways (Klatt et al., Environ. Microbiol. 9:2067-2078 (2007)). Enzyme candidates in other organisms including Roseiflexus castenholzii, Erythrobacter sp. NAP1 and marine gamma proteobacterium HTCC2080 can be inferred by sequence similarity.
TABLE-US-00096 Gene Accession No. GI No. Organism mcr AAS20429.1 42561982 Chloroflexus aurantiacus Rcas_2929 YP_001433009.1 156742880 Roseiflexus castenholzii NAP1_02720 ZP_01039179.1 85708113 Erythrobacter sp. NAP1 MGP2080_00535 ZP_01626393.1 119504313 marine gamma proteobacterium HTCC2080
[0586] Longer chain acyl-CoA molecules can be reduced by enzymes such as the jojoba (Simmondsia chinensis) FAR, which encodes an alcohol-forming fatty acyl-CoA reductase. Its overexpression in E. coli resulted in FAR activity and the accumulation of fatty alcohol (Metz et al., Plant Physiol. 122:635-644 2000)).
TABLE-US-00097 Gene Accession No. GI No. Organism FAR AAD38039.1 5020215 Simmondsia chinensis
1.2.1.b Oxidoreductase (Acyl-CoA to Aldehyde).
[0587] Step A of FIGS. 58, 62 and 63 involves the conversion of succinyl-CoA to succinate semialdehyde by an aldehyde forming succinyl-CoA reductase. Step Q of FIG. 62 depicts the conversion of 4-hydroxybutyryl-CoA to 4-hydroxybutanal by an aldehyde-forming 4-hydroxybutyryl-CoA reductase. Several acyl-CoA dehydrogenases are capable of reducing an acyl-CoA to its corresponding aldehyde. Exemplary genes that encode such enzymes include the Acinetobacter calcoaceticus acr1 encoding a fatty acyl-CoA reductase (Reiser and Somerville, J. Bacteriol. 179:2969-2975 (1997)), the Acinetobacter sp. M-1 fatty acyl-CoA reductase (Ishige et al., Appl. Environ. Microbiol. 68:1192-1195 (2002)), and a CoA- and NADP-dependent succinate semialdehyde dehydrogenase encoded by the sucD gene in Clostridium kluyveri (Sohling and Gottschalk, J. Bacteriol. 178:871-80 (1996); Sohling and Gottschalk, J. Bacteriol. 178:871-880 (1996)). SucD of P. gingivalis is another aldehyde-forming succinyl-CoA reductase (Takahashi et al., J. Bacteriol. 182:4704-4710 (2000)). The enzyme acylating acetaldehyde dehydrogenase in Pseudomonas sp, encoded by bphG, is yet another as it has been demonstrated to oxidize and acylate acetaldehyde, propionaldehyde, butyraldehyde, isobutyraldehyde and formaldehyde (Powlowski et al., J. Bacteriol. 175:377-385 (1993)). In addition to reducing acetyl-CoA to ethanol, the enzyme encoded by adhE in Leuconostoc mesenteroides has been shown to oxidize the branched chain compound isobutyraldehyde to isobutyryl-CoA (Koo et al., Biotechnol. Lett. 27:505-510 (2005)). Butyraldehyde dehydrogenase catalyzes a similar reaction, conversion of butyryl-CoA to butyraldehyde, in solventogenic organisms such as Clostridium saccharoperbutylacetonicum (Kosaka et al., Biosci. Biotechnol. Biochem. 71:58-68 (2007)).
TABLE-US-00098 Gene Accession No. GI No. Organism acr1 YP_047869.1 50086359 Acinetobacter calcoaceticus acr1 AAC45217 1684886 Acinetobacter baylyi acr1 BAB85476.1 18857901 Acinetobacter sp. Strain M-1 sucD P38947.1 730847 Clostridium kluyveri sucD NP_904963.1 34540484 Porphyromonas gingivalis bphG BAA03892.1 425213 Pseudomonas sp adhE AAV66076.1 55818563 Leuconostoc mesenteroides bld AAP42563.1 31075383 Clostridium saccharoperbutylacetonicum
[0588] An additional enzyme type that converts an acyl-CoA to its corresponding aldehyde is malonyl-CoA reductase, which transforms malonyl-CoA to malonic semialdehyde. Malonyl-CoA reductase is a key enzyme in autotrophic carbon fixation via the 3-hydroxypropionate cycle in thermoacidophilic archael bacteria (Berg et al., Science 318:1782-1786 (2007); Thauer, Science 318:1732-1733 (2007)). The enzyme utilizes NADPH as a cofactor and has been characterized in Metallosphaera and Sulfolobus spp (Alber et al., J. Bacteriol. 188:8551-8559 (2006); Hugler et al., J. Bacteriol. 184:2404-2410 (2002)). The enzyme is encoded by Msed_0709 in Metallosphaera sedula (Alber et al., J. Bacteriol. 188:8551-8559 (2006); Berg et al., Science 318:1782-1786 (2007)). A gene encoding a malonyl-CoA reductase from Sulfolobus tokodaii was cloned and heterologously expressed in E. coli (Alber et al., J. Bacteriol. 188:8551-8559 (2006)). Although the aldehyde dehydrogenase functionality of these enzymes is similar to the bifunctional dehydrogenase from Chloroflexus aurantiacus, there is little sequence similarity. Both malonyl-CoA reductase enzyme candidates have high sequence similarity to aspartate-semialdehyde dehydrogenase, an enzyme catalyzing the reduction and concurrent dephosphorylation of aspartyl-4-phosphate to aspartate semialdehyde. Additional gene candidates can be found by sequence homology to proteins in other organisms including Sulfolobus solfataricus and Sulfolobus acidocaldarius. Yet another candidate for CoA-acylating aldehyde dehydrogenase is the ald gene from Clostridium beijerinckii (Toth et al., Appl. Environ. Microbiol. 65:4973-4980 (1999)). This enzyme has been reported to reduce acetyl-CoA and butyryl-CoA to their corresponding aldehydes. This gene is very similar to eutE that encodes acetaldehyde dehydrogenase of Salmonella typhimurium and E. coli (Toth et al., Appl. Environ. Microbiol. 65:4973-4980 (1999)).
TABLE-US-00099 Gene Accession No. GI No. Organism Msed_0709 YP_001190808.1 146303492 Metallosphaera sedula mcr NP_378167.1 15922498 Sulfolobus tokodaii asd-2 NP_343563.1 15898958 Sulfolobus solfataricus Saci_2370 YP_256941.1 70608071 Sulfolobus acidocaldarius Ald AAT66436 49473535 Clostridium beijerinckii eutE AAA80209 687645 Salmonella typhimurium eutE P77445 2498347 Escherichia coli
1.2.1.c Oxidoreductase (2-Oxo Acid to Acyl-CoA, Decarboxylation).
[0589] Step AA in FIG. 62 depicts the conversion of 5-hydroxy-2-oxopentanoic acid to 4-hydroxybutyryl-CoA. Candidate enzymes for this transformation include 1) branched-chain 2-keto-acid dehydrogenase, 2) alpha-ketoglutarate dehydrogenase, and 3) the pyruvate dehydrogenase multienzyme complex (PDHC). These enzymes are multi-enzyme complexes that catalyze a series of partial reactions which result in acylating oxidative decarboxylation of 2-keto-acids. Each of the 2-keto-acid dehydrogenase complexes occupies key positions in intermediary metabolism, and enzyme activity is typically tightly regulated (Fries et al. Biochemistry 42:6996-7002 (2003)). The enzymes share a complex but common structure composed of multiple copies of three catalytic components: alpha-ketoacid decarboxylase (E1), dihydrolipoamide acyltransferase (E2) and dihydrolipoamide dehydrogenase (E3). The E3 component is shared among all 2-keto-acid dehydrogenase complexes in an organism, while the E1 and E2 components are encoded by different genes. The enzyme components are present in numerous copies in the complex and utilize multiple cofactors to catalyze a directed sequence of reactions via substrate channeling. The overall size of these dehydrogenase complexes is very large, with molecular masses between 4 and 10 million Da (that is, larger than a ribosome).
[0590] Activity of enzymes in the 2-keto-acid dehydrogenase family is normally low or limited under anaerobic conditions in E. coli. Increased production of NADH (or NADPH) could lead to a redox-imbalance, and NADH itself serves as an inhibitor to enzyme function. Engineering efforts have increased the anaerobic activity of the E. coli pyruvate dehydrogenase complex (Kim et al. Appl. Environ. Microbiol. 73:1766-1771 (2007); Kim et al. J. Bacteriol. 190:3851-3858) 2008); Zhou et al. Biotechnol. Lett. 30:335-342 (2008)). For example, the inhibitory effect of NADH can be overcome by engineering an H322Y mutation in the E3 component (Kim et al. J. Bacteriol. 190:3851-3858 (2008)). Structural studies of individual components and how they work together in complex provide insight into the catalytic mechanisms and architecture of enzymes in this family (Aevarsson et al. Nat. Struct. Biol. 6:785-792 (1999); Zhou et al. Proc. Natl. Acad. Sci. U.S.A. 98:14802-14807 (2001)). The substrate specificity of the dehydrogenase complexes varies in different organisms, but generally branched-chain keto-acid dehydrogenases have the broadest substrate range.
[0591] Alpha-ketoglutarate dehydrogenase (AKGD) converts alpha-ketoglutarate to succinyl-CoA and is the primary site of control of metabolic flux through the TCA cycle (Hansford, R. G. Curr. Top. Bioenerg. 10:217-278 (1980)). Encoded by genes sucA, sucB and lpd in E. coli, AKGD gene expression is downregulated under anaerobic conditions and during growth on glucose (Park et al. Mol. Microbiol. 15:473-482 (1995)). Although the substrate range of AKGD is narrow, structural studies of the catalytic core of the E2 component pinpoint specific residues responsible for substrate specificity (Knapp et al. J. Mol. Biol. 280:655-668 (1998)). The Bacillus subtilis AKGD, encoded by odhAB (E1 and E2) and pdhD (E3, shared domain), is regulated at the transcriptional level and is dependent on the carbon source and growth phase of the organism (Resnekov et al. Mol. Gen. Genet. 234:285-296 (1992)). In yeast, the LPD1 gene encoding the E3 component is regulated at the transcriptional level by glucose (Roy and Dawes J. Gen. Microbiol. 133:925-933 (1987)). The E1 component, encoded by KGD1, is also regulated by glucose and activated by the products of HAP2 and HAP3 (Repetto and Tzagoloff Mol. Cell Biol. 9:2695-2705 (1989)). The AKGD enzyme complex, inhibited by products NADH and succinyl-CoA, is well-studied in mammalian systems, as impaired function of has been linked to several neurological diseases (Tretter and dam-Vizi Philos. Trans. R. Soc. Lond B Biol. Sci. 360:2335-2345 (2005)).
TABLE-US-00100 Gene Accession No. GI No. Organism sucA NP_415254.1 16128701 Escherichia coli str. K12 substr. MG1655 sucB NP_415255.1 16128702 Escherichia coli str. K12 substr. MG1655 lpd NP_414658.1 16128109 Escherichia coli str. K12 substr. MG1655 odhA P23129.2 51704265 Bacillus subtilis odhB P16263.1 129041 Bacillus subtilis pdhD P21880.1 118672 Bacillus subtilis KGD1 NP_012141.1 6322066 Saccharomyces cerevisiae KGD2 NP_010432.1 6320352 Saccharomyces cerevisiae LPD1 NP_116635.1 14318501 Saccharomyces cerevisiae
[0592] Branched-chain 2-keto-acid dehydrogenase complex (BCKAD), also known as 2-oxoisovalerate dehydrogenase, participates in branched-chain amino acid degradation pathways, converting 2-keto acids derivatives of valine, leucine and isoleucine to their acyl-CoA derivatives and CO.sub.2. The complex has been studied in many organisms including Bacillus subtilis (Wang et al. Eur. J. Biochem. 213:1091-1099 (1993)), Rattus norvegicus (Namba et al. J. Biol. Chem. 244:4437-4447 (1969)) and Pseudomonas putida (Sokatch J. Bacteriol. 148:647-652 (1981)). In Bacillus subtilis the enzyme is encoded by genes pdhD (E3 component), bfmBB (E2 component), bfmBAA and bfmBAB (E1 component) (Wang et al. Eur. J. Biochem. 213:1091-1099 (1993)). In mammals, the complex is regulated by phosphorylation by specific phosphatases and protein kinases. The complex has been studied in rat hepatocites (Chicco et al. J. Biol. Chem. 269:19427-19434 (1994)) and is encoded by genes Bckdha (E1 alpha), Bckdhb (E1 beta), Dbt (E2), and Dld (E3). The E1 and E3 components of the Pseudomonas putida BCKAD complex have been crystallized (Aevarsson et al. Nat. Struct. Biol. 6:785-792 (1999); Mattevi Science 255:1544-1550 (1992)) and the enzyme complex has been studied (Sokatch et al. J. Bacteriol. 148:647-652 (1981)). Transcription of the P. putida BCKAD genes is activated by the gene product of bkdR (Hester et al. Eur. J. Biochem. 233:828-836 (1995)). In some organisms including Rattus norvegicus (Paxton et al. Biochem. J. 234:295-303 (1986)) and Saccharomyces cerevisiae (Sinclair et al. Biochem. Mol. Biol. Int. 31:911-922 (1993)), this complex has been shown to have a broad substrate range that includes linear oxo-acids such as 2-oxobutanoate and alpha-ketoglutarate, in addition to the branched-chain amino acid precursors. The active site of the bovine BCKAD was engineered to favor alternate substrate acetyl-CoA (Meng and Chuang, Biochemistry 33:12879-12885 (1994)).
TABLE-US-00101 Gene Accession No. GI No. Organism bfmBB NP_390283.1 16079459 Bacillus subtilis bfmBAA NP_390285.1 16079461 Bacillus subtilis bfmBAB NP_390284.1 16079460 Bacillus subtilis pdhD P21880.1 118672 Bacillus subtilis lpdV P09063.1 118677 Pseudomonas putida bkdB P09062.1 129044 Pseudomonas putida bkdA1 NP_746515.1 26991090 Pseudomonas putida bkdA2 NP_746516.1 26991091 Pseudomonas putida Bckdha NP_036914.1 77736548 Rattus norvegicus Bckdhb NP_062140.1 158749538 Rattus norvegicus Dbt NP_445764.1 158749632 Rattus norvegicus Dld NP_955417.1 40786469 Rattus norvegicus
[0593] The pyruvate dehydrogenase complex, catalyzing the conversion of pyruvate to acetyl-CoA, has also been extensively studied. In the E. coli enzyme, specific residues in the E1 component are responsible for substrate specificity (Bisswanger, H. J Biol Chem. 256:815-822 (1981); Bremer, J. Eur. J Biochem. 8:535-540 (1969); Gong et al. J Biol Chem. 275:13645-13653 (2000)). As mentioned previously, enzyme engineering efforts have improved the E. coli PDH enzyme activity under anaerobic conditions (Kim et al. Appl. Environ. Microbiol. 73:1766-1771 (2007); Kim J. Bacteriol. 190:3851-3858 (2008); Zhou et al. Biotechnol. Lett. 30:335-342 (2008)). In contrast to the E. coli PDH, the B. subtilis complex is active and required for growth under anaerobic conditions (Nakano J. Bacteriol. 179:6749-6755 (1997)). The Klebsiella pneumoniae PDH, characterized during growth on glycerol, is also active under anaerobic conditions (Menzel et al. J. Biotechnol. 56:135-142 (1997)). Crystal structures of the enzyme complex from bovine kidney (Zhou et al. Proc. Natl. Acad. Sci. U.S.A. 98:14802-14807 (2001)) and the E2 catalytic domain from Azotobacter vinelandii are available (Mattevi et al. Science 255:1544-1550 (1992)). Some mammalian PDH enzymes complexes can react on alternate substrates such as 2-oxobutanoate, although comparative kinetics of Rattus norvegicus PDH and BCKAD indicate that BCKAD has higher activity on 2-oxobutanoate as a substrate (Paxton et al. Biochem. J. 234:295-303 (1986)).
TABLE-US-00102 Gene Accession No. GI No. Organism aceE NP_414656.1 16128107 Escherichia coli str. K12 substr. MG1655 aceF NP_414657.1 16128108 Escherichia coli str. K12 substr. MG1655 lpd NP_414658.1 16128109 Escherichia coli str. K12 substr. MG1655 pdhA P21881.1 3123238 Bacillus subtilis pdhB P21882.1 129068 Bacillus subtilis pdhC P21883.2 129054 Bacillus subtilis pdhD P21880.1 118672 Bacillus subtilis aceE YP_001333808.1 152968699 Klebsiella pneumonia MGH78578 aceF YP_001333809.1 152968700 Klebsiella pneumonia MGH78578 lpdA YP_001333810.1 152968701 Klebsiella pneumonia MGH78578 Pdha1 NP_001004072.2 124430510 Rattus norvegicus Pdha2 NP_446446.1 16758900 Rattus norvegicus Dlat NP_112287.1 78365255 Rattus norvegicus Dld NP_955417.1 40786469 Rattus norvegicus
[0594] As an alternative to the large multienzyme 2-keto-acid dehydrogenase complexes described above, some anaerobic organisms utilize enzymes in the 2-ketoacid oxidoreductase family (OFOR) to catalyze acylating oxidative decarboxylation of 2-keto-acids. Unlike the dehydrogenase complexes, these enzymes contain iron-sulfur clusters, utilize different cofactors, and use ferredoxin or flavodixin as electron acceptors in lieu of NAD(P)H. While most enzymes in this family are specific to pyruvate as a substrate (POR) some 2-keto-acid:ferredoxin oxidoreductases have been shown to accept a broad range of 2-ketoacids as substrates including alpha-ketoglutarate and 2-oxobutanoate (Fukuda and Wakagi Biochim. Biophys. Acta 1597:74-80 (2002); Zhang et al. J. Biochem. 120:587-599 (1996)). One such enzyme is the OFOR from the thermoacidophilic archaeon Sulfolobus tokodaii 7, which contains an alpha and beta subunit encoded by gene ST2300 (Fukuda and Wakagi Biochim. Biophys. Acta 1597:74-80 (2002); Zhang et al. J. Biochem. 120:587-599 (1996)). A plasmid-based expression system has been developed for efficiently expressing this protein in E. coli (Fukuda et al. Eur. J. Biochem. 268:5639-5646 (2001)) and residues involved in substrate specificity were determined (Fukuda and Wakagi Biochim. Biophys. Acta 1597:74-80 (2002)). Two OFORs from Aeropyrum pernix str. K1 have also been recently cloned into E. coli, characterized, and found to react with a broad range of 2-oxoacids (Nishizawa et al. FEBS Lett. 579:2319-2322 (2005)). The gene sequences of these OFOR candidates are available, although they do not have GenBank identifiers assigned to date. There is bioinformatic evidence that similar enzymes are present in all archaea, some anaerobic bacteria and amitochondrial eukarya (Fukuda and Wakagi Biochim. Biophys. Acta 1597:74-80 (2005)). This class of enzyme is also interesting from an energetic standpoint, as reduced ferredoxin could be used to generate NADH by ferredoxin-NAD reductase (Petitdemange et al. Biochim. Biophys. Acta 421:334-337 (1976)). Also, since most of the enzymes are designed to operate under anaerobic conditions, less enzyme engineering may be required relative to enzymes in the 2-keto-acid dehydrogenase complex family for activity in an anaerobic environment.
TABLE-US-00103 Gene Accession No. GI No. Organism ST2300 NP_378302.1 15922633 Sulfolobus tokodaii 7
1.2.1.d Oxidoreductase (Phosphonate Reductase).
[0595] The conversion of 4-hydroxybutyryl-phosphate to 4-hydroxybutanal can be catalyzed by an oxidoreductase in the EC class 1.2.1. Aspartate semialdehyde dehydrogenase (ASD, EC 1.2.1.11) catalyzes the NADPH-dependent reduction of 4-aspartyl phosphate to aspartate-4-semialdehyde. ASD participates in amino acid biosynthesis and recently has been studied as an antimicrobial target (Hadfield et al., Biochemistry 40:14475-14483 (2001). The E. coli ASD structure has been solved (Hadfield et al., J. Mol. Biol. 289:991-1002 (1999)) and the enzyme has been shown to accept the alternate substrate beta-3-methylaspartyl phosphate (Shames et al., J. Biol. Chem. 259:15331-15339 (1984)). The Haemophilus influenzae enzyme has been the subject of enzyme engineering studies to alter substrate binding affinities at the active site (Blanco et al., Acta Crystallogr. D. Biol. Crystallogr. 60:1388-1395 (2004); Blanco et al., Acta Crystallogr. D. Biol. Crystallogr. 60:1808-1815 (2004)). Other ASD candidates are found in Mycobacterium tuberculosis (Shafiani et al., J. Appl. Microbiol. 98:832-838 (2005), Methanococcus jannaschii (Faehnle et al., J. Mol. Biol. 353:1055-1068 (2005)), and the infectious microorganisms Vibrio cholera and Heliobacter pylori (Moore et al., Protein Expr. Purif. 25:189-194 (2002)). A related enzyme candidate is acetylglutamylphosphate reductase (EC 1.2.1.38), an enzyme that naturally reduces acetylglutamylphosphate to acetylglutamate-5-semialdehyde, found in S. cerevisiae (Pauwels et al., Eur. J. Biochem. 270:1014-1024 (2003), B. subtilis (O'Reilly and Devine, Microbiology 140 (Pt 5):1023-1025 (1994)). and other organisms.
TABLE-US-00104 Gene Accession No. GI No. Organism asd NP_417891.1 16131307 Escherichia coli asd YP_248335.1 68249223 Haemophilus influenzae asd AAB49996 1899206 Mycobacterium tuberculosis VC2036 NP_231670 15642038 Vibrio cholera asd YP_002301787.1 210135348 Heliobacter pylori ARG5,6 NP_010992.1 6320913 Saccharomyces cerevisiae argC NP_389001.1 16078184 Bacillus subtilis
[0596] Other exemplary enzymes in this class include glyceraldehyde 3-phosphate dehydrogenase which converts glyceraldehyde-3-phosphate into D-glycerate 1,3-bisphosphate (for example, E. coli gapA (Branlant and Branlant, Eur. J. Biochem. 150:61-66 (1985)), N-acetyl-gamma-glutamyl-phosphate reductase which converts N-acetyl-L-glutamate-5-semialdehyde into N-acetyl-L-glutamyl-5-phosphate (for example, E. coli argC (Parsot et al., Gene 68:275-283 (1988)), and glutamate-5-semialdehyde dehydrogenase, which converts L-glutamate-5-semialdehyde into L-glutamyl-5-phosphate (for example, E. coli proA (Smith et al., J. Bacteriol. 157:545-551 (1984)). Genes encoding glutamate-5-semialdehyde dehydrogenase enzymes from Salmonella typhimurium (Mahan and Csonka, J. Bacteriol. 156:1249-1262 (1983)) and Campylobacter jejuni (Louie and Chan, Mol. Gen. Genet. 240:29-35 (1993)) were cloned and expressed in E. coli.
TABLE-US-00105 Gene Accession No. GI No. Organism gapA P0A9B2.2 71159358 Escherichia coli argC NP_418393.1 16131796 Escherichia coli proA NP_414778.1 16128229 Escherichia coli proA NP_459319.1 16763704 Salmonella typhimurium proA P53000.2 9087222 Campylobacter jejuni
1.2.1.e Acid Reductase.
[0597] Several steps in FIGS. 58, 62 and 63 depict the conversion of unactivated acids to aldehydes by an acid reductase. These include the conversion of 4-hydroxybutyrate, succinate, alpha-ketoglutarate, and 4-aminobutyrate to 4-hydroxybutanal, succinate semialdehyde, 2,5-dioxopentanoate, and 4-aminobutanal, respectively. One notable carboxylic acid reductase can be found in Nocardia iowensis which catalyzes the magnesium, ATP and NADPH-dependent reduction of carboxylic acids to their corresponding aldehydes (Venkitasubramanian et al., J. Biol. Chem. 282:478-485 (2007)). This enzyme is encoded by the car gene and was cloned and functionally expressed in E. coli (Venkitasubramanian et al., J. Biol. Chem. 282:478-485 (2007)). Expression of the npt gene product improved activity of the enzyme via post-transcriptional modification. The npt gene encodes a specific phosphopantetheine transferase (PPTase) that converts the inactive apo-enzyme to the active holo-enzyme. The natural substrate of this enzyme is vanillic acid, and the enzyme exhibits broad acceptance of aromatic and aliphatic substrates (Venkitasubramanian et al., in Biocatalysis in the Pharmaceutical and Biotechnology Industires, ed. R. N. Patel, Chapter 15, pp. 425-440, CRC Press LLC, Boca Raton, Fla. (2006)).
TABLE-US-00106 Accession Gene No. GI No. Organism car AAR91681.1 40796035 Nocardia iowensis (sp. NRRL 5646) npt ABI83656.1 114848891 Nocardia iowensis (sp. NRRL 5646)
[0598] Additional car and npt genes can be identified based on sequence homology.
TABLE-US-00107 Gene Accession No. GI No. Organism fadD9 YP_978699.1 121638475 Mycobacterium bovis BCG BCG_2812c YP_978898.1 121638674 Mycobacterium bovis BCG nfa20150 YP_118225.1 54023983 Nocardia farcinica IFM 10152 nfa40540 YP_120266.1 54026024 Nocardia farcinica IFM 10152 SGR_6790 YP_001828302.1 182440583 Streptomyces griseus subsp. griseus NBRC 13350 SGR_665 YP_001822177.1 182434458 Streptomyces griseus subsp. griseus NBRC 13350
[0599] An additional enzyme candidate found in Streptomyces griseus is encoded by the griC and griD genes. This enzyme is believed to convert 3-amino-4-hydroxybenzoic acid to 3-amino-4-hydroxybenzaldehyde as deletion of either griC or griD led to accumulation of extracellular 3-acetylamino-4-hydroxybenzoic acid, a shunt product of 3-amino-4-hydroxybenzoic acid metabolism (Suzuki, et al., J. Antibiot. 60(6):380-387 (2007)). Co-expression of griC and griD with SGR_665, an enzyme similar in sequence to the Nocardia iowensis npt, can be beneficial.
TABLE-US-00108 Gene Accession No. GI No. Organism griC 182438036 YP_001825755.1 Streptomyces griseus subsp. griseus NBRC 13350 griD 182438037 YP_001825756.1 Streptomyces griseus subsp. griseus NBRC 13350 MSMEG_2956 YP_887275.1 YP_887275.1 Mycobacterium smegmatis MC2 155 MSMEG_5739 YP_889972.1 118469671 Mycobacterium smegmatis MC2 155 MSMEG_2648 YP_886985.1 118471293 Mycobacterium smegmatis MC2 155 MAP1040c NP_959974.1 41407138 Mycobacterium avium subsp. paratuberculosis K-10 MAP2899c NP_961833.1 41408997 Mycobacterium avium subsp. paratuberculosis K-10 MMAR_2117 YP_001850422.1 183982131 Mycobacterium marinum M MMAR_2936 YP_001851230.1 183982939 Mycobacterium marinum M MMAR_1916 YP_001850220.1 183981929 Mycobacterium marinum M TpauDRAFT_33060 ZP_04027864.1 227980601 Tsukamurella paurometabola DSM 20162 TpauDRAFT_20920 ZP_04026660.1 227979396 Tsukamurella paurometabola DSM 20162 CPCC7001_1320 ZP_05045132.1 254431429 Cyanobium PCC7001 DDBDRAFT_0187729 XP_636931.1 66806417 Dictyostelium discoideum AX4
[0600] An enzyme with similar characteristics, alpha-aminoadipate reductase (AAR, EC 1.2.1.31), participates in lysine biosynthesis pathways in some fungal species. This enzyme naturally reduces alpha-aminoadipate to alpha-aminoadipate semialdehyde. The carboxyl group is first activated through the ATP-dependent formation of an adenylate that is then reduced by NAD(P)H to yield the aldehyde and AMP. Like CAR, this enzyme utilizes magnesium and requires activation by a PPTase. Enzyme candidates for AAR and its corresponding PPTase are found in Saccharomyces cerevisiae (Morris et al., Gene 98:141-145 (1991)), Candida albicans (Guo et al., Mol. Genet. Genomics 269:271-279 (2003)), and Schizosaccharomyces pombe (Ford et al., Curr. Genet. 28:131-137 (1995)). The AAR from S. pombe exhibited significant activity when expressed in E. coli (Guo et al., Yeast 21:1279-1288 (2004)). The AAR from Penicillium chrysogenum accepts S-carboxymethyl-L-cysteine as an alternate substrate, but did not react with adipate, L-glutamate or diaminopimelate (Hijarrubia et al., J. Biol. Chem. 278:8250-8256 (2003)). The gene encoding the P. chrysogenum PPTase has not been identified to date.
TABLE-US-00109 Gene Accession No. GI No. Organism LYS2 AAA34747.1 171867 Saccharomyces cerevisiae LYS5 P50113.1 1708896 Saccharomyces cerevisiae LYS2 AAC02241.1 2853226 Candida albicans LYS5 AAO26020.1 28136195 Candida albicans Lys1p P40976.3 13124791 Schizosaccharomyces pombe Lys7p Q10474.1 1723561 Schizosaccharomyces pombe Lys2 CAA74300.1 3282044 Penicillium chrysogenum
1.4.1.a Oxidoreductase (Aminating).
[0601] Glutamate dehydrogenase (Step J, FIGS. 62 and 63), 4-aminobutyrate dehydrogenase (Step M, FIGS. 62 and 63), putrescine dehydrogenase (Step D, FIG. 63), 5-amino-2-oxopentanoate dehydrogenase (Step P, FIG. 63), and ornithine dehydrogenase (Step S, FIG. 63) can be catalyzed by aminating oxidoreductases. Enzymes in this EC class catalyze the oxidative deamination of alpha-amino acids with NAD+ or NADP+ as acceptor, and the reactions are typically reversible. Exemplary oxidoreductases operating on amino acids include glutamate dehydrogenase (deaminating), encoded by gdhA, leucine dehydrogenase (deaminating), encoded by ldh, and aspartate dehydrogenase (deaminating), encoded by nadX. The gdhA gene product from Escherichia coli (Korber et al., J. Mol. Biol. 234:1270-1273 (1993); McPherson and Wootton, Nucleic Acids Res. 11:5257-5266 (1983)), gdh from Thermotoga maritima (Kort et al., Extremophiles 1:52-60 (1997); Lebbink, et al. J. Mol. Biol. 280:287-296 (1998); Lebbink et al. J. Mol. Biol. 289:357-369 (1999)), and gdhA1 from Halobacterium salinarum (Ingoldsby et al., Gene 349:237-244 (2005)) catalyze the reversible interconversion of glutamate to 2-oxoglutarate and ammonia, while favoring NADP(H), NAD(H), or both, respectively. The ldh gene of Bacillus cereus encodes the LeuDH protein that has a wide of range of substrates including leucine, isoleucine, valine, and 2-aminobutanoate (Ansorge and Kula, Biotechnol. Bioeng. 68:557-562 (2000); Stoyan et al. J. Biotechnol. 54:77-80 (1997)). The nadX gene from Thermotoga maritime encoding for the aspartate dehydrogenase is involved in the biosynthesis of NAD (Yang et al., J. Biol. Chem. 278:8804-8808 (2003)).
TABLE-US-00110 Gene Accession No. GI No. Organism gdhA P00370 118547 Escherichia coli gdh P96110.4 6226595 Thermotoga maritima gdhA1 NP_279651.1 15789827 Halobacterium salinarum ldh P0A393 61222614 Bacillus cereus nadX NP_229443.1 15644391 Thermotoga maritima
[0602] Additional glutamate dehydrogenase gene candidates are found in Bacillus subtilis (Khan et al., Biosci. Biotechnol. Biochem. 69:1861-1870 (2005)), Nicotiana tabacum (Purnell et al., Planta 222:167-180 (2005)), Oryza sativa (Abiko et al., Plant Cell Physiol. 46:1724-1734 (2005)), Haloferax mediterranei (Diaz et al., Extremophiles 10:105-115 (2006)) and Halobactreium salinarum (Hayden et al., FEMS Microbiol. Lett. 211:37-41 (2002)). The Nicotiana tabacum enzyme is composed of alpha and beta subunits encoded by gdh1 and gdh2 (Purnell et al., Planta 222:167-180 (2005)). Overexpression of the NADH-dependent glutamate dehydrogenase was found to improve ethanol production in engineered strains of S. cerevisiae (Roca et al., Appl. Environ. Microbiol. 69:4732-4736 (2003)).
TABLE-US-00111 Gene Accession No. GI No. Organism rocG NP_391659.1 16080831 Bacillus subtilis gdh1 AAR11534.1 38146335 Nicotiana tabacum gdh2 AAR11535.1 38146337 Nicotiana tabacum GDH Q852M0 75243660 Oryza sativa GDH Q977U6 74499858 Haloferax mediterranei GDH P29051 118549 Halobactreium salinarum GDH2 NP_010066.1 6319986 Saccharomyces cerevisiae
[0603] An exemplary enzyme for catalyzing the conversion of aldehydes to their corresponding primary amines is lysine 6-dehydrogenase (EC 1.4.1.18), encoded by the lysDH genes. The lysine 6-dehydrogenase (deaminating), encoded by lysDH gene, catalyze the oxidative deamination of the .epsilon.-amino group of L-lysine to form 2-aminoadipate-6-semialdehyde, which in turn nonenzymatically cyclizes to form .DELTA.1-piperideine-6-carboxylate (Misono and Nagasaki, J. Bacteriol. 150:398-401 (1982)). The lysDH gene from Geobacillus stearothermophilus encodes a thermophilic NAD-dependent lysine 6-dehydrogenase (Heydari et al., Appl. Environ. Microbiol 70:937-942 (2004)). The lysDH gene from Aeropyrum pernix K1 is identified through homology from genome projects. Additional enzymes can be found in Agrobacterium tumefaciens (Hashimoto et al., J. Biochem. 106:76-80 (1989); Misono and Nagasaki, J. Bacteriol. 150:398-401 (1982)) and Achromobacter denitrificans (Ruldeekulthamrong et al., BMB. Rep. 41:790-795 (2008)).
TABLE-US-00112 Gene Accession No. GI No. Organism lysDH BAB39707 13429872 Geobacillus stearothermophilus lysDH NP_147035.1 14602185 Aeropyrum pernix K1 lysDH NP_353966 15888285 Agrobacterium tumefaciens lysDH AAZ94428 74026644 Achromobacter denitrificans
[0604] An enzyme that converts 3-oxoacids to 3-amino acids is 3,5-diaminohexanoate dehydrogenase (EC 1.4.1.11), an enzyme found in organisms that ferment lysine. The gene encoding this enzyme, kdd, was recently identified in Fusobacterium nucleatum (Kreimeyer et al., J. Biol. Chem. 282:7191-7197 (2007)). The enzyme has been purified and characterized in other organisms (Baker et al., J. Biol. Chem. 247:7724-7734 (1972); Baker and van der Drift, Biochemistry 13:292-299 (1974)), but the genes associated with these enzymes are not known. Candidates in Myxococcus xanthus, Porphyromonas gingivalis W83 and other sequenced organisms can be inferred by sequence homology.
TABLE-US-00113 Gene Accession No. GI No. Organism kdd AAL93966.1 19713113 Fusobacterium nucleatum mxan_4391 ABF87267.1 108462082 Myxococcus xanthus pg_1069 AAQ66183.1 34397119 Porphyromonas gingivalis
2.3.1.a Acyltransferase (Transferring Phosphate Group to CoA).
[0605] Step P of FIG. 62 depicts the transformation of 4-hydroxybutyryl-CoA to 4-hydroxybutyryl-Pi. Exemplary phosphate transferring acyltransferases include phosphotransacetylase, encoded by pta, and phosphotransbutyrylase, encoded by ptb. The pta gene from E. coli encodes an enzyme that can convert acetyl-CoA into acetyl-phosphate, and vice versa (Suzuki, Biochim. Biophys. Acta 191:559-569 (1969)). This enzyme can also utilize propionyl-CoA instead of acetyl-CoA forming propionate in the process (Hesslinger et al., Mol. Microbiol. 27:477-492 (1998)). Similarly, the ptb gene from C. acetobutylicum encodes an enzyme that can convert butyryl-CoA into butyryl-phosphate (Walter et al., Gene 134:107-111 (1993)); Huang et al., J Mol. Microbiol. Biotechnol. 2:33-38 (2000). Additional ptb genes can be found in butyrate-producing bacterium L2-50 (Louis et al., J. Bacteriol. 186:2099-2106 (2004)) and Bacillus megaterium (Vazquez et al., Curr. Microbiol. 42:345-349 (2001)).
TABLE-US-00114 Gene Accession No. GI No. Organism pta NP_416800.1 16130232 Escherichia coli ptb NP_349676 15896327 Clostridium acetobutylicum ptb AAR19757.1 38425288 butyrate-producing bacterium L2-50 ptb CAC07932.1 10046659 Bacillus megaterium
2.6.1. Aminotransferase.
[0606] Aminotransferases reversibly convert an aldehyde or ketone to an amino group. Common amino donor/acceptor combinations include glutamate/alpha-ketoglutarate, alanine/pyruvate, and aspartate/oxaloacetate. Several enzymes have been shown to convert aldehydes to primary amines, and vice versa, such as 4-aminobutyrate, putrescine, and 5-amino-2-oxopentanoate. These enzymes are particularly well suited to carry out the following transformations: Step N in FIGS. 62 and 63, Steps E and Q in FIG. 63. Lysine-6-aminotransferase (EC 2.6.1.36) is one exemplary enzyme capable of forming a primary amine. This enzyme function, converting lysine to alpha-aminoadipate semialdehyde, has been demonstrated in yeast and bacteria. Candidates from Candida utilis (Hammer and Bode, J. Basic Microbiol. 32:21-27 (1992)), Flavobacterium lutescens (Fujii et al., J. Biochem. 128:391-397 (2000)) and Streptomyces clavuligenus (Romero et al., J. Ind. Microbiol. Biotechnol. 18:241-246 (1997)) have been characterized. A recombinant lysine-6-aminotransferase from S. clavuligenus was functionally expressed in E. coli (Tobin et al., J. Bacteriol. 173:6223-6229 (1991)). The F. lutescens enzyme is specific to alpha-ketoglutarate as the amino acceptor (Soda and Misono, Biochemistry 7:4110-4119 (1968)). Other enzymes which convert aldehydes to terminal amines include the dat gene product in Acinetobacter baumanii encoding 2,4-diaminobutanoate:2-ketoglutarate 4-transaminase (Ikai and Yamamoto, J. Bacteriol. 179:5118-5125 (1997)). In addition to its natural substrate, 2,4-diaminobutyrate, DAT transaminates the terminal amines of lysine, 4-aminobutyrate and ornithine.
TABLE-US-00115 Gene Accession No. GI No. Organism lat BAB13756.1 10336502 Flavobacterium lutescens lat AAA26777.1 153343 Streptomyces clavuligenus dat P56744.1 6685373 Acinetobacter baumanii
[0607] The conversion of an aldehyde to a terminal amine can also be catalyzed by gamma-aminobutyrate transaminase (GABA transaminase or 4-aminobutyrate transaminase). This enzyme naturally interconverts succinic semialdehyde and glutamate to 4-aminobutyrate and alpha-ketoglutarate and is known to have a broad substrate range (Liu et al., Biochemistry 43:10896-10905 2004); Schulz et al., Appl. Environ. Microbiol. 56:1-6 (1990)). The two GABA transaminases in E. coli are encoded by gabT (Bartsch et al., J. Bacteriol. 172:7035-7042 (1990)) and puuE (Kurihara et al., J. Biol. Chem. 280:4602-4608. (2005)). GABA transaminases in Mus musculus, Pseudomonas fluorescens, and Sus scrofa have been shown to react with a range of alternate substrates including 6-aminocaproic acid (Cooper, Methods Enzymol. 113:80-82 (1985); Scott and Jakoby, J. Biol. Chem. 234:932-936 (1959)).
TABLE-US-00116 Gene Accession No. GI No. Organism gabT NP_417148.1 16130576 Escherichia coli puuE NP_415818.1 16129263 Escherichia coli abat NP_766549.2 37202121 Mus musculus gabT YP_257332.1 70733692 Pseudomonas fluorescens abat NP_999428.1 47523600 Sus scrofa
[0608] Additional enzyme candidates for interconverting aldehydes and primary amines are putrescine transminases or other diamine aminotransferases. The E. coli putrescine aminotransferase is encoded by the ygjG gene, and the purified enzyme also was able to transaminate cadaverine and spermidine (Samsonova et al., BMC Microbiol. 3:2 (2003)). In addition, activity of this enzyme on 1,7-diaminoheptane and with amino acceptors other than 2-oxoglutarate (for example, pyruvate, 2-oxobutanoate) has been reported (Kim, J. Biol. Chem. 239:783-786 (1964); Samsonova et al., BMC Microbiol. 3:2 (2003)). A putrescine aminotransferase with higher activity with pyruvate as the amino acceptor than alpha-ketoglutarate is the spuC gene of Pseudomonas aeruginosa (Lu et al., J. Bacteriol. 184:3765-3773 (2002)).
TABLE-US-00117 Gene Accession No. GI No. Organism ygjG NP_417544 145698310 Escherichia coli spuC AAG03688 9946143 Pseudomonas aeruginosa
[0609] Enzymes that transaminate 3-oxoacids include GABA aminotransferase (described above), beta-alanine/alpha-ketoglutarate aminotransferase and 3-amino-2-methylpropionate aminotransferase. Beta-alanine/alpha-ketoglutarate aminotransferase (WO08027742) reacts with beta-alanine to form malonic semialdehyde, a 3-oxoacid. The gene product of SkPYD4 in Saccharomyces kluyveri was shown to preferentially use beta-alanine as the amino group donor (Andersen and Hansen, Gene 124:105-109 (1993)). SkUGA1 encodes a homologue of Saccharomyces cerevisiae GABA aminotransferase, UGA1 (Ramos et al., Eur. J. Biochem. 149:401-404 (1985)), whereas SkPYD4 encodes an enzyme involved in both beta-alanine and GABA transamination (Andersen and Hansen, Gene 124:105-109 (1993)). 3-Amino-2-methylpropionate transaminase catalyzes the transformation from methylmalonate semialdehyde to 3-amino-2-methylpropionate. The enzyme has been characterized in Rattus norvegicus and Sus scrofa and is encoded by Abat (Kakimoto et al., Biochim. Biophys. Acta 156:374-380 (1968); Tamaki et al., Methods Enzymol. 324:376-389 (2000)).
TABLE-US-00118 Gene Accession No. GI No. Organism SkyPYD4 ABF58893.1 98626772 Lachancea kluyveri SkUGA1 ABF58894.1 98626792 Lachancea kluyveri UGA1 NP_011533.1 6321456 Saccharomyces cerevisiae Abat P50554.3 122065191 Rattus norvegicus Abat P80147.2 120968 Sus scrofa
[0610] Several aminotransferases transaminate the amino groups of amino acids to form 2-oxoacids. Aspartate aminotransferase is an enzyme that naturally transfers an oxo group from oxaloacetate to glutamate, forming alpha-ketoglutarate and aspartate. Aspartate is similar in structure to OHED and 2-AHD. Aspartate aminotransferase activity is catalyzed by, for example, the gene products of aspC from Escherichia coli (Yagi et al., FEBS Lett. 100:81-84 (1979); Yagi et al., Methods Enzymol. 113:83-89 (1985)), AAT2 from Saccharomyces cerevisiae (Yagi et al., J. Biochem. 92:35-43 (1982)) and ASPS from Arabidopsis thaliana (de la Torre et al., Plant J. 46:414-425 (2006); Kwok and Hanson. J. Exp. Bot. 55:595-604 (2004); Wilkie and Warren, Protein Expr. Purif. 12:381-389 (1998)). The enzyme from Rattus norvegicus has been shown to transaminate alternate substrates such as 2-aminohexanedioic acid and 2,4-diaminobutyric acid (Recasens et al., Biochemistry 19:4583-4589 (1980)). Aminotransferases that work on other amino-acid substrates can also be able to catalyze this transformation. Valine aminotransferase catalyzes the conversion of valine and pyruvate to 2-ketoisovalerate and alanine. The E. coli gene, avtA, encodes one such enzyme (Whalen and Berg, J. Bacteriol. 150:739-746 (1982)). This gene product also catalyzes the transamination of .alpha.-ketobutyrate to generate .alpha.-aminobutyrate, although the amine donor in this reaction has not been identified (Whalen and Berg, J. Bacteriol. 158:571-574 1984)). The gene product of the E. coli serC catalyzes two reactions, phosphoserine aminotransferase and phosphohydroxythreonine aminotransferase (Lam and Winkler, J. Bacteriol. 172:6518-6528 (1990)), and activity on non-phosphorylated substrates could not be detected (Drewke et al., FEBS Lett. 390:179-182 (1996)).
TABLE-US-00119 Gene Accession No. GI No. Organism aspC NP_415448.1 16128895 Escherichia coli AAT2 P23542.3 1703040 Saccharomyces cerevisiae ASP5 P46248.2 20532373 Arabidopsis thaliana Got2 P00507 112987 Rattus norvegicus avtA YP_026231.1 49176374 Escherichia coli serC NP_415427.1 16128874 Escherichia coli
[0611] Another enzyme candidate is alpha-aminoadipate aminotransferase (EC 2.6.1.39), an enzyme that participates in lysine biosynthesis and degradation in some organisms. This enzyme interconverts 2-aminoadipate and 2-oxoadipate, using alpha-ketoglutarate as the amino acceptor. Gene candidates are found in Homo sapiens (Okuno et al., Enzyme Protein 47:136-148 (1993)) and Thermus thermophilus (Miyazaki et al., Microbiology 150:2327-2334 (2004)). The Thermus thermophilus enzyme, encoded by lysN, is active with several alternate substrates including oxaloacetate, 2-oxoisocaproate, 2-oxoisovalerate, and 2-oxo-3-methylvalerate.
TABLE-US-00120 Gene Accession No. GI No. Organism lysN BAC76939.1 31096548 Thermus thermophilus AadAT-II Q8N5Z0.2 46395904 Homo sapiens
2.7.2.a Phosphotransferase (Carboxy Acceptor).
[0612] Phosphotransferase enzymes in the EC class 2.7.2 transform carboxylic acids to phosphonic acids with concurrent hydrolysis of one ATP. Step 0 of FIG. 62 involves the conversion of 4-hydroxybutyrate to 4-hydroxybutyryl-phosphate by such an enzyme. Butyrate kinase (EC 2.7.2.7) carries out the reversible conversion of butyryl-phosphate to butyrate during acidogenesis in C. acetobutylicum (Cary et al., Appl. Environ. Microbiol. 56:1576-1583 (1990)). This enzyme is encoded by either of the two buk gene products (Huang et al., J. Mol. Microbiol. Biotechnol. 2:33-38 (2000)). Other butyrate kinase enzymes are found in C. butyricum and C. tetanomorphum (Twarog and Wolfe, J. Bacteriol. 86:112-117 (1963)). Related enzyme isobutyrate kinase from Thermotoga maritima has also been expressed in E. coli and crystallized (Diao et al., Acta Crystallogr. D. Biol. Crystallogr. 59:1100-1102 (2003); Diao and Hasson, J. Bacteriol. 191:2521-2529 (2009)). Aspartokinase catalyzes the ATP-dependent phosphorylation of aspartate and participates in the synthesis of several amino acids. The aspartokinase III enzyme in E. coli, encoded by lysC, has a broad substrate range, and the catalytic residues involved in substrate specificity have been elucidated (Keng and Viola, Arch. Biochem. Biophys. 335:73-81 (1996)). Two additional kinases in E. coli are also good candidates: acetate kinase and gamma-glutamyl kinase. The E. coli acetate kinase, encoded by ackA (Skarstedt and Silverstein, J. Biol. Chem. 251:6775-6783 (1976)), phosphorylates propionate in addition to acetate (Hesslinger et al., Mol. Microbiol. 27:477-492 (1998)). The E. coli gamma-glutamyl kinase, encoded by proB (Smith et al., J. Bacteriol. 157:545-551 (1984)), phosphorylates the gamma carbonic acid group of glutamate.
TABLE-US-00121 Gene Accession No. GI No. Organism buk1 NP_349675 15896326 Clostridium acetobutylicum buk2 Q97II1 20137415 Clostridium acetobutylicum buk2 Q9X278.1 6685256 Thermotoga maritima lysC NP_418448.1 16131850 Escherichia coli ackA NP_416799.1 16130231 Escherichia coli proB NP_414777.1 16128228 Escherichia coli
[0613] Acetylglutamate kinase phosphorylates acetylated glutamate during arginine biosynthesis. This enzyme is not known to accept alternate substrates; however, several residues of the E. coli enzyme involved in substrate binding and phosphorylation have been elucidated by site-directed mutagenesis (Marco-Marin et al., J. Mol. Biol. 334:459-476 (2003); Ramon-Maiques et al., Structure 10:329-342 (2002)). The enzyme is encoded by argB in Bacillus subtilis and E. coli (Parsot et al., Gene 68:275-283 (1988)), and ARG5,6 in S. cerevisiae (Pauwels et al., Eur. Biochem. 270:1014-1024 (2003)). The ARG5,6 gene of S. cerevisiae encodes a polyprotein precursor that is matured in the mitochondrial matrix to become acetylglutamate kinase and acetylglutamylphosphate reductase.
TABLE-US-00122 Gene Accession No. GI No. Organism argB NP_418394.3 145698337 Escherichia coli argB NP_389003.1 16078186 Bacillus subtilis ARG5,6 NP_010992.1 6320913 Saccharomyces cerevisiae
2.8.3.a CoA Transferase.
[0614] The gene products of cat1, cat2, and cat3 of Clostridium kluyveri have been shown to exhibit succinyl-CoA (Step G, FIGS. 62 and 63), 4-hydroxybutyryl-CoA (Step T, FIG. 62), and butyryl-CoA acetyltransferase activity, respectively (Seedorf et al., Proc. Natl. Acad. Sci. USA 105:2128-2133 (2008); Sohling and Gottschalk, J. Bacteriol 178:871-880 (1996)). Similar CoA transferase activities are also present in Trichomonas vaginalis (van Grinsven et al., J. Biol. Chem. 283:1411-1418 (2008)) and Trypanosoma brucei (Riviere et al., J Biol. Chem. 279:45337-45346 (2004)).
TABLE-US-00123 Gene Accession No. GI No. Organism cat1 P38946.1 729048 Clostridium kluyveri cat2 P38942.2 1705614 Clostridium kluyveri cat3 EDK35586.1 146349050 Clostridium kluyveri TVAG_395550 XP_001330176 123975034 Trichomonas vaginalis G3 Tb11.02.0290 XP_828352 71754875 Trypanosoma brucei
[0615] An additionally useful enzyme for this type of transformation is acyl-CoA:acetate-CoA transferase, also known as acetate-CoA transferase (EC 2.8.3.8), which has been shown to transfer the CoA moiety to acetate from a variety of branched and linear acyl-CoA substrates, including isobutyrate (Matthies and Schink, Appl. Environ. Microbiol. 58:1435-1439 (1992)), valerate (Vanderwinkel et al., Biochem. Biophys. Res. Commun. 33:902-908 (1968)) and butanoate (Vanderwinkel, supra (1968)). This enzyme is encoded by atoA (alpha subunit) and atoD (beta subunit) in E. coli sp. K12 (Korolev et al., Acta Crystallogr. D Biol. Crystallo. 58:2116-2121 (2002); Vanderwinkel, supra (1968)). Similar enzymes exist in Corynebacterium glutamicum ATCC 13032 (Duncan et al., Appl. Environ. Microbiol. 68:5186-5190 (2002)), Clostridium acetobutylicum (Cary et al., Appl. Environ. Microbiol. 56:1576-1583 (1990); Wiesenborn et al., Appl. Environ. Microbiol. 55:323-329 (1989)), and Clostridium saccharoperbutylacetonicum (Kosaka et al., Biosci. Biotechnol. Biochem. 71:58-68 (2007)).
TABLE-US-00124 Gene Accession No. GI No. Organism atoA P76459.1 2492994 Escherichia coli K12 atoD P76458.1 2492990 Escherichia coli K12 actA YP_226809.1 62391407 Corynebacterium glutamicum cg0592 YP_224801.1 62389399 Corynebacterium glutamicum ctfA NP_149326.1 15004866 Clostridium acetobutylicum ctfB NP_149327.1 15004867 Clostridium acetobutylicum ctfA AAP42564.1 31075384 Clostridium saccharoperbutyl- acetonicum ctfB AAP42565.1 31075385 Clostridium saccharoperbutyl- acetonicum
[0616] The glutaconate-CoA-transferase (EC 2.8.3.12) enzyme from anaerobic bacterium Acidaminococcus fermentans reacts with diacid glutaconyl-CoA and 3-butenoyl-CoA (Mack and Buckel, FEBS Lett. 405:209-212 (1997)). The genes encoding this enzyme are gctA and gctB. This enzyme has reduced but detectable activity with other CoA derivatives including glutaryl-CoA, 2-hydroxyglutaryl-CoA, adipyl-CoA and acrylyl-CoA (Buckel et al., Eur. J. Biochem. 118:315-321 (1981)). The enzyme has been cloned and expressed in E. coli (Mac et al., Eur. J. Biochem. 226:41-51 (1994)).
TABLE-US-00125 Gene Accession No. GI No. Organism gctA CAA57199.1 559392 Acidaminococcus fermentans gctB CAA57200.1 559393 Acidaminococcus fermentans
3.1.2.a CoA Hydrolase.
[0617] Enzymes in the 3.1.2 family hydrolyze acyl-CoA molecules to their corresponding acids. However, such enzymes can be modified to empart CoA-ligase or synthetase functionality if coupled to an energy source such as a proton pump or direct ATP hydrolysis. Several eukaryotic acetyl-CoA hydrolases (EC 3.1.2.1) have broad substrate specificity. For example, the enzyme from Rattus norvegicus brain (Robinson et al., Biochem. Biophys. Res. Commun. 71:959-965 (1976)) can react with butyryl-CoA, hexanoyl-CoA and malonyl-CoA. Though its sequence has not been reported, the enzyme from the mitochondrion of the pea leaf also has a broad substrate specificity, with demonstrated activity on acetyl-CoA, propionyl-CoA, butyryl-CoA, palmitoyl-CoA, oleoyl-CoA, succinyl-CoA, and crotonyl-CoA (Zeiher and Randall, Plant. Physiol. 94:20-27 (1990)). The acetyl-CoA hydrolase, ACH1, from S. cerevisiae represents another candidate hydrolase (Buu et al., J. Biol. Chem. 278:17203-17209 (2003)).
TABLE-US-00126 Gene Accession No. GI No. Organism acot12 NP_570103.1 18543355 Rattus norvegicus ACH1 NP_009538 6319456 Saccharomyces cerevisiae
[0618] Another candidate hydrolase is the human dicarboxylic acid thioesterase, acot8, which exhibits activity on glutaryl-CoA, adipyl-CoA, suberyl-CoA, sebacyl-CoA, and dodecanedioyl-CoA (Westin et al., J. Biol. Chem. 280:38125-38132 (2005)) and the closest E. coli homolog, tesB, which can also hydrolyze a broad range of CoA thioesters (Naggert et al., J. Biol. Chem. 266:11044-11050 (1991)). A similar enzyme has also been characterized in the rat liver (Deana, Biochem. Int. 26:767-773 (1992)). Other potential E. coli thioester hydrolases include the gene products of tesA (Bonner and Bloch, J. Biol. Chem. 247:3123-3133 (1972)), ybgC (Kuznetsova et al., FEMS Microbiol. Rev. 29:263-279 (2005); Zhuang et al., FEBS Lett. 516:161-163 (2002)), paaI (Song et al., J. Biol. Chem. 281:11028-11038 (2006)), and ybdB (Leduc et al., J. Bacteriol. 189:7112-7126 (2007)).
TABLE-US-00127 Gene Accession No. GI No. Organism acot8 CAA15502 3191970 Homo sapiens tesB NP_414986 16128437 Escherichia coli acot8 NP_570112 51036669 Rattus norvegicus tesA NP_415027 16128478 Escherichia coli ybgC NP_415264 16128711 Escherichia coli paaI NP_415914 16129357 Escherichia coli ybdB NP_415129 16128580 Escherichia coli
[0619] Yet another candidate hydrolase is the glutaconate CoA-transferase from Acidaminococcus fermentans. This enzyme was transformed by site-directed mutagenesis into an acyl-CoA hydrolase with activity on glutaryl-CoA, acetyl-CoA and 3-butenoyl-CoA (Mack and Buckel, FEBS Lett. 405:209-212 (1997)). This indicates that the enzymes encoding succinyl-CoA:3-ketoacid-CoA transferases and acetoacetyl-CoA:acetyl-CoA transferases can also serve as candidates for this reaction step but would likely require certain mutations to change their function.
TABLE-US-00128 Gene Accession No. GI No. Organism gctA CAA57199.1 559392 Acidaminococcus fermentans gctB CAA57200.1 559393 Acidaminococcus fermentans
[0620] Additional hydrolase enzymes include 3-hydroxyisobutyryl-CoA hydrolase which has been described to efficiently catalyze the conversion of 3-hydroxyisobutyryl-CoA to 3-hydroxyisobutyrate during valine degradation (Shimomura et al., J. Biol. Chem. 269:14248-14253 (1994)). Genes encoding this enzyme include hibch of Rattus norvegicus (Shimomura et al., supra (1994); Shimomura et al., Methods Enzymol. 324:229-240 (2000)) and Homo sapiens (Shimomura et al., supra (1994). Candidate genes by sequence homology include hibch of Saccharomyces cerevisiae and BC_2292 of Bacillus cereus.
TABLE-US-00129 Gene Accession No. GI No. Organism hibch Q5XIE6.2 146324906 Rattus norvegicus hibch Q6NVY1.2 146324905 Homo sapiens hibch P28817.2 2506374 Saccharomyces cerevisiae BC_2292 AP09256 29895975 Bacillus cereus
4.1.1.a Carboxy-Lyase.
[0621] Decarboxylation of Alpha-Keto Acids.
[0622] Alpha-ketoglutarate decarboxylase (Step B, FIGS. 58, 62 and 63), 5-hydroxy-2-oxopentanoic acid decarboxylase (Step Z, FIG. 62), and 5-amino-2-oxopentanoate decarboxylase (Step R, FIG. 63) all involve the decarboxylation of an alpha-ketoacid. The decarboxylation of keto-acids is catalyzed by a variety of enzymes with varied substrate specificities, including pyruvate decarboxylase (EC 4.1.1.1), benzoylformate decarboxylase (EC 4.1.1.7), alpha-ketoglutarate decarboxylase and branched-chain alpha-ketoacid decarboxylase.
[0623] Pyruvate decarboxylase (PDC), also termed keto-acid decarboxylase, is a key enzyme in alcoholic fermentation, catalyzing the decarboxylation of pyruvate to acetaldehyde. The enzyme from Saccharomyces cerevisiae has a broad substrate range for aliphatic 2-keto acids including 2-ketobutyrate, 2-ketovalerate, 3-hydroxypyruvate and 2-phenylpyruvate (Davie et al., J. Biol. Chem. 267:16601-16606 (1992)). This enzyme has been extensively studied, engineered for altered activity, and functionally expressed in E. coli (Killenberg-Jabs et al., Eur. J. Biochem. 268:1698-1704 (2001); Li and Jordan, Biochemistry 38:10004-10012 (1999); ter Schure et al., Appl. Environ. Microbiol. 64:1303-1307 (1998)). The PDC from Zymomonas mobilus, encoded by pdc, also has a broad substrate range and has been a subject of directed engineering studies to alter the affinity for different substrates (Siegert et al., Protein Eng. Des. Sel. 18:345-357 (2005)). The crystal structure of this enzyme is available (Killenberg-Jabs et al., Eur. J. Biochem. 268:1698-1704 (2001)). Other well-characterized PDC candidates include the enzymes from Acetobacter pasteurians (Chandra et al., Arch. Microbiol. 176:443-451 (2001)) and Kluyveromyces lactis (Krieger et al., Eur. J. Biochem. 269:3256-3263 (2002)).
TABLE-US-00130 Gene Accession No. GI No. Organism pdc P06672.1 118391 Zymomonas mobilus pdc1 P06169 30923172 Saccharomyces cerevisiae pdc AM21208 20385191 Acetobacter pasteurians pdc1 Q12629 52788279 Kluyveromyces lactis
[0624] Like PDC, benzoylformate decarboxylase (EC 4.1.1.7) has a broad substrate range and has been the target of enzyme engineering studies. The enzyme from Pseudomonas putida has been extensively studied and crystal structures of this enzyme are available (Hasson et al., Biochemistry 37:9918-9930 (1998); Polovnikova et al., Biochemistry 42:1820-1830 (2003). Site-directed mutagenesis of two residues in the active site of the Pseudomonas putida enzyme altered the affinity (Km) of naturally and non-naturally occurring substrates (Siegert et al., Protein Eng. Des. Sel. 18:345-357 (2005)). The properties of this enzyme have been further modified by directed engineering (Lingen et al., Protein Eng. 15:585-593 (2002); Lingen et al., Chembiochem. 4:721-726 (2003)). The enzyme from Pseudomonas aeruginosa, encoded by mdlC, has also been characterized experimentally (Barrowman et al., FEMS Microbiol. Lett. 34:57-60 (1986)). Additional gene candidates from Pseudomonas stutzeri, Pseudomonas fluorescens and other organisms can be inferred by sequence homology or identified using a growth selection system developed in Pseudomonas putida (Henning et al., Appl. Environ. Microbiol. 72:7510-7517 (2006)).
TABLE-US-00131 Gene Accession No. GI No. Organism mdlC P20906.2 3915757 Pseudomonas putida mdlC Q9HUR2.1 81539678 Pseudomonas aeruginosa dpgB ABN80423.1 126202187 Pseudomonas stutzeri ilvB-1 YP_260581.1 70730840 Pseudomonas fluorescens
[0625] A third enzyme capable of decarboxylating 2-oxoacids is alpha-ketoglutarate decarboxylase (KGD). The substrate range of this class of enzymes has not been studied to date. The KDC from Mycobacterium tuberculosis (Tian et al., Proc. Natl. Acad. Sci. USA 102:10670-10675 (2005)) has been cloned and functionally expressed. However, it is not an ideal candidate for strain engineering because it is large (.about.130 kD) and GC-rich. KDC enzyme activity has been detected in several species of rhizobia including Bradyrhizobium japonicum and Mesorhizobium loti (Green et al., J. Bacteriol. 182:2838-2844 (2000). Although the KDC-encoding gene(s) have not been isolated in these organisms, the genome sequences are available, and several genes in each genome are annotated as putative KDCs. A KDC from Euglena gracilis has also been characterized, but the gene associated with this activity has not been identified to date (Shigeoka and Nakano, Arch. Biochem. Biophys. 288:22-28 (1991)). The first twenty amino acids starting from the N-terminus were sequenced (MTYKAPVKDVKFLLDKVFKV; SEQ ID NO:45) (Shigeoka and Nakano, Arch. Biochem. Biophys. 288:22-28 (1991)). The gene can be identified by testing candidate genes containing this N-terminal sequence for KDC activity.
TABLE-US-00132 Gene Accession No. GI No. Organism kgd O50463.4 160395583 Mycobacterium tuberculosis kgd NP_767092.1 27375563 Bradyrhizobium japonicum kgd NP_105204.1 13473636 Mesorhizobium loti
[0626] A fourth candidate enzyme for catalyzing this reaction is branched chain alpha-ketoacid decarboxylase (BCKA). This class of enzyme has been shown to act on a variety of compounds varying in chain length from 3 to 6 carbons (Oku and Kaneda, J. Biol. Chem. 263:18386-18396 (1988); Smit et al., B. A., J. E. Hylckama Vlieg, W. J. Engels, L. Meijer, J. T. Wouters, and G. Smit. Identification, cloning, and characterization of a Lactococcus lactis branched-chain alpha-keto acid decarboxylase involved in flavor formation. Appl. Environ. Microbiol. 71:303-311 (2005)). The enzyme in Lactococcus lactis has been characterized on a variety of branched and linear substrates including 2-oxobutanoate, 2-oxohexanoate, 2-oxopentanoate, 3-methyl-2-oxobutanoate, 4-methyl-2-oxobutanoate and isocaproate (Smit et al., Appl. Environ. Microbiol. 71:303-311 (2005)). The enzyme has been structurally characterized (Berg et al., Science 318:1782-1786 (2007)). Sequence alignments between the Lactococcus lactis enzyme and the pyruvate decarboxylase of Zymomonas mobilus indicate that the catalytic and substrate recognition residues are nearly identical (Siegert et al., Protein Eng. Des. Sel. 18:345-357 (2005)), so this enzyme would be a promising candidate for directed engineering. Decarboxylation of alpha-ketoglutarate by a BCKA was detected in Bacillus subtilis; however, this activity was low (5%) relative to activity on other branched-chain substrates (Oku and Kaneda. Biosynthesis of branched-chain fatty acids in Bacillus subtilis. A decarboxylase is essential for branched-chain fatty acid synthetase. J. Biol. Chem. 263:18386-18396 (1988)), and the gene encoding this enzyme has not been identified to date. Additional BCKA gene candidates can be identified by homology to the Lactococcus lactis protein sequence. Many of the high-scoring BLASTp hits to this enzyme are annotated as indolepyruvate decarboxylases (EC 4.1.1.74). Indolepyruvate decarboxylase (IPDA) is an enzyme that catalyzes the decarboxylation of indolepyruvate to indoleacetaldehyde in plants and plant bacteria.
TABLE-US-00133 Gene Accession No. GI No. Organism kdcA AAS49166.1 44921617 Lactococcus lactis
[0627] Recombinant branched chain alpha-keto acid decarboxylase enzymes derived from the E1 subunits of the mitochondrial branched-chain keto acid dehydrogenase complex from Homo sapiens and Bos taurus have been cloned and functionally expressed in E. coli (Davie et al., J. Biol. Chem. 267:16601-16606 1992); Wynn et al., J. Biol. Chem. 267:1881-1887 (1992); Wynn et al., J. Biol. Chem. 267:12400-12403 (1992)). In these studies, the authors found that co-expression of chaperonins GroEL and GroES enhanced the specific activity of the decarboxylase by 500-fold (Wynn et al., J. Biol. Chem. 267:12400-12403 (1992)). These enzymes are composed of two alpha and two beta subunits.
TABLE-US-00134 Gene Accession No. GI No. Organism BCKDHB NP_898871.1 34101272 Homo sapiens BCKDHA NP_000700.1 11386135 Homo sapiens BCKDHB P21839 115502434 Bos taurus BCKDHA P11178 129030 Bos taurus
[0628] Decarboxylation of Alpha-Keto Acids.
[0629] Several ornithine decarboxylase (Step U, FIG. 63) enzymes also exhibit activity on lysine and other similar compounds. Such enzymes are found in Nicotiana glutinosa (Lee and Cho, Biochem. J. 360:657-665 (2001)), Lactobacillus sp. 30a (Guirard and Snell, J. Biol. Chem. 255:5960-5964 (1980)) and Vibrio vulnificus (Lee et al., J. Biol. Chem. 282:27115-27125 (2007)). The enzymes from Lactobacillus sp. 30a (Momany et al., J. Mol. Biol. 252:643-655 (1995)) and V. vulnificus have been crystallized. The V. vulnificus enzyme efficiently catalyzes lysine decarboxylation, and the residues involved in substrate specificity have been elucidated (Lee et al., J. Biol. Chem. 282:27115-27125 (2007)). A similar enzyme has been characterized in Trichomonas vaginalis, but the gene encoding this enzyme is not known (Yarlett et al., Biochem. J. 293 (Pt 2):487-493 (1993)).
TABLE-US-00135 Gene Accession No. GI No. Organism AF323910.1:1 . . . 1299 AAG45222.1 12007488 Nicotiana glutinosa odc1 P43099.2 1169251 Lactobacillus sp. 30a VV2_1235 NP_763142.1 27367615 Vibrio vulnificus
[0630] Glutamate decarboxylase enzymes (Step L, FIGS. 62 and 63) are also well-characterized. Exemplary glutamate decarboxylases can be found in E. coli (De Biase et al., Protein Expr. Purif. 8:430-438 (1996)), S. cerevisiae (Coleman et al., J. Biol. Chem. 276:244-250 (2001)), and Homo sapiens (Bu et al., Proc. Natl. Acad. Sci. USA 89:2115-2119 (1992); Bu and Tobin, Genomics 21:222-228 (1994)).
TABLE-US-00136 Gene Accession No. GI No. Organism GAD1 NP_000808 58331246 Homo sapiens GAD2 NP_001127838 197276620 Homo sapiens gadA NP_417974 16131389 Escherichia coli gadB NP_416010 16129452 Escherichia coli GAD1 NP_013976 6323905 Saccharomyces cerevisiae
[0631] Lysine decarboxylase (EC 4.1.1.18) catalyzes the decarboxylation of lysine to cadaverine. Two isozymes of this enzyme are encoded in the E. coli genome by genes cadA and ldcC. CadA is involved in acid resistance and is subject to positive regulation by the cadC gene product (Lemonnier and Lane, Microbiology 144 (Pt 3):751-760 (1998)). CadC accepts hydroxylysine and S-aminoethylcysteine as alternate substrates, and 2-Aminopimelate and 6-ACA act as competitive inhibitors to this enzyme (Sabo et al., Biochemistry 13:662-670 (1974)). Directed evolution or other enzyme engineering methods can be utilized to increase the activity for this enzyme to decarboxylate 2-aminopimelate. The constitutively expressed ldc gene product is less active than CadA (Lemonnier and Lane, Microbiology 144 (Pt 3):751-760 (1998)). A lysine decarboxylase analogous to CadA was recently identified in Vibrio parahaemolyticus (Tanaka et al., J. Appl. Microbiol. 104:1283-1293 (2008)). The lysine decarboxylase from Selenomonas ruminantium, encoded by ldc, bears sequence similarity to eukaryotic ornithine decarboxylases, and accepts both L-lysine and L-ornithine as substrates (Takatsuka et al., Biosci. Biotechnol. Biochem. 63:1843-1846 (1999)). Active site residues were identified and engineered to alter the substrate specificity of the enzyme (Takatsuka et al., J. Bacteriol. 182:6732-6741 (2000)).
TABLE-US-00137 Gene Accession No. GI No. Organism cadA AAA23536.1 145458 Escherichia coli ldcC AAC73297.1 1786384 Escherichia coli ldc O50657.1 13124043 Selenomonas ruminantium cadA AB124819.1 44886078 Vibrio parahaemolyticus
6.2.1.a CoA Synthetase.
[0632] CoA synthetase or ligase reactions are required by Step I of FIGS. 62 and 63, and Step V of FIG. 62. Succinate or 4-hydroxybutyrate are the required substrates. Exemplary genes encoding enzymes likely to carry out these transformations include the sucCD genes of E. coli, which naturally form a succinyl-CoA synthetase complex. This enzyme complex naturally catalyzes the formation of succinyl-CoA from succinate with the concomitant consumption of one ATP, a reaction which is reversible in vivo (Buck et al., Biochem. 24:6245-6252 (1985)).
TABLE-US-00138 Gene Accession No. GI No. Organism sucC NP_415256.1 16128703 Escherichia coli sucD AAC73823.1 1786949 Escherichia coli
[0633] Additional exemplary CoA-ligases include the rat dicarboxylate-CoA ligase for which the sequence is yet uncharacterized (Vamecq et al., Biochemical J. 230:683-693 (1985)), either of the two characterized phenylacetate-CoA ligases from P. chrysogenum (Lamas-Maceiras et al., Biochem. J. 395:147-155 (2005); Wang et al., Biochem Biophy Res Commun 360(2):453-458 (2007)), the phenylacetate-CoA ligase from Pseudomonas putida (Martinez-Blanco et al., J. Biol. Chem. 265:7084-7090 (1990)), and the 6-carboxyhexanoate-CoA ligase from Bacillus subtilis (Bower et al., J. Bacteriol. 178(14):4122-4130 (1996)). Additional candidate enzymes are acetoacetyl-CoA synthetases from Mus musculus (Hasegawa et al., Biochim. Biophys. Acta 1779:414-419 (2008)) and Homo sapiens (Ohgami et al., Biochem. Pharmacol. 65:989-994 (2003)), which naturally catalyze the ATP-dependant conversion of acetoacetate into acetoacetyl-CoA. 4-Hydroxybutyryl-CoA synthetase activity has been demonstrated in Metallosphaera sedula (Berg et al., Science 318:1782-1786 (2007)). This function has been tentatively assigned to the Msed_1422 gene.
TABLE-US-00139 Gene Accession No. GI No. Organism phl CAJ15517.1 77019264 Penicillium chrysogenum phlB ABS19624.1 152002983 Penicillium chrysogenum paaF AAC24333.2 22711873 Pseudomonas putida bioW NP_390902.2 50812281 Bacillus subtilis AACS NP_084486.1 21313520 Mus musculus AACS NP_076417.2 31982927 Homo sapiens Msed_1422 YP_001191504 146304188 Metallosphaera sedula
[0634] ADP-forming acetyl-CoA synthetase (ACD, EC 6.2.1.13) is another candidate enzyme that couples the conversion of acyl-CoA esters to their corresponding acids with the concurrent synthesis of ATP. Several enzymes with broad substrate specificities have been described in the literature. ACD I from Archaeoglobus fulgidus, encoded by AF1211, was shown to operate on a variety of linear and branched-chain substrates including acetyl-CoA, propionyl-CoA, butyryl-CoA, acetate, propionate, butyrate, isobutyryate, isovalerate, succinate, fumarate, phenylacetate, indoleacetate (Musfeldt et al., J. Bacteriol. 184:636-644 (2002)). The enzyme from Haloarcula marismortui (annotated as a succinyl-CoA synthetase) accepts propionate, butyrate, and branched-chain acids (isovalerate and isobutyrate) as substrates, and was shown to operate in the forward and reverse directions (Brasen et al., Arch. Microbiol. 182:277-287 (2004)). The ACD encoded by PAE3250 from hyperthermophilic crenarchaeon Pyrobaculum aerophilum showed the broadest substrate range of all characterized ACDs, reacting with acetyl-CoA, isobutyryl-CoA (preferred substrate) and phenylacetyl-CoA (Brasen et al., supra (2004)). The enzymes from A. fulgidus, H. marismortui and P. aerophilum have all been cloned, functionally expressed, and characterized in E. coli (Musfeldt et al., supra; Brasen et al., supra (2004)).
TABLE-US-00140 Gene Accession No. GI No. Organism AF1211 NP_070039.1 11498810 Archaeoglobus fulgidus DSM 4304 scs YP_135572.1 55377722 Haloarcula marismortui ATCC 43049 PAE3250 NP_560604.1 18313937 Pyrobaculum aerophilum str. IM2
[0635] Throughout this application various publications have been referenced. The disclosures of these publications in their entireties, including GenBank and GI number publications, are hereby incorporated by reference in this application in order to more fully describe the state of the art to which this invention pertains. Although the invention has been described with reference to the examples provided above, it should be understood that various modifications can be made without departing from the spirit of the invention.
Sequence CWU 1
1
93159DNAArtificial SequenceDescription of Artificial Sequence
Synthetic primer 1gacgaattcg ctagcaagag gagaagtcga catgtccaat
tcactggccg tcgttttac 59247DNAArtificial SequenceDescription of
Artificial Sequence Synthetic primer 2gaccctagga agctttctag
agtcgaccta tgcggcatca gagcaga 47318DNAArtificial
SequenceDescription of Artificial Sequence Synthetic primer
3atgtaccgca agttccgc 18418DNAArtificial SequenceDescription of
Artificial Sequence Synthetic primer 4caatttgccg atgcccag
18520DNAArtificial SequenceDescription of Artificial Sequence
Synthetic primer 5gctgaccact gaagactttg 20619DNAArtificial
SequenceDescription of Artificial Sequence Synthetic primer
6gatcagggct tcggtgtag 19727DNAArtificial SequenceDescription of
Artificial Sequence Synthetic primer 7ttggtgcggg ccaagcagga tctgctc
27830DNAArtificial SequenceDescription of Artificial Sequence
Synthetic primer 8tcagccgaac gcctcgtcga ggatctcctg
30932DNAArtificial SequenceDescription of Artificial Sequence
Synthetic primer 9tggccaacat aagttcacca ttcgggcaaa ac
321029DNAArtificial SequenceDescription of Artificial Sequence
Synthetic primer 10tctcttcaac cagccattcg ttttgcccg
291127DNAClostridium acetobutylicum 11attaaagtta agtggaggaa tgttaac
271222DNAArtificial SequenceDescription of Artificial Sequence
Synthetic primer 12acacgcggat ccaacgtccc gg 221323DNAArtificial
SequenceDescription of Artificial Sequence Synthetic primer
13agcggctccg ctagccgctt atg 231424DNAArtificial SequenceDescription
of Artificial Sequence Synthetic primer 14aagccgttgc tgcagctctt
gagc 241523DNAArtificial SequenceDescription of Artificial Sequence
Synthetic primer 15atctccggcg gtcggatccg tcg 231620DNAArtificial
SequenceDescription of Artificial Sequence Synthetic primer
16aaagcggcta gccacgccgc 201720DNAArtificial SequenceDescription of
Artificial Sequence Synthetic primer 17attacacgag gtacccaacg
201822DNAArtificial SequenceDescription of Artificial Sequence
Synthetic primer 18atgctggcgt acaaaggtgt cc 221922DNAArtificial
SequenceDescription of Artificial Sequence Synthetic primer
19ggacaccttt gtacgccagc at 222025DNAArtificial SequenceDescription
of Artificial Sequence Synthetic primer 20atcgcctaca ctaaaccaga
agtgg 252125DNAArtificial SequenceDescription of Artificial
Sequence Synthetic primer 21ccacttctgg tttagtgtag gcgat
252220DNAArtificial SequenceDescription of Artificial Sequence
Synthetic primer 22aggcagttcc ataggatggc 202355DNAArtificial
SequenceDescription of Artificial Sequence Synthetic primer
23tgacatgtaa cacctacctt ctgtgcctgt gccagtggtt gctgtgatat agaag
552448DNAArtificial SequenceDescription of Artificial Sequence
Synthetic primer 24ataataatac atatgaacca tgcgagttac gggcctataa
gccaggcg 482542DNAArtificial SequenceDescription of Artificial
Sequence Synthetic primer 25agtttttcga tatctgcatc agacaccggc
acattgaaac gg 422660DNAArtificial SequenceDescription of Artificial
Sequence Synthetic primer 26ctggcacagg cacagaaggt aggtgttaca
tgtcagaacg tttacacaat gacgtggatc 602749DNAArtificial
SequenceDescription of Artificial Sequence Synthetic primer
27agacaaatcg gttgccgttt gttaagccag gcgagatatg atctatatc
492854DNAArtificial SequenceDescription of Artificial Sequence
Synthetic primer 28gagttttgat ttcagtactc atcatgtaac acctaccttc
ttgctgtgat atag 542954DNAArtificial SequenceDescription of
Artificial Sequence Synthetic primer 29ctatatcaca gcaagaaggt
aggtgttaca tgatgagtac tgaaatcaaa actc 543049DNAArtificial
SequenceDescription of Artificial Sequence Synthetic primer
30gatatagatc atatctcgcc tggcttaaca aacggcaacc gatttgtct
493170DNAArtificial SequenceDescription of Artificial Sequence
Synthetic oligonucleotide 31tattgtgcat acagatgaat ttttatgcaa
acagtcagcc ctgaagaagg gtgtaggctg 60gagctgcttc 703270DNAArtificial
SequenceDescription of Artificial Sequence Synthetic
oligonucleotide 32caaaaaaccg gagtctgtgc tccggttttt tattatccgc
taatcaatta catatgaata 60tcctccttag 703351DNAArtificial
SequenceDescription of Artificial Sequence Synthetic primer
33ataataatag aattcgtttg ctacctaaat tgccaactaa atcgaaacag g
513447DNAArtificial SequenceDescription of Artificial Sequence
Synthetic primer 34tattattatg gtaccaatat catgcagcaa acggtgcaac
attgccg 473547DNAArtificial SequenceDescription of Artificial
Sequence Synthetic primer 35tgatctggaa gaattcatcg gctttaccac
cgtcaaaaaa aacggcg 473648DNAArtificial SequenceDescription of
Artificial Sequence Synthetic primer 36ataaaaccct gcagcggaaa
cgaagtttta tccatttttg gttacctg 483735DNAArtificial
SequenceDescription of Artificial Sequence Synthetic primer
37ggaagagagg ctggtaccca gaagccacag cagga 353838DNAArtificial
SequenceDescription of Artificial Sequence Synthetic primer
38gtaatcactg cgtaagcgcc atgccccggc gttaattc 383925DNAArtificial
SequenceDescription of Artificial Sequence Synthetic primer
39attgccgcgt tcctcctgct gtcga 254024DNAArtificial
SequenceDescription of Artificial Sequence Synthetic primer
40cgacagcagg aggaacgcgg caat 244175DNAArtificial
SequenceDescription of Artificial Sequence Synthetic primer
41gtttgcacgc tatagctgag gttgttgtct tccagcaacg taccgtatac aataggcgta
60tcacgaggcc ctttc 754270DNAArtificial SequenceDescription of
Artificial Sequence Synthetic primer 42gctacagcat gtcacacgat
ctcaacggtc ggatgaccaa tctggctggt atgggaatta 60gccatggtcc
704373DNAArtificial SequenceDescription of Artificial Sequence
Synthetic oligonucleotide 43tgtgagtgaa agtcacctgc cttaatatct
caaaactcat cttcgggtga cgaaatatgg 60cgtgactcga tac
734470DNAArtificial SequenceDescription of Artificial Sequence
Synthetic oligonucleotide 44tctgtatcag gctgaaaatc ttctctcatc
cgccaaaaca gcttcggcgt taagatgcgc 60gctcaaggac 704520PRTEuglena
gracilis 45Met Thr Tyr Lys Ala Pro Val Lys Asp Val Lys Phe Leu Leu
Asp Lys 1 5 10 15 Val Phe Lys Val 20 462036DNAEscherichia coli
46atgaacttac atgaatatca ggcaaaacaa ctttttgccc gctatggctt accagcaccg
60gtgggttatg cctgtactac tccgcgcgaa gcagaagaag ccgcttcaaa aatcggtgcc
120ggtccgtggg tagtgaaatg tcaggttcac gctggtggcc gcggtaaagc
gggcggtgtg 180aaagttgtaa acagcaaaga agacatccgt gcttttgcag
aaaactggct gggcaagcgt 240ctggtaacgt atcaaacaga tgccaatggc
caaccggtta accagattct ggttgaagca 300gcgaccgata tcgctaaaga
gctgtatctc ggtgccgttg ttgaccgtag ttcccgtcgt 360gtggtcttta
tggcctccac cgaaggcggc gtggaaatcg aaaaagtggc ggaagaaact
420ccgcacctga tccataaagt tgcgcttgat ccgctgactg gcccgatgcc
gtatcaggga 480cgcgagctgg cgttcaaact gggtctggaa ggtaaactgg
ttcagcagtt caccaaaatc 540ttcatgggcc tggcgaccat tttcctggag
cgcgacctgg cgttgatcga aatcaacccg 600ctggtcatca ccaaacaggg
cgatctgatt tgcctcgacg gcaaactggg cgctgacggc 660aacgcactgt
tccgccagcc tgatctgcgc gaaatgcgtg accagtcgca ggaagatccg
720cgtgaagcac aggctgcaca gtgggaactg aactacgttg cgctggacgg
taacatcggt 780tgtatggtta acggcgcagg tctggcgatg ggtacgatgg
acatcgttaa actgcacggc 840ggcgaaccgg ctaacttcct tgacgttggc
ggcggcgcaa ccaaagaacg tgtaaccgaa 900gcgttcaaaa tcatcctctc
tgacgacaaa gtgaaagccg ttctggttaa catcttcggc 960ggtatcgttc
gttgcgacct gatcgctgac ggtatcatcg gcgcggtagc agaagtgggt
1020gttaacgtac cggtcgtggt acgtctggaa ggtaacaacg ccgaactcgg
cgcgaagaaa 1080ctggctgaca gcggcctgaa tattattgca gcaaaaggtc
tgacggatgc agctcagcag 1140gttgttgccg cagtggaggg gaaataatgt
ccattttaat cgataaaaac accaaggtta 1200tctgccaggg ctttaccggt
agccagggga ctttccactc agaacaggcc attgcatacg 1260gcactaaaat
ggttggcggc gtaaccccag gtaaaggcgg caccacccac ctcggcctgc
1320cggtgttcaa caccgtgcgt gaagccgttg ctgccactgg cgctaccgct
tctgttatct 1380acgtaccagc accgttctgc aaagactcca ttctggaagc
catcgacgca ggcatcaaac 1440tgattatcac catcactgaa ggcatcccga
cgctggatat gctgaccgtg aaagtgaagc 1500tggatgaagc aggcgttcgt
atgatcggcc cgaactgccc aggcgttatc actccgggtg 1560aatgcaaaat
cggtatccag cctggtcaca ttcacaaacc gggtaaagtg ggtatcgttt
1620cccgttccgg tacactgacc tatgaagcgg ttaaacagac cacggattac
ggtttcggtc 1680agtcgacctg tgtcggtatc ggcggtgacc cgatcccggg
ctctaacttt atcgacattc 1740tcgaaatgtt cgaaaaagat ccgcagaccg
aagcgatcgt gatgatcggt gagatcggcg 1800gtagcgctga agaagaagca
gctgcgtaca tcaaagagca cgttaccaag ccagttgtgg 1860gttacatcgc
tggtgtgact gcgccgaaag gcaaacgtat gggccacgcg ggtgccatca
1920ttgccggtgg gaaagggact gcggatgaga aattcgctgc tctggaagcc
gcaggcgtga 1980aaaccgttcg cagcctggcg gatatcggtg aagcactgaa
aactgttctg aaataa 203647388PRTEscherichia coli 47Met Asn Leu His
Glu Tyr Gln Ala Lys Gln Leu Phe Ala Arg Tyr Gly 1 5 10 15 Leu Pro
Ala Pro Val Gly Tyr Ala Cys Thr Thr Pro Arg Glu Ala Glu 20 25 30
Glu Ala Ala Ser Lys Ile Gly Ala Gly Pro Trp Val Val Lys Cys Gln 35
40 45 Val His Ala Gly Gly Arg Gly Lys Ala Gly Gly Val Lys Val Val
Asn 50 55 60 Ser Lys Glu Asp Ile Arg Ala Phe Ala Glu Asn Trp Leu
Gly Lys Arg 65 70 75 80 Leu Val Thr Tyr Gln Thr Asp Ala Asn Gly Gln
Pro Val Asn Gln Ile 85 90 95 Leu Val Glu Ala Ala Thr Asp Ile Ala
Lys Glu Leu Tyr Leu Gly Ala 100 105 110 Val Val Asp Arg Ser Ser Arg
Arg Val Val Phe Met Ala Ser Thr Glu 115 120 125 Gly Gly Val Glu Ile
Glu Lys Val Ala Glu Glu Thr Pro His Leu Ile 130 135 140 His Lys Val
Ala Leu Asp Pro Leu Thr Gly Pro Met Pro Tyr Gln Gly 145 150 155 160
Arg Glu Leu Ala Phe Lys Leu Gly Leu Glu Gly Lys Leu Val Gln Gln 165
170 175 Phe Thr Lys Ile Phe Met Gly Leu Ala Thr Ile Phe Leu Glu Arg
Asp 180 185 190 Leu Ala Leu Ile Glu Ile Asn Pro Leu Val Ile Thr Lys
Gln Gly Asp 195 200 205 Leu Ile Cys Leu Asp Gly Lys Leu Gly Ala Asp
Gly Asn Ala Leu Phe 210 215 220 Arg Gln Pro Asp Leu Arg Glu Met Arg
Asp Gln Ser Gln Glu Asp Pro 225 230 235 240 Arg Glu Ala Gln Ala Ala
Gln Trp Glu Leu Asn Tyr Val Ala Leu Asp 245 250 255 Gly Asn Ile Gly
Cys Met Val Asn Gly Ala Gly Leu Ala Met Gly Thr 260 265 270 Met Asp
Ile Val Lys Leu His Gly Gly Glu Pro Ala Asn Phe Leu Asp 275 280 285
Val Gly Gly Gly Ala Thr Lys Glu Arg Val Thr Glu Ala Phe Lys Ile 290
295 300 Ile Leu Ser Asp Asp Lys Val Lys Ala Val Leu Val Asn Ile Phe
Gly 305 310 315 320 Gly Ile Val Arg Cys Asp Leu Ile Ala Asp Gly Ile
Ile Gly Ala Val 325 330 335 Ala Glu Val Gly Val Asn Val Pro Val Val
Val Arg Leu Glu Gly Asn 340 345 350 Asn Ala Glu Leu Gly Ala Lys Lys
Leu Ala Asp Ser Gly Leu Asn Ile 355 360 365 Ile Ala Ala Lys Gly Leu
Thr Asp Ala Ala Gln Gln Val Val Ala Ala 370 375 380 Val Glu Gly Lys
385 48289PRTEscherichia coli 48Met Ser Ile Leu Ile Asp Lys Asn Thr
Lys Val Ile Cys Gln Gly Phe 1 5 10 15 Thr Gly Ser Gln Gly Thr Phe
His Ser Glu Gln Ala Ile Ala Tyr Gly 20 25 30 Thr Lys Met Val Gly
Gly Val Thr Pro Gly Lys Gly Gly Thr Thr His 35 40 45 Leu Gly Leu
Pro Val Phe Asn Thr Val Arg Glu Ala Val Ala Ala Thr 50 55 60 Gly
Ala Thr Ala Ser Val Ile Tyr Val Pro Ala Pro Phe Cys Lys Asp 65 70
75 80 Ser Ile Leu Glu Ala Ile Asp Ala Gly Ile Lys Leu Ile Ile Thr
Ile 85 90 95 Thr Glu Gly Ile Pro Thr Leu Asp Met Leu Thr Val Lys
Val Lys Leu 100 105 110 Asp Glu Ala Gly Val Arg Met Ile Gly Pro Asn
Cys Pro Gly Val Ile 115 120 125 Thr Pro Gly Glu Cys Lys Ile Gly Ile
Gln Pro Gly His Ile His Lys 130 135 140 Pro Gly Lys Val Gly Ile Val
Ser Arg Ser Gly Thr Leu Thr Tyr Glu 145 150 155 160 Ala Val Lys Gln
Thr Thr Asp Tyr Gly Phe Gly Gln Ser Thr Cys Val 165 170 175 Gly Ile
Gly Gly Asp Pro Ile Pro Gly Ser Asn Phe Ile Asp Ile Leu 180 185 190
Glu Met Phe Glu Lys Asp Pro Gln Thr Glu Ala Ile Val Met Ile Gly 195
200 205 Glu Ile Gly Gly Ser Ala Glu Glu Glu Ala Ala Ala Tyr Ile Lys
Glu 210 215 220 His Val Thr Lys Pro Val Val Gly Tyr Ile Ala Gly Val
Thr Ala Pro 225 230 235 240 Lys Gly Lys Arg Met Gly His Ala Gly Ala
Ile Ile Ala Gly Gly Lys 245 250 255 Gly Thr Ala Asp Glu Lys Phe Ala
Ala Leu Glu Ala Ala Gly Val Lys 260 265 270 Thr Val Arg Ser Leu Ala
Asp Ile Gly Glu Ala Leu Lys Thr Val Leu 275 280 285 Lys
493696DNAMycobacterium bovis 49atggccaaca taagttcacc attcgggcaa
aacgaatggc tggttgaaga gatgtaccgc 60aagttccgcg acgacccctc ctcggtcgat
cccagctggc acgagttcct ggttgactac 120agccccgaac ccacctccca
accagctgcc gaaccaaccc gggttacctc gccactcgtt 180gccgagcggg
ccgctgcggc cgccccgcag gcacccccca agccggccga caccgcggcc
240gcgggcaacg gcgtggtcgc cgcactggcc gccaaaactg ccgttccccc
gccagccgaa 300ggtgacgagg tagcggtgct gcgcggcgcc gccgcggccg
tcgtcaagaa catgtccgcg 360tcgttggagg tgccgacggc gaccagcgtc
cgggcggtcc cggccaagct actgatcgac 420aaccggatcg tcatcaacaa
ccagttgaag cggacccgcg gcggcaagat ctcgttcacg 480catttgctgg
gctacgccct ggtgcaggcg gtgaagaaat tcccgaacat gaaccggcac
540tacaccgaag tcgacggcaa gcccaccgcg gtcacgccgg cgcacaccaa
tctcggcctg 600gcgatcgacc tgcaaggcaa ggacgggaag cgttccctgg
tggtggccgg catcaagcgg 660tgcgagacca tgcgattcgc gcagttcgtc
acggcctacg aagacatcgt acgccgggcc 720cgcgacggca agctgaccac
tgaagacttt gccggcgtga cgatttcgct gaccaatccc 780ggaaccatcg
gcaccgtgca ttcggtgccg cggctgatgc ccggccaggg cgccatcatc
840ggcgtgggcg ccatggaata ccccgccgag tttcaaggcg ccagcgagga
acgcatcgcc 900gagctgggca tcggcaaatt gatcactttg acctccacct
acgaccaccg catcatccag 960ggcgcggaat cgggcgactt cctgcgcacc
atccacgagt tgctgctctc ggatggcttc 1020tgggacgagg tcttccgcga
actgagcatc ccatatctgc cggtgcgctg gagcaccgac 1080aaccccgact
cgatcgtcga caagaacgct cgcgtcatga acttgatcgc ggcctaccgc
1140aaccgcggcc atctgatggc cgataccgac ccgctgcggt tggacaaagc
tcggttccgc 1200agtcaccccg acctcgaagt gctgacccac ggcctgacgc
tgtgggatct cgatcgggtg 1260ttcaaggtcg acggctttgc cggtgcgcag
tacaagaaac tgcgcgacgt gctgggcttg 1320ctgcgcgatg cctactgccg
ccacatcggc gtggagtacg cccatatcct cgaccccgaa 1380caaaaggagt
ggctcgaaca acgggtcgag accaagcacg tcaaacccac tgtggcccaa
1440cagaaataca tcctcagcaa gctcaacgcc gccgaggcct ttgaaacgtt
cctacagacc 1500aagtacgtcg gccagaagcg gttctcgctg gaaggcgccg
aaagcgtgat cccgatgatg 1560gacgcggcga tcgaccagtg cgctgagcac
ggcctcgacg aggtggtcat cgggatgccg 1620caccggggcc ggctcaacgt
gctggccaac atcgtcggca agccgtactc gcagatcttc 1680accgagttcg
agggcaacct gaatccgtcg caggcgcacg gctccggtga cgtcaagtac
1740cacctgggcg ccaccgggct gtacctgcag atgttcggcg acaacgacat
tcaggtgtcg 1800ctgaccgcca acccgtcgca tctggaggcc gtcgacccgg
tgctggaggg attggtgcgg 1860gccaagcagg atctgctcga ccacggaagc
atcgacagcg acggccaacg ggcgttctcg 1920gtggtgccgc tgatgttgca
tggcgatgcc gcgttcgccg gtcagggtgt ggtcgccgag 1980acgctgaacc
tggcgaatct gccgggctac cgcgtcggcg gcaccatcca catcatcgtc
2040aacaaccaga
tcggcttcac caccgcgccc gagtattcca ggtccagcga gtactgcacc
2100gacgtcgcaa agatgatcgg ggcaccgatc tttcacgtca acggcgacga
cccggaggcg 2160tgtgtctggg tggcgcggtt ggcggtggac ttccgacaac
ggttcaagaa ggacgtcgtc 2220atcgacatgc tgtgctaccg ccgccgcggg
cacaacgagg gtgacgaccc gtcgatgacc 2280aacccctaca tgtacgacgt
cgtcgacacc aagcgcgggg cccgcaaaag ctacaccgaa 2340gccctgatcg
gacgtggcga catctcgatg aaggaggccg aggacgcgct gcgcgactac
2400cagggccagc tggaacgggt gttcaacgaa gtgcgcgagc tggagaagca
cggtgtgcag 2460ccgagcgagt cggtcgagtc cgaccagatg attcccgcgg
ggctggccac tgcggtggac 2520aagtcgctgc tggcccggat cggcgatgcg
ttcctcgcct tgccgaacgg cttcaccgcg 2580cacccgcgag tccaaccggt
gctggagaag cgccgggaga tggcctatga aggcaagatc 2640gactgggcct
ttggcgagct gctggcgctg ggctcgctgg tggccgaagg caagctggtg
2700cgcttgtcgg ggcaggacag ccgccgcggc accttctccc agcggcattc
ggttctcatc 2760gaccgccaca ctggcgagga gttcacacca ctgcagctgc
tggcgaccaa ctccgacggc 2820agcccgaccg gcggaaagtt cctggtctac
gactcgccac tgtcggagta cgccgccgtc 2880ggcttcgagt acggctacac
tgtgggcaat ccggacgccg tggtgctctg ggaggcgcag 2940ttcggcgact
tcgtcaacgg cgcacagtcg atcatcgacg agttcatcag ctccggtgag
3000gccaagtggg gccaattgtc caacgtcgtg ctgctgttac cgcacgggca
cgaggggcag 3060ggacccgacc acacttctgc ccggatcgaa cgcttcttgc
agttgtgggc ggaaggttcg 3120atgaccatcg cgatgccgtc gactccgtcg
aactacttcc acctgctacg ccggcatgcc 3180ctggacggca tccaacgccc
gctgatcgtg ttcacgccca agtcgatgtt gcgtcacaag 3240gccgccgtca
gcgaaatcaa ggacttcacc gagatcaagt tccgctcagt gctggaggaa
3300cccacctatg aggacggcat cggagaccgc aacaaggtca gccggatcct
gctgaccagt 3360ggcaagctgt attacgagct ggccgcccgc aaggccaagg
acaaccgcaa tgacctcgcg 3420atcgtgcggc ttgaacagct cgccccgctg
cccaggcgtc gactgcgtga aacgctggac 3480cgctacgaga acgtcaagga
gttcttctgg gtccaagagg aaccggccaa ccagggtgcg 3540tggccgcgat
tcgggctcga actacccgag ctgctgcctg acaagttggc cgggatcaag
3600cgaatctcgc gccgggcgat gtcagccccg tcgtcaggct cgtcgaaggt
gcacgccgtc 3660gaacagcagg agatcctcga cgaggcgttc ggctaa
3696501231PRTMycobacterium bovis 50Met Ala Asn Ile Ser Ser Pro Phe
Gly Gln Asn Glu Trp Leu Val Glu 1 5 10 15 Glu Met Tyr Arg Lys Phe
Arg Asp Asp Pro Ser Ser Val Asp Pro Ser 20 25 30 Trp His Glu Phe
Leu Val Asp Tyr Ser Pro Glu Pro Thr Ser Gln Pro 35 40 45 Ala Ala
Glu Pro Thr Arg Val Thr Ser Pro Leu Val Ala Glu Arg Ala 50 55 60
Ala Ala Ala Ala Pro Gln Ala Pro Pro Lys Pro Ala Asp Thr Ala Ala 65
70 75 80 Ala Gly Asn Gly Val Val Ala Ala Leu Ala Ala Lys Thr Ala
Val Pro 85 90 95 Pro Pro Ala Glu Gly Asp Glu Val Ala Val Leu Arg
Gly Ala Ala Ala 100 105 110 Ala Val Val Lys Asn Met Ser Ala Ser Leu
Glu Val Pro Thr Ala Thr 115 120 125 Ser Val Arg Ala Val Pro Ala Lys
Leu Leu Ile Asp Asn Arg Ile Val 130 135 140 Ile Asn Asn Gln Leu Lys
Arg Thr Arg Gly Gly Lys Ile Ser Phe Thr 145 150 155 160 His Leu Leu
Gly Tyr Ala Leu Val Gln Ala Val Lys Lys Phe Pro Asn 165 170 175 Met
Asn Arg His Tyr Thr Glu Val Asp Gly Lys Pro Thr Ala Val Thr 180 185
190 Pro Ala His Thr Asn Leu Gly Leu Ala Ile Asp Leu Gln Gly Lys Asp
195 200 205 Gly Lys Arg Ser Leu Val Val Ala Gly Ile Lys Arg Cys Glu
Thr Met 210 215 220 Arg Phe Ala Gln Phe Val Thr Ala Tyr Glu Asp Ile
Val Arg Arg Ala 225 230 235 240 Arg Asp Gly Lys Leu Thr Thr Glu Asp
Phe Ala Gly Val Thr Ile Ser 245 250 255 Leu Thr Asn Pro Gly Thr Ile
Gly Thr Val His Ser Val Pro Arg Leu 260 265 270 Met Pro Gly Gln Gly
Ala Ile Ile Gly Val Gly Ala Met Glu Tyr Pro 275 280 285 Ala Glu Phe
Gln Gly Ala Ser Glu Glu Arg Ile Ala Glu Leu Gly Ile 290 295 300 Gly
Lys Leu Ile Thr Leu Thr Ser Thr Tyr Asp His Arg Ile Ile Gln 305 310
315 320 Gly Ala Glu Ser Gly Asp Phe Leu Arg Thr Ile His Glu Leu Leu
Leu 325 330 335 Ser Asp Gly Phe Trp Asp Glu Val Phe Arg Glu Leu Ser
Ile Pro Tyr 340 345 350 Leu Pro Val Arg Trp Ser Thr Asp Asn Pro Asp
Ser Ile Val Asp Lys 355 360 365 Asn Ala Arg Val Met Asn Leu Ile Ala
Ala Tyr Arg Asn Arg Gly His 370 375 380 Leu Met Ala Asp Thr Asp Pro
Leu Arg Leu Asp Lys Ala Arg Phe Arg 385 390 395 400 Ser His Pro Asp
Leu Glu Val Leu Thr His Gly Leu Thr Leu Trp Asp 405 410 415 Leu Asp
Arg Val Phe Lys Val Asp Gly Phe Ala Gly Ala Gln Tyr Lys 420 425 430
Lys Leu Arg Asp Val Leu Gly Leu Leu Arg Asp Ala Tyr Cys Arg His 435
440 445 Ile Gly Val Glu Tyr Ala His Ile Leu Asp Pro Glu Gln Lys Glu
Trp 450 455 460 Leu Glu Gln Arg Val Glu Thr Lys His Val Lys Pro Thr
Val Ala Gln 465 470 475 480 Gln Lys Tyr Ile Leu Ser Lys Leu Asn Ala
Ala Glu Ala Phe Glu Thr 485 490 495 Phe Leu Gln Thr Lys Tyr Val Gly
Gln Lys Arg Phe Ser Leu Glu Gly 500 505 510 Ala Glu Ser Val Ile Pro
Met Met Asp Ala Ala Ile Asp Gln Cys Ala 515 520 525 Glu His Gly Leu
Asp Glu Val Val Ile Gly Met Pro His Arg Gly Arg 530 535 540 Leu Asn
Val Leu Ala Asn Ile Val Gly Lys Pro Tyr Ser Gln Ile Phe 545 550 555
560 Thr Glu Phe Glu Gly Asn Leu Asn Pro Ser Gln Ala His Gly Ser Gly
565 570 575 Asp Val Lys Tyr His Leu Gly Ala Thr Gly Leu Tyr Leu Gln
Met Phe 580 585 590 Gly Asp Asn Asp Ile Gln Val Ser Leu Thr Ala Asn
Pro Ser His Leu 595 600 605 Glu Ala Val Asp Pro Val Leu Glu Gly Leu
Val Arg Ala Lys Gln Asp 610 615 620 Leu Leu Asp His Gly Ser Ile Asp
Ser Asp Gly Gln Arg Ala Phe Ser 625 630 635 640 Val Val Pro Leu Met
Leu His Gly Asp Ala Ala Phe Ala Gly Gln Gly 645 650 655 Val Val Ala
Glu Thr Leu Asn Leu Ala Asn Leu Pro Gly Tyr Arg Val 660 665 670 Gly
Gly Thr Ile His Ile Ile Val Asn Asn Gln Ile Gly Phe Thr Thr 675 680
685 Ala Pro Glu Tyr Ser Arg Ser Ser Glu Tyr Cys Thr Asp Val Ala Lys
690 695 700 Met Ile Gly Ala Pro Ile Phe His Val Asn Gly Asp Asp Pro
Glu Ala 705 710 715 720 Cys Val Trp Val Ala Arg Leu Ala Val Asp Phe
Arg Gln Arg Phe Lys 725 730 735 Lys Asp Val Val Ile Asp Met Leu Cys
Tyr Arg Arg Arg Gly His Asn 740 745 750 Glu Gly Asp Asp Pro Ser Met
Thr Asn Pro Tyr Met Tyr Asp Val Val 755 760 765 Asp Thr Lys Arg Gly
Ala Arg Lys Ser Tyr Thr Glu Ala Leu Ile Gly 770 775 780 Arg Gly Asp
Ile Ser Met Lys Glu Ala Glu Asp Ala Leu Arg Asp Tyr 785 790 795 800
Gln Gly Gln Leu Glu Arg Val Phe Asn Glu Val Arg Glu Leu Glu Lys 805
810 815 His Gly Val Gln Pro Ser Glu Ser Val Glu Ser Asp Gln Met Ile
Pro 820 825 830 Ala Gly Leu Ala Thr Ala Val Asp Lys Ser Leu Leu Ala
Arg Ile Gly 835 840 845 Asp Ala Phe Leu Ala Leu Pro Asn Gly Phe Thr
Ala His Pro Arg Val 850 855 860 Gln Pro Val Leu Glu Lys Arg Arg Glu
Met Ala Tyr Glu Gly Lys Ile 865 870 875 880 Asp Trp Ala Phe Gly Glu
Leu Leu Ala Leu Gly Ser Leu Val Ala Glu 885 890 895 Gly Lys Leu Val
Arg Leu Ser Gly Gln Asp Ser Arg Arg Gly Thr Phe 900 905 910 Ser Gln
Arg His Ser Val Leu Ile Asp Arg His Thr Gly Glu Glu Phe 915 920 925
Thr Pro Leu Gln Leu Leu Ala Thr Asn Ser Asp Gly Ser Pro Thr Gly 930
935 940 Gly Lys Phe Leu Val Tyr Asp Ser Pro Leu Ser Glu Tyr Ala Ala
Val 945 950 955 960 Gly Phe Glu Tyr Gly Tyr Thr Val Gly Asn Pro Asp
Ala Val Val Leu 965 970 975 Trp Glu Ala Gln Phe Gly Asp Phe Val Asn
Gly Ala Gln Ser Ile Ile 980 985 990 Asp Glu Phe Ile Ser Ser Gly Glu
Ala Lys Trp Gly Gln Leu Ser Asn 995 1000 1005 Val Val Leu Leu Leu
Pro His Gly His Glu Gly Gln Gly Pro Asp 1010 1015 1020 His Thr Ser
Ala Arg Ile Glu Arg Phe Leu Gln Leu Trp Ala Glu 1025 1030 1035 Gly
Ser Met Thr Ile Ala Met Pro Ser Thr Pro Ser Asn Tyr Phe 1040 1045
1050 His Leu Leu Arg Arg His Ala Leu Asp Gly Ile Gln Arg Pro Leu
1055 1060 1065 Ile Val Phe Thr Pro Lys Ser Met Leu Arg His Lys Ala
Ala Val 1070 1075 1080 Ser Glu Ile Lys Asp Phe Thr Glu Ile Lys Phe
Arg Ser Val Leu 1085 1090 1095 Glu Glu Pro Thr Tyr Glu Asp Gly Ile
Gly Asp Arg Asn Lys Val 1100 1105 1110 Ser Arg Ile Leu Leu Thr Ser
Gly Lys Leu Tyr Tyr Glu Leu Ala 1115 1120 1125 Ala Arg Lys Ala Lys
Asp Asn Arg Asn Asp Leu Ala Ile Val Arg 1130 1135 1140 Leu Glu Gln
Leu Ala Pro Leu Pro Arg Arg Arg Leu Arg Glu Thr 1145 1150 1155 Leu
Asp Arg Tyr Glu Asn Val Lys Glu Phe Phe Trp Val Gln Glu 1160 1165
1170 Glu Pro Ala Asn Gln Gly Ala Trp Pro Arg Phe Gly Leu Glu Leu
1175 1180 1185 Pro Glu Leu Leu Pro Asp Lys Leu Ala Gly Ile Lys Arg
Ile Ser 1190 1195 1200 Arg Arg Ala Met Ser Ala Pro Ser Ser Gly Ser
Ser Lys Val His 1205 1210 1215 Ala Val Glu Gln Gln Glu Ile Leu Asp
Glu Ala Phe Gly 1220 1225 1230 511356DNAPorphyromonas gingivalis
51atggaaatca aagaaatggt gagccttgca cgcaaggctc agaaggagta tcaagctacc
60cataaccaag aagcagttga caacatttgc cgagctgcag caaaagttat ttatgaaaat
120gcagctattc tggctcgcga agcagtagac gaaaccggca tgggcgttta
cgaacacaaa 180gtggccaaga atcaaggcaa atccaaaggt gtttggtaca
acctccacaa taaaaaatcg 240attggtatcc tcaatataga cgagcgtacc
ggtatgatcg agattgcaaa gcctatcgga 300gttgtaggag ccgtaacgcc
gacgaccaac ccgatcgtta ctccgatgag caatatcatc 360tttgctctta
agacctgcaa tgccatcatt attgcccccc accccagatc caaaaaatgc
420tctgcacacg cagttcgtct gatcaaagaa gctatcgctc cgttcaacgt
accggaaggt 480atggttcaga tcatcgaaga acccagcatc gagaagacgc
aggaactcat gggcgccgta 540gacgtagtag ttgctacggg tggtatgggc
atggtgaagt ctgcatattc ttcaggaaag 600ccttctttcg gtgttggagc
cggtaacgtt caggtgatcg tggatagcaa catcgatttc 660gaagctgctg
cagaaaaaat catcaccggt cgtgctttcg acaacggtat catctgctca
720ggcgaacaga gcatcatcta caacgaggct gacaaggaag cagttttcac
agcattccgc 780aaccacggtg catatttctg tgacgaagcc gaaggagatc
gggctcgtgc agctatcttc 840gaaaatggag ccatcgcgaa agatgtagta
ggtcagagcg ttgccttcat tgccaagaaa 900gcaaacatca atatccccga
gggtacccgt attctcgttg ttgaagctcg cggcgtagga 960gcagaagacg
ttatctgtaa ggaaaagatg tgtcccgtaa tgtgcgccct cagctacaag
1020cacttcgaag aaggtgtaga aatcgcacgt acgaacctcg ccaacgaagg
taacggccac 1080acctgtgcta tccactccaa caatcaggca cacatcatcc
tcgcaggatc agagctgacg 1140gtatctcgta tcgtagtgaa tgctccgagt
gccactacag caggcggtca catccaaaac 1200ggtcttgccg taaccaatac
gctcggatgc ggatcatggg gtaataactc tatctccgag 1260aacttcactt
acaagcacct cctcaacatt tcacgcatcg caccgttgaa ttcaagcatt
1320cacatccccg atgacaaaga aatctgggaa ctctaa
135652451PRTPorphyromonas gingivalis 52Met Glu Ile Lys Glu Met Val
Ser Leu Ala Arg Lys Ala Gln Lys Glu 1 5 10 15 Tyr Gln Ala Thr His
Asn Gln Glu Ala Val Asp Asn Ile Cys Arg Ala 20 25 30 Ala Ala Lys
Val Ile Tyr Glu Asn Ala Ala Ile Leu Ala Arg Glu Ala 35 40 45 Val
Asp Glu Thr Gly Met Gly Val Tyr Glu His Lys Val Ala Lys Asn 50 55
60 Gln Gly Lys Ser Lys Gly Val Trp Tyr Asn Leu His Asn Lys Lys Ser
65 70 75 80 Ile Gly Ile Leu Asn Ile Asp Glu Arg Thr Gly Met Ile Glu
Ile Ala 85 90 95 Lys Pro Ile Gly Val Val Gly Ala Val Thr Pro Thr
Thr Asn Pro Ile 100 105 110 Val Thr Pro Met Ser Asn Ile Ile Phe Ala
Leu Lys Thr Cys Asn Ala 115 120 125 Ile Ile Ile Ala Pro His Pro Arg
Ser Lys Lys Cys Ser Ala His Ala 130 135 140 Val Arg Leu Ile Lys Glu
Ala Ile Ala Pro Phe Asn Val Pro Glu Gly 145 150 155 160 Met Val Gln
Ile Ile Glu Glu Pro Ser Ile Glu Lys Thr Gln Glu Leu 165 170 175 Met
Gly Ala Val Asp Val Val Val Ala Thr Gly Gly Met Gly Met Val 180 185
190 Lys Ser Ala Tyr Ser Ser Gly Lys Pro Ser Phe Gly Val Gly Ala Gly
195 200 205 Asn Val Gln Val Ile Val Asp Ser Asn Ile Asp Phe Glu Ala
Ala Ala 210 215 220 Glu Lys Ile Ile Thr Gly Arg Ala Phe Asp Asn Gly
Ile Ile Cys Ser 225 230 235 240 Gly Glu Gln Ser Ile Ile Tyr Asn Glu
Ala Asp Lys Glu Ala Val Phe 245 250 255 Thr Ala Phe Arg Asn His Gly
Ala Tyr Phe Cys Asp Glu Ala Glu Gly 260 265 270 Asp Arg Ala Arg Ala
Ala Ile Phe Glu Asn Gly Ala Ile Ala Lys Asp 275 280 285 Val Val Gly
Gln Ser Val Ala Phe Ile Ala Lys Lys Ala Asn Ile Asn 290 295 300 Ile
Pro Glu Gly Thr Arg Ile Leu Val Val Glu Ala Arg Gly Val Gly 305 310
315 320 Ala Glu Asp Val Ile Cys Lys Glu Lys Met Cys Pro Val Met Cys
Ala 325 330 335 Leu Ser Tyr Lys His Phe Glu Glu Gly Val Glu Ile Ala
Arg Thr Asn 340 345 350 Leu Ala Asn Glu Gly Asn Gly His Thr Cys Ala
Ile His Ser Asn Asn 355 360 365 Gln Ala His Ile Ile Leu Ala Gly Ser
Glu Leu Thr Val Ser Arg Ile 370 375 380 Val Val Asn Ala Pro Ser Ala
Thr Thr Ala Gly Gly His Ile Gln Asn 385 390 395 400 Gly Leu Ala Val
Thr Asn Thr Leu Gly Cys Gly Ser Trp Gly Asn Asn 405 410 415 Ser Ile
Ser Glu Asn Phe Thr Tyr Lys His Leu Leu Asn Ile Ser Arg 420 425 430
Ile Ala Pro Leu Asn Ser Ser Ile His Ile Pro Asp Asp Lys Glu Ile 435
440 445 Trp Glu Leu 450 531116DNAPorphyromonas gingivalis
53atgcaacttt tcaaactcaa gagtgtaaca catcactttg acacttttgc agaatttgcc
60aaggaattct gtcttggaga acgcgacttg gtaattacca acgagttcat ctatgaaccg
120tatatgaagg catgccagct cccctgccat tttgttatgc aggagaaata
tgggcaaggc 180gagccttctg acgaaatgat gaataacatc ttggcagaca
tccgtaatat ccagttcgac 240cgcgtaatcg gtatcggagg aggtacggtt
attgacatct ctaaactttt cgttctgaaa 300ggattaaatg atgtactcga
tgcattcgac cgcaaaatac ctcttatcaa agagaaagaa 360ctgatcattg
tgcccacaac atgcggaacg ggtagcgagg tgacgaacat ttctatcgca
420gaaatcaaaa gccgtcacac caaaatggga ttggctgacg atgccattgt
tgcagaccat 480gccatcatca tacctgaact tctgaagagc ttgcctttcc
acttctacgc atgcagtgca 540atcgatgctc ttatccatgc catcgagtca
tacgtatctc ctaaagccag tccatattct 600cgtctgttca gtgaggcggc
ttgggacatt atcctggaag tattcaagaa aatcgccgaa 660cacggccctg
aataccgctt cgaaaagctg ggagaaatga tcatggccag caactatgcc
720ggtatagcct tcggaaatgc aggagtagga gccgtccacg cactatccta
cccgttggga 780ggcaactatc acgtgccgca tggagaagca
aactatcagt tcttcacaga ggtattcaaa 840gtataccaaa agaagaatcc
tttcggctat atagtcgaac tcaactggaa gctctccaag 900atactgaact
gccagcccga atacgtatat ccgaagctgg atgaacttct cggatgcctt
960cttaccaaga aacctttgca cgaatacggc atgaaggacg aagaggtaag
aggctttgcg 1020gaatcagtgc ttaagacaca gcaaagattg ctcgccaaca
actacgtaga gcttactgta 1080gatgagatcg aaggtatcta cagaagactc tactaa
111654371PRTPorphyromonas gingivalis 54Met Gln Leu Phe Lys Leu Lys
Ser Val Thr His His Phe Asp Thr Phe 1 5 10 15 Ala Glu Phe Ala Lys
Glu Phe Cys Leu Gly Glu Arg Asp Leu Val Ile 20 25 30 Thr Asn Glu
Phe Ile Tyr Glu Pro Tyr Met Lys Ala Cys Gln Leu Pro 35 40 45 Cys
His Phe Val Met Gln Glu Lys Tyr Gly Gln Gly Glu Pro Ser Asp 50 55
60 Glu Met Met Asn Asn Ile Leu Ala Asp Ile Arg Asn Ile Gln Phe Asp
65 70 75 80 Arg Val Ile Gly Ile Gly Gly Gly Thr Val Ile Asp Ile Ser
Lys Leu 85 90 95 Phe Val Leu Lys Gly Leu Asn Asp Val Leu Asp Ala
Phe Asp Arg Lys 100 105 110 Ile Pro Leu Ile Lys Glu Lys Glu Leu Ile
Ile Val Pro Thr Thr Cys 115 120 125 Gly Thr Gly Ser Glu Val Thr Asn
Ile Ser Ile Ala Glu Ile Lys Ser 130 135 140 Arg His Thr Lys Met Gly
Leu Ala Asp Asp Ala Ile Val Ala Asp His 145 150 155 160 Ala Ile Ile
Ile Pro Glu Leu Leu Lys Ser Leu Pro Phe His Phe Tyr 165 170 175 Ala
Cys Ser Ala Ile Asp Ala Leu Ile His Ala Ile Glu Ser Tyr Val 180 185
190 Ser Pro Lys Ala Ser Pro Tyr Ser Arg Leu Phe Ser Glu Ala Ala Trp
195 200 205 Asp Ile Ile Leu Glu Val Phe Lys Lys Ile Ala Glu His Gly
Pro Glu 210 215 220 Tyr Arg Phe Glu Lys Leu Gly Glu Met Ile Met Ala
Ser Asn Tyr Ala 225 230 235 240 Gly Ile Ala Phe Gly Asn Ala Gly Val
Gly Ala Val His Ala Leu Ser 245 250 255 Tyr Pro Leu Gly Gly Asn Tyr
His Val Pro His Gly Glu Ala Asn Tyr 260 265 270 Gln Phe Phe Thr Glu
Val Phe Lys Val Tyr Gln Lys Lys Asn Pro Phe 275 280 285 Gly Tyr Ile
Val Glu Leu Asn Trp Lys Leu Ser Lys Ile Leu Asn Cys 290 295 300 Gln
Pro Glu Tyr Val Tyr Pro Lys Leu Asp Glu Leu Leu Gly Cys Leu 305 310
315 320 Leu Thr Lys Lys Pro Leu His Glu Tyr Gly Met Lys Asp Glu Glu
Val 325 330 335 Arg Gly Phe Ala Glu Ser Val Leu Lys Thr Gln Gln Arg
Leu Leu Ala 340 345 350 Asn Asn Tyr Val Glu Leu Thr Val Asp Glu Ile
Glu Gly Ile Tyr Arg 355 360 365 Arg Leu Tyr 370
551296DNAPorphyromonas gingivalis 55atgaaagacg tattagcgga
atatgcctcc cgaattgttt cggccgaaga agccgtaaaa 60catatcaaaa atggagaacg
ggtagctttg tcacatgctg ccggagttcc tcagagttgt 120gttgatgcac
tggtacaaca ggccgacctt ttccagaatg tcgaaattta tcacatgctt
180tgtctcggcg aaggaaaata tatggcacct gaaatggccc ctcacttccg
acacataacc 240aattttgtag gtggtaattc tcgtaaagca gttgaggaaa
atagagccga cttcattccg 300gtattctttt atgaagtgcc atcaatgatt
cgcaaagaca tccttcacat agatgtcgcc 360atcgttcagc tttcaatgcc
tgatgagaat ggttactgta gttttggagt atcttgcgat 420tatagcaaac
cggcagcaga aagcgctcat ttagttatag gggaaatcaa ccgtcaaatg
480ccatatgtac atggcgacaa cttgattcac atatcgaagt tggattacat
cgtgatggca 540gactacccta tctattctct tgcaaagccc aaaatcggag
aagtagaaga agctatcggg 600cgtaattgtg ccgagcttat tgaagatggt
gccacactcc aactcggtat cggcgcgatt 660cctgatgcag ccctgttatt
cctcaaggac aaaaaagatc tggggatcca taccgagatg 720ttctccgatg
gtgttgtcga attagttcgc agtggagtaa ttacaggaaa gaaaaagaca
780cttcaccccg gaaagatggt cgcaaccttc ttaatgggaa gcgaagacgt
atatcatttc 840atcgacaaaa atcccgatgt agaactttat ccggtagatt
acgtcaatga tccgcgagta 900atcgctcaaa atgataatat ggtcagcatc
aatagctgta tcgaaatcga tcttatggga 960caagtcgtgt ccgaatgtat
aggaagcaag caattcagcg gaaccggcgg tcaagtagat 1020tatgttcgtg
gagcagcatg gtctaaaaac ggcaaaagca tcatggcaat tccctcaaca
1080gccaaaaacg gtactgcatc tcgaattgta cctataattg cagagggagc
tgctgtaaca 1140accctccgca acgaagtcga ttacgttgta accgaatacg
gtatagcaca actcaaagga 1200aagagtttgc gccagcgagc agaagctctt
attgccatag cccacccgga tttcagagag 1260gaactaacga aacatctccg
caaacgtttc ggataa 129656431PRTPorphyromonas gingivalis 56Met Lys
Asp Val Leu Ala Glu Tyr Ala Ser Arg Ile Val Ser Ala Glu 1 5 10 15
Glu Ala Val Lys His Ile Lys Asn Gly Glu Arg Val Ala Leu Ser His 20
25 30 Ala Ala Gly Val Pro Gln Ser Cys Val Asp Ala Leu Val Gln Gln
Ala 35 40 45 Asp Leu Phe Gln Asn Val Glu Ile Tyr His Met Leu Cys
Leu Gly Glu 50 55 60 Gly Lys Tyr Met Ala Pro Glu Met Ala Pro His
Phe Arg His Ile Thr 65 70 75 80 Asn Phe Val Gly Gly Asn Ser Arg Lys
Ala Val Glu Glu Asn Arg Ala 85 90 95 Asp Phe Ile Pro Val Phe Phe
Tyr Glu Val Pro Ser Met Ile Arg Lys 100 105 110 Asp Ile Leu His Ile
Asp Val Ala Ile Val Gln Leu Ser Met Pro Asp 115 120 125 Glu Asn Gly
Tyr Cys Ser Phe Gly Val Ser Cys Asp Tyr Ser Lys Pro 130 135 140 Ala
Ala Glu Ser Ala His Leu Val Ile Gly Glu Ile Asn Arg Gln Met 145 150
155 160 Pro Tyr Val His Gly Asp Asn Leu Ile His Ile Ser Lys Leu Asp
Tyr 165 170 175 Ile Val Met Ala Asp Tyr Pro Ile Tyr Ser Leu Ala Lys
Pro Lys Ile 180 185 190 Gly Glu Val Glu Glu Ala Ile Gly Arg Asn Cys
Ala Glu Leu Ile Glu 195 200 205 Asp Gly Ala Thr Leu Gln Leu Gly Ile
Gly Ala Ile Pro Asp Ala Ala 210 215 220 Leu Leu Phe Leu Lys Asp Lys
Lys Asp Leu Gly Ile His Thr Glu Met 225 230 235 240 Phe Ser Asp Gly
Val Val Glu Leu Val Arg Ser Gly Val Ile Thr Gly 245 250 255 Lys Lys
Lys Thr Leu His Pro Gly Lys Met Val Ala Thr Phe Leu Met 260 265 270
Gly Ser Glu Asp Val Tyr His Phe Ile Asp Lys Asn Pro Asp Val Glu 275
280 285 Leu Tyr Pro Val Asp Tyr Val Asn Asp Pro Arg Val Ile Ala Gln
Asn 290 295 300 Asp Asn Met Val Ser Ile Asn Ser Cys Ile Glu Ile Asp
Leu Met Gly 305 310 315 320 Gln Val Val Ser Glu Cys Ile Gly Ser Lys
Gln Phe Ser Gly Thr Gly 325 330 335 Gly Gln Val Asp Tyr Val Arg Gly
Ala Ala Trp Ser Lys Asn Gly Lys 340 345 350 Ser Ile Met Ala Ile Pro
Ser Thr Ala Lys Asn Gly Thr Ala Ser Arg 355 360 365 Ile Val Pro Ile
Ile Ala Glu Gly Ala Ala Val Thr Thr Leu Arg Asn 370 375 380 Glu Val
Asp Tyr Val Val Thr Glu Tyr Gly Ile Ala Gln Leu Lys Gly 385 390 395
400 Lys Ser Leu Arg Gln Arg Ala Glu Ala Leu Ile Ala Ile Ala His Pro
405 410 415 Asp Phe Arg Glu Glu Leu Thr Lys His Leu Arg Lys Arg Phe
Gly 420 425 430 57906DNAClostridium acetobutylicum 57atgattaaga
gttttaatga aattatcatg aaggtaaaga gcaaagaaat gaaaaaagtt 60gctgttgctg
tagcacaaga cgagccagta cttgaagcag taagagatgc taagaaaaat
120ggtattgcag atgctattct tgttggagac catgacgaaa tcgtgtcaat
cgcgcttaaa 180ataggaatgg atgtaaatga ttttgaaata gtaaacgagc
ctaacgttaa gaaagctgct 240ttaaaggcag tagagcttgt atcaactgga
aaagctgata tggtaatgaa gggacttgta 300aatacagcaa ctttcttaag
atctgtatta aacaaagaag ttggacttag aacaggaaaa 360actatgtctc
acgttgcagt atttgaaact gagaaatttg atagactatt atttttaaca
420gatgttgctt tcaatactta tcctgaatta aaggaaaaaa ttgatatagt
aaacaattca 480gttaaggttg cacatgcaat aggaattgaa aatccaaagg
ttgctccaat ttgtgcagtt 540gaggttataa accctaaaat gccatcaaca
cttgatgcag caatgctttc aaaaatgagt 600gacagaggac aaattaaagg
ttgtgtagtt gacggacctt tagcacttga tatagcttta 660tcagaagaag
cagcacatca taagggagta acaggagaag ttgctggaaa agctgatatc
720ttcttaatgc caaacataga aacaggaaat gtaatgtata agactttaac
atatacaact 780gattcaaaaa atggaggaat cttagttgga acttctgcac
cagttgtttt aacttcaaga 840gctgacagcc atgaaacaaa aatgaactct
atagcacttg cagctttagt tgcaggcaat 900aaataa 90658301PRTClostridium
acetobutylicum 58Met Ile Lys Ser Phe Asn Glu Ile Ile Met Lys Val
Lys Ser Lys Glu 1 5 10 15 Met Lys Lys Val Ala Val Ala Val Ala Gln
Asp Glu Pro Val Leu Glu 20 25 30 Ala Val Arg Asp Ala Lys Lys Asn
Gly Ile Ala Asp Ala Ile Leu Val 35 40 45 Gly Asp His Asp Glu Ile
Val Ser Ile Ala Leu Lys Ile Gly Met Asp 50 55 60 Val Asn Asp Phe
Glu Ile Val Asn Glu Pro Asn Val Lys Lys Ala Ala 65 70 75 80 Leu Lys
Ala Val Glu Leu Val Ser Thr Gly Lys Ala Asp Met Val Met 85 90 95
Lys Gly Leu Val Asn Thr Ala Thr Phe Leu Arg Ser Val Leu Asn Lys 100
105 110 Glu Val Gly Leu Arg Thr Gly Lys Thr Met Ser His Val Ala Val
Phe 115 120 125 Glu Thr Glu Lys Phe Asp Arg Leu Leu Phe Leu Thr Asp
Val Ala Phe 130 135 140 Asn Thr Tyr Pro Glu Leu Lys Glu Lys Ile Asp
Ile Val Asn Asn Ser 145 150 155 160 Val Lys Val Ala His Ala Ile Gly
Ile Glu Asn Pro Lys Val Ala Pro 165 170 175 Ile Cys Ala Val Glu Val
Ile Asn Pro Lys Met Pro Ser Thr Leu Asp 180 185 190 Ala Ala Met Leu
Ser Lys Met Ser Asp Arg Gly Gln Ile Lys Gly Cys 195 200 205 Val Val
Asp Gly Pro Leu Ala Leu Asp Ile Ala Leu Ser Glu Glu Ala 210 215 220
Ala His His Lys Gly Val Thr Gly Glu Val Ala Gly Lys Ala Asp Ile 225
230 235 240 Phe Leu Met Pro Asn Ile Glu Thr Gly Asn Val Met Tyr Lys
Thr Leu 245 250 255 Thr Tyr Thr Thr Asp Ser Lys Asn Gly Gly Ile Leu
Val Gly Thr Ser 260 265 270 Ala Pro Val Val Leu Thr Ser Arg Ala Asp
Ser His Glu Thr Lys Met 275 280 285 Asn Ser Ile Ala Leu Ala Ala Leu
Val Ala Gly Asn Lys 290 295 300 591068DNAClostridium acetobutylicum
59atgtatagat tactaataat caatcctggc tcgacctcaa ctaaaattgg tatttatgac
60gatgaaaaag agatatttga gaagacttta agacattcag ctgaagagat agaaaaatat
120aacactatat ttgatcaatt tcaattcaga aagaatgtaa ttttagatgc
gttaaaagaa 180gcaaacatag aagtaagttc tttaaatgct gtagttggaa
gaggcggact cttaaagcca 240atagtaagtg gaacttatgc agtaaatcaa
aaaatgcttg aagaccttaa agtaggagtt 300caaggtcagc atgcgtcaaa
tcttggtgga attattgcaa atgaaatagc aaaagaaata 360aatgttccag
catacatagt tgatccagtt gttgtggatg agcttgatga agtttcaaga
420atatcaggaa tggctgacat tccaagaaaa agtatattcc atgcattaaa
tcaaaaagca 480gttgctagaa gatatgcaaa agaagttgga aaaaaatacg
aagatcttaa tttaatcgta 540gtccacatgg gtggaggtac ttcagtaggt
actcataaag atggtagagt aatagaagtt 600aataatacac ttgatggaga
aggtccattc tcaccagaaa gaagtggtgg agttccaata 660ggagatcttg
taagattgtg cttcagcaac aaatatactt atgaagaagt aatgaaaaag
720ataaacggca aaggcggagt tgttagttac ttaaatacta tcgattttaa
ggctgtagtt 780gataaagctc ttgaaggaga taagaaatgt gcacttatat
atgaagcttt cacattccag 840gtagcaaaag agataggaaa atgttcaacc
gttttaaaag gaaatgtaga tgcaataatc 900ttaacaggcg gaattgcgta
caacgagcat gtatgtaatg ccatagagga tagagtaaaa 960ttcatagcac
ctgtagttag atatggtgga gaagatgaac ttcttgcact tgcagaaggt
1020ggacttagag ttttaagagg agaagaaaaa gctaaggaat acaaataa
106860355PRTClostridium acetobutylicum 60Met Tyr Arg Leu Leu Ile
Ile Asn Pro Gly Ser Thr Ser Thr Lys Ile 1 5 10 15 Gly Ile Tyr Asp
Asp Glu Lys Glu Ile Phe Glu Lys Thr Leu Arg His 20 25 30 Ser Ala
Glu Glu Ile Glu Lys Tyr Asn Thr Ile Phe Asp Gln Phe Gln 35 40 45
Phe Arg Lys Asn Val Ile Leu Asp Ala Leu Lys Glu Ala Asn Ile Glu 50
55 60 Val Ser Ser Leu Asn Ala Val Val Gly Arg Gly Gly Leu Leu Lys
Pro 65 70 75 80 Ile Val Ser Gly Thr Tyr Ala Val Asn Gln Lys Met Leu
Glu Asp Leu 85 90 95 Lys Val Gly Val Gln Gly Gln His Ala Ser Asn
Leu Gly Gly Ile Ile 100 105 110 Ala Asn Glu Ile Ala Lys Glu Ile Asn
Val Pro Ala Tyr Ile Val Asp 115 120 125 Pro Val Val Val Asp Glu Leu
Asp Glu Val Ser Arg Ile Ser Gly Met 130 135 140 Ala Asp Ile Pro Arg
Lys Ser Ile Phe His Ala Leu Asn Gln Lys Ala 145 150 155 160 Val Ala
Arg Arg Tyr Ala Lys Glu Val Gly Lys Lys Tyr Glu Asp Leu 165 170 175
Asn Leu Ile Val Val His Met Gly Gly Gly Thr Ser Val Gly Thr His 180
185 190 Lys Asp Gly Arg Val Ile Glu Val Asn Asn Thr Leu Asp Gly Glu
Gly 195 200 205 Pro Phe Ser Pro Glu Arg Ser Gly Gly Val Pro Ile Gly
Asp Leu Val 210 215 220 Arg Leu Cys Phe Ser Asn Lys Tyr Thr Tyr Glu
Glu Val Met Lys Lys 225 230 235 240 Ile Asn Gly Lys Gly Gly Val Val
Ser Tyr Leu Asn Thr Ile Asp Phe 245 250 255 Lys Ala Val Val Asp Lys
Ala Leu Glu Gly Asp Lys Lys Cys Ala Leu 260 265 270 Ile Tyr Glu Ala
Phe Thr Phe Gln Val Ala Lys Glu Ile Gly Lys Cys 275 280 285 Ser Thr
Val Leu Lys Gly Asn Val Asp Ala Ile Ile Leu Thr Gly Gly 290 295 300
Ile Ala Tyr Asn Glu His Val Cys Asn Ala Ile Glu Asp Arg Val Lys 305
310 315 320 Phe Ile Ala Pro Val Val Arg Tyr Gly Gly Glu Asp Glu Leu
Leu Ala 325 330 335 Leu Ala Glu Gly Gly Leu Arg Val Leu Arg Gly Glu
Glu Lys Ala Lys 340 345 350 Glu Tyr Lys 355 61906DNAArtificial
SequenceDescription of Artificial Sequence Synthetic polynucleotide
61atgattaaga gttttaatga aattatcatg aaggtaaaga gcaaagaaat gaaaaaagtt
60gctgttgctg tagcacaaga cgagccagta cttgaagcag tacgcgatgc taagaaaaat
120ggtattgcag atgctattct tgttggcgac catgacgaaa tcgtgtcaat
cgcgcttaaa 180ataggcatgg atgtaaatga ttttgaaata gtaaacgagc
ctaacgttaa gaaagctgct 240ttaaaggcag tagagctggt atcaactgga
aaagctgata tggtaatgaa gggacttgta 300aatacagcaa ctttcttacg
ctctgtatta aacaaagaag ttggactgag aacaggaaaa 360actatgtctc
acgttgcagt atttgaaact gagaaatttg atcgtctgtt atttttaaca
420gatgttgctt tcaatactta tcctgaatta aaggaaaaaa ttgatatcgt
aaacaattca 480gttaaggttg cacatgcaat aggtattgaa aatccaaagg
ttgctccaat ttgtgcagtt 540gaggttataa accctaaaat gccatcaaca
cttgatgcag caatgctttc aaaaatgagt 600gacagaggac aaattaaagg
ttgtgtagtt gacggaccgt tagcacttga tatcgcttta 660tcagaagaag
cagcacatca taagggcgta acaggagaag ttgctggaaa agctgatatc
720ttcttaatgc caaacattga aacaggaaat gtaatgtata agactttaac
atatacaact 780gatagcaaaa atggcggaat cttagttgga acttctgcac
cagttgtttt aacttcacgc 840gctgacagcc atgaaacaaa aatgaactct
attgcacttg cagctttagt tgcaggcaat 900aaataa 90662906DNAArtificial
SequenceDescription of Artificial Sequence Synthetic polynucleotide
62atgattaaga gttttaatga aattatcatg aaggtaaaga gcaaagaaat gaaaaaagtt
60gctgttgctg tagcacaaga cgagccagta cttgaagcag tacgcgatgc taagaaaaat
120ggtattgccg atgctattct ggttggcgac catgacgaaa tcgtgtctat
cgcgctgaaa 180ataggcatgg atgtaaatga ttttgaaatt gttaacgagc
ctaacgttaa gaaagctgcg 240ttaaaggcag tagagctggt atcaactgga
aaagctgata tggtaatgaa gggactggta 300aataccgcaa ctttcttacg
ctctgtatta aacaaagaag ttggtctgcg tacaggaaaa 360accatgtctc
acgttgcagt atttgaaact gagaaatttg atcgtctgtt atttttaaca
420gatgttgctt tcaatactta tcctgaatta aaggaaaaaa ttgatatcgt
taacaatagc
480gttaaggttg cacatgccat tggtattgaa aatccaaagg ttgctccaat
ttgtgcagtt 540gaggttatta acccgaaaat gccatcaaca cttgatgcag
caatgctttc aaaaatgagt 600gaccgcggac aaattaaagg ttgtgtagtt
gacggaccgc tggcacttga tatcgcttta 660tcagaagaag cagcacatca
taaaggcgta acaggagaag ttgctggaaa agctgatatc 720ttcttaatgc
caaacattga aacaggaaat gtaatgtata agacgttaac ctataccact
780gatagcaaaa atggcggcat cctggttgga acttctgcac cagttgtttt
aacttcacgc 840gctgacagcc atgaaacaaa aatgaactct attgcactgg
cagcgctggt tgcaggcaat 900aaataa 90663906DNAArtificial
SequenceDescription of Artificial Sequence Synthetic polynucleotide
63atgattaaga gttttaatga aattatcatg aaggtaaaga gcaaagaaat gaaaaaagtt
60gctgttgctg ttgcacaaga cgagccggta ctggaagcgg tacgcgatgc taagaaaaat
120ggtattgccg atgctattct ggttggcgac catgacgaaa tcgtctctat
cgcgctgaaa 180attggcatgg atgttaatga ttttgaaatt gttaacgagc
ctaacgttaa gaaagctgcg 240ctgaaggcgg tagagctggt ttccaccgga
aaagctgata tggtaatgaa agggctggtg 300aataccgcaa ctttcttacg
cagcgtactg aacaaagaag ttggtctgcg taccggaaaa 360accatgagtc
acgttgcggt atttgaaact gagaaatttg atcgtctgct gtttctgacc
420gatgttgctt tcaatactta tcctgaatta aaagaaaaaa ttgatatcgt
taacaatagc 480gttaaggttg cgcatgccat tggtattgaa aatccaaagg
ttgctccaat ttgtgcagtt 540gaggttatta acccgaaaat gccatcaaca
cttgatgccg caatgcttag caaaatgagt 600gaccgcggac aaattaaagg
ttgtgtggtt gacggcccgc tggcactgga tatcgcgtta 660agcgaagaag
cggcacatca taaaggcgta accggcgaag ttgctggaaa agctgatatc
720ttcctgatgc caaacattga aacaggcaat gtaatgtata aaacgttaac
ctataccact 780gatagcaaaa atggcggcat cctggttgga acttctgcac
cagttgtttt aacctcacgc 840gctgacagcc atgaaaccaa aatgaacagc
attgcactgg cagcgctggt tgcaggcaat 900aaataa 90664906DNAArtificial
SequenceDescription of Artificial Sequence Synthetic polynucleotide
64atgattaaaa gttttaacga aattatcatg aaagtgaaaa gcaaagagat gaaaaaagtg
60gcggttgcgg ttgcgcagga tgaaccggtg ctggaagcgg tgcgcgatgc caaaaaaaac
120ggtattgccg atgccattct ggtgggcgat cacgatgaaa ttgtctctat
tgcgctgaaa 180attggcatgg atgttaacga ttttgaaatt gttaatgaac
cgaacgtgaa aaaagcggcg 240ctgaaagcgg ttgaactggt ttccaccggt
aaagccgata tggtgatgaa agggctggtg 300aataccgcaa ccttcctgcg
cagcgtgctg aataaagaag tgggtctgcg taccggtaaa 360accatgagtc
atgttgcggt gtttgaaacc gaaaaatttg accgtctgct gtttctgacc
420gatgttgcgt ttaataccta tccggaactg aaagagaaaa ttgatatcgt
taataacagc 480gtgaaagtgg cgcatgccat tggtattgaa aacccgaaag
tggcgccgat ttgcgcggtt 540gaagtgatta acccgaaaat gccgtcaacg
ctggatgccg cgatgctcag caaaatgagc 600gatcgcggtc aaatcaaagg
ctgtgtggtt gatggcccgc tggcgctgga tatcgcgctt 660agcgaagaag
cggcgcatca taaaggcgtg accggcgaag tggccggtaa agccgatatt
720ttcctgatgc cgaatattga aaccggcaac gtgatgtata aaacgctgac
ctataccacc 780gacagcaaaa acggcggcat tctggtgggt accagcgcgc
cggtggtgct gacctcgcgc 840gccgacagcc atgaaaccaa aatgaacagc
attgcgctgg cggcgctggt ggccggtaat 900aaataa 906651068DNAArtificial
SequenceDescription of Artificial Sequence Synthetic polynucleotide
65atgtatcgtt tactgattat caatcctggc tcgacctcaa ctaaaattgg tatttatgac
60gatgaaaaag agatatttga gaagacttta cgtcattcag ctgaagagat agaaaaatat
120aacactatat ttgatcaatt tcagttcaga aagaatgtaa ttctcgatgc
gttaaaagaa 180gcaaacattg aagtaagttc tttaaatgct gtagttggac
gcggcggact gttaaagcca 240atagtaagtg gaacttatgc agtaaatcaa
aaaatgcttg aagaccttaa agtaggcgtt 300caaggtcagc atgcgtcaaa
tcttggtgga attattgcaa atgaaatagc aaaagaaata 360aatgttccag
catacatcgt tgatccagtt gttgtggatg agcttgatga agtttcacgt
420atatcaggaa tggctgacat tccacgtaaa agtatattcc atgcattaaa
tcaaaaagca 480gttgctagac gctatgcaaa agaagttgga aaaaaatacg
aagatcttaa tttaatcgtg 540gtccacatgg gtggcggtac ttcagtaggt
actcataaag atggtagagt aattgaagtt 600aataatacac ttgatggaga
aggtccattc tcaccagaaa gaagtggtgg cgttccaata 660ggcgatcttg
tacgtttgtg cttcagcaac aaatatactt atgaagaagt aatgaaaaag
720ataaacggca aaggcggcgt tgttagttac ttaaatacta tcgattttaa
ggctgtagtt 780gataaagctc ttgaaggcga taagaaatgt gcacttatat
atgaagcttt cacattccag 840gtagcaaaag agataggaaa atgttcaacc
gttttaaaag gaaatgtaga tgcaataatc 900ttaacaggcg gaattgcgta
caacgagcat gtatgtaatg ccatagagga tagagtaaaa 960ttcattgcac
ctgtagttcg ttatggtgga gaagatgaac ttcttgcact tgcagaaggt
1020ggactgcgcg ttttacgcgg agaagaaaaa gctaaggaat acaaataa
1068661068DNAArtificial SequenceDescription of Artificial Sequence
Synthetic polynucleotide 66atgtatcgtt tactgattat caatcctggc
tcgacctcaa ctaaaattgg tatttatgac 60gatgaaaaag agatatttga gaagacgtta
cgtcattcag ctgaagagat tgaaaaatat 120aacactatat ttgatcaatt
tcagttccgc aagaatgtga ttctcgatgc gttaaaagaa 180gcaaacattg
aagtcagttc tttaaatgct gtagttggac gcggcggact gttaaagcca
240attgtcagtg gaacttatgc agtaaatcaa aaaatgcttg aagaccttaa
agtgggcgtt 300caaggtcagc atgccagcaa tcttggtggc attattgcca
atgaaatcgc aaaagaaatc 360aatgttccag catacatcgt tgatccggtt
gttgtggatg agcttgatga agttagccgt 420ataagcggaa tggctgacat
tccacgtaaa agtatattcc atgcattaaa tcaaaaagca 480gttgctcgtc
gctatgcaaa agaagttggt aaaaaatacg aagatcttaa tttaatcgtg
540gtccacatgg gtggcggtac ttcagtaggt actcataaag atggtcgcgt
gattgaagtt 600aataatacac ttgatggcga aggtccattc tcaccagaac
gtagtggtgg cgttccaatt 660ggcgatctgg tacgtttgtg cttcagcaac
aaatatactt atgaagaagt gatgaaaaag 720ataaacggca aaggcggcgt
tgttagttac ctgaatacta tcgattttaa ggctgtagtt 780gataaagcgc
ttgaaggcga taagaaatgt gcactgattt atgaagcttt caccttccag
840gtagcaaaag agattggtaa atgttcaacc gttttaaaag gaaatgttga
tgccattatc 900ttaacaggcg gcattgctta caacgagcat gtatgtaatg
ccattgagga tcgcgtaaaa 960ttcattgcac ctgtagttcg ttatggtggc
gaagatgaac tgctggcact ggcagaaggt 1020ggactgcgcg ttttacgcgg
cgaagaaaaa gcgaaggaat acaaataa 1068671068DNAArtificial
SequenceDescription of Artificial Sequence Synthetic polynucleotide
67atgtatcgtc tgctgattat caatcctggc tcgacctcaa ctaaaattgg tatttatgac
60gatgaaaaag agatatttga gaaaacgtta cgtcatagcg ctgaagagat tgaaaaatat
120aacactattt ttgatcaatt tcagttccgc aagaatgtga ttctcgatgc
gctgaaagaa 180gcaaacattg aagtcagttc gctgaatgcg gtagttggtc
gcggcggtct gctgaagcca 240attgtcagcg gcacttatgc ggtaaatcaa
aaaatgctgg aagacctgaa agtgggcgtt 300caggggcagc atgccagcaa
tcttggtggc attattgcca atgaaatcgc caaagaaatc 360aatgttccgg
catacatcgt tgatccggtt gttgtggatg agctggatga agttagccgt
420atcagcggaa tggctgacat tccacgtaaa agtattttcc atgcactgaa
tcaaaaagcg 480gttgcgcgtc gctatgcaaa agaagttggt aaaaaatacg
aagatcttaa tctgatcgtg 540gtgcatatgg gtggcggtac tagcgtcggt
actcataaag atggtcgcgt gattgaagtt 600aataatacac ttgatggcga
aggtccattc tcaccagaac gtagcggtgg cgttccaatt 660ggcgatctgg
tacgtttgtg cttcagcaac aaatatacct atgaagaagt gatgaaaaag
720ataaacggca aaggcggcgt tgttagttac ctgaatacta tcgattttaa
ggcggtagtt 780gataaagcgc tggaaggcga taagaaatgt gcactgattt
atgaagcgtt caccttccag 840gtggcaaaag agattggtaa atgttcaacc
gttctgaaag gcaatgttga tgccattatc 900ctgaccggcg gcattgctta
caacgagcat gtttgtaatg ccattgagga tcgcgtaaaa 960ttcattgcac
ctgtggttcg ttatggtggc gaagatgaac tgctggcact ggcagaaggt
1020ggtctgcgcg ttttacgcgg cgaagaaaaa gcgaaagaat acaaataa
1068681068DNAArtificial SequenceDescription of Artificial Sequence
Synthetic polynucleotide 68atgtatcgtc tgctgattat caacccgggc
agcacctcaa ccaaaattgg tatttacgac 60gatgaaaaag agatttttga aaaaacgctg
cgtcacagcg cagaagagat tgaaaaatac 120aacaccattt tcgatcagtt
ccagttccgc aaaaacgtga ttctcgatgc gctgaaagaa 180gccaatattg
aagtctcctc gctgaatgcg gtggtcggtc gcggcggtct gctgaaaccg
240attgtcagcg gcacttatgc ggttaatcag aaaatgctgg aagatctgaa
agtgggcgtg 300caggggcagc atgccagcaa tctcggcggc attatcgcca
atgaaatcgc caaagagatc 360aacgtgccgg cttatatcgt cgatccggtg
gtggttgatg aactggatga agtcagccgt 420atcagcggca tggcggatat
tccgcgtaaa agcattttcc atgcgctgaa tcagaaagcg 480gttgcgcgtc
gctatgccaa agaagtgggt aaaaaatatg aagatctcaa tctgattgtg
540gtgcatatgg gcggcggcac cagcgtcggt acgcataaag atggtcgcgt
gattgaagtg 600aataacacgc tggatggcga agggccgttc tcgccggaac
gtagcggcgg cgtgccgatt 660ggcgatctgg tgcgtctgtg tttcagcaat
aaatacacct acgaagaagt gatgaaaaaa 720atcaacggca aaggcggcgt
ggttagctat ctgaatacca tcgattttaa agcggtggtt 780gataaagcgc
tggaaggcga taaaaaatgc gcgctgattt atgaagcgtt taccttccag
840gtggcgaaag agattggtaa atgttcaacc gtgctgaaag gcaacgttga
tgccattatt 900ctgaccggcg gcattgctta taacgaacat gtttgtaatg
ccattgaaga tcgcgtgaaa 960tttattgcgc cggtggtgcg ttacggcggc
gaagatgaac tgctggcgct ggcggaaggc 1020ggtctgcgcg tgctgcgcgg
cgaagaaaaa gcgaaagagt acaaataa 1068691407DNAClostridium
biejerinckii 69atgaataaag acacactaat acctacaact aaagatttaa
aagtaaaaac aaatggtgaa 60aacattaatt taaagaacta caaggataat tcttcatgtt
tcggagtatt cgaaaatgtt 120gaaaatgcta taagcagcgc tgtacacgca
caaaagatat tatcccttca ttatacaaaa 180gagcaaagag aaaaaatcat
aactgagata agaaaggccg cattacaaaa taaagaggtc 240ttggctacaa
tgattctaga agaaacacat atgggaagat atgaggataa aatattaaaa
300catgaattgg tagctaaata tactcctggt acagaagatt taactactac
tgcttggtca 360ggtgataatg gtcttacagt tgtagaaatg tctccatatg
gtgttatagg tgcaataact 420ccttctacga atccaactga aactgtaata
tgtaatagca taggcatgat agctgctgga 480aatgctgtag tatttaacgg
acacccatgc gctaaaaaat gtgttgcctt tgctgttgaa 540atgataaata
aggcaattat ttcatgtggc ggtcctgaaa atctagtaac aactataaaa
600aatccaacta tggagtctct agatgcaatt attaagcatc cttcaataaa
acttctttgc 660ggaactgggg gtccaggaat ggtaaaaacc ctcttaaatt
ctggtaagaa agctataggt 720gctggtgctg gaaatccacc agttattgta
gatgatactg ctgatataga aaaggctggt 780aggagcatca ttgaaggctg
ttcttttgat aataatttac cttgtattgc agaaaaagaa 840gtatttgttt
ttgagaatgt tgcagatgat ttaatatcta acatgctaaa aaataatgct
900gtaattataa atgaagatca agtatcaaaa ttaatagatt tagtattaca
aaaaaataat 960gaaactcaag aatactttat aaacaaaaaa tgggtaggaa
aagatgcaaa attattctta 1020gatgaaatag atgttgagtc tccttcaaat
gttaaatgca taatctgcga agtaaatgca 1080aatcatccat ttgttatgac
agaactcatg atgccaatat tgccaattgt aagagttaaa 1140gatatagatg
aagctattaa atatgcaaag atagcagaac aaaatagaaa acatagtgcc
1200tatatttatt ctaaaaatat agacaaccta aatagatttg aaagagaaat
agatactact 1260atttttgtaa agaatgctaa atcttttgct ggtgttggtt
atgaagcaga aggatttaca 1320actttcacta ttgctggatc tactggtgag
ggaataacct ctgcaaggaa ttttacaaga 1380caaagaagat gtgtacttgc cggctaa
140770468PRTClostridium biejerinckii 70Met Asn Lys Asp Thr Leu Ile
Pro Thr Thr Lys Asp Leu Lys Val Lys 1 5 10 15 Thr Asn Gly Glu Asn
Ile Asn Leu Lys Asn Tyr Lys Asp Asn Ser Ser 20 25 30 Cys Phe Gly
Val Phe Glu Asn Val Glu Asn Ala Ile Ser Ser Ala Val 35 40 45 His
Ala Gln Lys Ile Leu Ser Leu His Tyr Thr Lys Glu Gln Arg Glu 50 55
60 Lys Ile Ile Thr Glu Ile Arg Lys Ala Ala Leu Gln Asn Lys Glu Val
65 70 75 80 Leu Ala Thr Met Ile Leu Glu Glu Thr His Met Gly Arg Tyr
Glu Asp 85 90 95 Lys Ile Leu Lys His Glu Leu Val Ala Lys Tyr Thr
Pro Gly Thr Glu 100 105 110 Asp Leu Thr Thr Thr Ala Trp Ser Gly Asp
Asn Gly Leu Thr Val Val 115 120 125 Glu Met Ser Pro Tyr Gly Val Ile
Gly Ala Ile Thr Pro Ser Thr Asn 130 135 140 Pro Thr Glu Thr Val Ile
Cys Asn Ser Ile Gly Met Ile Ala Ala Gly 145 150 155 160 Asn Ala Val
Val Phe Asn Gly His Pro Cys Ala Lys Lys Cys Val Ala 165 170 175 Phe
Ala Val Glu Met Ile Asn Lys Ala Ile Ile Ser Cys Gly Gly Pro 180 185
190 Glu Asn Leu Val Thr Thr Ile Lys Asn Pro Thr Met Glu Ser Leu Asp
195 200 205 Ala Ile Ile Lys His Pro Ser Ile Lys Leu Leu Cys Gly Thr
Gly Gly 210 215 220 Pro Gly Met Val Lys Thr Leu Leu Asn Ser Gly Lys
Lys Ala Ile Gly 225 230 235 240 Ala Gly Ala Gly Asn Pro Pro Val Ile
Val Asp Asp Thr Ala Asp Ile 245 250 255 Glu Lys Ala Gly Arg Ser Ile
Ile Glu Gly Cys Ser Phe Asp Asn Asn 260 265 270 Leu Pro Cys Ile Ala
Glu Lys Glu Val Phe Val Phe Glu Asn Val Ala 275 280 285 Asp Asp Leu
Ile Ser Asn Met Leu Lys Asn Asn Ala Val Ile Ile Asn 290 295 300 Glu
Asp Gln Val Ser Lys Leu Ile Asp Leu Val Leu Gln Lys Asn Asn 305 310
315 320 Glu Thr Gln Glu Tyr Phe Ile Asn Lys Lys Trp Val Gly Lys Asp
Ala 325 330 335 Lys Leu Phe Leu Asp Glu Ile Asp Val Glu Ser Pro Ser
Asn Val Lys 340 345 350 Cys Ile Ile Cys Glu Val Asn Ala Asn His Pro
Phe Val Met Thr Glu 355 360 365 Leu Met Met Pro Ile Leu Pro Ile Val
Arg Val Lys Asp Ile Asp Glu 370 375 380 Ala Ile Lys Tyr Ala Lys Ile
Ala Glu Gln Asn Arg Lys His Ser Ala 385 390 395 400 Tyr Ile Tyr Ser
Lys Asn Ile Asp Asn Leu Asn Arg Phe Glu Arg Glu 405 410 415 Ile Asp
Thr Thr Ile Phe Val Lys Asn Ala Lys Ser Phe Ala Gly Val 420 425 430
Gly Tyr Glu Ala Glu Gly Phe Thr Thr Phe Thr Ile Ala Gly Ser Thr 435
440 445 Gly Glu Gly Ile Thr Ser Ala Arg Asn Phe Thr Arg Gln Arg Arg
Cys 450 455 460 Val Leu Ala Gly 465 711407DNAArtificial
SequenceDescription of Artificial Sequence Synthetic polynucleotide
71atgaataaag acacactaat acctacaact aaagatttaa aagtaaaaac aaatggtgaa
60aacattaatt taaagaacta caaggataat tcttcatgtt tcggcgtatt cgaaaatgtt
120gaaaatgcta taagcagcgc tgtacacgca caaaagatat tatcccttca
ttatacaaaa 180gagcaacgtg aaaaaatcat aactgagata agaaaggccg
cattacaaaa taaagaggtc 240ttggctacaa tgattctgga agaaacacat
atgggacgtt atgaggataa aatattaaaa 300catgaattgg tagctaaata
tactcctggt acagaagatt taactactac tgcctggtca 360ggtgataatg
gtctgacagt tgtagaaatg tctccatatg gtgttattgg tgcaataact
420ccttctacga atccaactga aactgtaata tgtaatagca taggcatgat
tgctgctgga 480aatgctgtag tatttaacgg acacccatgc gctaaaaaat
gtgttgcctt tgctgttgaa 540atgataaata aggcaattat ttcatgtggc
ggtcctgaaa atctggtaac aactataaaa 600aatccaacca tggagtctct
ggatgcaatt attaagcatc cttcaataaa acttctttgc 660ggaactgggg
gtccaggaat ggtaaaaacc ctgttaaatt ctggtaagaa agctataggt
720gctggtgctg gaaatccacc agttattgtc gatgatactg ctgatataga
aaaggctggt 780cgtagcatca ttgaaggctg ttcttttgat aataatttac
cttgtattgc agaaaaagaa 840gtatttgttt ttgagaatgt tgcagatgat
ttaatatcta acatgctaaa aaataatgct 900gtaattataa atgaagatca
agtatcaaaa ttaatcgatt tagtattaca aaaaaataat 960gaaactcaag
aatactttat aaacaaaaaa tgggtaggaa aagatgcaaa attattcctc
1020gatgaaatag atgttgagtc tccttcaaat gttaaatgca taatctgcga
agtaaatgca 1080aatcatccat ttgttatgac agaactgatg atgccaatat
tgccaattgt acgcgttaaa 1140gatatcgatg aagctattaa atatgcaaag
atagcagaac aaaatagaaa acatagtgcc 1200tatatttatt ctaaaaatat
cgacaacctg aatcgctttg aacgtgaaat agatactact 1260atttttgtaa
agaatgctaa atcttttgct ggtgttggtt atgaagcaga aggatttaca
1320actttcacta ttgctggatc tactggtgag ggaataacct ctgcacgtaa
ttttacacgc 1380caacgtcgct gtgtacttgc cggctaa
1407721407DNAArtificial SequenceDescription of Artificial Sequence
Synthetic polynucleotide 72atgaataaag acacactgat ccctacaact
aaagatttaa aagtaaaaac aaatggtgaa 60aacattaatt taaagaacta caaagataat
agcagttgtt tcggcgtatt cgaaaatgtt 120gaaaatgcta tcagcagcgc
tgtacacgca caaaagatat tatcgctgca ttatacaaaa 180gagcaacgtg
aaaaaatcat cactgagata cgtaaggccg cattacaaaa taaagaggtg
240ctggctacaa tgattctgga agaaacacat atgggacgtt atgaggataa
aatattaaaa 300catgaactgg tagctaaata tactcctggt acagaagatt
taactactac tgcctggagc 360ggtgataatg gtctgacagt tgtagaaatg
tctccatatg gtgttattgg tgcaataact 420ccttctacca atccaactga
aactgtaatt tgtaatagca ttggcatgat tgctgctgga 480aatgctgtag
tatttaacgg acacccatgc gctaaaaaat gtgttgcctt tgctgttgaa
540atgatcaata aggcaattat tagctgtggc ggtccggaaa atctggtaac
aactataaaa 600aatccaacca tggagtctct ggatgccatt attaagcatc
cttcaataaa actgctttgc 660ggaactggcg gtccaggaat ggtaaaaacc
ctgttaaatt ctggtaagaa agctattggt 720gctggtgctg gaaatccacc
agttattgtc gatgatactg ctgatattga aaaggctggt 780cgtagcatca
ttgaaggctg ttcttttgat aataatttac cttgtattgc agaaaaagaa
840gtatttgttt ttgagaatgt tgcagatgat ttaatatcta acatgctgaa
aaataatgct 900gtaattatca atgaagatca ggtatcaaaa ttaatcgatt
tagtattaca aaaaaataat 960gaaactcaag aatactttat caacaaaaaa
tgggtaggta aagatgcaaa attattcctc 1020gatgaaatcg atgttgagtc
tccttcaaat gttaaatgca ttatctgcga agtgaatgcc 1080aatcatccat
ttgttatgac agaactgatg atgccaatat tgccaattgt gcgcgttaaa
1140gatatcgatg aagctattaa atatgcaaag attgcagaac aaaatagaaa
acatagtgcc 1200tatatttata gcaaaaatat cgacaacctg aatcgctttg
aacgtgaaat cgatactact 1260atttttgtaa agaatgctaa atcttttgct
ggtgttggtt atgaagcaga aggatttacc 1320actttcacta ttgctggatc
tactggtgag ggcataacct ctgcacgtaa ttttacccgc 1380caacgtcgct
gtgtactggc cggctaa 1407731407DNAArtificial SequenceDescription of
Artificial Sequence Synthetic polynucleotide 73atgaataaag
acacgctgat cccgacaact aaagatctga aagtaaaaac caatggtgaa 60aacattaatc
tgaagaacta caaagataat agcagttgtt tcggcgtatt cgaaaatgtt
120gaaaatgcta tcagcagcgc ggtacacgca caaaagatac tctcgctgca
ttataccaaa 180gagcaacgtg aaaaaatcat cactgagatc cgtaaggccg
cattacaaaa taaagaggtg 240ctggcaacaa tgattctgga agaaacacat
atgggacgtt atgaggataa aatactgaaa 300catgaactgg tggcgaaata
tacgcctggt actgaagatt taaccaccac tgcctggagc 360ggtgataatg
gtctgaccgt tgtggaaatg tcgccttatg gtgttattgg tgcaattacg
420ccttcaacca atccaactga aacggtaatt tgtaatagca ttggcatgat
tgctgctgga 480aatgcggtag tatttaacgg tcacccctgc gctaaaaaat
gtgttgcctt tgctgttgaa 540atgatcaata aagcgattat tagctgtggc
ggtccggaaa atctggtaac cactataaaa 600aatccaacca tggagtcgct
ggatgccatt attaagcatc cttcaatcaa actgctgtgc 660ggcactggcg
gtccaggaat ggtgaaaacc ctgctgaata gcggtaagaa agcgattggt
720gctggtgctg gaaatccacc agttattgtc gatgatactg ctgatattga
aaaagcgggt 780cgtagcatca ttgaaggctg ttcttttgat aataatttac
cttgtattgc agaaaaagaa 840gtatttgttt ttgagaatgt tgccgatgat
ctgatctcta acatgctgaa aaataatgcg 900gtgattatca atgaagatca
ggttagcaaa ctgatcgatc tggtattaca aaaaaataat 960gaaactcaag
aatactttat caacaaaaaa tgggtaggta aagatgcaaa actgttcctc
1020gatgaaatcg atgttgagtc gccttcaaat gttaaatgca ttatctgcga
agtgaatgcc 1080aatcatccat ttgtgatgac cgaactgatg atgccaattt
tgccgattgt gcgcgttaaa 1140gatatcgatg aagcgattaa atatgcaaag
attgcagaac aaaatcgtaa acatagtgcc 1200tatatttata gcaaaaatat
cgacaacctg aatcgctttg aacgtgaaat cgataccact 1260atttttgtga
agaatgctaa atcttttgct ggtgttggtt atgaagcaga aggttttacc
1320actttcacta ttgctggaag caccggtgaa ggcattacct ctgcacgtaa
ttttacccgc 1380caacgtcgct gtgtactggc cggctaa
1407741407DNAArtificial SequenceDescription of Artificial Sequence
Synthetic polynucleotide 74atgaataaag atacgctgat cccgaccacc
aaagatctga aagtgaaaac caacggcgaa 60aatatcaacc tgaaaaacta taaagataac
agcagttgct ttggcgtgtt tgaaaacgtt 120gaaaacgcca tctccagcgc
ggtgcatgcg caaaaaattc tctcgctgca ttacaccaaa 180gagcagcgtg
aaaaaattat caccgaaatc cgtaaagcgg cgctgcaaaa caaagaagtg
240ctggcaacca tgatcctgga agaaacgcat atggggcgtt atgaagataa
aattctgaaa 300catgaactgg tggcgaaata cacgccgggc actgaagatc
tgaccaccac cgcctggagc 360ggcgataacg gcctgaccgt ggtggagatg
tcgccttatg gcgtgattgg cgcgattacg 420ccgtcaacca acccgaccga
aacggtgatt tgtaacagca ttggcatgat tgccgcgggt 480aatgcggtgg
tgtttaacgg tcatccctgc gcgaaaaaat gtgtggcgtt tgccgttgag
540atgatcaaca aagcgattat cagctgcggc ggcccggaaa atctggtgac
caccatcaaa 600aatccgacca tggaatcgct ggatgccatt atcaaacatc
cttccatcaa actgctgtgc 660ggcaccggcg gcccgggcat ggtgaaaacg
ctgctgaaca gcggtaaaaa agcgattggc 720gcgggcgcgg gtaacccgcc
ggtgattgtc gatgacaccg ccgatattga aaaagcgggg 780cgtagcatta
ttgaaggctg ttcttttgat aacaacctgc cctgcattgc cgaaaaagaa
840gtgtttgtct ttgaaaacgt cgccgatgat ctgatcagca atatgctgaa
aaacaacgcg 900gtgattatca atgaagatca ggttagcaaa ctgatcgatc
tggtgctgca aaaaaacaac 960gaaacgcagg aatattttat caacaaaaaa
tgggttggta aagatgccaa actgtttctc 1020gatgaaatcg atgttgaatc
gccgtctaac gtgaaatgta ttatctgcga agtgaacgcc 1080aaccatccgt
ttgtgatgac cgaactgatg atgccgattc tgccgattgt gcgcgtgaaa
1140gatatcgatg aagcgattaa atatgccaaa attgccgaac aaaaccgtaa
acacagcgcc 1200tatatttaca gcaaaaatat cgataacctg aaccgctttg
aacgtgaaat cgataccacc 1260atttttgtga aaaatgccaa aagttttgcc
ggcgttggtt atgaagcgga aggttttacc 1320acctttacca ttgccggtag
caccggcgaa ggcattacca gcgcccgtaa ttttacccgc 1380cagcgtcgct
gcgtgctggc gggctaa 1407751023DNAGeobacillus thermoglucosidasius
75atgaaagctg cagtagtaga gcaatttaag gaaccattaa aaattaaaga agtggaaaag
60ccatctattt catatggcga agtattagtc cgcattaaag catgcggtgt atgccatacg
120gacttgcacg ccgctcatgg cgattggcca gtaaaaccaa aacttccttt
aatccctggc 180catgaaggag tcggaattgt tgaagaagtc ggtccggggg
taacccattt aaaagtggga 240gaccgcgttg gaattccttg gttatattct
gcgtgcggcc attgcgaata ttgtttaagc 300ggacaagaag cattatgtga
acatcaacaa aacgccggct actcagtcga cgggggttat 360gcagaatatt
gcagagctgc gccagattat gtggtgaaaa ttcctgacaa cttatcgttt
420gaagaagctg ctcctatttt ctgcgccgga gttactactt ataaagcgtt
aaaagtcaca 480ggtacaaaac cgggagaatg ggtagcgatc tatggcatcg
gcggccttgg acatgttgcc 540gtccagtatg cgaaagcgat ggggcttcat
gttgttgcag tggatatcgg cgatgagaaa 600ctggaacttg caaaagagct
tggcgccgat cttgttgtaa atcctgcaaa agaaaatgcg 660gcccaattta
tgaaagagaa agtcggcgga gtacacgcgg ctgttgtgac agctgtatct
720aaacctgctt ttcaatctgc gtacaattct atccgcagag gcggcacgtg
cgtgcttgtc 780ggattaccgc cggaagaaat gcctattcca atctttgata
cggtattaaa cggaattaaa 840attatcggtt ccattgtcgg cacgcggaaa
gacttgcaag aagcgcttca gttcgctgca 900gaaggtaaag taaaaaccat
tattgaagtg caacctcttg aaaaaattaa cgaagtattt 960gacagaatgc
taaaaggaga aattaacgga cgggttgttt taacgttaga aaataataat 1020taa
102376340PRTGeobacillus thermoglucosidasius 76Met Lys Ala Ala Val
Val Glu Gln Phe Lys Glu Pro Leu Lys Ile Lys 1 5 10 15 Glu Val Glu
Lys Pro Ser Ile Ser Tyr Gly Glu Val Leu Val Arg Ile 20 25 30 Lys
Ala Cys Gly Val Cys His Thr Asp Leu His Ala Ala His Gly Asp 35 40
45 Trp Pro Val Lys Pro Lys Leu Pro Leu Ile Pro Gly His Glu Gly Val
50 55 60 Gly Ile Val Glu Glu Val Gly Pro Gly Val Thr His Leu Lys
Val Gly 65 70 75 80 Asp Arg Val Gly Ile Pro Trp Leu Tyr Ser Ala Cys
Gly His Cys Glu 85 90 95 Tyr Cys Leu Ser Gly Gln Glu Ala Leu Cys
Glu His Gln Gln Asn Ala 100 105 110 Gly Tyr Ser Val Asp Gly Gly Tyr
Ala Glu Tyr Cys Arg Ala Ala Pro 115 120 125 Asp Tyr Val Val Lys Ile
Pro Asp Asn Leu Ser Phe Glu Glu Ala Ala 130 135 140 Pro Ile Phe Cys
Ala Gly Val Thr Thr Tyr Lys Ala Leu Lys Val Thr 145 150 155 160 Gly
Thr Lys Pro Gly Glu Trp Val Ala Ile Tyr Gly Ile Gly Gly Leu 165 170
175 Gly His Val Ala Val Gln Tyr Ala Lys Ala Met Gly Leu His Val Val
180 185 190 Ala Val Asp Ile Gly Asp Glu Lys Leu Glu Leu Ala Lys Glu
Leu Gly 195 200 205 Ala Asp Leu Val Val Asn Pro Ala Lys Glu Asn Ala
Ala Gln Phe Met 210 215 220 Lys Glu Lys Val Gly Gly Val His Ala Ala
Val Val Thr Ala Val Ser 225 230 235 240 Lys Pro Ala Phe Gln Ser Ala
Tyr Asn Ser Ile Arg Arg Gly Gly Thr 245 250 255 Cys Val Leu Val Gly
Leu Pro Pro Glu Glu Met Pro Ile Pro Ile Phe 260 265 270 Asp Thr Val
Leu Asn Gly Ile Lys Ile Ile Gly Ser Ile Val Gly Thr 275 280 285 Arg
Lys Asp Leu Gln Glu Ala Leu Gln Phe Ala Ala Glu Gly Lys Val 290 295
300 Lys Thr Ile Ile Glu Val Gln Pro Leu Glu Lys Ile Asn Glu Val Phe
305 310 315 320 Asp Arg Met Leu Lys Gly Glu Ile Asn Gly Arg Val Val
Leu Thr Leu 325 330 335 Glu Asn Asn Asn 340 774090DNAArtificial
SequenceDescription of Artificial Sequence Synthetic polynucleotide
77atggctatcg aaatcaaagt accggacatc ggggctgatg aagttgaaat caccgagatc
60ctggtcaaag tgggcgacaa agttgaagcc gaacagtcgc tgatcaccgt agaaggcgac
120aaagcctcta tggaagttcc gtctccgcag gcgggtatcg ttaaagagat
caaagtctct 180gttggcgata aaacccagac cggcgcactg attatgattt
tcgattccgc cgacggtgca 240gcagacgctg cacctgctca ggcagaagag
aagaaagaag cagctccggc agcagcacca 300gcggctgcgg cggcaaaaga
cgttaacgtt ccggatatcg gcagcgacga agttgaagtg 360accgaaatcc
tggtgaaagt tggcgataaa gttgaagctg aacagtcgct gatcaccgta
420gaaggcgaca aggcttctat ggaagttccg gctccgtttg ctggcaccgt
gaaagagatc 480aaagtgaacg tgggtgacaa agtgtctacc ggctcgctga
ttatggtctt cgaagtcgcg 540ggtgaagcag gcgcggcagc tccggccgct
aaacaggaag cagctccggc agcggcccct 600gcaccagcgg ctggcgtgaa
agaagttaac gttccggata tcggcggtga cgaagttgaa 660gtgactgaag
tgatggtgaa agtgggcgac aaagttgccg ctgaacagtc actgatcacc
720gtagaaggcg acaaagcttc tatggaagtt ccggcgccgt ttgcaggcgt
cgtgaaggaa 780ctgaaagtca acgttggcga taaagtgaaa actggctcgc
tgattatgat cttcgaagtt 840gaaggcgcag cgcctgcggc agctcctgcg
aaacaggaag cggcagcgcc ggcaccggca 900gcaaaagctg aagccccggc
agcagcacca gctgcgaaag cggaaggcaa atctgaattt 960gctgaaaacg
acgcttatgt tcacgcgact ccgctgatcc gccgtctggc acgcgagttt
1020ggtgttaacc ttgcgaaagt gaagggcact ggccgtaaag gtcgtatcct
gcgcgaagac 1080gttcaggctt acgtgaaaga agctatcaaa cgtgcagaag
cagctccggc agcgactggc 1140ggtggtatcc ctggcatgct gccgtggccg
aaggtggact tcagcaagtt tggtgaaatc 1200gaagaagtgg aactgggccg
catccagaaa atctctggtg cgaacctgag ccgtaactgg 1260gtaatgatcc
cgcatgttac tcacttcgac aaaaccgata tcaccgagtt ggaagcgttc
1320cgtaaacagc agaacgaaga agcggcgaaa cgtaagctgg atgtgaagat
caccccggtt 1380gtcttcatca tgaaagccgt tgctgcagct cttgagcaga
tgcctcgctt caatagttcg 1440ctgtcggaag acggtcagcg tctgaccctg
aagaaataca tcaacatcgg tgtggcggtg 1500gataccccga acggtctggt
tgttccggta ttcaaagacg tcaacaagaa aggcatcatc 1560gagctgtctc
gcgagctgat gactatttct aagaaagcgc gtgacggtaa gctgactgcg
1620ggcgaaatgc agggcggttg cttcaccatc tccagcatcg gcggcctggg
tactacccac 1680ttcgcgccga ttgtgaacgc gccggaagtg gctatcctcg
gcgtttccaa gtccgcgatg 1740gagccggtgt ggaatggtaa agagttcgtg
ccgcgtctga tgctgccgat ttctctctcc 1800ttcgaccacc gcgtgatcga
cggtgctgat ggtgcccgtt tcattaccat cattaacaac 1860acgctgtctg
acattcgccg tctggtgatg taagtaaaag agccggccca acggccggct
1920tttttctggt aatctcatga atgtattgag gttattagcg aatagacaaa
tcggttgccg 1980tttgttgttt aaaaattgtt aacaattttg taaaataccg
acggatagaa cgacccggtg 2040gtggttaggg tattacttca cataccctat
ggatttctgg gtgcagcaag gtagcaagcg 2100ccagaatccc caggagctta
cataagtaag tgactggggt gagggcgtga agctaacgcc 2160gctgcggcct
gaaagacgac gggtatgacc gccggagata aatatataga ggtcatgatg
2220agtactgaaa tcaaaactca ggtcgtggta cttggggcag gccccgcagg
ttactccgct 2280gccttccgtt gcgctgattt aggtctggaa accgtaatcg
tagaacgtta caacaccctt 2340ggcggtgttt gtctgaacgt gggttgtatc
ccttctaaag cgctgctgca cgtggcaaaa 2400gttatcgaag aagcgaaagc
gctggccgaa cacggcatcg ttttcggcga accgaaaact 2460gacattgaca
agatccgcac ctggaaagaa aaagtcatca ctcagctgac cggtggtctg
2520gctggcatgg ccaaaggtcg taaagtgaag gtggttaacg gtctgggtaa
atttaccggc 2580gctaacaccc tggaagtgga aggcgaaaac ggcaaaaccg
tgatcaactt cgacaacgcc 2640atcatcgcgg cgggttcccg tccgattcag
ctgccgttta tcccgcatga agatccgcgc 2700gtatgggact ccaccgacgc
gctggaactg aaatctgtac cgaaacgcat gctggtgatg 2760ggcggcggta
tcatcggtct ggaaatgggt accgtatacc atgcgctggg ttcagagatt
2820gacgtggtgg aaatgttcga ccaggttatc ccggctgccg acaaagacgt
ggtgaaagtc 2880ttcaccaaac gcatcagcaa gaaatttaac ctgatgctgg
aagccaaagt gactgccgtt 2940gaagcgaaag aagacggtat ttacgtttcc
atggaaggta aaaaagcacc ggcggaagcg 3000cagcgttacg acgcagtgct
ggtcgctatc ggccgcgtac cgaatggtaa aaacctcgat 3060gcaggtaaag
ctggcgtgga agttgacgat cgcggcttca tccgcgttga caaacaaatg
3120cgcaccaacg tgccgcacat ctttgctatc ggcgatatcg tcggtcagcc
gatgctggcg 3180cacaaaggtg tccatgaagg ccacgttgcc gcagaagtta
tctccggtct gaaacactac 3240ttcgatccga aagtgatccc atccatcgcc
tacactaaac cagaagtggc atgggtcggt 3300ctgaccgaga aagaagcgaa
agagaaaggc atcagctacg aaaccgccac cttcccgtgg 3360gctgcttccg
gccgtgctat cgcttctgac tgcgcagatg gtatgaccaa actgatcttc
3420gacaaagaga cccaccgtgt tatcggcggc gcgattgtcg gcaccaacgg
cggcgagctg 3480ctgggtgaga tcggcctggc tatcgagatg ggctgtgacg
ctgaagacat cgccctgacc 3540atccacgctc acccgactct gcacgagtcc
gttggcctgg cggcggaagt gttcgaaggc 3600agcatcaccg acctgccaaa
cgccaaagcg aagaaaaagt aactttttct ttcaggaaaa 3660aagcataagc
ggctccggga gccgcttttt ttatgcctga tgtttagaac tatgtcactg
3720ttcataaacc gctacacctc atacatactt taagggcgaa ttctgcagat
atccatcaca 3780ctggcggccg ctcgagcatg catctagcac atccggcaat
taaaaaagcg gctaaccacg 3840ccgctttttt tacgtctgca atttaccttt
ccagtcttct tgctccacgt tcagagagac 3900gttcgcatac tgctgaccgt
tgctcgttat tcagcctgac agtatggtta ctgtcgttta 3960gacgttgtgg
gcggctctcc tgaactttct cccgaaaaac ctgacgttgt tcaggtgatg
4020ccgattgaac acgctggcgg gcgttatcac gttgctgttg attcagtggg
cgctgctgta 4080ctttttcctt 409078475PRTEscherichia coli 78Met Met
Ser Thr Glu Ile Lys Thr Gln Val Val Val Leu Gly Ala Gly 1 5 10 15
Pro Ala Gly Tyr Ser Ala Ala Phe Arg Cys Ala Asp Leu Gly Leu Glu 20
25 30 Thr Val Ile Val Glu Arg Tyr Asn Thr Leu Gly Gly Val Cys Leu
Asn 35 40 45 Val Gly Cys Ile Pro Ser Lys Ala Leu Leu His Val Ala
Lys Val Ile 50 55 60 Glu Glu Ala Lys Ala Leu Ala Glu His Gly Ile
Val Phe Gly Glu Pro 65 70 75 80 Lys Thr Asp Ile Asp Lys Ile Arg Thr
Trp Lys Glu Lys Val Ile Asn 85 90 95 Gln Leu Thr Gly Gly Leu Ala
Gly Met Ala Lys Gly Arg Lys Val Lys 100 105 110 Val Val Asn Gly Leu
Gly Lys Phe Thr Gly Ala Asn Thr Leu Glu Val 115 120 125 Glu Gly Glu
Asn Gly Lys Thr Val Ile Asn Phe Asp Asn Ala Ile Ile 130 135 140 Ala
Ala Gly Ser Arg Pro Ile Gln Leu Pro Phe Ile Pro His Glu Asp 145 150
155 160 Pro Arg Ile Trp Asp Ser Thr Asp Ala Leu Glu Leu Lys Glu Val
Pro 165 170 175 Glu Arg Leu Leu Val Met Gly Gly Gly Ile Ile Gly Leu
Glu Met Gly 180 185 190 Thr Val Tyr His Ala Leu Gly Ser Gln Ile Asp
Val Val Glu Met Phe 195 200 205 Asp Gln Val Ile Pro Ala Ala Asp Lys
Asp Ile Val Lys Val Phe Thr 210 215 220 Lys Arg Ile Ser Lys Lys Phe
Asn Leu Met Leu Glu Thr Lys Val Thr 225 230 235 240 Ala Val Glu Ala
Lys Glu Asp Gly Ile Tyr Val Thr Met Glu Gly Lys 245 250 255 Lys Ala
Pro Ala Glu Pro Gln Arg Tyr Asp Ala Val Leu Val Ala Ile 260 265 270
Gly Arg Val Pro Asn Gly Lys Asn Leu Asp Ala Gly Lys Ala Gly Val 275
280 285 Glu Val Asp Asp Arg Gly Phe Ile Arg Val Asp Lys Gln Leu Arg
Thr 290 295 300 Asn Val Pro His Ile Phe Ala Ile Gly Asp Ile Val Gly
Gln Pro Met 305 310 315 320 Leu Ala His Lys Gly Val His Glu Gly His
Val Ala Ala Glu Val Ile 325 330 335 Ala Gly Lys Lys His Tyr Phe Asp
Pro Lys Val Ile Pro Ser Ile Ala 340 345 350 Tyr Thr Glu Pro Glu Val
Ala Trp Val Gly Leu Thr Glu Lys Glu Ala 355 360 365 Lys Glu Lys Gly
Ile Ser Tyr Glu Thr Ala Thr Phe Pro Trp Ala Ala 370 375 380 Ser Gly
Arg Ala Ile Ala Ser Asp Cys Ala Asp Gly Met Thr Lys Leu 385 390 395
400 Ile Phe Asp Lys Glu Ser His Arg Val Ile Gly Gly Ala Ile Val Gly
405 410 415 Thr Asn Gly Gly Glu Leu Leu Gly Glu Ile Gly Leu Ala Ile
Glu Met 420 425 430 Gly Cys Asp Ala Glu Asp Ile Ala Leu Thr Ile His
Ala His Pro Thr 435 440 445 Leu His Glu Ser Val Gly Leu Ala Ala Glu
Val Phe Glu Gly Ser Ile 450 455 460 Thr Asp Leu Pro Asn Pro Lys Ala
Lys Lys Lys 465 470 475 79475PRTKlebsiella pneumoniae 79Met Met Ser
Thr Glu Ile Lys Thr Gln Val Val Val Leu Gly Ala Gly 1 5 10 15 Pro
Ala Gly Tyr Ser Ala Ala Phe Arg Cys Ala Asp Leu Gly Leu Glu 20 25
30 Thr Val Ile Val Glu Arg Tyr Ser Thr Leu Gly Gly Val Cys Leu Asn
35 40 45 Val Gly Cys Ile Pro Ser Lys Ala Leu Leu His Val Ala Lys
Val Ile 50 55 60 Glu Glu Ala Lys Ala Leu Ala Glu His Gly Ile Val
Phe Gly Glu Pro 65 70 75 80 Lys Thr Asp Ile Asp Lys Ile Arg Thr Trp
Lys Glu Lys Val Ile Thr 85 90 95 Gln Leu Thr Gly Gly Leu Ala Gly
Met Ala Lys Gly Arg Lys Val Lys 100 105 110 Val Val Asn Gly Leu Gly
Lys Phe Thr Gly Ala Asn Thr Leu Glu Val 115 120 125 Glu Gly Glu Asn
Gly Lys Thr Val Ile Asn Phe Asp Asn Ala Ile Ile 130 135 140 Ala Ala
Gly Ser Arg Pro Ile Gln Leu Pro Phe Ile Pro His Glu Asp 145 150 155
160 Pro Arg Val Trp Asp Ser Thr Asp Ala Leu Glu Leu Lys Ser Val Pro
165 170 175 Lys Arg Met Leu Val Met Gly Gly Gly Ile Ile Gly Leu Glu
Met Gly 180 185 190 Thr Val Tyr His Ala Leu Gly Ser Glu Ile Asp Val
Val Glu Met Phe 195 200 205 Asp Gln Val Ile Pro Ala Ala Asp Lys Asp
Val Val Lys Val Phe Thr 210 215 220
Lys Arg Ile Ser Lys Lys Phe Asn Leu Met Leu Glu Ala Lys Val Thr 225
230 235 240 Ala Val Glu Ala Lys Glu Asp Gly Ile Tyr Val Ser Met Glu
Gly Lys 245 250 255 Lys Ala Pro Ala Glu Ala Gln Arg Tyr Asp Ala Val
Leu Val Ala Ile 260 265 270 Gly Arg Val Pro Asn Gly Lys Asn Leu Asp
Ala Gly Lys Ala Gly Val 275 280 285 Glu Val Asp Asp Arg Gly Phe Ile
Arg Val Asp Lys Gln Met Arg Thr 290 295 300 Asn Val Pro His Ile Phe
Ala Ile Gly Asp Ile Val Gly Gln Pro Met 305 310 315 320 Leu Ala His
Lys Gly Val His Glu Gly His Val Ala Ala Glu Val Ile 325 330 335 Ser
Gly Leu Lys His Tyr Phe Asp Pro Lys Val Ile Pro Ser Ile Ala 340 345
350 Tyr Thr Lys Pro Glu Val Ala Trp Val Gly Leu Thr Glu Lys Glu Ala
355 360 365 Lys Glu Lys Gly Ile Ser Tyr Glu Thr Ala Thr Phe Pro Trp
Ala Ala 370 375 380 Ser Gly Arg Ala Ile Ala Ser Asp Cys Ala Asp Gly
Met Thr Lys Leu 385 390 395 400 Ile Phe Asp Lys Glu Thr His Arg Val
Ile Gly Gly Ala Ile Val Gly 405 410 415 Thr Asn Gly Gly Glu Leu Leu
Gly Glu Ile Gly Leu Ala Ile Glu Met 420 425 430 Gly Cys Asp Ala Glu
Asp Ile Ala Leu Thr Ile His Ala His Pro Thr 435 440 445 Leu His Glu
Ser Val Gly Leu Ala Ala Glu Val Phe Glu Gly Ser Ile 450 455 460 Thr
Asp Leu Pro Asn Ala Lys Ala Lys Lys Lys 465 470 475
80347DNAArtificial SequenceDescription of Artificial Sequence
Synthetic polynucleotide 80ataataatac atatgaacca tgcgagttac
gggcctataa gccaggcgag atatgatcta 60tatcaatttc tcatctataa tgctttgtta
gtatctcgtc gccgacttaa taaagagaga 120gttagtgtga aagctgacaa
cccttttgat cttttacttc ctgctgcaat ggccaaagtg 180gccgaagagg
cgggtgtcta taaagcaacg aaacatccgc ttaagacttt ctatctggcg
240attaccgccg gtgttttcat ctcaatcgca ttcaccactg gcacaggcac
agaaggtagg 300tgttacatgt cagaacgttt acacaatgac gtggatccta ttattat
347814678DNAArtificial SequenceDescription of Artificial Sequence
Synthetic polynucleotide 81aagaggtaaa agaataatgg ctatcgaaat
caaagtaccg gacatcgggg ctgatgaagt 60tgaaatcacc gagatcctgg tcaaagtggg
cgacaaagtt gaagccgaac agtcgctgat 120caccgtagaa ggcgacaaag
cctctatgga agttccgtct ccgcaggcgg gtatcgttaa 180agagatcaaa
gtctctgttg gcgataaaac ccagaccggc gcactgatta tgattttcga
240ttccgccgac ggtgcagcag acgctgcacc tgctcaggca gaagagaaga
aagaagcagc 300tccggcagca gcaccagcgg ctgcggcggc aaaagacgtt
aacgttccgg atatcggcag 360cgacgaagtt gaagtgaccg aaatcctggt
gaaagttggc gataaagttg aagctgaaca 420gtcgctgatc accgtagaag
gcgacaaggc ttctatggaa gttccggctc cgtttgctgg 480caccgtgaaa
gagatcaaag tgaacgtggg tgacaaagtg tctaccggct cgctgattat
540ggtcttcgaa gtcgcgggtg aagcaggcgc ggcagctccg gccgctaaac
aggaagcagc 600tccggcagcg gcccctgcac cagcggctgg cgtgaaagaa
gttaacgttc cggatatcgg 660cggtgacgaa gttgaagtga ctgaagtgat
ggtgaaagtg ggcgacaaag ttgccgctga 720acagtcactg atcaccgtag
aaggcgacaa agcttctatg gaagttccgg cgccgtttgc 780aggcgtcgtg
aaggaactga aagtcaacgt tggcgataaa gtgaaaactg gctcgctgat
840tatgatcttc gaagttgaag gcgcagcgcc tgcggcagct cctgcgaaac
aggaagcggc 900agcgccggca ccggcagcaa aagctgaagc cccggcagca
gcaccagctg cgaaagcgga 960aggcaaatct gaatttgctg aaaacgacgc
ttatgttcac gcgactccgc tgatccgccg 1020tctggcacgc gagtttggtg
ttaaccttgc gaaagtgaag ggcactggcc gtaaaggtcg 1080tatcctgcgc
gaagacgttc aggcttacgt gaaagaagct atcaaacgtg cagaagcagc
1140tccggcagcg actggcggtg gtatccctgg catgctgccg tggccgaagg
tggacttcag 1200caagtttggt gaaatcgaag aagtggaact gggccgcatc
cagaaaatct ctggtgcgaa 1260cctgagccgt aactgggtaa tgatcccgca
tgttactcac ttcgacaaaa ccgatatcac 1320cgagttggaa gcgttccgta
aacagcagaa cgaagaagcg gcgaaacgta agctggatgt 1380gaagatcacc
ccggttgtct tcatcatgaa agccgttgct gcagctcttg agcagatgcc
1440tcgcttcaat agttcgctgt cggaagacgg tcagcgtctg accctgaaga
aatacatcaa 1500catcggtgtg gcggtggata ccccgaacgg tctggttgtt
ccggtattca aagacgtcaa 1560caagaaaggc atcatcgagc tgtctcgcga
gctgatgact atttctaaga aagcgcgtga 1620cggtaagctg actgcgggcg
aaatgcaggg cggttgcttc accatctcca gcatcggcgg 1680cctgggtact
acccacttcg cgccgattgt gaacgcgccg gaagtggcta tcctcggcgt
1740ttccaagtcc gcgatggagc cggtgtggaa tggtaaagag ttcgtgccgc
gtctgatgct 1800gccgatttct ctctccttcg accaccgcgt gatcgacggt
gctgatggtg cccgtttcat 1860taccatcatt aacaacacgc tgtctgacat
tcgccgtctg gtgatgtaag taaaagagcc 1920ggcccaacgg ccggcttttt
tctggtaatc tcatgaatgt attgaggtta ttagcgaata 1980gacaaatcgg
ttgccgtttg ttaagccagg cgagatatga tctatatcaa tttctcatct
2040ataatgcttt gttagtatct cgtcgccgac ttaataaaga gagagttagt
cttctatatc 2100acagcaagaa ggtaggtgtt acatgatgag tactgaaatc
aaaactcagg tcgtggtact 2160tggggcaggc cccgcaggtt actctgcagc
cttccgttgc gctgatttag gtctggaaac 2220cgtcatcgta gaacgttaca
gcaccctcgg tggtgtttgt ctgaacgtgg gttgtatccc 2280ttctaaagcg
ctgctgcacg tggcaaaagt tatcgaagaa gcgaaagcgc tggccgaaca
2340cggcatcgtt ttcggcgaac cgaaaactga cattgacaag atccgcacct
ggaaagaaaa 2400agtcatcact cagctgaccg gtggtctggc tggcatggcc
aaaggtcgta aagtgaaggt 2460ggttaacggt ctgggtaaat ttaccggcgc
taacaccctg gaagtggaag gcgaaaacgg 2520caaaaccgtg atcaacttcg
acaacgccat catcgcggcg ggttcccgtc cgattcagct 2580gccgtttatc
ccgcatgaag atccgcgcgt atgggactcc accgacgcgc tggaactgaa
2640atctgtaccg aaacgcatgc tggtgatggg cggcggtatc atcggtctgg
aaatgggtac 2700cgtataccat gcgctgggtt cagagattga cgtggtggaa
atgttcgacc aggttatccc 2760ggctgccgac aaagacgtgg tgaaagtctt
caccaaacgc atcagcaaga aatttaacct 2820gatgctggaa gccaaagtga
ctgccgttga agcgaaagaa gacggtattt acgtttccat 2880ggaaggtaaa
aaagcaccgg cggaagcgca gcgttacgac gcagtgctgg tcgctatcgg
2940ccgcgtaccg aatggtaaaa acctcgatgc aggtaaagct ggcgtggaag
ttgacgatcg 3000cggcttcatc cgcgttgaca aacaaatgcg caccaacgtg
ccgcacatct ttgctatcgg 3060cgatatcgtc ggtcagccga tgctggcgca
caaaggtgtc catgaaggcc acgttgccgc 3120agaagttatc tccggtctga
aacactactt cgatccgaaa gtgatcccat ccatcgccta 3180cactaaacca
gaagtggcat gggtcggtct gaccgagaaa gaagcgaaag agaaaggcat
3240cagctacgaa accgccacct tcccgtgggc tgcttccggc cgtgctatcg
cttctgactg 3300cgcagatggt atgaccaaac tgatcttcga caaagagacc
caccgtgtta tcggcggcgc 3360gattgtcggc accaacggcg gcgagctgct
gggtgagatc ggcctggcta tcgagatggg 3420ctgtgacgct gaagacatcg
ccctgaccat ccacgctcac ccgactctgc acgagtccgt 3480tggcctggcg
gcggaagtgt tcgaaggcag catcaccgac ctgccaaacg ccaaagcgaa
3540gaaaaagtaa ctttttcttt caggaaaaaa gcataagcgg ctccgggagc
cgcttttttt 3600atgcctgatg tttagaacta tgtcactgtt cataaaccgc
tacacctcat acatacttta 3660agggcgaatt ctgcagatat ccatcacact
ggcggccgct cgagcatgca tctagcacat 3720ccggcaatta aaaaagcggc
taaccacgcc gcttttttta cgtctgcaat ttacctttcc 3780agtcttcttg
ctccacgttc agagagacgt tcgcatactg ctgaccgttg ctcgttattc
3840agcctgacag tatggttact gtcgtttaga cgttgtgggc ggctctcctg
aactttctcc 3900cgaaaaacct gacgttgttc aggtgatgcc gattgaacac
gctggcgggc gttatcacgt 3960tgctgttgat tcagtgggcg ctgctgtact
ttttccttaa acacctggcg ctgctctggt 4020gatgcggact gaatacgctc
acgcgctgcg tctcttcgct gctggttctg cgggttagtc 4080tgcattttct
cgcgaaccgc ctggcgctgc tcaggcgagg cggactgaat gcgctcacgc
4140gctgcctctc ttcgctgctg gatcttcggg ttagtctgca ttctctcgcg
aactgcctgg 4200cgctgctcag gcgaggcgga ctgataacgc tgacgagcgg
cgtccttttg ttgctgggtc 4260agtggttggc gacggctgaa gtcgtggaag
tcgtcatagc tcccatagtg ttcagcttca 4320ttaaaccgct gtgccgctgc
ctgacgttgg gtacctcgtg taatgactgg tgcggcgtgt 4380gttcgttgct
gaaactgatt tgctgccgcc tgacgctggc tgtcgcgcgt tggggcaggt
4440aattgcgtgg cgctcattcc gccgttgaca tcggtttgat gaaaccgctt
tgccatatcc 4500tgatcatgat agggcacacc attacggtag tttggattgt
gccgccatgc catattctta 4560tcagtaagat gctcaccggt gatacggttg
aaattgttga cgtcgatatt gatgttgtcg 4620ccgttgtgtt gccagccatt
accgtcacga tgaccgccat cgtggtgatg ataatcat 4678821114DNAArtificial
SequenceDescription of Artificial Sequence Synthetic polynucleotide
82caaaaaaccg gagtctgtgc tccggttttt tattatccgc taatcaatta catatgaata
60tcctccttag ttcctattcc gaagttccta ttctctagaa agtataggaa cttcggcgcg
120cctacctgtg acggaagatc acttcgcaga ataaataaat cctggtgtcc
ctgttgatac 180cgggaagccc tgggccaact tttggcgaaa atgagacgtt
gatcggcacg taagaggttc 240caactttcac cataatgaaa taagatcact
accgggcgta ttttttgagt tgtcgagatt 300ttcaggagct aaggaagcta aa atg
gag aaa aaa atc act gga tat acc acc 352 Met Glu Lys Lys Ile Thr Gly
Tyr Thr Thr 1 5 10 gtt gat ata tcc caa tgg cat cgt aaa gaa cat ttt
gag gca ttt cag 400Val Asp Ile Ser Gln Trp His Arg Lys Glu His Phe
Glu Ala Phe Gln 15 20 25 tca gtt gct caa tgt acc tat aac cag acc
gtt cag ctg gat att acg 448Ser Val Ala Gln Cys Thr Tyr Asn Gln Thr
Val Gln Leu Asp Ile Thr 30 35 40 gcc ttt tta aag acc gta aag aaa
aat aag cac aag ttt tat ccg gcc 496Ala Phe Leu Lys Thr Val Lys Lys
Asn Lys His Lys Phe Tyr Pro Ala 45 50 55 ttt att cac att ctt gcc
cgc ctg atg aat gct cat ccg gaa tta cgt 544Phe Ile His Ile Leu Ala
Arg Leu Met Asn Ala His Pro Glu Leu Arg 60 65 70 atg gca atg aaa
gac ggt gag ctg gtg ata tgg gat agt gtt cac cct 592Met Ala Met Lys
Asp Gly Glu Leu Val Ile Trp Asp Ser Val His Pro 75 80 85 90 tgt tac
acc gtt ttc cat gag caa act gaa acg ttt tca tcg ctc tgg 640Cys Tyr
Thr Val Phe His Glu Gln Thr Glu Thr Phe Ser Ser Leu Trp 95 100 105
agt gaa tac cac gac gat ttc cgg cag ttt cta cac ata tat tcg caa
688Ser Glu Tyr His Asp Asp Phe Arg Gln Phe Leu His Ile Tyr Ser Gln
110 115 120 gat gtg gcg tgt tac ggt gaa aac ctg gcc tat ttc cct aaa
ggg ttt 736Asp Val Ala Cys Tyr Gly Glu Asn Leu Ala Tyr Phe Pro Lys
Gly Phe 125 130 135 att gag aat atg ttt ttc gtc tca gcc aat ccc tgg
gtg agt ttc acc 784Ile Glu Asn Met Phe Phe Val Ser Ala Asn Pro Trp
Val Ser Phe Thr 140 145 150 agt ttt gat tta aac gtg gcc aat atg gac
aac ttc ttc gcc ccc gtt 832Ser Phe Asp Leu Asn Val Ala Asn Met Asp
Asn Phe Phe Ala Pro Val 155 160 165 170 ttc acc atg ggc aaa tat tat
acg caa ggc gac aag gtg ctg atg ccg 880Phe Thr Met Gly Lys Tyr Tyr
Thr Gln Gly Asp Lys Val Leu Met Pro 175 180 185 ctg gcg att cag gtt
cat cat gcc gtt tgt gat ggc ttc cat gtc ggc 928Leu Ala Ile Gln Val
His His Ala Val Cys Asp Gly Phe His Val Gly 190 195 200 aga tgc tta
atg aat aca aca gta ctg cga tgagtggcag ggcggggcgt 978Arg Cys Leu
Met Asn Thr Thr Val Leu Arg 205 210 aaggcgcgcc atttaaatga
agttcctatt ccgaagttcc tattctctag aaagtatagg 1038aacttcgaag
cagctccagc ctacaccctt cttcagggct gactgtttgc ataaaaattc
1098atctgtatgc acaata 111483212PRTArtificial SequenceDescription of
Artificial Sequence Synthetic polypeptide 83Met Glu Lys Lys Ile Thr
Gly Tyr Thr Thr Val Asp Ile Ser Gln Trp 1 5 10 15 His Arg Lys Glu
His Phe Glu Ala Phe Gln Ser Val Ala Gln Cys Thr 20 25 30 Tyr Asn
Gln Thr Val Gln Leu Asp Ile Thr Ala Phe Leu Lys Thr Val 35 40 45
Lys Lys Asn Lys His Lys Phe Tyr Pro Ala Phe Ile His Ile Leu Ala 50
55 60 Arg Leu Met Asn Ala His Pro Glu Leu Arg Met Ala Met Lys Asp
Gly 65 70 75 80 Glu Leu Val Ile Trp Asp Ser Val His Pro Cys Tyr Thr
Val Phe His 85 90 95 Glu Gln Thr Glu Thr Phe Ser Ser Leu Trp Ser
Glu Tyr His Asp Asp 100 105 110 Phe Arg Gln Phe Leu His Ile Tyr Ser
Gln Asp Val Ala Cys Tyr Gly 115 120 125 Glu Asn Leu Ala Tyr Phe Pro
Lys Gly Phe Ile Glu Asn Met Phe Phe 130 135 140 Val Ser Ala Asn Pro
Trp Val Ser Phe Thr Ser Phe Asp Leu Asn Val 145 150 155 160 Ala Asn
Met Asp Asn Phe Phe Ala Pro Val Phe Thr Met Gly Lys Tyr 165 170 175
Tyr Thr Gln Gly Asp Lys Val Leu Met Pro Leu Ala Ile Gln Val His 180
185 190 His Ala Val Cys Asp Gly Phe His Val Gly Arg Cys Leu Met Asn
Thr 195 200 205 Thr Val Leu Arg 210 842521DNAArtificial
SequenceDescription of Artificial Sequence Synthetic polynucleotide
84ttatttggtg atattggtac caatatcatg cagcaaacgg tgcaacattg ccgtgtctcg
60ttgctctaaa agccccaggc gttgttgtaa ccagtcgacc agttttatgt catctgccac
120tgccagagtc gtcagcaatg tcatggctcg ttcgcgtaaa gcttgcagtt
gatgttggtc 180tgccgttgca tcacttttcg ccggttgttg tattaatgtt
gctaattgat agcaatagac 240catcaccgcc tgccccagat tgagcgaagg
ataatccgcc accatcggca caccagtaag 300aacgtcagcc aacgctaact
cttcgttagt caacccggaa tcttcgcgac caaacaccag 360cgcggcatgg
ctcatccatg aagatttttc ctctaacagc ggcaccagtt caactggcgt
420ggcgtagtaa tgatatttcg cccgactgcg cgcagtggtg gcgacagtga
aatcgacatc 480gtgtaacgat tcagccaatg tcgggaaaac tttaatatta
tcaataatat caccagatcc 540atgtgcgacc cagcgggtgg ctggctccag
gtgtgcctga ctatcgacaa tccgcagatc 600gctaaacccc atcgttttca
ttgcccgcgc cgctgcccca atattttctg ctctggcggg 660tgcgaccaga
ataatcgtta tacgcatatt gccactcttc ttgatcaaat aaccgcgaac
720cgggtgatca ctgtcaactt attacgcggt gcgaatttac aaattcttaa
cgtaagtcgc 780agaaaaagcc ctttacttag cttaaaaaag gctaaactat
ttcctgactg tactaacggt 840tgagttgtta aaaaatgcta catatccttc
tgtttactta ggataatttt ataaaaaata 900aatctcgaca attggattca
ccacgtttat tagttgtatg atgcaactag ttggattatt 960aaaataatgt
gacgaaagct agcatttaga tacgatgatt tcatcaaact gttaacgtgc
1020tacaattgaa cttgatatat gtcaacgaag cgtagtttta ttgggtgtcc
ggcccctctt 1080agcctgttat gttgctgtta aaatggttag gatgacagcc
gtttttgaca ctgtcgggtc 1140ctgagggaaa gtacccacga ccaagctaat
gatgttgttg acgttgatgg aaagtgcatc 1200aagaacgcaa ttacgtactt
tagtcatgtt acgccgatca tgttaatttg cagcatgcat 1260caggcaggtc
agggactttt gtacttcctg tttcgattta gttggcaatt taggtagcaa
1320acgaattcat cggctttacc accgtcaaaa aaaacggcgc tttttagcgc
cgtttttatt 1380tttcaacctt atttccagat acgtaactca tcgtccgttg
taacttcttt actggctttc 1440attttcggca gtgaaaacgc ataccagtcg
atattacggg tcacaaacat catgccggcc 1500agcgccacca ccagcacact
ggttcccaac aacagcgcgc tatcggcaga gttgagcagt 1560ccccacatca
caccatccag caacaacagc gcgagggtaa acaacatgct gttgcaccaa
1620cctttcaata ccgcttgcaa ataaataccg ttcattatcg ccccaatcag
actggcgatt 1680atccatgcca cggtaaaacc ggtatgttca gaaagcgcca
gcaagagcaa ataaaacatc 1740accaatgaaa gccccaccag caaatattgc
attgggtgta aacgttgcgc ggtgagcgtt 1800tcaaaaacaa agaacgccat
aaaagtcagt gcaatcagca gaatggcgta cttagtcgcc 1860cggtcagtta
attggtattg atcggctggc gtcgttactg cgacgctaaa cgccgggaag
1920ttttcccagc cggtatcatt gcctgaagca aaacgctcac cgagattatt
agcaaaccag 1980ctgctttgcc agtgcgcctg aaaacctgac tcgctaactt
cccgtttggc tggtagaaaa 2040tcacctaaaa aactgggatg cggccagttg
ctggttaagg tcatttcgct attacgcccg 2100ccaggcacca cagaaagatc
gccggtaccg cttaaattca gggccatatt cagcttcagg 2160ttctgcttcc
gccagtcccc ttcaggtaaa gggatatgca cgccctgccc gccttgctct
2220aacccggtgc cgggttcaat ggtcagcgcc gttccgttaa cttcaggcgc
tttcaccaca 2280ccaataccac gcgcatcccc gacgctaatc acaataaatg
gcttgcctaa ggtgatattt 2340ggcgcgttga gttcgctaag acgcgaaaca
tcgaaatcgg cttttaacgt taaatcactg 2400tgccagacct gaccggtata
aatccctatc ttgcgttctt ccacgttctg attgccatca 2460accatcaatg
actcaggtaa ccaaaaatgg ataaaacttc gtttccgctg cagggtttta 2520t
2521853010DNAArtificial SequenceDescription of Artificial Sequence
Synthetic polynucleotide 85aagccacagc aggatgccca ctgcaacaaa
ggtgatcaca ccggaaacgc gatggagaat 60ggacgctatc gccgtgatgg ggaaccggat
ggtctgtagg tccagattaa caggtctttg 120ttttttcaca tttcttatca
tgaataacgc ccacatgctg ttcttattat tccctgggga 180ctacgggcac
agaggttaac tttctgttac ctggagacgt cgggatttcc ttcctccggt
240ctgcttgcgg gtcagacagc gtcctttcta taactgcgcg tcatgcaaaa
cactgcttcc 300agatgcgaaa acgacacgtt acaacgctgg gtggctcggg
attgcagggt gttccggaga 360cctggcggca gtataggctg ttcacaaaat
cattacaatt aacctacata tagtttgtcg 420ggttttatcc tgaacagtga
tccaggtcac gataacaaca tttatttaat ttttaatcat 480ctaatttgac
aatcattcaa caaagttgtt acaaacatta ccaggaaaag catataatgc
540gtaaaagtta tgaagtcggt atttcaccta agattaactt atgtaacagt
gtggaagtat 600tgaccaattc attcgggaca gttattagtg gtagacaagt
ttaataattc ggattgctaa 660gtacttgatt cgccatttat tcgtcatcaa
tggatccttt acctgcaagc gcccagagct 720ctgtacccag gttttcccct
ctttcacaga gcggcgagcc aaataaaaaa cgggtaaagc 780caggttgatg
tgcgaaggca aatttaagtt ccggcagtct tacgcaataa ggcgctaagg
840agaccttaaa tggctgatac aaaagcaaaa ctcaccctca acggggatac
agctgttgaa
900ctggatgtgc tgaaaggcac gctgggtcaa gatgttattg atatccgtac
tctcggttca 960aaaggtgtgt tcacctttga cccaggcttc acttcaaccg
catcctgcga atctaaaatt 1020acttttattg atggtgatga aggtattttg
ctgcaccgcg gtttcccgat cgatcagctg 1080gcgaccgatt ctaactacct
ggaagtttgt tacatcctgc tgaatggtga aaaaccgact 1140caggaacagt
atgacgaatt taaaactacg gtgacccgtc ataccatgat ccacgagcag
1200attacccgtc tgttccatgc tttccgtcgc gactcgcatc caatggcagt
catgtgtggt 1260attaccggcg cgctggcggc gttctatcac gactcgctgg
atgttaacaa tcctcgtcac 1320cgtgaaattg ccgcgttcct cctgctgtcg
aaaatgccga ccatggccgc gatgtgttac 1380aagtattcca ttggtcagcc
atttgtttac ccgcgcaacg atctctccta cgccggtaac 1440ttcctgaata
tgatgttctc cacgccgtgc gaaccgtatg aagttaatcc gattctggaa
1500cgtgctatgg accgtattct gatcctgcac gctgaccatg aacagaacgc
ctctacctcc 1560accgtgcgta ccgctggctc ttcgggtgcg aacccgtttg
cctgtatcgc agcaggtatt 1620gcttcactgt ggggacctgc gcacggcggt
gctaacgaag cggcgctgaa aatgctggaa 1680gaaatcagct ccgttaaaca
cattccggaa tttgttcgtc gtgcgaaaga caaaaatgat 1740tctttccgcc
tgatgggctt cggtcaccgc gtgtacaaaa attacgaccc gcgcgccacc
1800gtaatgcgtg aaacctgcca tgaagtgctg aaagagctgg gcacgaagga
tgacctgctg 1860gaagtggcta tggagctgga aaacatcgcg ctgaacgacc
cgtactttat cgagaagaaa 1920ctgtacccga acgtcgattt ctactctggt
atcatcctga aagcgatggg tattccgtct 1980tccatgttca ccgtcatttt
cgcaatggca cgtaccgttg gctggatcgc ccactggagc 2040gaaatgcaca
gtgacggtat gaagattgcc cgtccgcgtc agctgtatac aggatatgaa
2100aaacgcgact ttaaaagcga tatcaagcgt taatggttga ttgctaagtt
gtaaatattt 2160taacccgccg ttcatatggc gggttgattt ttatatgcct
aaacacaaaa aattgtaaaa 2220ataaaatcca ttaacagacc tatatagata
tttaaaaaga atagaacagc tcaaattatc 2280agcaacccaa tactttcaat
taaaaacttc atggtagtcg catttataac cctatgaaaa 2340tgacgtctat
ctataccccc ctatatttta ttcatcatac aacaaattca tgataccaat
2400aatttagttt tgcatttaat aaaactaaca atatttttaa gcaaaactaa
aaactagcaa 2460taatcaaata cgatattctg gcgtagctat acccctattc
tatatcctta aaggactctg 2520ttatgtttaa aggacaaaaa acattggccg
cactggccgt atctctgctg ttcactgcac 2580ctgtttatgc tgctgatgaa
ggttctggcg aaattcactt taagggggag gttattgaag 2640caccttgtga
aattcatcca gaagatattg ataaaaacat agatcttgga caagtcacga
2700caacccatat aaaccgggag catcatagca ataaagtggc cgtcgacatt
cgcttgatca 2760actgtgatct gcctgcttct gacaacggta gcggaatgcc
ggtatccaaa gttggcgtaa 2820ccttcgatag cacggctaag acaactggtg
ctacgccttt gttgagcaac accagtgcag 2880gcgaagcaac tggggtcggt
gtacgactga tggacaaaaa tgacggtaac atcgtattag 2940gttcagccgc
gccagatctt gacctggatg caagctcatc agaacagacg ctgaactttt
3000tcgcctggat 3010864180DNAArtificial SequenceDescription of
Artificial Sequence Synthetic polynucleotide 86cgcgatgtcg
acgtcacgaa actgaaaaaa ccgctctaca ttctggcgac tgctgatgaa 60gaaaccagta
tggccggagc gcgttatttt gccgaaacta ccgccctgcg cccggattgc
120gccatcattg gcgaaccgac gtcactacaa ccggtacgcg cacataaagg
tcatatctct 180aacgccatcc gtattcaggg ccagtcgggg cactccagcg
atccagcacg cggagttaac 240gctatcgaac taatgcacga cgccatcggg
catattttgc aattgcgcga taacctgaaa 300gaacgttatc actacgaagc
gtttaccgtg ccatacccta cgctcaacct cgggcatatt 360cacggtggcg
acgcttctaa ccgtatttgc gcttgctgtg agttgcatat ggatattcgt
420ccgctgcctg gcatgacact caatgaactt aatggtttgc tcaacgatgc
attggctccg 480gtgagcgaac gctggccggg tcgtctgacg gtcgacgagc
tgcatccgcc gatccctggc 540tatgaatgcc caccgaatca tcaactggtt
gaagtggttg agaaattgct cggagcaaaa 600accgaagtgg tgaactactg
taccgaagcg ccgtttattc aaacgttatg cccgacgctg 660gtgttggggc
ctggctcaat taatcaggct catcaacctg atgaatatct ggaaacacgg
720tttatcaagc ccacccgcga actgataacc caggtaattc accatttttg
ctggcattaa 780aacgtaggcc ggataaggcg ctcgcgccgc atccggcgct
gttgccaaac tccagtgccg 840caataatgtc ggatgcgatg cttgcgcatc
ttatccgacc tacagtgact caaacgatgc 900ccaaccgtag gccggataag
gcgctcgcgc cgcatccggc actgttgcca aactccagtg 960ccgcaataat
gtcggatgcg atacttgcgc atcttatccg accgacagtg actcaaacga
1020tgcccaactg taggccggat aaggcgctcg cgccgcatcc ggcactgttg
ccaaactcca 1080gtgccgcaat aatgtcggat gcgatacttg cgcatcttat
ccgacctaca cctttggtgt 1140tacttggggc gattttttaa catttccata
agttacgctt atttaaagcg tcgtgaattt 1200aatgacgtaa attcctgcta
tttattcgtt tgctgaagcg atttcgcagc atttgacgtc 1260accgctttta
cgtggcttta taaaagacga cgaaaagcaa agcccgagca tattcgcgcc
1320aatgctagca agaggagaag tcgacatgac agacttaaat aaagtggtaa
aagaacttga 1380agctcttggt atttatgacg taaaagaagt tgtttacaat
ccaagctacg agcaattgtt 1440cgaagaagaa actaaaccag gtttagaagg
ctttgaaaaa ggtactttaa ctacgactgg 1500tgcagtggca gtagatacag
gtatcttcac aggtcgttct ccaaaagata aatatatcgt 1560gttagatgaa
aaaaccaaag atactgtttg gtggacatct gaaacagcaa aaaacgacaa
1620caagccaatg aaccaagcta catggcaaag cttaaaagac ttggtaacca
accagctttc 1680tcgtaaacgc ttatttgtag ttgatggttt ctgtggtgcg
agcgaacacg accgtattgc 1740agtacgtatt gtcactgaag tagcgtggca
agcacatttt gtaaaaaata tgtttattcg 1800cccaactgaa gaacaactca
aaaattttga accagatttc gttgtaatga atggttctaa 1860agtaaccaat
ccaaactgga aagaacaagg tttaaattca gaaaactttg ttgctttcaa
1920cttgactgaa cgcattcaat taatcggtgg tacttggtac ggcggtgaaa
tgaaaaaagg 1980tatgttctca atcatgaact acttcctacc acttaaaggt
gttggtgcaa tgcactgctc 2040agctaacgtt ggtaaagatg gcgatgtagc
aatcttcttc ggcttatctg gcacaggtaa 2100aacaaccctt tcaacggatc
caaaacgtga attaatcggt gacgatgaac acggctggga 2160tgatgtgggt
atctttaact ttgaaggtgg ttgctatgcg aaaaccattc acctttcaga
2220agaaaatgaa ccagatattt accgcgctat ccgtcgcgac gcattattag
aaaacgtggt 2280tgttcgtgca gatggttctg ttgatttcga tgatggttca
aaaacagaaa atactcgcgt 2340gtcttaccca atttatcaca ttgataacat
tgtaaaacca gtttctcgtg caggtcacgc 2400aactaaagtg attttcttaa
ctgcagatgc atttggcgta ttaccaccag tatctaaatt 2460gacaccagaa
caaactaaat actacttctt atctggtttc acagcaaaat tagcaggtac
2520tgaacgtggt attactgaac caactccaac tttctcagca tgtttcggtg
ctgcgttctt 2580aacccttcac ccaactcaat atgcagaagt gttagtaaaa
cgtatgcaag cagtgggtgc 2640tgaagcttac ttagtaaata ctggttggaa
tggcacaggc aaacgtatct caatcaaaga 2700tactcgcgga atcattgatg
caatcttaga tggctcaatt gaaaaagctg aaatgggcga 2760attaccaatc
tttaacttag ccattcctaa agcattacca ggtgtagatt ctgcaatctt
2820agatcctcgc gatacttacg cagataaagc acaatggcaa tcaaaagctg
aagacttagc 2880aggtcgtttt gtgaaaaact ttgttaaata tgcaactaac
gaagaaggca aagctttaat 2940tgcagctggt cctaaagctt aatctagaaa
gcttcctaga ggcatcaaat aaaacgaaag 3000gctcagtcga aagactgggc
ctttcgtttt atctgttgtt tgtcggtgaa cgctctcctg 3060agtaggacga
attcacttct gttctaacac cctcgttttc aatatatttc tgtctgcatt
3120ttattcaaat tctgaatata ccttcagata tccttaagga attgtcgtta
cattcggcga 3180tattttttca agacaggttc ttactatgca ttccacagaa
gtccaggcta aacctctttt 3240tagctggaaa gccctgggtt gggcactgct
ctacttttgg tttttctcta ctctgctaca 3300ggccattatt tacatcagtg
gttatagtgg cactaacggc attcgcgact cgctgttatt 3360cagttcgctg
tggttgatcc cggtattcct ctttccgaag cggattaaaa ttattgccgc
3420agtaatcggc gtggtgctat gggcggcctc tctggcggcg ctgtgctact
acgtcatcta 3480cggtcaggag ttctcgcaga gcgttctgtt tgtgatgttc
gaaaccaaca ccaacgaagc 3540cagcgagtat ttaagccagt atttcagcct
gaaaattgtg cttatcgcgc tggcctatac 3600ggcggtggca gttctgctgt
ggacacgcct gcgcccggtc tatattccaa agccgtggcg 3660ttatgttgtc
tcttttgccc tgctttatgg cttgattctg catccgatcg ccatgaatac
3720gtttatcaaa aacaagccgt ttgagaaaac gttggataac ctggcctcgc
gtatggagcc 3780tgccgcaccg tggcaattcc tgaccggcta ttatcagtat
cgtcagcaac taaactcgct 3840aacaaagtta ctgaatgaaa ataatgcctt
gccgccactg gctaatttca aagatgaatc 3900gggtaacgaa ccgcgcactt
tagtgctggt gattggcgag tcgacccagc gcggacgcat 3960gagtctgtac
ggttatccgc gtgaaaccac gccggagctg gatgcgctgc ataaaaccga
4020tccgaatctg accgtgttta ataacgtagt tacgtctcgt ccgtacacca
ttgaaatcct 4080gcaacaggcg ctgacctttg ccaatgaaaa gaacccggat
ctgtatctga cgcagccgtc 4140gctgatgaac atgatgaaac aggcgggtta
taaaaccttc 4180874960DNAArtificial SequenceDescription of
Artificial Sequence Synthetic polynucleotide 87aataggcgta
tcacgaggcc ctttcgtctt cacctcgaga attgtgagcg gataacaatt 60gacattgtga
gcggataaca agatactgag cacatcagca ggacgcactg accgaattca
120attaagctag caagaggaga agtcgagatg aacttacatg aatatcaggc
aaaacaactt 180tttgcccgct atggcttacc agcaccggtg ggttatgcct
gtactactcc gcgcgaagca 240gaagaagccg cttcaaaaat cggtgccggt
ccgtgggtag tgaaatgtca ggttcacgct 300ggtggccgcg gtaaagcggg
cggtgtgaaa gttgtaaaca gcaaagaaga catccgtgct 360tttgcagaaa
actggctggg caagcgtctg gtaacgtatc aaacagatgc caatggccaa
420ccggttaacc agattctggt tgaagcagcg accgatatcg ctaaagagct
gtatctcggt 480gccgttgttg accgtagttc ccgtcgtgtg gtctttatgg
cctccaccga aggcggcgtg 540gaaatcgaaa aagtggcgga agaaactccg
cacctgatcc ataaagttgc gcttgatccg 600ctgactggcc cgatgccgta
tcagggacgc gagctggcgt tcaaactggg tctggaaggt 660aaactggttc
agcagttcac caaaatcttc atgggcctgg cgaccatttt cctggagcgc
720gacctggcgt tgatcgaaat caacccgctg gtcatcacca aacagggcga
tctgatttgc 780ctcgacggca aactgggcgc tgacggcaac gcactgttcc
gccagcctga tctgcgcgaa 840atgcgtgacc agtcgcagga agatccgcgt
gaagcacagg ctgcacagtg ggaactgaac 900tacgttgcgc tggacggtaa
catcggttgt atggttaacg gcgcaggtct ggcgatgggt 960acgatggaca
tcgttaaact gcacggcggc gaaccggcta acttccttga cgttggcggc
1020ggcgcaacca aagaacgtgt aaccgaagcg ttcaaaatca tcctctctga
cgacaaagtg 1080aaagccgttc tggttaacat cttcggcggt atcgttcgtt
gcgacctgat cgctgacggt 1140atcatcggcg cggtagcaga agtgggtgtt
aacgtaccgg tcgtggtacg tctggaaggt 1200aacaacgccg aactcggcgc
gaagaaactg gctgacagcg gcctgaatat tattgcagca 1260aaaggtctga
cggatgcagc tcagcaggtt gttgccgcag tggaggggaa ataatgtcca
1320ttttaatcga taaaaacacc aaggttatct gccagggctt taccggtagc
caggggactt 1380tccactcaga acaggccatt gcatacggca ctaaaatggt
tggcggcgta accccaggta 1440aaggcggcac cacccacctc ggcctgccgg
tgttcaacac cgtgcgtgaa gccgttgctg 1500ccactggcgc taccgcttct
gttatctacg taccagcacc gttctgcaaa gactccattc 1560tggaagccat
cgacgcaggc atcaaactga ttatcaccat cactgaaggc atcccgacgc
1620tggatatgct gaccgtgaaa gtgaagctgg atgaagcagg cgttcgtatg
atcggcccga 1680actgcccagg cgttatcact ccgggtgaat gcaaaatcgg
tatccagcct ggtcacattc 1740acaaaccggg taaagtgggt atcgtttccc
gttccggtac actgacctat gaagcggtta 1800aacagaccac ggattacggt
ttcggtcagt cgacctgtgt cggtatcggc ggtgacccga 1860tcccgggctc
taactttatc gacattctcg aaatgttcga aaaagatccg cagaccgaag
1920cgatcgtgat gatcggtgag atcggcggta gcgctgaaga agaagcagct
gcgtacatca 1980aagagcacgt taccaagcca gttgtgggtt acatcgctgg
tgtgactgcg ccgaaaggca 2040aacgtatggg ccacgcgggt gccatcattg
ccggtgggaa agggactgcg gatgagaaat 2100tcgctgctct ggaagccgca
ggcgtgaaaa ccgttcgcag cctggcggat atcggtgaag 2160cactgaaaac
tgttctgaaa taatctagca agaggagaag tcgacatgga aatcaaagaa
2220atggtgagcc ttgcacgcaa ggctcagaag gagtatcaag ctacccataa
ccaagaagca 2280gttgacaaca tttgccgagc tgcagcaaaa gttatttatg
aaaatgcagc tattctggct 2340cgcgaagcag tagacgaaac cggcatgggc
gtttacgaac acaaagtggc caagaatcaa 2400ggcaaatcca aaggtgtttg
gtacaacctc cacaataaaa aatcgattgg tatcctcaat 2460atagacgagc
gtaccggtat gatcgagatt gcaaagccta tcggagttgt aggagccgta
2520acgccgacga ccaacccgat cgttactccg atgagcaata tcatctttgc
tcttaagacc 2580tgcaatgcca tcattattgc cccccacccc agatccaaaa
aatgctctgc acacgcagtt 2640cgtctgatca aagaagctat cgctccgttc
aacgtaccgg aaggtatggt tcagatcatc 2700gaagaaccca gcatcgagaa
gacgcaggaa ctcatgggcg ccgtagacgt agtagttgct 2760acgggtggta
tgggcatggt gaagtctgca tattcttcag gaaagccttc tttcggtgtt
2820ggagccggta acgttcaggt gatcgtggat agcaacatcg atttcgaagc
tgctgcagaa 2880aaaatcatca ccggtcgtgc tttcgacaac ggtatcatct
gctcaggcga acagagcatc 2940atctacaacg aggctgacaa ggaagcagtt
ttcacagcat tccgcaacca cggtgcatat 3000ttctgtgacg aagccgaagg
agatcgggct cgtgcagcta tcttcgaaaa tggagccatc 3060gcgaaagatg
tagtaggtca gagcgttgcc ttcattgcca agaaagcaaa catcaatatc
3120cccgagggta cccgtattct cgttgttgaa gctcgcggcg taggagcaga
agacgttatc 3180tgtaaggaaa agatgtgtcc cgtaatgtgc gccctcagct
acaagcactt cgaagaaggt 3240gtagaaatcg cacgtacgaa cctcgccaac
gaaggtaacg gccacacctg tgctatccac 3300tccaacaatc aggcacacat
catcctcgca ggatcagagc tgacggtatc tcgtatcgta 3360gtgaatgctc
cgagtgccac tacagcaggc ggtcacatcc aaaacggtct tgccgtaacc
3420aatacgctcg gatgcggatc atggggtaat aactctatct ccgagaactt
cacttacaag 3480cacctcctca acatttcacg catcgcaccg ttgaattcaa
gcattcacat ccccgatgac 3540aaagaaatct gggaactcta atctagcaag
aggagaagtc gacatgcaac ttttcaaact 3600caagagtgta acacatcact
ttgacacttt tgcagaattt gccaaggaat tctgtcttgg 3660agaacgcgac
ttggtaatta ccaacgagtt catctatgaa ccgtatatga aggcatgcca
3720gctcccctgc cattttgtta tgcaggagaa atatgggcaa ggcgagcctt
ctgacgaaat 3780gatgaataac atcttggcag acatccgtaa tatccagttc
gaccgcgtaa tcggtatcgg 3840aggaggtacg gttattgaca tctctaaact
tttcgttctg aaaggattaa atgatgtact 3900cgatgcattc gaccgcaaaa
tacctcttat caaagagaaa gaactgatca ttgtgcccac 3960aacatgcgga
acgggtagcg aggtgacgaa catttctatc gcagaaatca aaagccgtca
4020caccaaaatg ggattggctg acgatgccat tgttgcagac catgccatca
tcatacctga 4080acttctgaag agcttgcctt tccacttcta cgcatgcagt
gcaatcgatg ctcttatcca 4140tgccatcgag tcatacgtat ctcctaaagc
cagtccatat tctcgtctgt tcagtgaggc 4200ggcttgggac attatcctgg
aagtattcaa gaaaatcgcc gaacacggcc ctgaataccg 4260cttcgaaaag
ctgggagaaa tgatcatggc cagcaactat gccggtatag ccttcggaaa
4320tgcaggagta ggagccgtcc acgcactatc ctacccgttg ggaggcaact
atcacgtgcc 4380gcatggagaa gcaaactatc agttcttcac agaggtattc
aaagtatacc aaaagaagaa 4440tcctttcggc tatatagtcg aactcaactg
gaagctctcc aagatactga actgccagcc 4500cgaatacgta tatccgaagc
tggatgaact tctcggatgc cttcttacca agaaaccttt 4560gcacgaatac
ggcatgaagg acgaagaggt aagaggcttt gcggaatcag tgcttaagac
4620acagcaaaga ttgctcgcca acaactacgt agagcttact gtagatgaga
tcgaaggtat 4680ctacagaaga ctctactaat ctagaaagct tcctagaggc
atcaaataaa acgaaaggct 4740cagtcgaaag actgggcctt tcgttttatc
tgttgtttgt cggtgaacgc tctcctgagt 4800aggacaaatc cgccgcccta
gacctaggcg ttcggctgcg acacgtcttg agcgattgtg 4860taggctggag
ctgcttcgaa gttcctatac tttctagaga ataggaactt cggaatagga
4920actaaggagg atattcatat ggaccatggc taattcccat
4960885083DNAArtificial SequenceDescription of Artificial Sequence
Synthetic polynucleotide 88tcgagaaatt tatcaaaaag agtgttgact
tgtgagcgga taacaatgat acttagattc 60aattgtgagc ggataacaat ttcacacaga
attcaattaa gctagcaaga ggagaagtcg 120acatggccaa cataagttca
ccattcgggc aaaacgaatg gctggttgaa gagatgtacc 180gcaagttccg
cgacgacccc tcctcggtcg atcccagctg gcacgagttc ctggttgact
240acagccccga acccacctcc caaccagctg ccgaaccaac ccgggttacc
tcgccactcg 300ttgccgagcg ggccgctgcg gccgccccgc aggcaccccc
caagccggcc gacaccgcgg 360ccgcgggcaa cggcgtggtc gccgcactgg
ccgccaaaac tgccgttccc ccgccagccg 420aaggtgacga ggtagcggtg
ctgcgcggcg ccgccgcggc cgtcgtcaag aacatgtccg 480cgtcgttgga
ggtgccgacg gcgaccagcg tccgggcggt cccggccaag ctactgatcg
540acaaccggat cgtcatcaac aaccagttga agcggacccg cggcggcaag
atctcgttca 600cgcatttgct gggctacgcc ctggtgcagg cggtgaagaa
attcccgaac atgaaccggc 660actacaccga agtcgacggc aagcccaccg
cggtcacgcc ggcgcacacc aatctcggcc 720tggcgatcga cctgcaaggc
aaggacggga agcgttccct ggtggtggcc ggcatcaagc 780ggtgcgagac
catgcgattc gcgcagttcg tcacggccta cgaagacatc gtacgccggg
840cccgcgacgg caagctgacc actgaagact ttgccggcgt gacgatttcg
ctgaccaatc 900ccggaaccat cggcaccgtg cattcggtgc cgcggctgat
gcccggccag ggcgccatca 960tcggcgtggg cgccatggaa taccccgccg
agtttcaagg cgccagcgag gaacgcatcg 1020ccgagctggg catcggcaaa
ttgatcactt tgacctccac ctacgaccac cgcatcatcc 1080agggcgcgga
atcgggcgac ttcctgcgca ccatccacga gttgctgctc tcggatggct
1140tctgggacga ggtcttccgc gaactgagca tcccatatct gccggtgcgc
tggagcaccg 1200acaaccccga ctcgatcgtc gacaagaacg ctcgcgtcat
gaacttgatc gcggcctacc 1260gcaaccgcgg ccatctgatg gccgataccg
acccgctgcg gttggacaaa gctcggttcc 1320gcagtcaccc cgacctcgaa
gtgctgaccc acggcctgac gctgtgggat ctcgatcggg 1380tgttcaaggt
cgacggcttt gccggtgcgc agtacaagaa actgcgcgac gtgctgggct
1440tgctgcgcga tgcctactgc cgccacatcg gcgtggagta cgcccatatc
ctcgaccccg 1500aacaaaagga gtggctcgaa caacgggtcg agaccaagca
cgtcaaaccc actgtggccc 1560aacagaaata catcctcagc aagctcaacg
ccgccgaggc ctttgaaacg ttcctacaga 1620ccaagtacgt cggccagaag
cggttctcgc tggaaggcgc cgaaagcgtg atcccgatga 1680tggacgcggc
gatcgaccag tgcgctgagc acggcctcga cgaggtggtc atcgggatgc
1740cgcaccgggg ccggctcaac gtgctggcca acatcgtcgg caagccgtac
tcgcagatct 1800tcaccgagtt cgagggcaac ctgaatccgt cgcaggcgca
cggctccggt gacgtcaagt 1860accacctggg cgccaccggg ctgtacctgc
agatgttcgg cgacaacgac attcaggtgt 1920cgctgaccgc caacccgtcg
catctggagg ccgtcgaccc ggtgctggag ggattggtgc 1980gggccaagca
ggatctgctc gaccacggaa gcatcgacag cgacggccaa cgggcgttct
2040cggtggtgcc gctgatgttg catggcgatg ccgcgttcgc cggtcagggt
gtggtcgccg 2100agacgctgaa cctggcgaat ctgccgggct accgcgtcgg
cggcaccatc cacatcatcg 2160tcaacaacca gatcggcttc accaccgcgc
ccgagtattc caggtccagc gagtactgca 2220ccgacgtcgc aaagatgatc
ggggcaccga tctttcacgt caacggcgac gacccggagg 2280cgtgtgtctg
ggtggcgcgg ttggcggtgg acttccgaca acggttcaag aaggacgtcg
2340tcatcgacat gctgtgctac cgccgccgcg ggcacaacga gggtgacgac
ccgtcgatga 2400ccaaccccta catgtacgac gtcgtcgaca ccaagcgcgg
ggcccgcaaa agctacaccg 2460aagccctgat cggacgtggc gacatctcga
tgaaggaggc cgaggacgcg ctgcgcgact 2520accagggcca gctggaacgg
gtgttcaacg aagtgcgcga gctggagaag cacggtgtgc 2580agccgagcga
gtcggtcgag tccgaccaga tgattcccgc ggggctggcc actgcggtgg
2640acaagtcgct gctggcccgg atcggcgatg cgttcctcgc cttgccgaac
ggcttcaccg 2700cgcacccgcg agtccaaccg gtgctggaga agcgccggga
gatggcctat gaaggcaaga 2760tcgactgggc ctttggcgag ctgctggcgc
tgggctcgct ggtggccgaa ggcaagctgg 2820tgcgcttgtc ggggcaggac
agccgccgcg gcaccttctc ccagcggcat tcggttctca 2880tcgaccgcca
cactggcgag gagttcacac cactgcagct gctggcgacc aactccgacg
2940gcagcccgac cggcggaaag ttcctggtct acgactcgcc actgtcggag
tacgccgccg 3000tcggcttcga gtacggctac actgtgggca atccggacgc
cgtggtgctc tgggaggcgc 3060agttcggcga cttcgtcaac ggcgcacagt
cgatcatcga cgagttcatc agctccggtg 3120aggccaagtg gggccaattg
tccaacgtcg tgctgctgtt accgcacggg cacgaggggc 3180agggacccga
ccacacttct gcccggatcg aacgcttctt gcagttgtgg gcggaaggtt
3240cgatgaccat cgcgatgccg tcgactccgt cgaactactt ccacctgcta
cgccggcatg 3300ccctggacgg catccaacgc ccgctgatcg tgttcacgcc
caagtcgatg ttgcgtcaca 3360aggccgccgt cagcgaaatc aaggacttca
ccgagatcaa gttccgctca gtgctggagg 3420aacccaccta tgaggacggc
atcggagacc
gcaacaaggt cagccggatc ctgctgacca 3480gtggcaagct gtattacgag
ctggccgccc gcaaggccaa ggacaaccgc aatgacctcg 3540cgatcgtgcg
gcttgaacag ctcgccccgc tgcccaggcg tcgactgcgt gaaacgctgg
3600accgctacga gaacgtcaag gagttcttct gggtccaaga ggaaccggcc
aaccagggtg 3660cgtggccgcg attcgggctc gaactacccg agctgctgcc
tgacaagttg gccgggatca 3720agcgaatctc gcgccgggcg atgtcagccc
cgtcgtcagg ctcgtcgaag gtgcacgccg 3780tcgaacagca ggagatcctc
gacgaggcgt tcggctaatc tagcaagagg agaagtcgac 3840atgaagttat
taaaattggc acctgatgtt tataaatttg atactgcaga ggagtttatg
3900aaatacttta aggttggaaa aggtgacttt atacttacta atgaattttt
atataaacct 3960ttccttgaga aattcaatga tggtgcagat gctgtatttc
aggagaaata tggactcggt 4020gaaccttctg atgaaatgat aaacaatata
attaaggata ttggagataa acaatataat 4080agaattattg ctgtaggggg
aggatctgta atagatatag ccaaaatcct cagtcttaag 4140tatactgatg
attcattgga tttgtttgag ggaaaagtac ctcttgtaaa aaacaaagaa
4200ttaattatag ttccaactac atgtggaaca ggttcagaag ttacaaatgt
atcagttgca 4260gaattaaaga gaagacatac taaaaaagga attgcttcag
acgaattata tgcaacttat 4320gcagtacttg taccagaatt tataaaagga
cttccatata agttttttgt aaccagctcc 4380gtagatgcct taatacatgc
aacagaagct tatgtatctc caaatgcaaa tccttatact 4440gatatgttta
gtgtaaaagc tatggagtta attttaaatg gatacatgca aatggtagag
4500aaaggaaatg attacagagt tgaaataatt gaggattttg ttataggcag
caattatgca 4560ggtatagctt ttggaaatgc aggagtggga gcggttcacg
cactctcata tccaataggc 4620ggaaattatc atgtgcctca tggagaagca
aattatctgt tttttacaga aatatttaaa 4680acttattatg agaaaaatcc
aaatggcaag attaaagatg taaataaact attagcaggc 4740atactaaaat
gtgatgaaag tgaagcttat gacagtttat cacaactttt agataaatta
4800ttgtcaagaa aaccattaag agaatatgga atgaaagagg aagaaattga
aacttttgct 4860gattcagtaa tagaaggaca gcagagactg ttggtaaaca
attatgaacc tttttcaaga 4920gaagacatag taaacacata taaaaagtta
tattaatcta gaaagcttcc tagaggcatc 4980aaataaaacg aaaggctcag
tcgaaagact gggcctttcg ttttatctgt tgtttgtcgg 5040tgaacgctct
cctgagtagg acaaatccgc cgccctagac cta 5083895097DNAArtificial
SequenceDescription of Artificial Sequence Synthetic polynucleotide
89tctgtatcag gctgaaaatc ttctctcatc cgccaaaaca gcttcggcgt taagatgcgc
60gctcaaggac gtaagccgtc gactctcgcc gtgctggcgc aggacacggc taccactcct
120ttctctgttg atattctgct tgccattgag caaaccgcca gcgagttcgg
ctggaatagt 180tttttaatca atattttttc tgaagatgac gctgcccgcg
cggcacgtca gctgcttgcc 240caccgtccgg atggcattat ctatactaca
atggggctgc gacatatcac gctgcctgag 300tctctgtatg gtgaaaatat
tgtattggcg aactgtgtgg cggatgaccc agcgttaccc 360agttatatcc
ctgatgatta cactgcacaa tatgaatcaa cacagcattt gctcgcggcg
420ggctatcgtc aaccgttatg cttctggcta ccggaaagtg cgttggcaac
agggtatcgt 480cggcagggat ttgagcaggc ctggcgtgat gctggacgag
atctggctga ggtgaaacaa 540tttcacatgg caacaggtga tgatcactac
accgatctcg caagtttact caatgcccac 600ttcaaaccgg gcaaaccaga
ttttgatgtt ctgatatgtg gtaacgatcg cgcagccttt 660gtggcttatc
aggttcttct ggcgaagggg gtacgaatcc cgcaggatgt cgccgtaatg
720ggctttgata atctggttgg cgtcgggcat ctgtttttac cgccgctgac
cacaattcag 780cttccacatg acattatcgg gcgggaagct gcattgcata
ttattgaagg tcgtgaaggg 840ggaagagtga cgcggatccc ttgcccgctg
ttgatccgtt gttccacctg atattatgtt 900aacccagtag ccagagtgct
ccatgttgca gcacagccac tccgtgggag gcataaagcg 960acagttcccg
ttcttctggc tgcggataga ttcgactact catcaccgct tccccgtcgt
1020taataaatac ttccacggat gatgtatcga taaatatcct tagggcgagc
gtgtcacgct 1080gcgggagggg aatactacgg tagccgtcta aattctcgtg
tgggtaatac cgccacaaaa 1140caagtcgctc agattggtta tcaatataca
gccgcattcc agtgccgagc tgtaatccgt 1200aatgttcggc atcactgttc
ttcagcgccc actgcaactg aatctcaact gcttgcgcgt 1260tttcctgcaa
aacatattta ttgctgattg tgcggggaga gacagattga tgctgctggc
1320gtaacgactc agcttcgtgt accgggcgtt gtagaagttt gccattgctc
tctgatagct 1380cgcgcgccag cgtcatgcag cctgcccatc cttcacgttt
tgagggcatt ggcgattccc 1440acatatccat ccagccgata acaatacgcc
gaccatcctt cgctaaaaag ctttgtggtg 1500cataaaagtc atgcccgtta
tcaagttcag taaaatgccc ggattgtgca aaaagtcgtc 1560ctggcgacca
cattccgggt attacgccac tttgaaagcg atttcggtaa ctgtatccct
1620cggcattcat tccctgcggg gaaaacatca gataatgctg atcgccaagg
ctgaaaaagt 1680ccggacattc ccacatatag ctttcacccg catcagcgtg
ggccagtacg cgatcgaagg 1740tccattcacg caacgaactg ccgcgataaa
gcaggatctg ccccgtgttg cctggatctt 1800tcgccccgac taccatccac
catgtgtcgg cttcacgcca cactttagga tcgcggaagt 1860gcatgattcc
ttctggtgga gtgaggatca caccctgttt ctcgaaatga ataccatccc
1920gactggtagc cagacattgt acttcgcgaa ttgcatcgtc attacctgca
ccatcgagcc 1980agacgtgtcc ggtgtagata agtgagagga caccattgtc
atcgacagca ctacctgaaa 2040aacacccgtc tttgtcatta tcgtctcctg
gcgctagcgc aataggctca tgctgccagt 2100ggatcatatc gtcgctggtg
gcatgtcccc agtgcattgg cccccagtgt tcgctcatcg 2160gatgatgttg
ataaaacgcg tgataacgat cgttaaacca gatcaggccg tttggatcgt
2220tcatccaccc ggcaggaggc gcgaggtgaa aatggggata gaaagtgtta
ccccggtgct 2280catgaagttt tgctagggcg ttttgcgccg catgcaatcg
agattgcgtc attttaatca 2340tcctggttaa gcaaatttgg tgaattgtta
acgttaactt ttataaaaat aaagtccctt 2400actttcataa atgcgatgaa
tatcacaaat gttaacgtta actatgacgt tttgtgatcg 2460aatatgcatg
ttttagtaaa tccatgacga ttttgcgaaa aagaggttta tcactatgcg
2520taactcagat gaatttaagg gaaaaaaatg tcagccaaag tatgggtttt
aggggatgcg 2580gtcgtagatc tcttgccaga atcagacggg cgcctactgc
cttgtcctgg cggcgcgcca 2640gctaacgttg cggtgggaat cgccagatta
ggcggaacaa gtgggtttat aggtcgggtg 2700ggggatgatc cttttggtgc
gttaatgcaa agaacgctgc taactgaggg agtcgatatc 2760acgtatctga
agcaagatga atggcaccgg acatccacgg tgcttgtcga tctgaacgat
2820caaggggaac gttcatttac gtttatggtc cgccccagtg ccgatctttt
tttagagacg 2880acagacttgc cctgctggcg acatggcgaa tggttacatc
tctgttcaat tgcgttgtct 2940gccgagcctt cgcgtaccag cgcatttact
gcgatgacgg cgatccggca tgccggaggt 3000tttgtcagct tcgatcctaa
tattcgtgaa gatctatggc aagacgagca tttgctccgc 3060ttgtgtttgc
ggcaggcgct acaactggcg gatgtcgtca agctctcgga agaagaatgg
3120cgacttatca gtggaaaaac acagaacgat caggatatat gcgccctggc
aaaagagtat 3180gagatcgcca tgctgttggt gactaaaggt gcagaagggg
tggtggtctg ttatcgagga 3240caagttcacc attttgctgg aatgtctgtg
aattgtgtcg atagcacggg ggcgggagat 3300gcgttcgttg ccgggttact
cacaggtctg tcctctacgg gattatctac agatgagaga 3360gaaatgcgac
gaattatcga tctcgctcaa cgttgcggag cgcttgcagt aacggcgaaa
3420ggggcaatga cagcgctgcc atgtcgacaa gaactggaat agtgagaagt
aaacggcgaa 3480gtcgctctta tctctaaata ggacgtgaat tttttaacga
caggcaggta attatggcac 3540tgaatattcc attcagaaat gcgtactatc
gttttgcatc cagttactca tttctctttt 3600ttatttcctg gtcgctgtgg
tggtcgttat acgctatttg gctgaaagga catctagggt 3660tgacagggac
ggaattaggt acactttatt cggtcaacca gtttaccagc attctattta
3720tgatgttcta cggcatcgtt caggataaac tcggtctgaa gaaaccgctc
atctggtgta 3780tgagtttcat cctggtcttg accggaccgt ttatgattta
cgtttatgaa ccgttactgc 3840aaagcaattt ttctgtaggt ctaattctgg
gggcgctatt ttttggcttg gggtatctgg 3900cgggatgcgg tttgcttgat
agcttcaccg aaaaaatggc gcgaaatttt catttcgaat 3960atggaacagc
gcgcgcctgg ggatcttttg gctatgctat tggcgcgttc tttgccggca
4020tattttttag tatcagtccc catatcaact tctggttggt ctcgctattt
ggcgctgtat 4080ttatgatgat caacatgcgt tttaaagata aggatcacca
gtgcgtagcg gcagatgcgg 4140gaggggtaaa aaaagaggat tttatcgcag
ttttcaagga tcgaaacttc tgggttttcg 4200tcatatttat tgtggggacg
tggtctttct ataacatttt tgatcaacaa ctttttcctg 4260tcttttattc
aggtttattc gaatcacacg atgtaggaac gcgcctgtat ggttatctca
4320actcattcca ggtggtactc gaagcgctgt gcatggcgat tattcctttc
tttgtgaatc 4380gggtagggcc aaaaaatgca ttacttatcg gagttgtgat
tatggcgttg cgtatccttt 4440cctgcgcgct gttcgttaac ccctggatta
tttcattagt gaagttgtta catgccattg 4500aggttccact ttgtgtcata
tccgtcttca aatacagcgt ggcaaacttt gataagcgcc 4560tgtcgtcgac
gatctttctg attggttttc aaattgccag ttcgcttggg attgtgctgc
4620tttcaacgcc gactgggata ctctttgacc acgcaggcta ccagacagtt
ttcttcgcaa 4680tttcgggtat tgtctgcctg atgttgctat ttggcatttt
cttcttgagt aaaaaacgcg 4740agcaaatagt tatggaaacg cctgtacctt
cagcaatata gacgtaaact ttttccggtt 4800gttgtcgata gctctatatc
cctcaaccgg aaaataataa tagtaaaatg cttagccctg 4860ctaataatcg
cctaatccaa acgcctcatt catgttctgg tacagtcgct caaatgtact
4920tcagatgcgc ggttcgctga tttccaggac attgtcgtca ttcagtgacc
tgtcccgtgt 4980atcacggtcc tgcgaattca tcaaggaatg cattgcggag
tgaagtatcg agtcacgcca 5040tatttcgtca cccgaagatg agttttgaga
tattaaggca ggtgactttc actcaca 5097903525DNANocardia iowensis
90atggcagtgg attcaccgga tgagcggcta cagcgccgca ttgcacagtt gtttgcagaa
60gatgagcagg tcaaggccgc acgtccgctc gaagcggtga gcgcggcggt gagcgcgccc
120ggtatgcggc tggcgcagat cgccgccact gttatggcgg gttacgccga
ccgcccggcc 180gccgggcagc gtgcgttcga actgaacacc gacgacgcga
cgggccgcac ctcgctgcgg 240ttacttcccc gattcgagac catcacctat
cgcgaactgt ggcagcgagt cggcgaggtt 300gccgcggcct ggcatcatga
tcccgagaac cccttgcgcg caggtgattt cgtcgccctg 360ctcggcttca
ccagcatcga ctacgccacc ctcgacctgg ccgatatcca cctcggcgcg
420gttaccgtgc cgttgcaggc cagcgcggcg gtgtcccagc tgatcgctat
cctcaccgag 480acttcgccgc ggctgctcgc ctcgaccccg gagcacctcg
atgcggcggt cgagtgccta 540ctcgcgggca ccacaccgga acgactggtg
gtcttcgact accaccccga ggacgacgac 600cagcgtgcgg ccttcgaatc
cgcccgccgc cgccttgccg acgcgggcag cttggtgatc 660gtcgaaacgc
tcgatgccgt gcgtgcccgg ggccgcgact taccggccgc gccactgttc
720gttcccgaca ccgacgacga cccgctggcc ctgctgatct acacctccgg
cagcaccgga 780acgccgaagg gcgcgatgta caccaatcgg ttggccgcca
cgatgtggca ggggaactcg 840atgctgcagg ggaactcgca acgggtcggg
atcaatctca actacatgcc gatgagccac 900atcgccggtc gcatatcgct
gttcggcgtg ctcgctcgcg gtggcaccgc atacttcgcg 960gccaagagcg
acatgtcgac actgttcgaa gacatcggct tggtacgtcc caccgagatc
1020ttcttcgtcc cgcgcgtgtg cgacatggtc ttccagcgct atcagagcga
gctggaccgg 1080cgctcggtgg cgggcgccga cctggacacg ctcgatcggg
aagtgaaagc cgacctccgg 1140cagaactacc tcggtgggcg cttcctggtg
gcggtcgtcg gcagcgcgcc gctggccgcg 1200gagatgaaga cgttcatgga
gtccgtcctc gatctgccac tgcacgacgg gtacgggtcg 1260accgaggcgg
gcgcaagcgt gctgctcgac aaccagatcc agcggccgcc ggtgctcgat
1320tacaagctcg tcgacgtgcc cgaactgggt tacttccgca ccgaccggcc
gcatccgcgc 1380ggtgagctgt tgttgaaggc ggagaccacg attccgggct
actacaagcg gcccgaggtc 1440accgcggaga tcttcgacga ggacggcttc
tacaagaccg gcgatatcgt ggccgagctc 1500gagcacgatc ggctggtcta
tgtcgaccgt cgcaacaatg tgctcaaact gtcgcagggc 1560gagttcgtga
ccgtcgccca tctcgaggcc gtgttcgcca gcagcccgct gatccggcag
1620atcttcatct acggcagcag cgaacgttcc tatctgctcg cggtgatcgt
ccccaccgac 1680gacgcgctgc gcggccgcga caccgccacc ttgaaatcgg
cactggccga atcgattcag 1740cgcatcgcca aggacgcgaa cctgcagccc
tacgagattc cgcgcgattt cctgatcgag 1800accgagccgt tcaccatcgc
caacggactg ctctccggca tcgcgaagct gctgcgcccc 1860aatctgaagg
aacgctacgg cgctcagctg gagcagatgt acaccgatct cgcgacaggc
1920caggccgatg agctgctcgc cctgcgccgc gaagccgccg acctgccggt
gctcgaaacc 1980gtcagccggg cagcgaaagc gatgctcggc gtcgcctccg
ccgatatgcg tcccgacgcg 2040cacttcaccg acctgggcgg cgattccctt
tccgcgctgt cgttctcgaa cctgctgcac 2100gagatcttcg gggtcgaggt
gccggtgggt gtcgtcgtca gcccggcgaa cgagctgcgc 2160gatctggcga
attacattga ggcggaacgc aactcgggcg cgaagcgtcc caccttcacc
2220tcggtgcacg gcggcggttc cgagatccgc gccgccgatc tgaccctcga
caagttcatc 2280gatgcccgca ccctggccgc cgccgacagc attccgcacg
cgccggtgcc agcgcagacg 2340gtgctgctga ccggcgcgaa cggctacctc
ggccggttcc tgtgcctgga atggctggag 2400cggctggaca agacgggtgg
cacgctgatc tgcgtcgtgc gcggtagtga cgcggccgcg 2460gcccgtaaac
ggctggactc ggcgttcgac agcggcgatc ccggcctgct cgagcactac
2520cagcaactgg ccgcacggac cctggaagtc ctcgccggtg atatcggcga
cccgaatctc 2580ggtctggacg acgcgacttg gcagcggttg gccgaaaccg
tcgacctgat cgtccatccc 2640gccgcgttgg tcaaccacgt ccttccctac
acccagctgt tcggccccaa tgtcgtcggc 2700accgccgaaa tcgtccggtt
ggcgatcacg gcgcggcgca agccggtcac ctacctgtcg 2760accgtcggag
tggccgacca ggtcgacccg gcggagtatc aggaggacag cgacgtccgc
2820gagatgagcg cggtgcgcgt cgtgcgcgag agttacgcca acggctacgg
caacagcaag 2880tgggcggggg aggtcctgct gcgcgaagca cacgatctgt
gtggcttgcc ggtcgcggtg 2940ttccgttcgg acatgatcct ggcgcacagc
cggtacgcgg gtcagctcaa cgtccaggac 3000gtgttcaccc ggctgatcct
cagcctggtc gccaccggca tcgcgccgta ctcgttctac 3060cgaaccgacg
cggacggcaa ccggcagcgg gcccactatg acggcttgcc ggcggacttc
3120acggcggcgg cgatcaccgc gctcggcatc caagccaccg aaggcttccg
gacctacgac 3180gtgctcaatc cgtacgacga tggcatctcc ctcgatgaat
tcgtcgactg gctcgtcgaa 3240tccggccacc cgatccagcg catcaccgac
tacagcgact ggttccaccg tttcgagacg 3300gcgatccgcg cgctgccgga
aaagcaacgc caggcctcgg tgctgccgtt gctggacgcc 3360taccgcaacc
cctgcccggc ggtccgcggc gcgatactcc cggccaagga gttccaagcg
3420gcggtgcaaa cagccaaaat cggtccggaa caggacatcc cgcatttgtc
cgcgccactg 3480atcgataagt acgtcagcga tctggaactg cttcagctgc tctaa
3525911174PRTNocardia iowensis 91Met Ala Val Asp Ser Pro Asp Glu
Arg Leu Gln Arg Arg Ile Ala Gln 1 5 10 15 Leu Phe Ala Glu Asp Glu
Gln Val Lys Ala Ala Arg Pro Leu Glu Ala 20 25 30 Val Ser Ala Ala
Val Ser Ala Pro Gly Met Arg Leu Ala Gln Ile Ala 35 40 45 Ala Thr
Val Met Ala Gly Tyr Ala Asp Arg Pro Ala Ala Gly Gln Arg 50 55 60
Ala Phe Glu Leu Asn Thr Asp Asp Ala Thr Gly Arg Thr Ser Leu Arg 65
70 75 80 Leu Leu Pro Arg Phe Glu Thr Ile Thr Tyr Arg Glu Leu Trp
Gln Arg 85 90 95 Val Gly Glu Val Ala Ala Ala Trp His His Asp Pro
Glu Asn Pro Leu 100 105 110 Arg Ala Gly Asp Phe Val Ala Leu Leu Gly
Phe Thr Ser Ile Asp Tyr 115 120 125 Ala Thr Leu Asp Leu Ala Asp Ile
His Leu Gly Ala Val Thr Val Pro 130 135 140 Leu Gln Ala Ser Ala Ala
Val Ser Gln Leu Ile Ala Ile Leu Thr Glu 145 150 155 160 Thr Ser Pro
Arg Leu Leu Ala Ser Thr Pro Glu His Leu Asp Ala Ala 165 170 175 Val
Glu Cys Leu Leu Ala Gly Thr Thr Pro Glu Arg Leu Val Val Phe 180 185
190 Asp Tyr His Pro Glu Asp Asp Asp Gln Arg Ala Ala Phe Glu Ser Ala
195 200 205 Arg Arg Arg Leu Ala Asp Ala Gly Ser Leu Val Ile Val Glu
Thr Leu 210 215 220 Asp Ala Val Arg Ala Arg Gly Arg Asp Leu Pro Ala
Ala Pro Leu Phe 225 230 235 240 Val Pro Asp Thr Asp Asp Asp Pro Leu
Ala Leu Leu Ile Tyr Thr Ser 245 250 255 Gly Ser Thr Gly Thr Pro Lys
Gly Ala Met Tyr Thr Asn Arg Leu Ala 260 265 270 Ala Thr Met Trp Gln
Gly Asn Ser Met Leu Gln Gly Asn Ser Gln Arg 275 280 285 Val Gly Ile
Asn Leu Asn Tyr Met Pro Met Ser His Ile Ala Gly Arg 290 295 300 Ile
Ser Leu Phe Gly Val Leu Ala Arg Gly Gly Thr Ala Tyr Phe Ala 305 310
315 320 Ala Lys Ser Asp Met Ser Thr Leu Phe Glu Asp Ile Gly Leu Val
Arg 325 330 335 Pro Thr Glu Ile Phe Phe Val Pro Arg Val Cys Asp Met
Val Phe Gln 340 345 350 Arg Tyr Gln Ser Glu Leu Asp Arg Arg Ser Val
Ala Gly Ala Asp Leu 355 360 365 Asp Thr Leu Asp Arg Glu Val Lys Ala
Asp Leu Arg Gln Asn Tyr Leu 370 375 380 Gly Gly Arg Phe Leu Val Ala
Val Val Gly Ser Ala Pro Leu Ala Ala 385 390 395 400 Glu Met Lys Thr
Phe Met Glu Ser Val Leu Asp Leu Pro Leu His Asp 405 410 415 Gly Tyr
Gly Ser Thr Glu Ala Gly Ala Ser Val Leu Leu Asp Asn Gln 420 425 430
Ile Gln Arg Pro Pro Val Leu Asp Tyr Lys Leu Val Asp Val Pro Glu 435
440 445 Leu Gly Tyr Phe Arg Thr Asp Arg Pro His Pro Arg Gly Glu Leu
Leu 450 455 460 Leu Lys Ala Glu Thr Thr Ile Pro Gly Tyr Tyr Lys Arg
Pro Glu Val 465 470 475 480 Thr Ala Glu Ile Phe Asp Glu Asp Gly Phe
Tyr Lys Thr Gly Asp Ile 485 490 495 Val Ala Glu Leu Glu His Asp Arg
Leu Val Tyr Val Asp Arg Arg Asn 500 505 510 Asn Val Leu Lys Leu Ser
Gln Gly Glu Phe Val Thr Val Ala His Leu 515 520 525 Glu Ala Val Phe
Ala Ser Ser Pro Leu Ile Arg Gln Ile Phe Ile Tyr 530 535 540 Gly Ser
Ser Glu Arg Ser Tyr Leu Leu Ala Val Ile Val Pro Thr Asp 545 550 555
560 Asp Ala Leu Arg Gly Arg Asp Thr Ala Thr Leu Lys Ser Ala Leu Ala
565 570 575 Glu Ser Ile Gln Arg Ile Ala Lys Asp Ala Asn Leu Gln Pro
Tyr Glu 580 585 590 Ile Pro Arg Asp Phe Leu Ile Glu Thr Glu Pro Phe
Thr Ile Ala Asn 595 600 605 Gly Leu Leu Ser Gly Ile Ala Lys Leu Leu
Arg Pro Asn Leu Lys Glu 610 615 620 Arg Tyr Gly Ala Gln Leu Glu Gln
Met Tyr Thr Asp Leu Ala Thr Gly 625 630 635 640 Gln Ala Asp Glu Leu
Leu Ala Leu Arg Arg Glu Ala Ala Asp Leu Pro 645 650 655 Val Leu Glu
Thr Val Ser Arg Ala Ala Lys Ala Met Leu Gly Val Ala 660 665 670 Ser
Ala Asp Met Arg Pro Asp Ala His Phe Thr Asp Leu Gly Gly Asp 675 680
685 Ser Leu Ser Ala Leu Ser Phe Ser Asn Leu Leu His Glu Ile Phe Gly
690 695 700 Val Glu Val Pro Val Gly Val Val
Val Ser Pro Ala Asn Glu Leu Arg 705 710 715 720 Asp Leu Ala Asn Tyr
Ile Glu Ala Glu Arg Asn Ser Gly Ala Lys Arg 725 730 735 Pro Thr Phe
Thr Ser Val His Gly Gly Gly Ser Glu Ile Arg Ala Ala 740 745 750 Asp
Leu Thr Leu Asp Lys Phe Ile Asp Ala Arg Thr Leu Ala Ala Ala 755 760
765 Asp Ser Ile Pro His Ala Pro Val Pro Ala Gln Thr Val Leu Leu Thr
770 775 780 Gly Ala Asn Gly Tyr Leu Gly Arg Phe Leu Cys Leu Glu Trp
Leu Glu 785 790 795 800 Arg Leu Asp Lys Thr Gly Gly Thr Leu Ile Cys
Val Val Arg Gly Ser 805 810 815 Asp Ala Ala Ala Ala Arg Lys Arg Leu
Asp Ser Ala Phe Asp Ser Gly 820 825 830 Asp Pro Gly Leu Leu Glu His
Tyr Gln Gln Leu Ala Ala Arg Thr Leu 835 840 845 Glu Val Leu Ala Gly
Asp Ile Gly Asp Pro Asn Leu Gly Leu Asp Asp 850 855 860 Ala Thr Trp
Gln Arg Leu Ala Glu Thr Val Asp Leu Ile Val His Pro 865 870 875 880
Ala Ala Leu Val Asn His Val Leu Pro Tyr Thr Gln Leu Phe Gly Pro 885
890 895 Asn Val Val Gly Thr Ala Glu Ile Val Arg Leu Ala Ile Thr Ala
Arg 900 905 910 Arg Lys Pro Val Thr Tyr Leu Ser Thr Val Gly Val Ala
Asp Gln Val 915 920 925 Asp Pro Ala Glu Tyr Gln Glu Asp Ser Asp Val
Arg Glu Met Ser Ala 930 935 940 Val Arg Val Val Arg Glu Ser Tyr Ala
Asn Gly Tyr Gly Asn Ser Lys 945 950 955 960 Trp Ala Gly Glu Val Leu
Leu Arg Glu Ala His Asp Leu Cys Gly Leu 965 970 975 Pro Val Ala Val
Phe Arg Ser Asp Met Ile Leu Ala His Ser Arg Tyr 980 985 990 Ala Gly
Gln Leu Asn Val Gln Asp Val Phe Thr Arg Leu Ile Leu Ser 995 1000
1005 Leu Val Ala Thr Gly Ile Ala Pro Tyr Ser Phe Tyr Arg Thr Asp
1010 1015 1020 Ala Asp Gly Asn Arg Gln Arg Ala His Tyr Asp Gly Leu
Pro Ala 1025 1030 1035 Asp Phe Thr Ala Ala Ala Ile Thr Ala Leu Gly
Ile Gln Ala Thr 1040 1045 1050 Glu Gly Phe Arg Thr Tyr Asp Val Leu
Asn Pro Tyr Asp Asp Gly 1055 1060 1065 Ile Ser Leu Asp Glu Phe Val
Asp Trp Leu Val Glu Ser Gly His 1070 1075 1080 Pro Ile Gln Arg Ile
Thr Asp Tyr Ser Asp Trp Phe His Arg Phe 1085 1090 1095 Glu Thr Ala
Ile Arg Ala Leu Pro Glu Lys Gln Arg Gln Ala Ser 1100 1105 1110 Val
Leu Pro Leu Leu Asp Ala Tyr Arg Asn Pro Cys Pro Ala Val 1115 1120
1125 Arg Gly Ala Ile Leu Pro Ala Lys Glu Phe Gln Ala Ala Val Gln
1130 1135 1140 Thr Ala Lys Ile Gly Pro Glu Gln Asp Ile Pro His Leu
Ser Ala 1145 1150 1155 Pro Leu Ile Asp Lys Tyr Val Ser Asp Leu Glu
Leu Leu Gln Leu 1160 1165 1170 Leu 92669DNAArtificial
SequenceDescription of Artificial Sequence Synthetic polynucleotide
92atgattgaaa ccattctgcc tgcaggcgtt gaaagcgcag aactgctgga atatccggaa
60gatctgaaag cacatccggc agaagaacat ctgattgcca aaagcgttga aaaacgtcgt
120cgtgatttta ttggtgcacg tcattgtgca cgtctggcac tggcagaact
gggtgaacct 180ccggttgcaa ttggtaaagg tgaacgtggt gcaccgattt
ggcctcgtgg tgttgttggt 240agcctgaccc attgtgatgg ttatcgtgca
gcagcagttg cacataaaat gcgctttcgc 300agcattggta ttgatgcaga
accgcatgca accctgccgg aaggtgttct ggatagcgtt 360agcctgccgc
cggaacgtga atggctgaaa accaccgata gcgcactgca tctggatcgt
420ctgctgtttt gtgcaaaaga agccacctat aaagcctggt ggccgctgac
agcacgttgg 480ctgggttttg aagaagccca tattaccttt gaaattgaag
atggtagcgc agatagcggt 540aatggcacct ttcatagcga actgctggtt
ccgggtcaga ccaatgatgg tggtacaccg 600ctgctgagct ttgatggtcg
ttggctgatt gcagatggtt ttattctgac cgcaattgcc 660tatgcctaa
66993222PRTArtificial SequenceDescription of Artificial Sequence
Synthetic polypeptide 93Met Ile Glu Thr Ile Leu Pro Ala Gly Val Glu
Ser Ala Glu Leu Leu 1 5 10 15 Glu Tyr Pro Glu Asp Leu Lys Ala His
Pro Ala Glu Glu His Leu Ile 20 25 30 Ala Lys Ser Val Glu Lys Arg
Arg Arg Asp Phe Ile Gly Ala Arg His 35 40 45 Cys Ala Arg Leu Ala
Leu Ala Glu Leu Gly Glu Pro Pro Val Ala Ile 50 55 60 Gly Lys Gly
Glu Arg Gly Ala Pro Ile Trp Pro Arg Gly Val Val Gly 65 70 75 80 Ser
Leu Thr His Cys Asp Gly Tyr Arg Ala Ala Ala Val Ala His Lys 85 90
95 Met Arg Phe Arg Ser Ile Gly Ile Asp Ala Glu Pro His Ala Thr Leu
100 105 110 Pro Glu Gly Val Leu Asp Ser Val Ser Leu Pro Pro Glu Arg
Glu Trp 115 120 125 Leu Lys Thr Thr Asp Ser Ala Leu His Leu Asp Arg
Leu Leu Phe Cys 130 135 140 Ala Lys Glu Ala Thr Tyr Lys Ala Trp Trp
Pro Leu Thr Ala Arg Trp 145 150 155 160 Leu Gly Phe Glu Glu Ala His
Ile Thr Phe Glu Ile Glu Asp Gly Ser 165 170 175 Ala Asp Ser Gly Asn
Gly Thr Phe His Ser Glu Leu Leu Val Pro Gly 180 185 190 Gln Thr Asn
Asp Gly Gly Thr Pro Leu Leu Ser Phe Asp Gly Arg Trp 195 200 205 Leu
Ile Ala Asp Gly Phe Ile Leu Thr Ala Ile Ala Tyr Ala 210 215 220
D00001
D00002
D00003
D00004
D00005
D00006
D00007
D00008
D00009
D00010
D00011
D00012
D00013
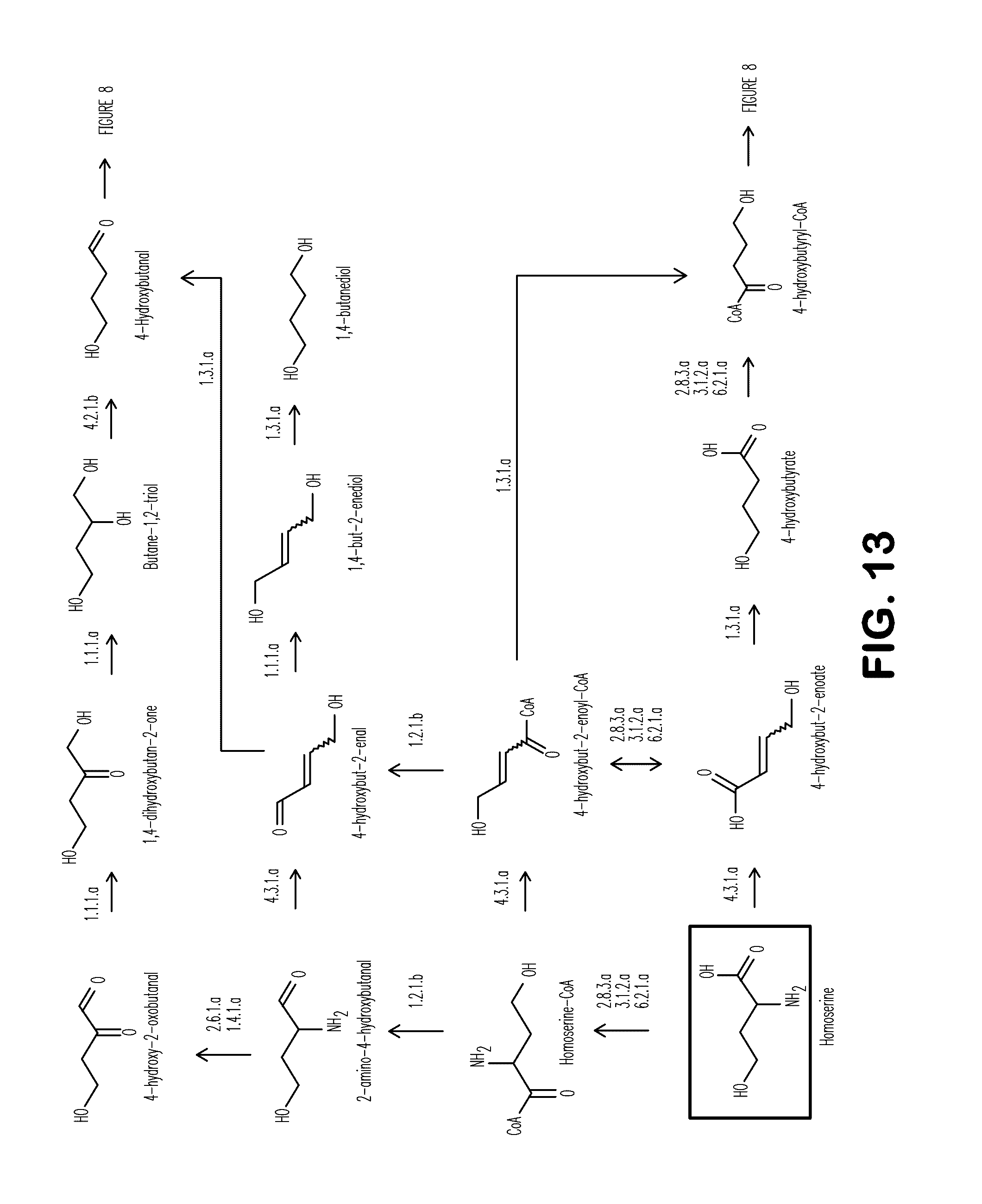
D00014

D00015

D00016

D00017

D00018

D00019

D00020

D00021

D00022

D00023

D00024

D00025

D00026
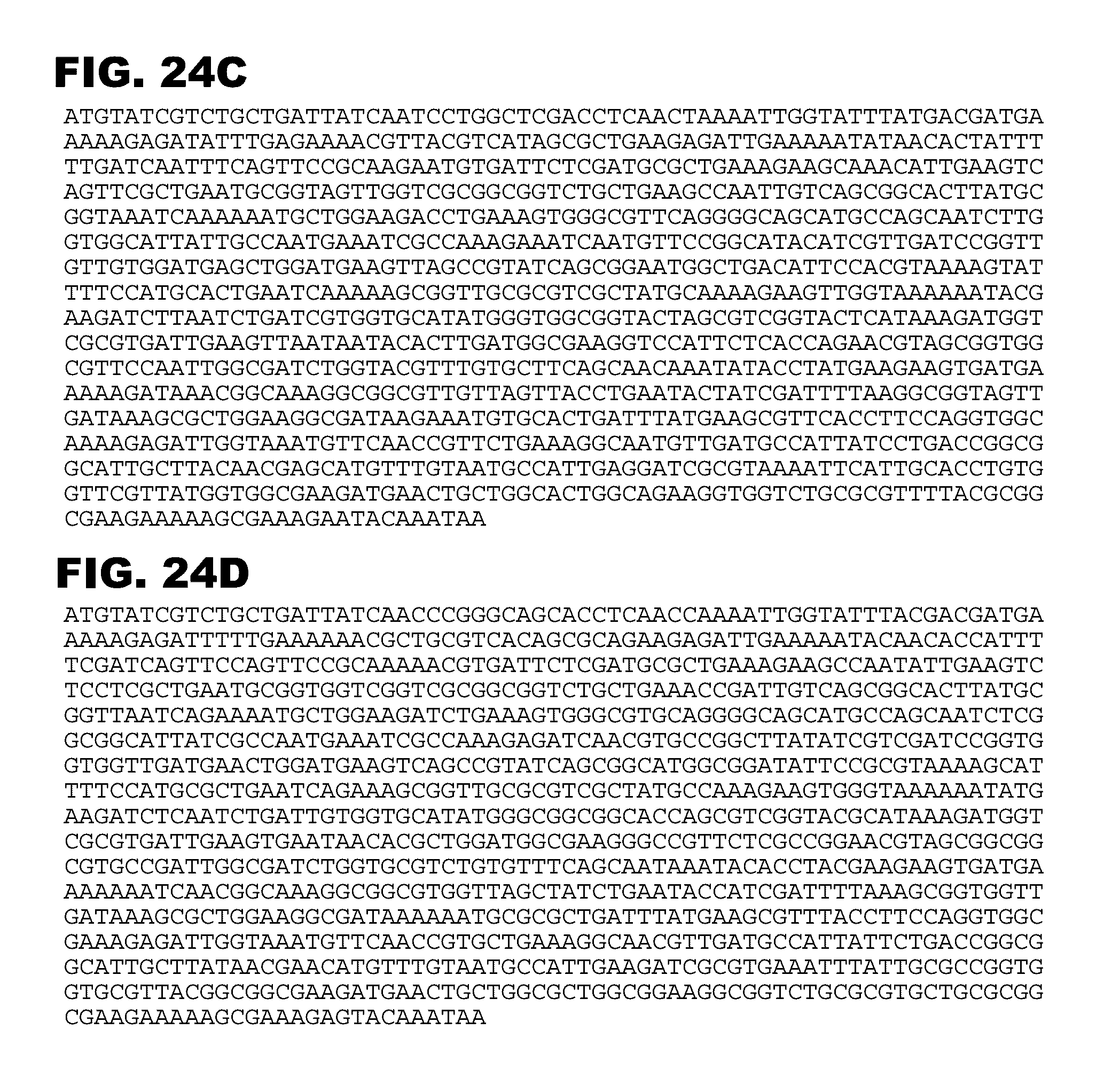
D00027
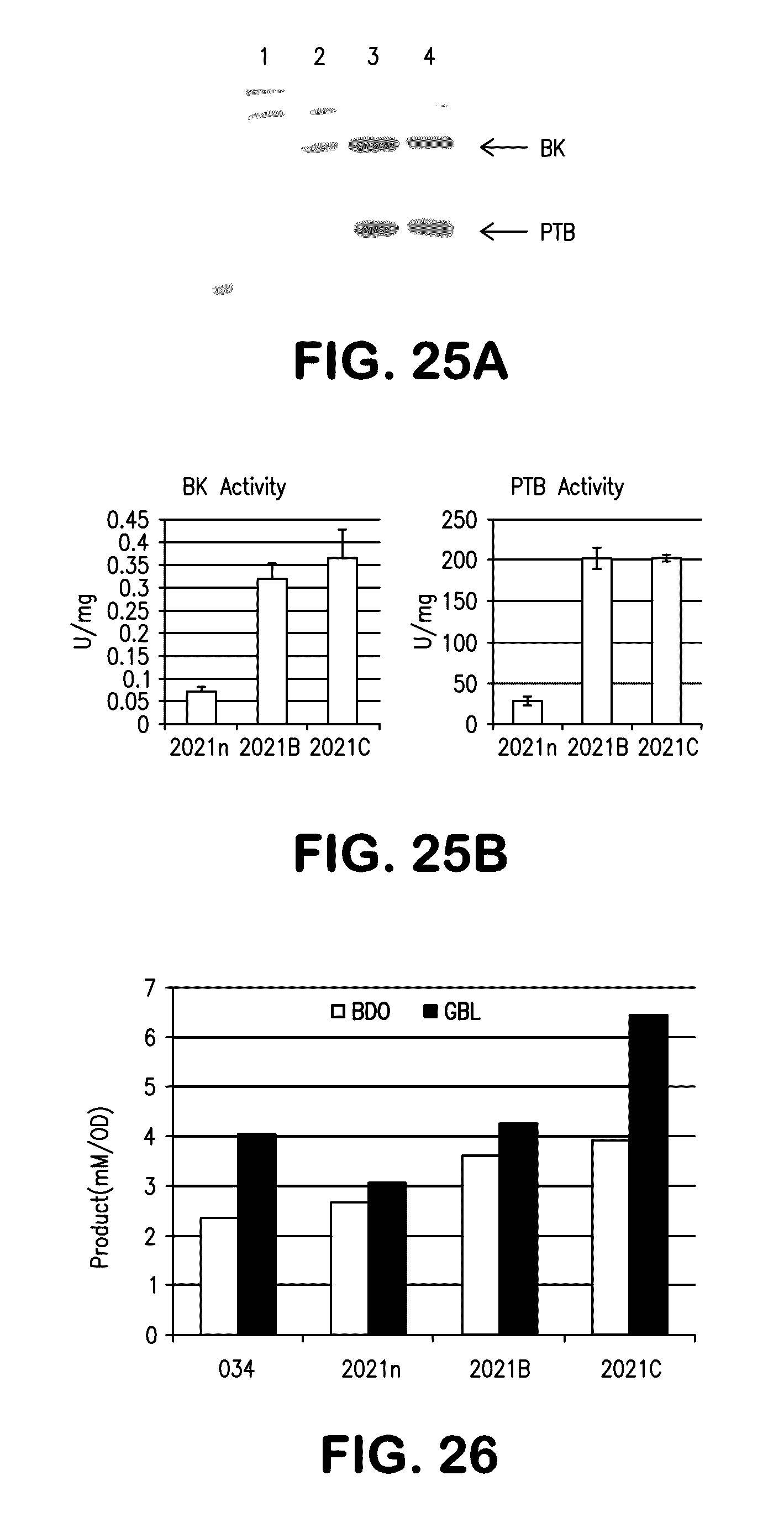
D00028

D00029

D00030

D00031

D00032
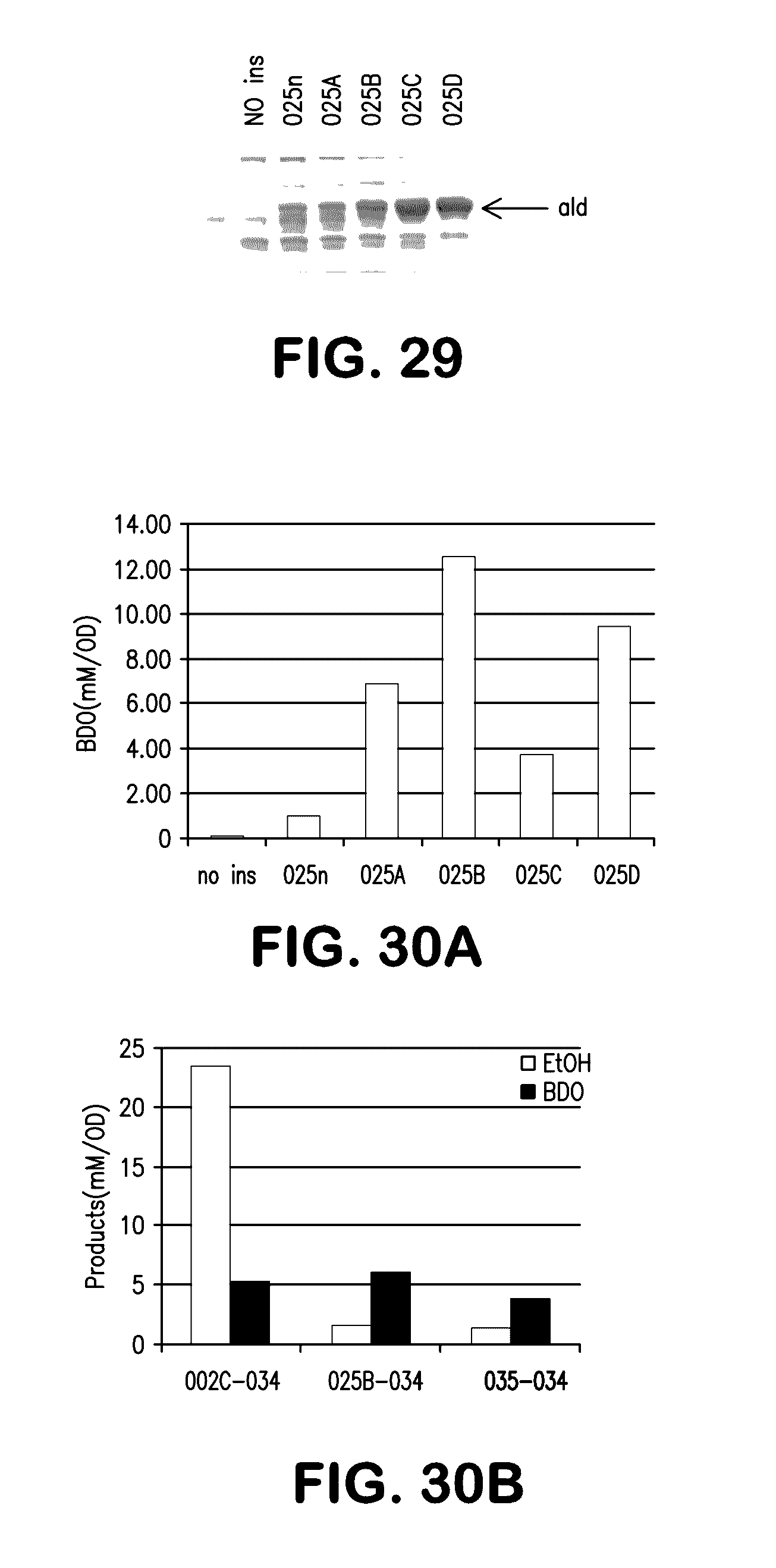
D00033

D00034
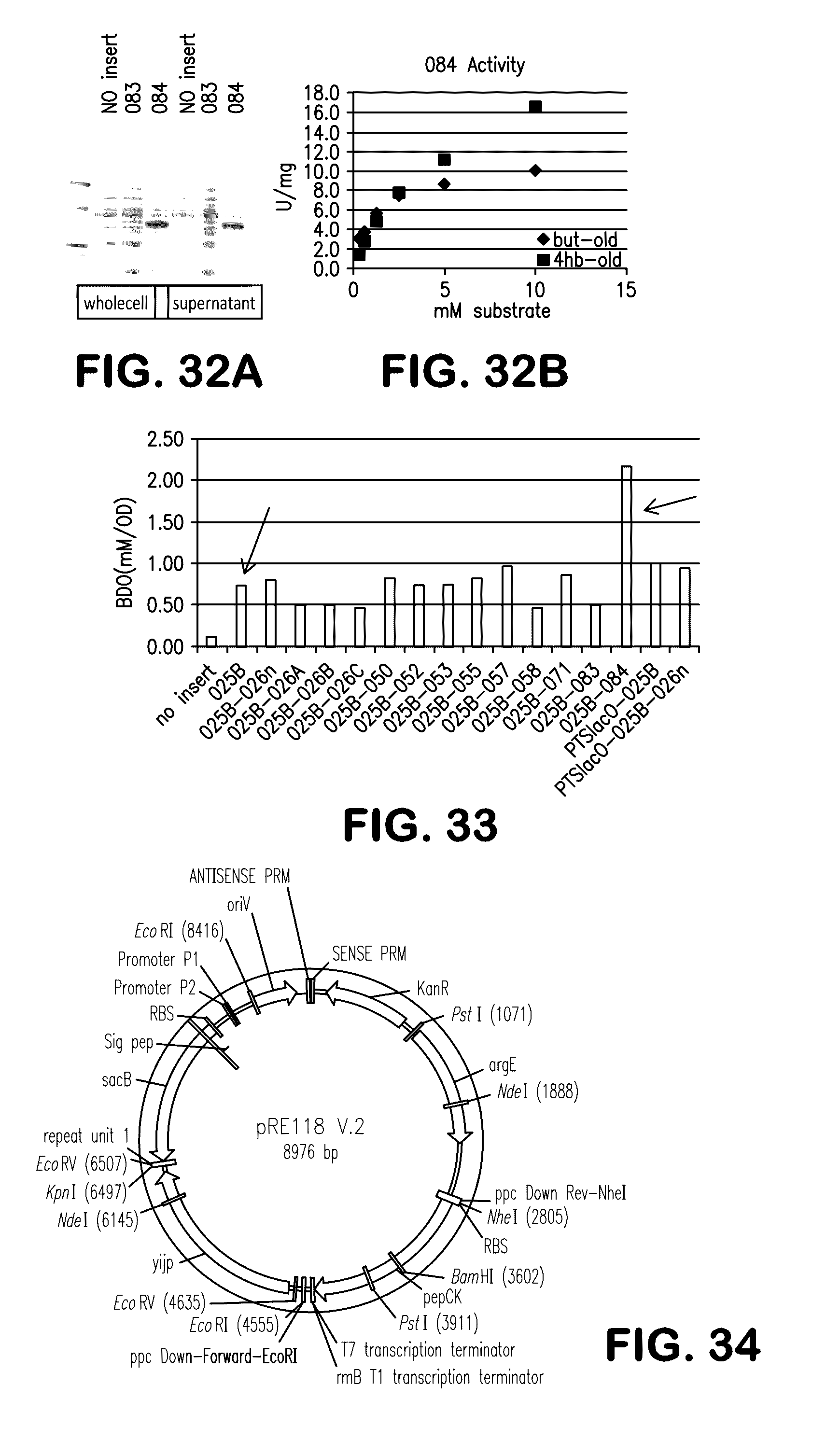
D00035

D00036

D00037

D00038

D00039

D00040

D00041
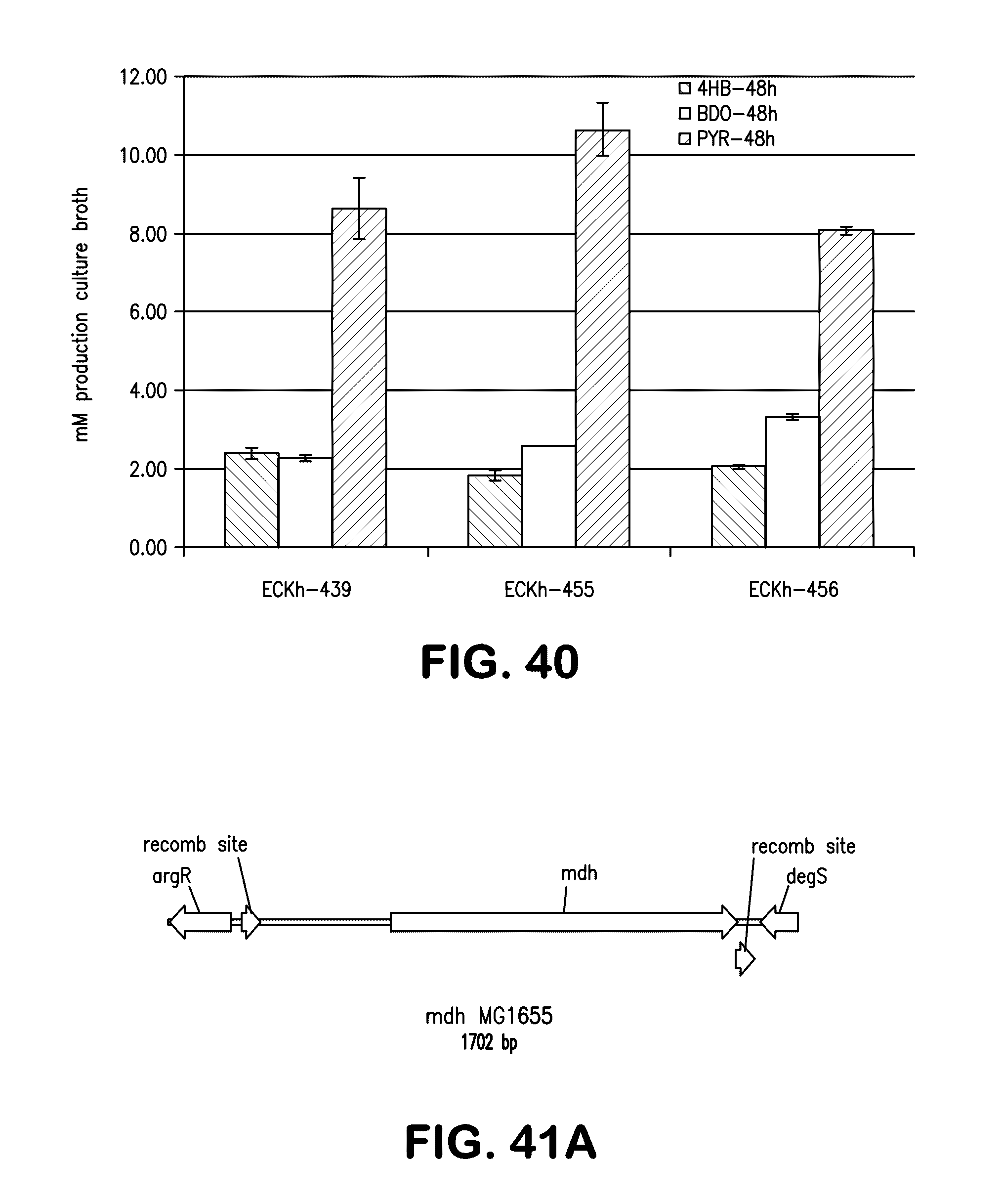
D00042

D00043

D00044

D00045

D00046

D00047

D00048

D00049

D00050

D00051

D00052

D00053

D00054

D00055
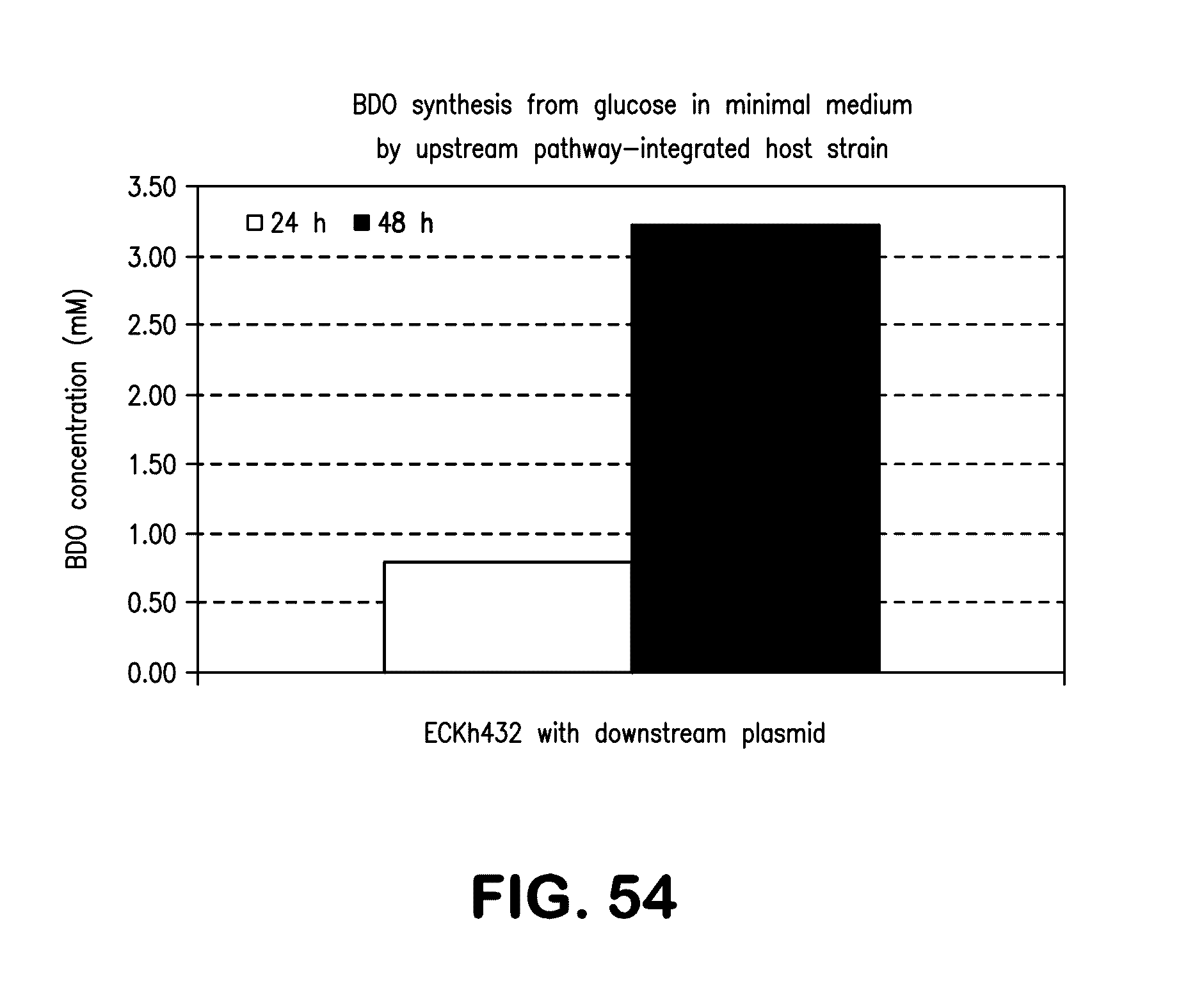
D00056

D00057

D00058

D00059

D00060

D00061

D00062

D00063

D00064

D00065

D00066

D00067

D00068

S00001
XML
uspto.report is an independent third-party trademark research tool that is not affiliated, endorsed, or sponsored by the United States Patent and Trademark Office (USPTO) or any other governmental organization. The information provided by uspto.report is based on publicly available data at the time of writing and is intended for informational purposes only.
While we strive to provide accurate and up-to-date information, we do not guarantee the accuracy, completeness, reliability, or suitability of the information displayed on this site. The use of this site is at your own risk. Any reliance you place on such information is therefore strictly at your own risk.
All official trademark data, including owner information, should be verified by visiting the official USPTO website at www.uspto.gov. This site is not intended to replace professional legal advice and should not be used as a substitute for consulting with a legal professional who is knowledgeable about trademark law.